Моделирование процесса регуляции времени сна по будням и выходным: опровержение мифов, связанных с социальным десинхронозом
- Аннотация
- Статья
- Ссылки
- English
Материал и методы. Для симуляции времени сна в будни и выходные были использованы 190 выборок из литературы (средний возраст испытуемых варьировался от полугода до 60 лет). Применялась классическая двухпроцессная модель регуляции сна и бодрствования, предложенная S. Daan и соавт. в 1984 г., в модификации автора 1995 г. Кроме того, для подтверждения некоторых из результатов первых 190 выборок использовались данные 117 дополнительных выборок из публикаций последних двух лет.
Результаты. Анализ эмпирических данных и их симуляция не поддержали ни одно из представлений, бытующих вокруг явления социального десинхроноза, в частности, что ранние пробуждения по будням приводят к аккумуляции долга сна и сон, потерянный из-за ранних пробуждений в эти дни, можно хотя бы частично компенсировать, отоспавшись в выходные. Подтверждена ошибочность мнения, что время начала и окончания сна в выходные дни можно использовать для идентификации «сов» и «жаворонков» и социальный десинхроноз наиболее выражен у «сов». Признано неверным допущение, что биологические часы начинают запаздывать в подростковом возрасте (что следует из запаздывания времени сна в выходные), но затем начинают снова спешить по мере дальнейшего взросления и старения. Наконец, опровергнуто мнение, что в течение недели биологические часы остаются синфазными 24-часовому режиму естественного освещения и только фаза цикла «сон – бодрствование» сдвигается вперед-назад.
Выводы. Очевидно, что представления о хронобиологических механизмах, лежащих в основе явления социального десинхроноза, нуждаются в основательном пересмотре.
Материал и методы. Для симуляции времени сна в будни и выходные были использованы 190 выборок из литературы (средний возраст испытуемых варьировался от полугода до 60 лет). Применялась классическая двухпроцессная модель регуляции сна и бодрствования, предложенная S. Daan и соавт. в 1984 г., в модификации автора 1995 г. Кроме того, для подтверждения некоторых из результатов первых 190 выборок использовались данные 117 дополнительных выборок из публикаций последних двух лет.
Результаты. Анализ эмпирических данных и их симуляция не поддержали ни одно из представлений, бытующих вокруг явления социального десинхроноза, в частности, что ранние пробуждения по будням приводят к аккумуляции долга сна и сон, потерянный из-за ранних пробуждений в эти дни, можно хотя бы частично компенсировать, отоспавшись в выходные. Подтверждена ошибочность мнения, что время начала и окончания сна в выходные дни можно использовать для идентификации «сов» и «жаворонков» и социальный десинхроноз наиболее выражен у «сов». Признано неверным допущение, что биологические часы начинают запаздывать в подростковом возрасте (что следует из запаздывания времени сна в выходные), но затем начинают снова спешить по мере дальнейшего взросления и старения. Наконец, опровергнуто мнение, что в течение недели биологические часы остаются синфазными 24-часовому режиму естественного освещения и только фаза цикла «сон – бодрствование» сдвигается вперед-назад.
Выводы. Очевидно, что представления о хронобиологических механизмах, лежащих в основе явления социального десинхроноза, нуждаются в основательном пересмотре.
Введение
Когда на протяжении рабочей или учебной недели приходится вставать пораньше, то ночной сон становится короче желаемого. В конце такой недели хочется компенсировать (хотя бы частично) потерянный сон, поспать подольше в выходные и благодаря некоторому «пересыпу» вернуться в нормальное состояние. Действительно, если удается поспать подольше в ночь с пятницы на субботу и/или с субботы на воскресенье, человек чувствует себя намного лучше, чем в будни. В чем ошибочность подобного рассуждения? В допущении того, что утерянный сон можно вернуть хотя бы частично.
