Работа поддержана грантами РФФИ № 13-04-00941 и 13-04-00741.
Медицинский портал для врачей
количество статей
6422
Загрузка...
Пожалуйста, авторизуйтесь:
Обзоры
Асинхронное развитие сна как вероятная причина снижения когнитивных функций и возникновения ряда патологических состояний, связанных с циклом «сон – бодрствование»
"ЭФФЕКТИВНАЯ ФАРМАКОТЕРАПИЯ. Неврология спецвыпуск (22)
- Аннотация
- Статья
- Ссылки
- English
В статье представлен краткий обзор теорий, рассматривающих сон как период восстановления эффективности работы мозга после периода бодрствования, и работ, подтверждающих снижение уровня когнитивной деятельности на фоне возросшей сонливости. Предложена альтернативная гипотеза, объясняющая снижение когнитивных функций не ухудшением работоспособности мозга, а отключением отдельных корковых зон от управления поведением из-за развития в них локального сна. Дан обзор исследований асинхронного развития сна в корковых зонах. Причины и условия развития локального сна рассмотрены в контексте разрабатываемой авторами висцеральной теории сна. В основе этой теории лежит предположение, что во время сна центральные отделы нервной системы, включая кору мозга, переключаются с анализа информации, поступающей из внешней среды, на анализ потока сигналов от висцеральных систем организма. Приведена предполагаемая схема переключений основных информационных потоков в цикле «сон – бодрствование». Показано, что асинхронность этих переключений может объяснить возникновение ряда патологических состояний, связанных с циклом «сон – бодрствование» (просоночные галлюцинации, вздрагивания во сне, сновидения и ночные кошмары, сомнамбулизм и паралич сна). Предложенная теория предсказывает появление «висцеральных галлюцинаций», которые могут лежать в основе таких состояний, как укачивание, болезнь движений, и ряда других психосоматических патологий.
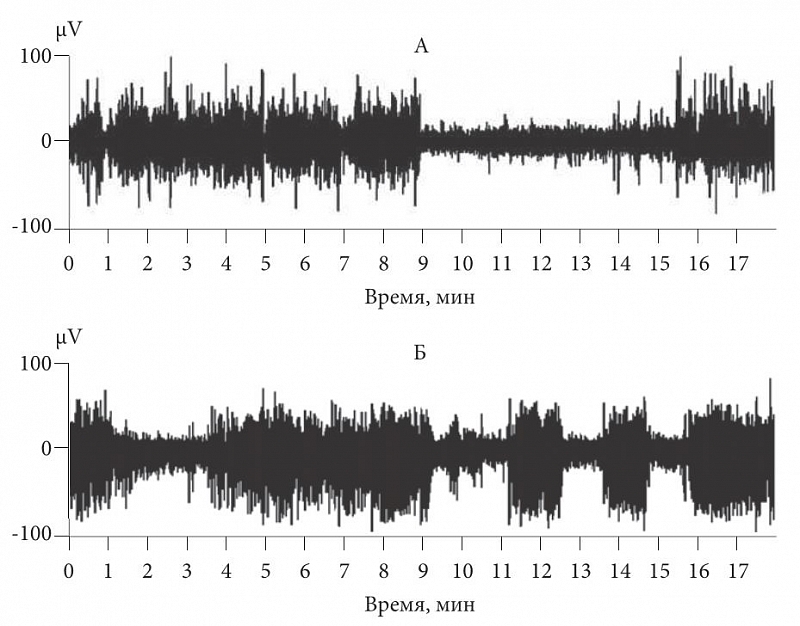
Рис. 1. Электроэнцефалограмма кролика: А – периоды медленного сна и бодрствования; Б – имитация дельта-волн сна зрительной стимуляцией в состоянии активного бодрствования
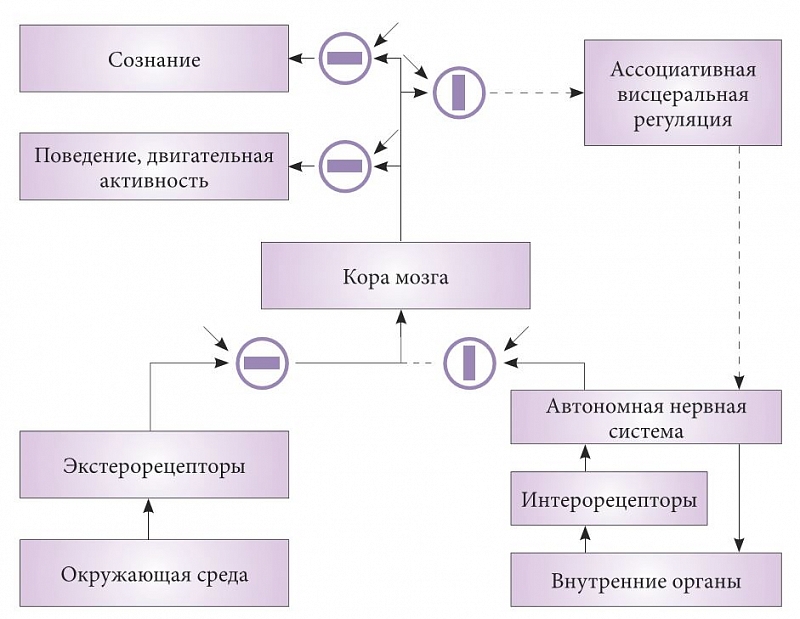
Рис. 2. Схема состояния информационных потоков организма в период бодрствования
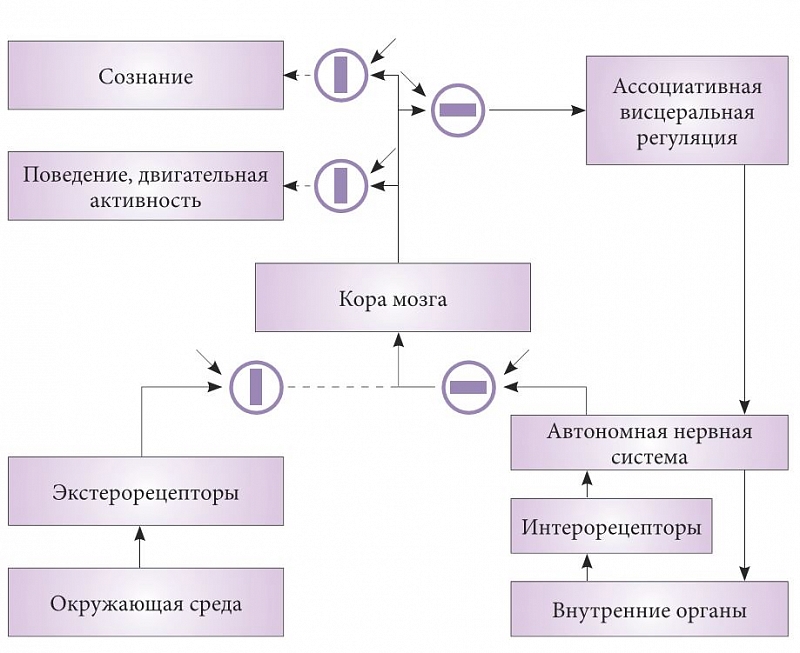
Рис. 3. Схема состояния информационных потоков организма в период сна
Введение
Давно замеченная связь техногенных катастроф, дорожных происшествий и медицинских ошибок с наступлением темного времени суток свидетельствует о снижении эффективности работы мозга в периоды повышенной сонливости [1–3]. Данные наблюдения подтверждались и результатами специальных исследований, в которых было показано, что депривация сна затрудняет способность к обучению и запоминание материала [4, 5]. Одни авторы связывали консолидацию памяти и, следовательно, обучаемость с медленноволновой фазой сна, тогда как другие – с фазой быстрого сна. При этом определенное влияние сна на процессы консолидации памяти ни у кого не вызывало сомнения.
