Менструальный цикл и энергетическая «политика» гипоталамуса
- Аннотация
- Статья
- Ссылки
- English
Введение
Менструальный цикл – это одно из наиболее загадочных явлений в человеческой биологии. Почему наш вид выработал ритмичную овуляцию, которая не зависит от коитуса? Каков биологический смысл менструального кровотечения, которое отсутствует у других млекопитающих? Есть и более практические вопросы: известно, что физические и психологические нагрузки, а также масса тела заметно влияют на менструальный цикл – но как и почему это происходит? В поисках ответов обратимся к процессам высокого уровня – к аналитической системе гипоталамуса.
Продление рода требует оптимальной внешней и внутренней среды. Женщина должна быть в безопасности, благоприятных социальных условиях, иметь неограниченный доступ к питательным веществам. В противном случае беременность может прерваться либо плод пострадает от дефектов развития.
Как организму оценить множество внешних и внутренних факторов и сделать выбор: пойти на риск зачатия сейчас – или пожертвовать частью фертильного времени, выжидая более подходящие условия? Принимая во внимание сложность задачи, можно сказать, что выбор физиологических состояний, подходящих для зачатия, не может осуществляться органами малого таза. Эту функцию должен выполнять орган, «знающий» в целом физиологический контекст.
Гипоталамус – это нейронный аппарат, связанный с корой головного мозга, эмоционально-поведенческой (лимбической) и эндокринной системами. Кроме того, через «окно» в гематоэнцефалическом барьере гипоталамус сканирует кровоток, следя за физиологическими сигналами. Благодаря такому обилию связей он может информированно управлять менструальным циклом, а при необходимости и предотвращать начало нецелесообразной беременности. Для этого гипоталамус использует единственный инструмент – гонадотропин-рилизинг-гормон (ГнРГ), или гонадолиберин.
ГнРГ: нейронная сеть и ее импульсный код
Гонадолиберин – это короткий пептид (10 аминокислот) с периодом жизни не более двух – четырех минут. Он поступает в портальную систему, импульсами переменной частоты и амплитуды образуя код, доступный нижележащему гипофизу. Частота и амплитуда импульсов меняются циклично, вначале обеспечивая выделение фолликулостимулирующего гормона, а затем подготавливая овуляторный выброс лютеинизирующего гормона. Именно эти фазные изменения и определяют механизм менструального цикла [1]. Монотонная стимуляция гипофиза, напротив, приводит к прекращению синтеза лютеинизирующего и фолликулостимулирующего гормонов [2], что можно использовать в терапевтических целях, назначая агонисты ГнРГ.
Гонадолиберин эффективно «настраивает» половую систему, но полностью зависит от слаженной работы своих нейронов. У млекопитающих из них образована сеть, причем удивительно малочисленная – всего несколько тысяч клеток [3]. Нейронная сеть распределена по нескольким областям гипоталамуса и из каждой направляет аксоны к воротным сосудам гипофиза [1]. Разветвленное строение сети позволяет собирать стимулирующие и подавляющие сигналы от физиологических процессов, за которыми наблюдает гипоталамус. Важный пример – это управляющее питанием аркуатное ядро, которое может менять работу ГнРГ-сети [4]. В неблагоприятном физиологическом контексте секреторный код ГнРГ искажен, а с ним и менструальный цикл, вплоть до функциональной аменореи.
Итак, каким принципом руководствуется гипоталамус, регулируя половую систему? Воспроизводство налагает на женский организм большие энергетические расходы и не способствует его выживанию, поэтому гипоталамус стремится «экономить» на половой функции при любых стрессовых условиях [5].
Стресс и продление рода: поиск подходящего времени
В современной биологии понятие «стресс» означает перегрузку адаптивных способностей организма [6]. В качестве универсального ответа на длительные неблагоприятные условия гипоталамус наращивает кортизол плазмы (и ликвора) через гипофизарно-надпочечниковую систему [7]. В ответ кортизол среди прочего подавляет секрецию ГнРГ [8]. Это происходит при множестве хронических состояний, различных по сути и происхождению, но единообразно истолкованных гипоталамусом как стресс: при депрессии и тревожных расстройствах, сахарном диабете, алкоголизме, нарушении режима «сон – бодрствование» (работа в ночные смены) [9]. Избыток кортизола также наблюдается у многих профессиональных спортсменов [10].