Поразительно, что эта мысль, вполне естественно возникающая на бытовом уровне, нашла воплощение в научной литературе под названием социального десинхроноза (social jet lag). Правда, это произошло сравнительно недавно (в 2006 г.) и, что уже само по себе подозрительно, намного позднее оформившейся в начале 1960-х гг. хронобиологии в качестве отдельной научной области, которая исследует биологические ритмы, то есть уже после научной фиксации всех основных изучаемых в ее рамках феноменов. К ним, в частности, относится явление собственно десинхроноза (jet lag), под которым понимается временное рассогласование фаз циркадианных (околосуточных) ритмов организма в процессе их вынужденного сдвига вслед за сдвигом фазы внешнего 24-часового синхронизирующего воздействия. Концепцию социального десинхроноза предложили M. Wittmann и соавт., которые определили его как специфическую (добавлю – видимо, ранее никому не известную) форму десинхроноза в результате рассогласования между временем работы/учебы и предпочитаемым временем сна и бодрствования. Последнее зависит от внутренних биологических часов, которые в свою очередь контролируются 24-часовым режимом освещения, а не какими-либо социальными сигналами времени, включая часы работы/учебы [1].
Исходя из такого определения, авторы пришли к выводу, что социальный десинхроноз наиболее выражен у позднего хронотипа (типа суточного ритма), то есть у «сов» [1]. Они вынуждены приспосабливать временные привычки к социальным требованиям. Иначе говоря, «совам» приходится вставать рано и они просто не способны сдвинуть на более раннее время начало своего сна, за которое отвечают внутренние часы, в свою очередь контролируемые 24-часовым режимом освещения. Таким образом, другая идея авторов состоит в том, что «совы» вынуждены отсыпаться на выходных дольше, чем «жаворонки» [1]. Соответственно они в большей степени страдают от социального десинхроноза, то есть от фазового рассогласования между временем сна и фазой собственных биологических часов. И, если верить авторам статьи, «совы» компенсируют в выходные недельный долг сна, который у них выше, чем у «жаворонков» [1].
T. Roenneberg и соавт. утверждают, что хронотип – специфическое для индивидуума отношение между фазой внутренних часов, индикаторами которой являются фазы циркадианных ритмов температуры тела и мелатонина, и внешним синхронизатором, например временем восхода и захода солнца [2]. По их мнению, с учетом этого отношения дети обычно имеют ранний хронотип, но, постепенно сдвигая время сна и достигая максимума в возрасте около 20 лет, переходят к позднему хронотипу, затем по мере дальнейшего взросления нередко возвращаясь к раннему хронотипу [3]. При этом заметим, речь идет уже о фазе сна, тогда как маркеры фазы биологических часов (циркадианные ритмы температуры тела и мелатонина) в этих работах, основанных на опроснике, не измерялись [3].
Хронобиология – фундаментальная и, можно даже сказать, точная наука, поскольку в основе всех ее теоретических представлений лежит математическая теория колебаний. Следовательно, объяснить какое-либо изучаемое хронобиологами явление можно только путем его описания и количественного предсказания в терминах этой теории. Например, можно предсказать временнóе поведение организма (в частности, время сна в будни и выходные) путем количественной симуляции с помощью математической модели регулятора конкретного циркадианного ритма (в данном случае цикла «сон – бодрствование»). Удивительно, но никто за более чем десяток лет не попытался применить такой традиционно-хронобиологический подход к тестированию явления социального десинхроноза. Поэтому в двух предыдущих статьях [4, 5] мы с коллегами использовали предложенную мной ранее модель регуляции цикла «сон – бодрствование» [6] для количественной симуляции различий по времени сна между разными хронотипами [5] и возрастами [4]. В качестве эмпирических данных были взяты опубликованные в литературе сведения о времени отхода ко сну и подъема по будням и в выходные дни для 190 выборок (всего четыре показателя). Еще 117 выборок дополнительно собраны из новых работ 2018 и 2019 гг. и использованы для подтверждения результатов, полученных ранее [4, 5].
Цель исследования
Проверить правдоподобие восьми допущений, вытекающих из концепции социального десинхроноза.
- Ранние пробуждения по будням приводят к аккумуляции долга сна.
- Сон, потерянный в будние дни из-за ранних пробуждений, можно хотя бы частично компенсировать, отоспавшись в выходные дни.
- В целом время начала и окончания сна в выходные дни можно использовать для идентификации «сов» и «жаворонков».
- Социальный десинхроноз наиболее выражен у «сов».