Однако результаты экспериментов по депривации сна были весьма противоречивы и зависели не только от типа предъявляемой задачи, времени сна, его циркадианной фазы, в которой тестировалось выполнение задачи, но и от индивидуальных особенностей испытуемых. В подробном обзоре литературы о влиянии депривации сна на обучение и память был сделан вывод: в результате нарушения сна ухудшается выполнение сложных задач, требующих принятия решения, гибкости, инновации или изменения плана действий [6]. Отмечаемое ухудшение кратковременной зрительной памяти после депривации сна можно объяснить не столько затруднением консолидации памяти, сколько снижением зрительного внимания и/или затруднением обработки зрительной информации [7]. Складывается впечатление, что даже небольшое различие в протоколе экспериментов приводило к существенному расхождению выводов относительно зависимости процессов памяти от характеристик сна. В пользу этого заключения свидетельствует и то, что некоторые результаты были получены только определенными группами исследователей [8].
Однако наблюдаемое влияние сна на запоминание было не столь значительным, и казалось маловероятным, что для небольших улучшений требовались столь глобальные изменения в работе головного мозга, да и всего организма, которые происходят в период сна.
В то же время редкие, но драматические последствия антропогенных катастроф и аварий на дорогах, которые являются результатом сонливости, а также фантастические картины сновидений поддерживали идею о том, что сон важен прежде всего для эффективного функционирования мозга. Авторы большинства теорий сна предлагали различные механизмы, которые могли быть использованы мозгом в этих целях: теория нейронных групп [9] или теория синаптического гомеостаза [10]. Исходя из этих гипотез, было бы логично ожидать, что в результате депривации сна в первую очередь должен страдать именно мозг.
Однако исследования A. Rechtschaffen и соавт. [11–13] показали, что тотальная депривация сна приводит прежде всего к нарушениям висцеральной системы (выпадение волос, появление язв на коже или слизистой желудка) и к неминуемой гибели животных. Поражало, что у крыс, погибших в результате депривации сна, единственным органом, не имевшим ярких дегенеративных изменений, был именно мозг [12]. Это наблюдение было удивительным, поскольку негативное воздействие депривации сна на работу мозга хорошо известно.
Феномен локального сна отдельных зон коры головного мозга
В ходе исследования активности нейронов коры головного мозга у поведенчески бодрствующих животных была выявлена причина снижения эффективности поведения, связанная со сном, но проявляющаяся при нормальной работе мозга. Было показано, что в условиях общего бодрствования при определенных условиях сон может развиваться локально – только в некоторых областях коры. Локальный сон опасен тем, что его появление и развитие не замечают ни сам человек, ни окружающие его люди.