Однако «политика» гипоталамуса сложнее столь прямолинейного ответа на угрозы. Не все стрессовые нагрузки невыгодны организму, некоторые из них являются частью образа жизни. Чтобы отдать предпочтение той или иной деятельности, эволюция породила так называемую систему нейробиологического вознаграждения (neurobiological reward system).
Так, вид Homo sapiens адаптирован к бегу на длинные дистанции [11], при котором в кровь поступают эндогенные каннабиноиды. На заре истории человечества они облегчали многочасовые забеги во времена миграции и охоты, оказывая обезболивающее и эйфорическое действие [12]. Разумеется, они также угнетают выделение ГнРГ [13], поскольку столь тяжелые испытания не сочетаются с продлением рода. Вероятно, в наши дни это отчасти объясняет случаи аменореи среди бегуний [14].
Эндорфины (эндогенные опиаты) дополняют систему «вознаграждения»: они тоже смягчают боль и улучшают настроение, но опять-таки подавляют ГнРГ [15].
Итак, «вознаграждающие» агенты указывают гипоталамусу на время, неподходящее для воспроизводства. Насколько это важно с клинической точки зрения, ведь лишь немногие пациенты подвержены долгим изнурительным нагрузкам? Дело в том, что эндорфины [16] и эндоканнабиноиды [17] выполняют и другую функцию: они стимулируют поиск пищи, а также чувственно окрашивают ее прием, поощряя восстановление ресурсов. К несчастью, если гипоталамус малочувствителен к сигналам насыщения (что наблюдается при ожирении), то «вознаграждающие» вещества выделяются в хроническом избытке [18] с теми же последствиями для ГнРГ.
Остановимся подробнее на гипоталамической взаимосвязи между половой системой и питанием.
ГнРГ и энергетический метаболизм: «экономика» воспроизводства
Исследования на приматах показывают, что голодание или чрезмерный расход энергии устойчиво вызывает аменорею и прерывание беременности [19]. Эта приспособительная реакция носит характер более общий, нежели обеспечение жизнедеятельности организма: увеличение популяции в голодное время нецелесообразно. Пытаясь оценить, безопасно ли для индивидуума и вида сохранение половой функции, гипоталамус ориентируется на сигналы голода и насыщения.
Важнейший из них – нейропептид Y (NY), вызывающий сильное чувство голода и снижающий выбросы ГнРГ [4]. Голод стихает, когда инсулин и лептин – главные гормоны насыщения – «отключают» NY-нейроны аркуатного ядра [20]. Инсулин поступает в кровоток пропорционально поглощенной глюкозе, лептин же выделяют адипоциты, сообщая тем самым о количестве жировой ткани [21].
Кроме того, при голодании в желудке выделяется гормон грелин. В краткосрочной перспективе он лишь стимулирует поиск пищи, но продолжительное воздействие может замедлять половое созревание, нарушая секрецию ГнРГ [22].
Ограничение половой функции при нехватке энергии легко объяснимо, но нарушения менструальной функции при ожирении выглядят парадоксально. В действительности ожирение – это «голод среди изобилия». Гипоталамус, пораженный резистентностью к инсулину [23] и лептину [24] (то есть нечувствительный к насыщению), принимает меры для сбережения энергии, снижая в том числе и выделение ГнРГ.
Ожирение – это и разновидность затяжного воспаления [25]. Растущие адипоциты выделяют провоспалительные цитокины, из которых два – фактор некроза опухоли альфа и интерлейкин 6 – подавляют импульсную секрецию ГнРГ [26]. Для гипоталамуса воспаление всегда служит сигналом о неблагополучии, о вовлечении иммунной системы в борьбу с внешними агрессорами, вероятно инфекционного происхождения. Хроническая инфекция – неподходящее время для вынашивания потомства, а ожирение «симулирует» ее в асептической форме.
Наконец, линолевая кислота, относящаяся к классу омега-6 ненасыщенных жирных кислот, – важный субстрат для синтеза каннабиноидов – в рационе полных людей присутствует в избытке, определяя высокий каннабиноидный фон [27].
Уровнем выше: клеточное окружение ГнРГ-нейронов
До сих пор мы говорили о том, как с помощью входящих физиологических сигналов ГнРГ-сеть «настраивает» или прерывает менструальный цикл. Но двусложный подход «стимул → команда» лишен всякой гибкости. Можно предположить, что решение об остановке менструального цикла должно приниматься на другом, надсетевом уровне. Где же?