- Биологические часы (фаза внутреннего синхронизатора циркадианных ритмов организма) начинают запаздывать в подростковом возрасте (что следует из запаздывания времени сна в выходные).
- В дальнейшем биологические часы начинают снова спешить по мере взросления и старения.
- В течение недели биологические часы остаются синфазными 24-часовому режиму естественного освещения.
- Только фаза цикла «сон – бодрствование» сдвигается вперед-назад в течение недели.
Материал и методы
Для симуляции циклов «сон – бодрствование» в будни и выходные была использована модель ритмостата [6], вариант классической двухпроцессной модели регуляции сна и бодрствования, предложенной S. Daan и соавт. [7]. Особенность этого варианта заключается в постулировании модулирующего эффекта биологических часов (синусоидальная функция с 24-часовым периодом) [6] на параметры собственно сомностата (чередование обратно-экспоненциальной и экспоненциальной функций, представляющих фазы бодрствования и сна в цикле «сон – бодрствование» соответственно) [7]. Если t1 – время отхода ко сну, а t2 – время подъема (начальные временные значения), то процесс регуляции сна и бодрствования описывается следующими уравнениями:

периодическая функция с периодом τ, приравненным к 24 часам [6].
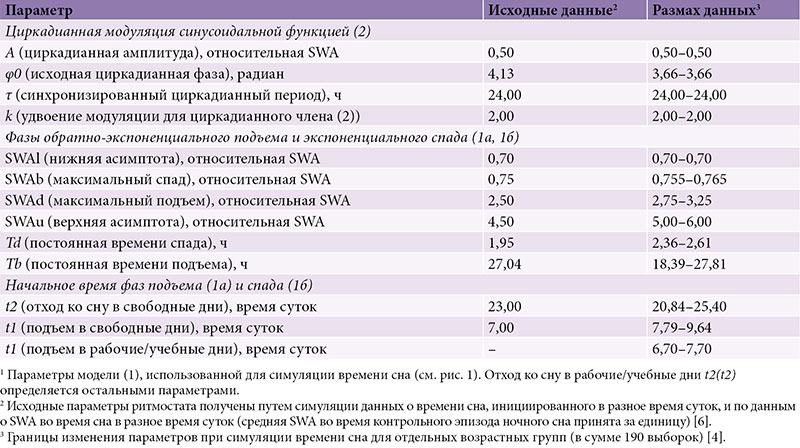
В таблице 1 перечислены исходные параметры, полученные при симулировании времени сна, инициированного на разных фазах циркадианного ритма, и уровня медленноволновой активности (Slow-Wave Activity – SWA) в экспериментах со сном, следующим за различными интервалами предшествующего ему бодрствования (более детально это рассматривается в работе A.A. Putilov 1995 г. [6]). Симуляции позволили выразить X в относительных единицах SWA (см. табл. 1) [6].
Эмпирические значения продолжительности сна в выходные (8,9 ± 0,4 часа) оказались близкими к максимальной емкости сна в экспериментах с людьми примерно того же возраста, по данным E. Klerman и D. Dijk (2008) [8]. По этой причине при симуляции было сделано допущение, что время подъема и отхода ко сну (t1 и t2) в выходные, равно как и время отхода ко сну по будням, определяется исключительно ритмостатом. Четвертый показатель (подъем в будни) задается не ритмостатом, а внешним влиянием.