Феномен локального (или частичного) сна был замечен давно, при исследовании зрительных зон лобной коры кошек [14]. Нейроны этой части мозга активно реагировали на зрительные стимулы в периоды повышенной поведенческой активности, но их ответы снижались и пропадали в периоды спокойного бодрствования. Наблюдаемая картина импульсации нейронов была типичной для периода сна. Данное явление мы постоянно отмечали в ходе продолжительных исследований по изучению зрительных зон теменной и височной коры головного мозга, а также ассоциативной соматосенсорной коры кошек.
Более детальное исследование феномена локального сна было проведено при изучении свойств нейронов зрительной ассоциативной зоны V4 обезьяны [15]. В процессе длительного эксперимента при выполнении обезьяной монотонной задачи – сравнения зрительных стимулов, появляющихся на экране монитора, – часто замедлялась реакция нейронов в зоне V4 на один и тот же зрительный стимул. Со временем ответы нейрона могли полностью прекратиться, хотя обезьяна продолжала выполнять поставленную задачу. Качество решения задачи при этом оставалось высоким и лишь иногда несколько снижалось по сравнению с таковым в начале исследования. Следовательно, по крайней мере первичная зрительная кора мозга обезьяны продолжала работать.
Фоновая импульсация нейронов в зоне V4 в такие периоды напоминала типичную картину активности нейронов в естественном сне. Когда задачу прерывали, обезьяна сразу глубоко засыпала. После короткого сна реакция нейронов на зрительные стимулы обычно восстанавливались. В том же исследовании было отмечено, что даже в пределах одной зрительной зоны V4 локальный сон развивался не одновременно. Он сначала появлялся в участках проекции на кору периферии поля зрения, а потом распространялся к зоне проекции фовеа (центрального углубления в центре желтого пятна сетчатой оболочки глаза).
В описанном случае, очевидно, распространение локального сна начинается от сенсорных областей «высшего порядка», наиболее важных для сложных поведенческих ситуаций. Именно поэтому следовало ожидать появления локального сна у животных с более выраженными множественными сенсорными представительствами, прежде всего у приматов. И действительно, лобно-затылочный тренд развития сна был обнаружен A. Borbely и соавт. в коре головного мозга человека [16]. Исследования динамики распространения сна в мозге человека представлены и проанализированы в обзоре 2011 г. [17].
Возможность обнаружить локальный сон у животных с более простой организацией коры и ограниченным количеством сенсорных зон казалась маловероятной. Однако недавно локальный сон был зафиксирован в лобной области мозга крыс, выполняющих двигательную задачу [18].
Принимая во внимание возможность развития локального сна, ухудшение когнитивных функций в результате депривации сна можно объяснить не снижением эффективности работы мозга, а отключением некоторых зон коры больших полушарий от выполнения функций, за которые они отвечают в состоянии бодрствования.
На первый взгляд эта гипотеза подтверждает мнение о необходимости восстановления работоспособности мозга после длительного периода бодрствования, что и происходит во время сна. После продолжительного бодрствования для восстановления функционирования все отделы мозга могут заснуть одновременно или же какие-то участки могут это сделать в первую очередь, тогда как другие будут еще некоторое время оставаться в состоянии бодрствования.
Тем не менее следует напомнить: лишенные сна животные погибали не потому, что становились слепыми, глухими, забывали путь к кормушке или испытывали серьезные трудности с принятием решений, а из-за множественных висцеральных расстройств во всех системах жизнеобеспечения, включая иммунную систему [13]. В то же время именно мозг оказывался органом, наиболее устойчивым к депривации сна.
Учитывая вышесказанное, можно предложить другой фундаментально отличающийся сценарий. Что если мозг, как и другие висцеральные органы, не нуждается в специальном восстановительном отдыхе, связанном с полной потерей функциональности? Что если мозг, как и компьютеры, может работать эффективно неопределенно долго, а наблюдаемая «сенсорная изоляция» мозга во время сна отражает не прекращение работы, а переключение для обработки другого потока входной информации?
Электроэнцефалограмма сна: отражение особого режима работы мозга во сне или характера входной афферентации?
Общепризнано, что картины электроэнцефалограммы (ЭЭГ) медленноволнового сна и бодрствования резко отличаются. Это может свидетельствовать о принципиально разном характере работы мозга в этих состояниях. Однако интерпретация этого наблюдения также неоднозначна. Как правило, ЭЭГ медленного сна сравнивают с ЭЭГ пассивного бодрствования, когда люди или животные находятся без движений и сенсорные стимулы практически отсутствуют. Именно поэтому наблюдаемые различия в картинах ЭЭГ могут быть связаны с различной корковой афферентацией.