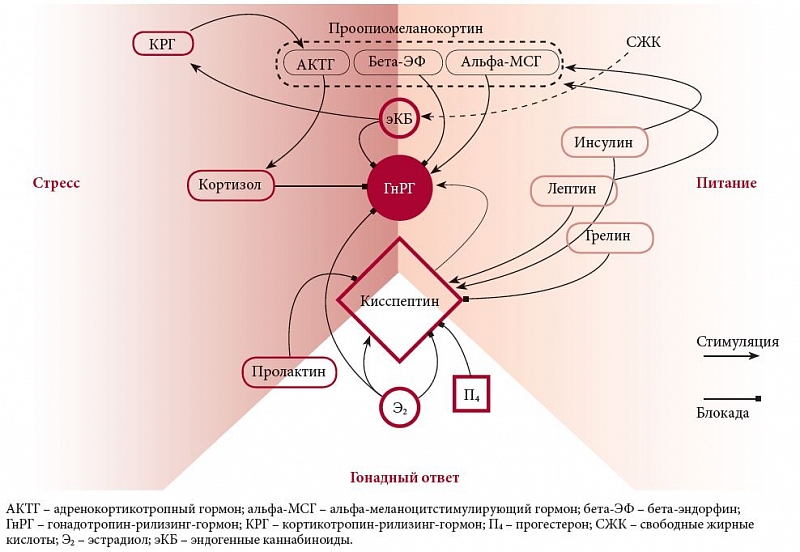
Мы обсуждали важность пищевых сигналов для выделения ГнРГ. Тем удивительнее, что сами ГнРГ-нейроны, похоже, почти лишены рецепторов к лептину [28], инсулину [29] и грелину [30]. Даже обратный эстрогеновый сигнал они получают только через рецептор эстрогенов типа бета, между тем преовуляторный скачок секреции лютеинизирующего гормона запускается через рецептор эстрогенов типа альфа [31]. Очевидно, что ГнРГ-сеть использует посредников, своеобразные «реле», задающие более гибкое управление (рисунок).
Первое место среди них принадлежит кисспептину (KiSS1): его нейроны прямо стимулируют секрецию ГнРГ, представляя в гипоталамусе «разрешительные» силы воспроизводства [32]. KiSS1-нейроны протягиваются к ГнРГ-синтезирующей сети незадолго до начала пубертата [33]. В дальнейшем именно через них, несущих рецепторы эстрогенов типа альфа, эстрогены оказывают обратное влияние на ГнРГ-сеть [34]. Интересно, что одна подгруппа KiSS1-нейронов замыкает обратную отрицательную связь [35], а другая, напротив, придает эстрогенам временный умножающий эффект незадолго до преовуляторного всплеска лютеинизирующего гормона [36]. По всей видимости, обратное отрицательное влияние прогестерона также хотя бы отчасти происходит через KiSS1-систему [37].
Через супрессию KiSS1-нейронов гипоталамус вызывает ановуляцию у кормящих матерей, ориентируясь на отрицательный баланс энергии из-за выделения молока. Судя по всему, в этом также участвует пролактин, напрямую подавляющий KiSS1-нейроны [38]. При голодании уровень кисспептина в гипоталамусе резко падает вместе с секрецией ГнРГ [39], а экспериментально восполнив кисспептин извне при хроническом недоедании, можно сократить задержку пубертата [40]. Вспоминая же «парадоксальное» бесплодие у полных женщин, нужно заметить, что при ожирении синтез кисспептина в аркуатном ядре опять-таки снижен [41].
Вполне вероятно, что KiSS1-нейроны в свою очередь лишь обобщают сигналы других нервных клеток, и новые исследования покажут истинный размах системы, сосредоточенной вокруг ГнРГ-сети [42].
В аркуатном ядре также богато представлена и вторая группа посредников ГнРГ-сети. Речь идет о нейронах проопиомеланокортина (proopiomelanocortin – POMC), которые в зависимости от обстоятельств могут распадаться до бета-эндорфина, адренокортикотропного гормона или меланоцитстимулирующего гормона (альфа-МСГ). Если KiSS1-нейроны смыкают вокруг ГнРГ-сети сигналы пищеварения и гонад, то POMCергические клетки работают с ней на стыке питания и стресса.
О роли эндогенных опиатов в системе «вознаграждения» и в подавлении ГнРГ-сети мы уже говорили выше. Адренокортикотропный гормон управляет надпочечниками, предрасполагая к подъему уровня кортизола, альфа-МСГ, напротив, прямо стимулирует работу ГнРГ-нейронов [4].