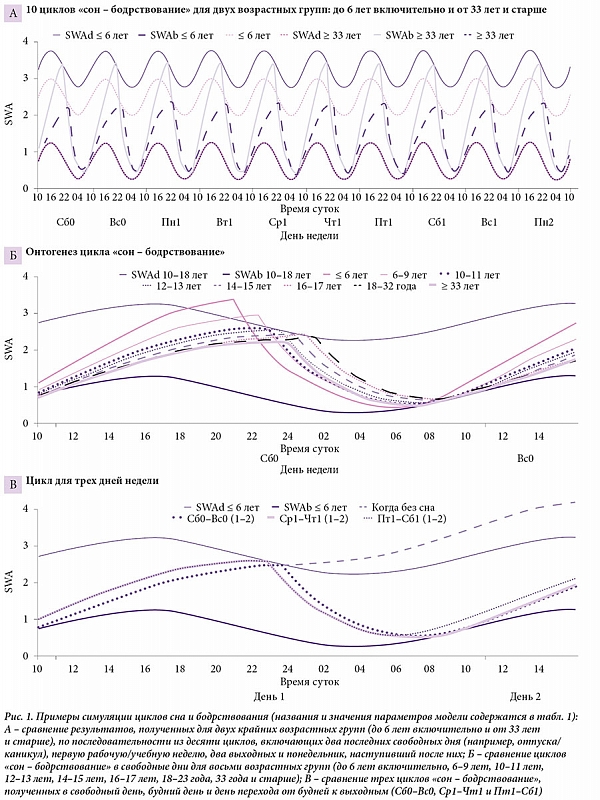
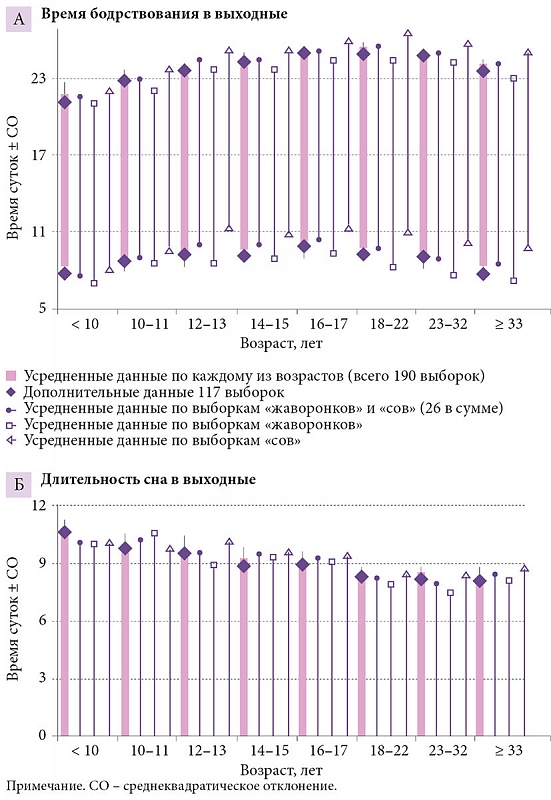
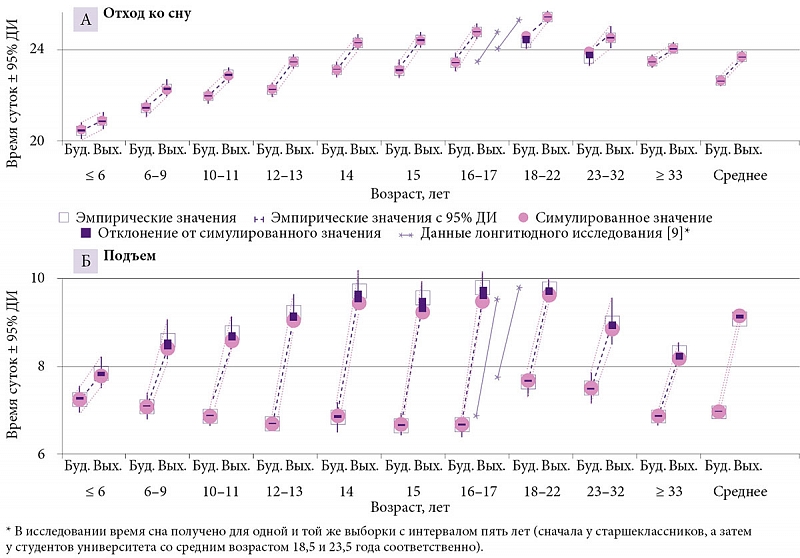
Для простоты и ясности эмпирические значения брались для симуляции с округлением до первого знака. Например, усредненное по 190 выборкам (средний возраст 17,0 года) время начала и окончания сна было принято как 9,1 и 23,7 часа (t1 и t2), а подъем в будни приравнен к 7,0 часа (табл. 2). В таблице 1 представлена область значений параметров, полученных при симуляции данных о времени сна в восьми возрастных группах. Разница между показателями, полученными эмпирически и путем симуляции этих эмпирических данных, указана в табл. 2 (для усредненных данных и данных по разным возрастам или хронотипам соответственно). Симулированные циклы «сон – бодрствование» представлены на рис. 1, а эмпирические данные – на рис. 2 и 3. Новые эмпирические данные и результаты их симуляции добавлены к ранее симулированным данным в рис. 2.