Для того чтобы проверить эту гипотезу, мы регистрировали нейронную активность и локальную электрокортикограмму у кошек и кроликов. Для бодрствования были подобраны стимулы (зрительный или соматический), вызывающие активную импульсацию нейронов, и эти стимулы предъявляли с частотой, соответствующей частоте дельта-волн. В состоянии активного бодрствования животного мы действительно наблюдали на ЭЭГ пачечную активность нейронов и медленные волны, не отличимые от активности в стадии медленного сна (рис. 1). На рисунке 1а показаны состояния бодрствования и медленного сна, отличающегося появлением высокоамплитудных дельта-волн. Рисунок 1б отражает состояние активного бодрствования – кролика стимулировали неяркими вспышками света. Эти вспышки вызывали появление вызванных ответов, которые, сливаясь, приводили к картине, неотличимой от таковой в медленном сне. Ответы на вспышки напоминали картину ЭЭГ при медленном сне. Как для зрительной, так и для соматосенсорной коры можно было подобрать такие условия сенсорной стимуляции, при которых спектры локальной электрокортикограммы в бодрствовании и во сне не отличались. Значит, характер ЭЭГ определяет именно входная корковая афферентация, а не состояние сна или бодрствования.
Феномен локального сна в контексте висцеральной теории сна
Переключение коры головного мозга на анализ другого потока входящей информации может лежать в основе перехода от состояния бодрствования ко сну. Какие же сигналы могли бы обеспечить такую периодическую афферентацию в период сна? С точки зрения физиологии ответ прост. Это могла быть активность разных висцеральных систем, например сердечно-сосудистой или дыхательной систем, желудочно-кишечного тракта.
Мы предположили, что во время сна нейроны мозга, которые в состоянии бодрствования обрабатывают экстероцептивную и проприоцептивную информацию, переключаются на анализ интероцептивной информации, приходящей от разных висцеральных систем. Ритмическая активность этих висцеральных систем определяет периодический афферентный поток к корковым зонам, что и отражает корковый ритм ЭЭГ медленноволнового сна. Таким образом, в период сна высшие отделы центральной нервной системы включаются в процессы висцеральной регуляции.
Следовательно, периоды локального сна – это не те периоды, когда усталый мозг прекращает обработку экстероцептивной информации для восстановления своей работоспособности, а периоды, когда нормально работающие зоны мозга, реагируя на сигналы тревоги, идущие от внутренних органов, переключаются на анализ поступающей от них висцеральной афферентации.
Это предположение, с одной стороны, представлялось фантастическим для общепринятой парадигмы мозга, сформированной на основе экспериментов, выполненных в состоянии бодрствования. С другой стороны, это открывало возможности для проверки данного предположения в достаточно простых условиях.
Результаты разноплановых экспериментов на живых моделях (кошки, обезьяны, кролики), проведенных с целью проверки высказанной гипотезы, опубликованы [19, 20]. Было подтверждено, что в периоды сна нейроны всех исследуемых зон коры головного мозга (затылочной, теменной и лобной) начинали реагировать на сигналы, приходящие из висцеральных систем. Сразу после пробуждения животного связь мозга с висцеральными системами прерывалась.
Этот факт позволяет говорить уже о висцеральной теории сна. Согласно этой теории именно афферентация от висцеральных органов с явно выраженной ритмичностью определяет волновую картину корковой активации в стадии медленного сна. Десинхронизация электроэнцефалограммы в периоды быстрого сна может свидетельствовать о поступлении афферентации от висцеральных органов, не имеющих явной периодической активности, – печени, почек, органов репродуктивной системы и мозга.
В рамках висцеральной теории сна мозг имеет двойственный статус: с одной стороны, это центральный информационный процессор, контролирующий поведение в окружающей среде во время бодрствования и обеспечивающий восстановление работоспособности висцеральных систем в период сна, а с другой – сложнейший висцеральный орган, который также нуждается в профилактическом обслуживании. Как и когда такое самообслуживание мозга осуществляется, пока не ясно. Можно предположить, что это происходит в определенные периоды быстрого или медленного сна или же осуществляется постоянно, например, с помощью глиальных клеток. Недавно открытый глимфатический (glymphatic) механизм может быть частью этого процесса [21, 22].
Предложенный подход проясняет запуск сна. Действительно, несоответствие текущих параметров, отражающих состояние любой висцеральной системы, и генетически заданных будет вызывать ощущение усталости или давления сна. Если ситуация допускает немедленный переход ко сну, последний будет происходить во всех корковых зонах. Когда в той или иной висцеральной системе потребность сна резко возрастает, но окружающая обстановка не позволяет перейти ко сну, сон может наступить лишь в некоторых корковых зонах при сохранении поведенческой активности. Развитие сна начинается с филогенетически наиболее молодых корковых зон «высокого уровня» (они преобладают в лобных отделах мозга). С этим, видимо, и связан выявленный в ЭЭГ лобно-затылочный тренд развития сна [16].
Поведенческие задачи, не требующие вовлечения высших корковых ресурсов, могут нормально реализовываться даже в условиях, когда часть корковых зон уже спит. Однако в ситуациях, когда всю мощь коры мозга необходимо задействовать для принятия сложных решений, локальный сон может привести к серьезным последствиям.
Условия возникновения локального сна указывают на то, что он наиболее вероятен в ситуациях, когда приходится поддерживать бодрствование на фоне высокого давления сна. Для людей это может происходить при работе в периоды максимальной сонливости.