Чувствительные к лептину, инсулину и нейропептиду Y POMC-нейроны – хорошие кандидаты для передачи пищевых сигналов ГнРГ-нейронам. И действительно, подобно KiSS1-нейронам, POMC-нейроны распространяют свои окончания к ГнРГ-сети, образуя с ней синаптические контакты [43]. Возможность POMC-нейронов расщепляться на три разнонаправленных продукта является ярким примером гибкой контекстуальной регуляции ГнРГ-сети, а через нее и всей половой системы.
Таким образом, управление половой системой включает минимум три физиологических аспекта: стресс, энергетический баланс и активность гонад. Секреция ГнРГ находится под тяжелым прессом отрицательных влияний, активных при той или иной угрозе выживанию. Некоторые факторы (например, пролактин, бета-эндорфин и эндоканнабиноиды) передают, хотя и в неравной мере, влияние двух аспектов. Популяция KiSS1-ергических нейронов воспринимает сигналы всех типов и является, по-видимому, основным стимулятором ГнРГ-сети. Огромный вес имеют сигналы голода и насыщения, подаваемые ради гибкости управления нескольким аналитическим узлам гипоталамуса.
Затяжной стресс закономерно вызывает дисфункцию половой системы, причем стрессовая нагрузка не всегда бывает болезненной. Нередко стресс является образом жизни, как, например, у балерин и профессиональных спортсменов [10]. Но с точки зрения гипоталамической системы даже менее экстремальная деятельность не всегда располагает к зачатию и беременности.
Общественные и межличностные конфликты, сопровождающиеся агрессией, тревожностью, депрессией, гипоталамус может распознать как отголосок древних видов стресса. К сожалению, психонейроэндокринология все еще находится в начале своего пути становления как науки, и пока мы имеем больше вопросов, чем ответов.
Социально-психологический стресс
Человеческое общество претерпело значительные изменения за последнее тысячелетие. Однако социальное неблагополучие, перемены в образе жизни и информационная перегрузка задействуют те же стрессовые механизмы, что и угроза физическому выживанию [44].
У многих млекопитающих самки ведут между собой борьбу за ресурсы и тех партнеров, что могли бы обеспечить тактические и генетические преимущества будущему потомству. У приматов подчиненное социальное положение вызывает увеличение в плазме кортизола. При этом снижается величина пиковых выбросов лютеинизирующего гормона, необходимых для овуляции [45]. По мере сокращения ресурсов и роста агрессивности внутри сообщества наиболее перспективно зачатие именно у доминирующей самки. Агрессия, в том числе между близкими родственниками, по-видимому, играет важную роль в подавлении овуляции среди подчиненных особей [46].
Есть основания связывать замедление ГнРГ-сети с психологическим неблагополучием и у людей, например с профессиональной перегрузкой, тревожностью, депрессией, негативными установками или фрустрацией [47].
Исследования на животных показывают, что даже относительно мягкие психологические испытания (например, смена жилища) могут нарушать менструальный цикл, если они сочетаются с небольшим дефицитом энергии (аналогом низкокалорийной диеты у людей) [48].
Острые психологические переживания (в экспериментах с иммобилизацией животных) показали быстрое и глубокое подавление секреции лютеинизирующего гормона, что сопровождалось нарастанием в плазме адренокортикотропного гормона и кортизола. При этом введение налоксона – блокатора опиоидных рецепторов – восстанавливало секрецию лютеинизирующего гормона еще до того, как животных выпускали на свободу [49]. Примечательно, что налоксон восстанавливал секрецию лютеинизирующего гормона и у некоторых женщин с аменореей [50].
Эндогенные каннабиноиды, вероятно, также играют роль в нарастании психологического стресса. В краткосрочных условиях это удалось убедительно показать с применением классического теста Триера, который вводит испытуемых в состояние социального стресса, имитируя собеседование при приеме на работу [51].
Психологический стресс нередко вызывает и перемены в питании. Так, при затяжном неблагополучии известно пристрастие к богатой углеводами пище. Возможная причина тому – хроническая кортизоловая стимуляция гипоталамуса [52] при участии все той же каннабиноидной системы «вознаграждения» [53]. Как уже обсуждалось выше, и кортизол, и каннабиноиды подавляют синтез ГнРГ.