Результаты
Опровержение первого и второго допущений о том, что ранние пробуждения по будням приводят к аккумуляции долга сна и сон, потерянный в будние дни из-за ранних пробуждений, можно хотя бы частично компенсировать, отоспавшись в выходные дни
На рисунке 1 представлены результаты симуляции циклов «сон – бодрствование» в будни и выходные при допущении, что аккумуляция долга сна в течение недели попросту невозможна. Как видно на рис. 1А, подъем SWA во время бодрствования никогда не выходит за верхнюю границу, которая задается ритмостатом, то есть исключительно изнутри. Рисунок 1В дополнительно иллюстрирует, что произойдет с этой активностью – индикатором накопления долга сна в случае перехода через границу при продолжении бодрствования после навязываемого ритмостатом времени отхода ко сну в поздневечерние часы (то есть при накоплении долга сна). В силу понижающего влияния циркадианной модуляции на рост SWA (спад синусоиды на графике) активность почти не возрастает в течение всей ночи, то есть долг практически не аккумулируется. Более того, если сон начнется после прохождения границы, он обязательно окажется не длиннее, а, наоборот, короче сна, начинающегося на границе, в силу того, что в течение такого запоздалого сна понижающее влияние циркадианного ритма сменится повышающим влиянием (подъем синусоиды на графике). Вместо дополнительного сна после удлинения времени бодрствования человек получит его укорочение. Следовательно, допущения, вытекающие из концепции социального десинхроноза (ранние пробуждения по будням приводят к аккумуляции долга сна и сон, потерянный в будние дни из-за ранних пробуждений, можно хотя бы частично компенсировать, отоспавшись в выходные дни), не подтверждаются результатами симуляции данных с помощью модели. Более того, результат не изменится даже в случае принятия каких-либо дополнительных допущений (например, если предположить, что человек не слушается своего ритмостата и продолжает бодрствовать в ночное время либо в течение одного-двух часов после ожидаемого отхода ко сну, либо вообще всю ночь).
Точность предсказаний времени сна для усредненного набора данных была высокой. В большинстве случаев различия между эмпирическими и симулированными значениями были меньше таковых, возникающих при округлении эмпирических значений для их дальнейшей симуляции. Особенно точно модель предсказывала время отхода ко сну в будни (нулевая разница). Из этого, в частности, следует, что если в качестве «ввода» в модель используется время, когда люди обычно ложатся спать и встают в выходные дни и когда они вынуждены вставать в будни, то в качестве «вывода» модели предсказывается точное время, когда они обычно ложатся спать в будни.
Опровержение третьего допущения о том, что в целом время начала и окончания сна в выходные дни можно использовать для идентификации «сов» и «жаворонков»
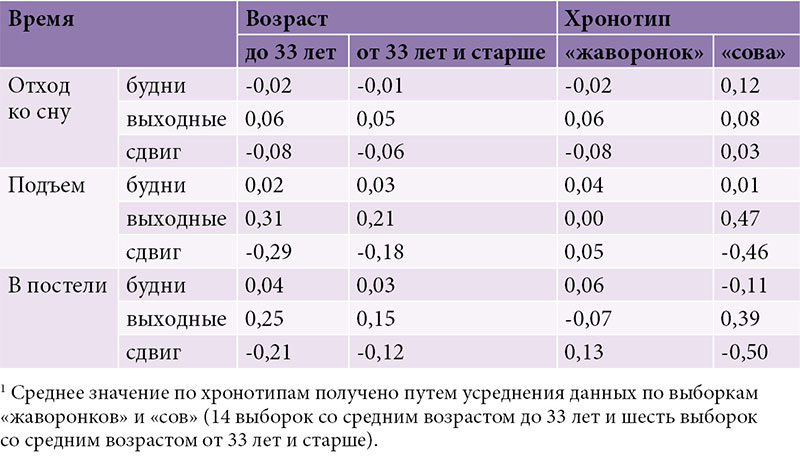
Рисунок 2 позволяет сравнить время бодрствования и сна у «сов» и «жаворонков» (26 выборок в целом, из них 20 из исходного набора в 190 выборок, которые были симулированы). Как по всем выборкам, так и по отдельным возрастным группам время сна в исходном наборе из 190 выборок существенно не отличалось от такового в дополнительном наборе из 117 выборок. При этом значения, усредненные по хронотипам, существенно не отличались от значений, усредненных по всем выборкам. Разница между «совами» и «жаворонками» состояла не в длительности сна (рис. 2Б), а почти исключительно во времени начала/окончания сна, которое у «сов» запаздывало минимум на час относительно такового у «жаворонков» (рис. 2А). Нетрудно заметить, что «совы» первой возрастной группы соответствовали «жаворонкам» второй, а «совы» второй возрастной группы в свою очередь соответствовали «жаворонкам» третьей. Иными словами, нет никаких оснований думать, что в целом время начала и окончания сна в выходные дни можно использовать для идентификации «сов» и «жаворонков». Возрастная изменчивость времени сна слишком велика, чтобы позволить делать это таким нехитрым путем.