У людей с привычным или вынужденным сокращенным ночным сном очень высок риск развития локального сна. Для некоторых видов деятельности локальный сон, вызванный хронической депривацией сна, может и не создавать особых трудностей. Однако для деятельности, связанной с принятием сложных и ответственных решений, особенно в периоды ночных смен, последствия возникновения локального сна могут быть печальны, поскольку увеличивается риск принятия ошибочных решений.
Кроме отрицательного влияния частичного сна на когнитивные функции, нужно учитывать и возможные последствия этого явления для висцерального здоровья. Как для успешного решения сложных проблем в бодрствовании важно участие всех корковых зон, так важно и вовлечение всех корковых зон в анализ висцеральной информации для эффективного управления всеми висцеральными системами. Появление локального сна в периоды бодрствования может свидетельствовать о недостаточной продолжительности общего сна в ночное время или указывать на вероятные скрытые проблемы в висцеральной сфере.
Рассмотренные выше возможности асинхронного развития сна в разных отделах коры головного мозга представляли варианты нормального функционирования мозга. Связанные с этим явлением негативные для организмов и окружения последствия отражали, скорее, нарушение «правил эксплуатации», то есть то, что принято называть человеческим фактором.
Организация основных информационных потоков в организме
Висцеральная теория сна предполагает и новые подходы к пониманию механизмов возникновения ряда патологических состояний организма, связанных также с нарушением синхронности работы структур, участвующих в управлении информационными потоками в цикле «сон – бодрствование».
Рассмотрим в рамках висцеральной теории сна основные информационные потоки в здоровом организме в состоянии бодрствования и сна, которые схематически отражены на рисунках 2 и 3. Открытые пути для проведения сигналов показаны черным цветом, пути, проведение сигналов по которым прекращено, – пунктиром. Структуры, блокирующие передачу сигналов, изображены в виде кружков. Прямоугольники внутри кружков отражают состояние структур: когда они расположены вдоль проводящих путей, проведение открыто, а поперек – закрыто. Каждый блок имеет второй вход, показанный стрелкой. Это вход управляющего сигнала, по команде которого происходит открытие или закрытие путей. Подобные структуры известны в нейрофизиологии, например триадные синапсы или системы пресинаптического торможения. Описанные модуляторы синаптической передачи нервной системы тоже, скорее всего, являются элементами таких управляющих систем.
На рисунке 2 представлено состояние проводящих путей нервной системы в бодрствовании. Сигналы из окружающей среды или о положении тела животного, преобразованные экстеро- и проприорецепторами в нервные импульсы, проходят через открытое блокирующее устройство и направляются для анализа в центральный блок («Кора мозга»). Результаты выполненного здесь анализа передаются в блок, отвечающий за поведение и двигательную активность. Параллельно выходящие из коры сигналы передаются в блок «Сознание», активация нейронов которого приводит к появлению ощущений себя и окружающего мира. Традиционно сознание связывают с активностью коры мозга. Однако результаты исследования сна определенно свидетельствуют, что это не так. Вопрос о вероятной локализации «структур сознания» в контексте результатов исследования сна уже рассмотрен [19], и мы не будем на нем останавливаться.
Во время бодрствования информация о состоянии внутренних органов, преобразованная интерорецепторами в нервные импульсы, поступает в структуры автономной нервной системы. Контроль внутренних органов в состоянии бодрствования осуществляется по линиям обратных проекций – от автономной нервной системы, которая организована сегментарно, что и препятствует взаимодействию сигналов из разных висцеральных систем [23]. Несмотря на это, автономная нервная система может поддерживать гомеостаз в течение ограниченного периода.
Однако со временем интерорецепторы начинают передавать сигналы об отклонении текущих параметров от генетически заданных норм. Видимо, именно эти сигналы воспринимаются животными как чувство усталости и заставляют их искать безопасное и удобное место для сна. Более детально динамика перехода ко сну в свете висцеральной теории рассмотрена ранее [20].
Во время сна происходит радикальное изменение информационных потоков в нервной системе (рис. 2). В идеале после получения разрешения на сон все блокирующие системы одновременно меняют положение. Во сне прекращается передача сигналов от экстеро- и проприорецепторов в кору мозга. Но по тем же входным каналам в кору начинает поступать информация из автономной нервной системы о состоянии всех висцеральных систем организма.
Скорее всего, основной структурой, в которой осуществляется переключение корковой афферентации с экстеро- и проприоцептивных входов на интероцептивные, является таламус. Весьма вероятно, что это и есть его основная функциональная нагрузка. При смене корковой афферентации с экстероцептивной на интероцептивную естественно ожидать изменений и в эфферентных корковых проекциях. Потоки информации, выходящие из коры во время сна, отражают результаты обработки корой головного мозга висцеральной информации, и они не должны направляться в структуры, связанные с двигательной активностью, поведением и сознанием. На схеме блокирующие системы на этих путях во время сна закрыты.
Факт отключения сознания от внешнего мира во время сна хорошо известен каждому из собственного опыта. Кроме того, из анатомии и физиологии известно, что во время сна активно блокируется передача сигналов из коры головного мозга на мотонейроны спинного мозга. Это приводит к расслаблению мышц тела, которое достигает максимума в периоды быстрого сна – так называемая атония сна [24].