Менструальный цикл и микроэкология питания
В предыдущих разделах мы подробно рассмотрели количественные нарушения питания и их тесную связь с высокоуровневыми гипоталамическими процессами. Но картина не была бы полной без качественных характеристик пищи, что особенно важно для женщин с неустойчивой функцией ГнРГ-синтезирующей сети, балансирующих на грани менструальной дисфункции.
Помимо трех основных питательных классов: белков, жиров и углеводов – с каждым приемом пищи человеческий организм получает микроскопические дозы регуляторных веществ, имеющих не питательную, а информационную ценность. Доступность витаминов растительного и животного происхождения исторически включала Homo sapiens в конкретные пищевые цепочки, а микроэлементный состав среды определял благоприятные зоны для обитания. Тем самым витамины и микроэлементы, чье значение часто недооценивается, сопрягают биогеоценоз с индивидуальной физиологией [54]. Женский организм, непрерывно ищущий оптимальные условия для продления рода, не может игнорировать такие данные.
В наши дни человечество распространилось по земному шару повсеместно, нередко и вовсе проводя большую часть жизни в переездах. Так, перед медицинской наукой встала задача компенсировать смену экологических подсистем, что в рыночных условиях доступно лишь путем разработки универсальных адаптивных комплексов. Степень их влияния индивидуальна, поскольку уникален и каждый организм. Однако такие заместительные препараты занимают свою терапевтическую нишу наряду с методами лечения более высокого уровня. Приведем несколько примеров.
Так, фолиевая кислота, необходимая для фундаментальных процессов жизни (удвоения, репарации и эпигенетического управления геномом), содержится преимущественно в зеленых лиственных растениях. Поскольку дефицит фолиевой кислоты наносит наибольший ущерб обновляющимся клеточным популяциям (костному мозгу, иммунной системе, эндометрию и фетоплацентарной системе при беременности), трудно переоценить ее значение. В какой-то мере фолиевую кислоту можно рассматривать как «разрешающий» фактор для поддержания нормального менструального цикла, ведь фолаты – функциональные партнеры эстрогенов, необходимые для своевременного менархе и нормальной пролиферации эндометрия. В прегравидарном периоде запас фолатов определяет оптимальное развитие плода в предстоящей беременности, поэтому их дефицит может отсрочить наступление зачатия. В развитых странах доступ к свежим овощам не является проблемой, но современная культура термической обработки приводит к утрате до 90% фолатов, что остро ставит вопрос об их дотации.
Витамин B6 поддерживает коммуникацию между тканями, включая отделы центральной нервной системы. Он участвует в обмене медиаторов, в том числе гамма-аминомасляной кислоты, катехоламинов, серотонина, простагландинов и гистамина. Организм Homo sapiens адаптировал молекулу B6 для подавления провоспалительных медиаторов, что проявляется падением острофазных показателей воспаления (C-реактивного белка и гомоцистеина). Как мы уже обсуждали выше, гипоталамус чутко реагирует на воспалительный фон торможением репродуктивной функции.
Витамины-антиоксиданты (токоферола ацетат, ретинола ацетат, аскорбиновая кислота) вместе с коэнзимом Q10 и цинком, замыкающими антиоксидантную систему, необходимы для защиты яичников от окислительного стресса, который возникает в момент овуляции. Вместе с кальцием аскорбиновая кислота регулирует в гонадотрофах гипофиза специальный трансмембранный канал, определяя секрецию лютеинизирующего и фолликулостимулирующего гормонов. Дефицит любого из компонентов антиоксидантной системы приводит к снижению фертильности. Например, недостаток токоферола ацетата может стать причиной ановуляции у женщин и азооспермии у мужчин [55]. Известны случаи восстановления фертильности у бесплодных женщин после приема курса витамина Е [56]. Экспериментально на самых разных моделях было показано, что витамин Е играет ключевую роль в защите плода от окислительного стресса [57].
Потребность в железе для кроветворения и в качестве кофактора ряда ферментов отчасти определяет доминирующее положение нашего вида в животных пищевых цепочках. При наличии кровотечений железо должно замещаться, причем ионы Fe должны пребывать в эволюционно обусловленном химическом микроокружении – иначе эффективного всасывания не произойдет. Такое окружение существует в животных тканях.
Наконец, некоторые более редкие растительные факторы могут принести пользу женщинам с гипоталамической аменореей. Например, экстракт витекса священного оказывает мягкое дофаминергическое действие, подавляя выработку стрессового гормона пролактина, нарушающего работу ГнРГ-синтезирующий сети (см. выше).