Опровержение четвертого допущения о том, что социальный десинхроноз наиболее выражен у «сов»
В целом данные о времени сна у «сов» и «жаворонков» в выходные, полученные для отдельных групп, не имеют общей тенденции удлинения продолжительности сна (отражающей стремление отоспаться) у первых относительно вторых. Более того, разница во времени подъема и длительности сна по будням между хронотипами обнаруживает достоверный возрастной тренд. Разница по времени подъема близка к нулю только тогда, когда все дети/подростки, независимо от хронотипа, вынужденно ходят в садик или школу, а начиная со старших классов разница между хронотипами постепенно увеличивается, достигая нескольких часов в пожилом возрасте. В результате знак различия между хронотипами в длительности сна в будни с возрастом меняется на противоположный, то есть, как это ни парадоксально звучит, по достижении возраста Христа именно «жаворонки», а не «совы» начинают терять больше сна в будни из-за ранних пробуждений. Поэтому представление о том, что социальный десинхроноз наиболее выражен у «сов», также неверно.
Опровержение пятого и шестого допущений о том, что биологические часы (фаза внутреннего синхронизатора циркадианных ритмов организма) начинают запаздывать в подростковом возрасте (что следует из запаздывания времени сна в выходные) и в дальнейшем начинают снова спешить по мере взросления и старения
В целом различие между хронотипами отсутствует, а кажущееся очевидным представление о существенном различии возникает из-за того, что исследования, как правило, проводятся на подростках. У них это явление ассоциируется с наибольшими жизненными трудностями, поскольку в старших классах сон сдвигается на позднее время, но все школьники вынуждены приходить в школу к одному и тому же часу, независимо от предпочитаемого времени сна.
Рисунок 3 иллюстрирует влияние раннего начала уроков в школе на различия между временем сна в будни и выходные. На границе 18–22-летнего возраста в графики специально добавлены данные (в виде двух отдельных линий) лонгитюдного исследования, выполненного M. Urner и соавт. (2009) [9], в котором значения времени сна были получены для одной и той же выборки с интервалом пять лет. Сначала участники опроса были старшеклассниками, а потом студентами университета. Эти данные напоминают результаты, полученные при усреднении всех выборок по двум соседствующим возрастным группам. Видимо, общая тенденция такова, что, освободившись от диктата школы, молодые люди начинали вставать в будни в среднем на час позже, чем во время учебы в старших классах. На рисунке 3 этот социально, а не биологически обусловленный сдвиг заметно искажает в целом плавное возрастное изменение времени сна в выходные, но подобная аномалия не помешала определить общие возрастные тенденции изменения параметров процессов регуляции времени сна с помощью модели (1).