Одновременно во сне должно открываться проведение по новому пути из коры мозга на структуры, связанные с ассоциативной обработкой висцеральной информации, где замыкается петля управления всеми висцеральными системами во время сна. Можно предположить, что центром ассоциативной висцеральной регуляции является гипоталамус.
Когда все висцеральные параметры в результате включения коры в процессы висцеральной интеграции будут приведены в норму, команда на сон снимается, все блокирующие системы меняют положение и животное пробуждается.
Так выглядит идеальная картина переключения информационных потоков в цикле «сон – бодрствование». Однако основные элементы этой схемы – системы, блокирующие проведение по тому или иному пути, – это не электронные реле, а химические синапсы. Эффективность их работы зависит от многих внешних по отношению к этому синапсу условий.
Таким образом, анализируя работу представленной схемы, нужно помнить, что переключение блокирующих систем при патологических факторах может развиваться асинхронно и растянуться на значительное время. В результате на фоне бодрствования часть структур мозга может быть переключена в состояние сна. И наоборот, в состоянии общего сна отдельные проводящие пути могут оказаться открытыми, как в бодрствовании. Возникающая из-за этого асинхронность переключения информационных потоков, безусловно, приведет к смешению экстероцептивной и интероцептивной информации на входе в кору мозга. Выходящие сигналы из коры мозга могут быть направлены не тем адресатам. Далее мы рассмотрим некоторые вероятные последствия таких ложных переадресаций.
Просоночные галлюцинации
Часто при переходе из состояния бодрствования ко сну наблюдаются просоночные (гипнагогические) галлюцинации в виде существ, ползущих по стене, или движущихся или деформирующихся предметов из окружающей обстановки [25]. Просоночные галлюцинации возможны, когда под давлением сна уже открывается путь для висцеральной афферентации в кору, но проведение зрительной информации еще не блокировано и сохранена связь коры с блоками поведения и сознания. Открывшийся поток висцеральных сигналов по интенсивности будет таким же, как и поток зрительных сигналов. Всплески импульсной активности от висцеральных входов, попав в сознание, вызовут зрительные образы, наложенные на все еще воспринимаемую реальную зрительную сцену.
Вздрагивание во сне (старты сна)
Иногда при переходе от бодрствования ко сну выход из коры головного мозга в сознание перекрывается нормально, а выход в структуры, связанные с двигательной активностью, остается открытым. При развитии сна активация в моторных отделах коры, вызванная уже висцеральными входами, не блокируется, а передается на мотонейроны спинного мозга. Возбуждение двигательных нейронов приводит к резким движениям туловища и конечностей, которые прерывают развивающийся сон. Это модель распространенного и известного многим состояния вздрагиваний во сне, часто сопровождающегося ощущением падения [26].
Сновидения
Самой часто встречающейся и безобидной патологией сна являются сновидения. Причина их возникновения, с нашей точки зрения, очень проста. Блокирующая система на пути из коры в сознание в состоянии сна или в процессе его развития оказывается не полностью перекрытой. Нельзя исключить и то, что висцеральные сигналы в определенные моменты достигают интенсивности, которая позволяет им преодолеть возросшие пороги проведения по каналам в сознание. Висцеральные системы не имеют представления в нашем сознании, поэтому для него их сигналы чаще всего являются просто шумом. А шум может возбудить нейроны с самыми низкими порогами ответов, то есть нейроны, которые наиболее активно работали в предыдущий период бодрствования.
Можно также предположить, что в определенные моменты времени пространственное распределение возбуждения, идущего из коры и отражающего результат анализа висцеральной информации, окажется близким к пространственному профилю сформированных в бодрствовании образов каких-либо реальных объектов. В результате вспыхнувший образ по ассоциативным связям в системе сознания запустит развитие фантастических сюжетов.
Скорее всего, сновидения представляют собой явления переходного периода от бодрствования ко сну или чаще от сна к бодрствованию, когда блокирующая система на входе в сознание еще не полностью закрыта или уже немного приоткрыта. Эта безвредная патология имеет свои неприятные стороны. Когда блок на входе в отдел сознания не удается перекрыть в течение длительного времени, возникают ночные кошмары, прерывающие течение нормального сна. Это встречается при наличии глубокой патологии.
Паралич сна
Нередко при переходе от сна к бодрствованию проявляется так называемый паралич сна. Человек нормально просыпается, адекватно воспринимает окружающую действительность и собственное тело, но не способен в течение некоторого времени (от нескольких десятков секунд до минуты) совершить какое-либо произвольное движение. Согласно нашей схеме вероятной причиной этого является запаздывание снятия блока передачи моторных команд из коры в отдел поведения и двигательной активности, когда сознание уже проснулось, а система управления движениями еще продолжает спать.
Сомнамбулизм (снохождение)
Встречаются и прямо противоположные случаи. Открывается проведение сигналов из коры в отдел поведения и двигательной активности, а путь в отдел сознания все еще закрыт (рис. 2 и 3). Это приводит к снохождениям – довольно часто встречающемуся явлению, особенно в детском возрасте. Люди встают ночью с постели и совершают разнообразные действия или прогулки, длительность которых может быть самой разной. Глаза при хождении во сне обычно открыты. Люди не натыкаются на препятствия, их движения хорошо координированы. Часто в конце таких прогулок они могут найти другое место для сна, а после пробуждения не понимают, как сюда попали [27]. Явление сомнамбулизма является еще одним аргументом в пользу того, что сознание отделено от коры мозга и структур, программирующих движения тела.