Однако при функциональной аменорее употребление этих и других микрокомпонентов питания может не привести к успеху. Как мы постарались показать, запретительные «инструкции» исходят от гипоталамуса через многоэтапную высокоуровневую оценку. Однако, работая с функциональным разобщением внешней и внутренней среды женщины (то есть занимаясь коррекцией образа жизни, психотерапией, нормализацией массы тела и регулярности питания и иной патогенетической защитой ГнРГ-синтезирующей сети), не следует забывать и о собственно экологических аспектах, в том числе о микроэкологии питания. В настоящее время разработано множество комплексов, содержащих витамины, макроэлементы, микроэлементы и растительные экстракты. Эти многокомпонентные комплексы предназначены для повышения адаптивных возможностей организма, осуществление конкретных задач предполагает вариабельность их состава.
К примеру, комплекс Миразенова разработан главным образом для уменьшения симптоматики такого дезадаптивного расстройства, как предменструальный синдром. В состав комплекса введены экстракт семян гриффонии (серотонинергическое действие), экстракт витекса священного (дофаминергическое действие), экстракт зверобоя продырявленного (комплексный антидепрессивный эффект), имеющие доказательную базу положительного влияния на функции центральной нервной системы. Комплекс содержит также и другие компоненты (экстракты дудника китайского и лугового клевера, L-фенилаланин, железа глюконат), которые важны не только для хорошего самочувствия женщины, но и для осуществления репродуктивной функции.
Конкретной направленностью на подготовку к процессу репродукции характеризуется комплекс Витазенова, в состав которого входят абсолютно необходимые для нормального наступления и течения беременности витамины группы В, в том числе фолиевая и никотиновая кислоты, витамины А, Е, D, К, биотин, коэнзим Q10, магний, кальций, железо, цинк, марганец, медь и L-таурин. Использование витаминно-минеральных и витаминно-растительных комплексов помогает отчасти разрешить острую современную проблему микроэкологии питания.
Заключение
Несмотря на успехи индустриальной цивилизации, мы живем в эпоху сниженной фертильности [58]. К сожалению, многие особенности современного образа жизни гипоталамус трактует как аналоги первобытных угроз. Вероятно, ожирение имитирует инфекции и голод, увлечение спортом – затяжную миграцию, а конфликты на рабочих местах и в семьях – конкуренцию внутри колонии за ограниченные ресурсы.
Не исключено, что и в случаях метаболического стресса, и при хронических психологических угрозах гипоталамус действует единообразно: он стремится защитить организм, снизив расходы энергии и не допустив рискованной беременности. Это необходимо учитывать у пациенток с функциональной аменореей. В таких случаях одной лишь заместительной терапии может быть недостаточно, поскольку она не влияет на гиперфункцию надпочечников, массу тела и психологические проблемы. Даже при искусственно вызванной овуляции последующей беременности угрожает продолжающийся психологический и/или метаболический стресс. Иными словами, полноценное восстановление половой функции требует коррекции фундаментальной проблемы – образа жизни [10, 59].
Menstrual Cycle and Energy ‘Policy’ of the Hypothalamus
A.S. Gorelyshev1, I.V. Kuznetsova2
1 I.M. Sechenov First Moscow State Medical University
2 Endocrinology Research Center
Contact person: Aleksandr Sergeyevich Gorelyshev, alexander.gorelyshev@gmail.com
Prolongation of progeny requires to be perfectly selected. Pregnancy is vulnerable to physical, social and energetic issues, explaining a reason why sex system in mammalians is dominated by excessive negative signals. A selection of physiological conditions suiting to conception must be performed by a specialized analytical organ in the body. Such role is played by hypothalamus that unites both nervous and endocrine systems. In different countries, natural obstacles to the childbirth such as famine, long-lasting diseases and physical exhaustion have been overcome. So, why we have now more and more women suffering from impaired menstrual cycle and infertility? Here, by using neuroendocrine and anthropological viewpoints, we discuss functional amenorrhea and idiopathic infertility. A hypothesis has been developed suggesting that improper lifestyle simulates primordial threats making hypothalamus to “switch off” sexual function for saving energy as it was at the dawn of human history. A concept of nutritional microecology as a method for optimizing lifestyle is proposed in the context of the abovementioned hypothesis.