Висцеральные галлюцинации
В разделе о просоночных галлюцинациях мы говорили об особенностях восприятия, вызванных смешением экстероцептивной и интероцептивной информации на входе в кору, которое оказывается возможным в переходные периоды от бодрствования ко сну, когда одни проводящие пути еще не полностью закрылись, а другие еще не полностью открылись. В результате интероцептивные сигналы проецируются на реальную жизненную ситуацию и воспринимаются людьми как галлюцинации.
Похожее явление теоретически может развиваться и в противоположном направлении. По аналогии мы назвали его висцеральными галлюцинациями. Можно предположить, что под давлением сна начнут открываться пути, связывающие висцеральные системы с корой мозга (рис. 2 и 3), но жизненные обстоятельства не позволяют немедленно перейти ко сну. Связь коры с экстероцептивными входами и выходы на сознание и двигательную активность остаются открытыми. При этом может оказаться, что результаты анализа экстероцептивных сигналов будут попадать в блок ассоциативной висцеральной интеграции и расцениваться там как сигналы коры анализа висцеральных входов. Теперь управлять висцеральными системами будут галлюцинаторные сигналы. Понятно, что ни к чему хорошему в отношении висцеральной системы это не приведет. Известные негативные висцеральные последствия острого стресса, возможно, имеют такой же механизм развития.
Данным механизмом можно объяснить и появление висцеральных компонентов (тошноты, рвоты), болезни движения или укачивания. Ритмичное покачивание, или периодическое, как на качелях, или постоянное, как в космическом полете, а также снижение гравитационной нагрузки резко меняют афферентную импульсацию от механорецепторов в стенках органов желудочно-кишечного тракта. Необходимость экстренного анализа причин такого необычного изменения афферентации приведет к усилению давления сна. Под этим давлением будет приоткрыт корковый выход в структуры висцерального анализа и, возможно, вход висцеральной информации в кору. Но высокая поведенческая активность в это время исключает переход ко сну, и кора мозга остается связанной с мощным потоком экстероцептивных входов. Этот поток экстероцептивных сигналов начинает забрасываться в отделы анализа висцеральной информации, что делает информационный поток непривычным. Система расценивает его как поступающий из органов желудочно-кишечного тракта. Естественной реакцией на эти сигналы будет команда экстренной очистки желудка от источников пугающих сигналов, то есть рвотный рефлекс.
Насколько нам известно, механизм, ведущий к висцеральным галлюцинациям, ранее никем не рассматривался. Вероятно, что в сонно-депривированном обществе негативные для висцеральных систем последствия работы подобного механизма могут быть существенными. Скорее всего, именно интерференция экстероцептивной и интероцептивной информации на пути в кору и на выходах из коры может быть причиной многих психосоматических заболеваний.
Детальное выяснение динамики участия высших отделов мозга в регуляции висцеральных функций в период сна – задача на будущее. Пока же смело можно утверждать, что эффективный и достаточный сон – это самый дешевый, безопасный и приятный способ улучшения как психического, так и физического здоровья.
1. Дорохов В.Б. Сомнология и безопасность профессио- нальной деятельности // Журнал высшей нервной де- ятельности им. И.П. Павлова. 2013. Т. 63. № 1. С. 33–47.
2. Ковальзон В.М. Основы сомнологии: физиология и нейрохимия цикла «бодрствование – сон». М.: Бином. Лаборатория знаний, 2012.
3. Akerstedt T., Hallvig D., Anund A. et al. Having to stop driving at night because of dangerous sleepiness – awareness, physiology and behaviour // J. Sleep Res. 2013. Vol. 22. № 4. P. 380–388.
4. Maquet P. The role of sleep in learning and memory // Science. 2001. Vol. 294. № 5544. P. 1048–1052.
5. Diekelmann S., Born J. The memory function of sleep // Nat. Rev. Neurosci. 2010. Vol. 11. № 2. P. 114–126.
6. Harrison Y., Horne J.A. The impact of sleep deprivation on decision making: a review // J. Exp. Psychol. Appl. 2000. Vol. 6. № 3. P. 236–249.
7. Chuah L.Y., Chee M.W. Cholinergic augmentation modulates visual task performance in sleep-deprived young adults // J. Neurosci. 2008. Vol. 28. № 44. P. 11369–11377.
8. Genzel L., Dresler M., Wehrle R. et al. Slow wave sleep and REM sleep awakenings do not affect sleep dependent memory consolidation // Sleep. 2009. Vol. 32. № 3. P. 302–310.
9. Krueger J.M., Obál F. A neuronal group theory of sleep function // J. Sleep Res. 1993. Vol. 2. № 2. P. 63–69.
10. Tononi G., Cirelli C. Sleep function and synaptic homeostasis // Sleep Med. Rev. 2006. Vol. 10. № 1. P. 49–62.
11. Everson C.A., Bergmann B.M., Rechtschaffen A. Sleep deprivation in the rat: III. Total sleep deprivation // Sleep. 1989. Vol. 12. № 1. P. 13–21.
12. Cirelli C., Shaw P.J., Rechtschaffen A. et al. No evidence of brain cell degeneration after long-term sleep deprivation in rats // Brain Res. 1999. Vol. 840. № 1–2. P. 184–193.
13. Rechtshaffen A., Bergmann B.M. Sleep deprivation in the rat: an update of the 1989 paper // Sleep. 2002. Vol. 25. № 1. P. 18–24.
14. Пигарев И.Н. Новая зрительная зона на ниж- ней стенке крестовидной борозды мозга кошек // Нейрофизиология. 1984. Т. 16. № 6. С. 766–773.
15. Pigarev I.N., Nothdurft H.C., Kastner S. Evidence for asynchronous development of sleep in cortical areas // Neuroreport. 1997. Vol. 8. № 11. P. 2557–2560.
16. Werth E., Achermann P., Borbely A.A. Brain topography of the human sleep EEG: antero-posterior shifts of spectral power // Neuroreport. 1996. Vol. 8. № 1. P. 123–127.
17. Ferrara M., De Gennaro L. Going local: insights from EEG and stereo-EEG studies of the human sleep-wake cycle // Curr. Top. Med. Chem. 2011. Vol. 11. № 19. P. 2423–2437.
18. Vyazovskiy V.V., Olcese U., Hanlon E.C. et al. Local sleep in awake rats // Nature. 2011. Vol. 472. № 6. P. 443–447.
19. Пигарев И.Н. Висцеральная теория сна // Журнал вы- сшей нервной деятельности им. И.П. Павлова. 2013. Т. 63. № 1. С. 86–104.
20. Пигарев И.Н., Пигарева М.Л. Сон, эмоции и висцераль- ный контроль // Физиология человека. 2013. Т. 39. № 6. С. 31–44.
21. Nedergaard M. Neuroscience. Garbage truck of the brain // Science. 2013. Vol. 340. № 6140. P. 1529–1530.
22. Xie L., Kang H., Xu Q. et al. Sleep drives metabolite clearance from the adult brain // Science. 2013. Vol. 342. № 6156. P. 373–377.
23. Ноздрачев А.Д. Физиология вегетативной нервной системы. Л.: Медицина, 1983.
24. Chase M.H. Confirmation of the consensus that glycinergic postsynaptic inhibition is responsible for the atonia of REM sleep // Sleep. 2008. Vol. 31. № 11. P. 1487–1491.
25. Sturzenegger C., Bassetti C.L. The clinical spectrum of narcolepsy with cataplexy: a reappraisal // J. Sleep Res. 2004. Vol. 13. № 4. P. 395–406.
26. Walters A.S. Clinical identification of the simple sleep-related movement disorders // Chest. 2007. Vol. 131. № 4. P. 1260–1266.
27. Kavey N.B., Whyte J., Resor S.R. Jr. et al. Somnambulism in adults // Neurology. 1990. Vol. 40. № 5. P. 749–752.
Asynchronous development of sleep – a plausible cause of cognitive impairment and a number of pathological conditions related to sleep-wake cycle
I.N. Pigarev1, M.L. Pigareva2
1 A.A. Kharkevich Institute of Information Transfer Issues of the Russian Academy of Sciences
2 Institute of Higher Nervous Function and Neurophysiology of the Russian Academy of Sciences
Contact person: Ivan Nikolayevich Pigarev, pigarev@iitp.ru
The article gives a review to several theories of sleep. According to them, sleep is regarded as a period of recovery of effective brain functioning after active wakefulness. Several studies have demonstrated the association between decreased cognition and increased sleepiness. According to the alternative hypothesis, cognitive impairment is explained by shutdown of certain cortical zones due to their local sleep rather than by impaired brain performance. The studies of asynchronous development of sleep in the cortical zones are described. Causes and conditions for the development of local sleep are discussed in the framework of authirs’ visceral theory of sleep. The theory is based upon an assumption that, during sleep, central parts of the nervous system shift from the analysis of external information to the analysis of internal (visceral) signals. Hypothetical switching network diagram of the main information pathways in the sleep-wake cycle is presented. The authors emphasize that asynchronous switching may explain the development of the number of the sleep-wake cycle-associated pathological conditions (i.e. waking hallucinations, sleep starts, dreams/anxiety dreams, sleepwaking and sleep paralysis). This theory may predict emergence of ‘visceral hallucinations’ which are regarded as a potential cause of seasickness, motion-sickness and some other psychosomatic disorders.
Новости на тему
26.01.2023
10.08.2022 01:00:00
Выявлены оптимальные диагностические критерии болезни Паркинсона с помощью искусственного интеллекта
15.07.2022 01:00:00
07.07.2022 10:00:00
СТАТЬИ по теме
01.11.2023
01.11.2023
Отправить статью по электронной почте
Ваш адрес электронной почты:
Адрес электронной почты получателя:
Разделите несколько адресов электронной почты запятой
Сообщение(не обязательно)
Не более 1500 символов
Анти спам:
Для предотвращения спама, пожалуйста, введите в поле слово, которое видите ниже.
Обновить код
* адреса предоставленные Вами будут использоваться только для отправки электронной почты.