Как иллюстрирует рис. 1Б, симуляция была проведена с допущением, что ни один из параметров циркадианной модуляции времени сна не меняется в течение всей жизни. В частности, циркадианная фаза остается одной и той же. Более того, было также принято допущение, что SWA остается неизменной в подростковом возрасте (хотя на самом деле она снижается и в детстве, и в подростковом возрасте, и по мере дальнейшего взросления, а потом и старения). Такие допущения позволили продемонстрировать, что возрастные сдвиги времени сна при переходе от детского к взрослому возрасту можно объяснить изменением всего одного параметра процесса регуляции цикла «сон – бодрствование» – постоянной времени для фазы бодрствования, то есть для фазы подъема SWA (Tb). Другие подробности моделирования возрастных различий можно найти в работе A. Putilov и E. Verevkin [4]. Для целей данной статьи важно отметить, что возможность симуляции возрастных изменений времени сна без постулирования каких-либо параллельных возрастных изменений фазы циркадианного влияния на сон находится в противоречии с допущениями авторов концепции социального десинхроноза о том, что биологические часы (фаза внутреннего синхронизатора циркадианных ритмов организма) начинают запаздывать в подростковом возрасте (что следует из запаздывания времени сна в выходные), но затем принимаются снова спешить по мере дальнейшего взросления и старения.
Опровержение седьмого допущения о том, что в течение недели биологические часы остаются синфазными 24-часовому режиму естественного освещения
Различия фазы циркадианной модуляции были постулированы только при симуляции различий во времени сна между «совами» и «жаворонками» (подробности можно найти в работе A. Putilov и соавт. [5]). Моделирование предсказывает отсутствие каких-либо существенных различий по длительности сна между двумя хронотипами. Как видно из табл. 2, расхождение между эмпирическими и симулированными значениями было минимальным для «жаворонков», тогда как «совы» указывали более позднее время подъема в выходные, чем время, предсказанное моделью. При этом расхождение было более выраженным у лиц моложе 33 лет. Кроме того, как видно на рис. 3, более позднее время подъема в выходные по сравнению с тем, что предсказала модель, наблюдалось и в старшем подростковом возрасте. Одно из возможных объяснений обнаруженного расхождения (о других расхождениях можно прочесть в работах A. Putilov и E. Verevkin [4], A. Putilov [5]) – сдвиг фазы циркадианного ритма под влиянием искусственного вечернего освещения у людей с поздним отходом ко сну, что позволяет этим людям спать дольше в выходные дни (свет вызывает запаздывание фазы, не учтенное в симуляции). Затем в рабочие/учебные дни фаза может сдвигаться на ту же величину вперед из-за влияния утреннего освещения, что вполне вероятно из-за раннего подъема. Поэтому для людей с поздним отходом ко сну может быть неверным представление о том, что в течение недели биологические часы остаются синфазными 24-часовому режиму естественного освещения. И эти люди могут еженедельно испытывать десинхроноз в его самой обычной форме.
Опровержение восьмого допущения о том, что только фаза цикла «сон – бодрствование» сдвигается вперед-назад в течение недели
Казалось бы, предположение о том, что только фаза цикла «сон – бодрствование» сдвигается вперед-назад в течение недели, трудно опровергнуть. Однако оно основано исключительно на факте, что время сна сдвигается сначала на ранние часы в будни, а потом на поздние часы в выходные. Модель отражает не только время сна, но и динамику ритмического процесса, лежащего в основе цикла «сон – бодрствование». Его индикатором считаются колебания SWA в этом цикле – обратно-экспоненциальный рост и экспоненциальный спад (см. рис. 1). На рисунке 1В видно, что в течение всей недели меняется только форма волны цикла колебаний SWA. Если в качестве его фазы выбрать уровень SWA накануне подъема в будни, то есть семь часов утра (фаза – это любая произвольно выбранная точка цикла), то эта утренняя фаза оказывается практически одинаковой по уровню SWA и в любом из циклов. Иными словами, у подавляющего большинства людей она не сдвигается в течение недели.
Обсуждение результатов
Симулирование времени сна в выходные и по будням с помощью модели регуляции цикла «сон – бодрствование» позволяет усомниться в достоверности любого из бытующих в научных кругах с 2006 г. представлений о явлении социального десинхроноза. Они лишь на первый взгляд кажутся очевидными, но их нельзя объяснить с позиций хронобиологической теории, базирующейся на математическом знании. Поскольку для симуляций были доступны только данные о времени сна, а данные о маркерах фазы циркадианного ритма для подавляющего большинства выборок отсутствовали, необходимо было поддержать результаты моделирования теми немногими литературными источниками, в которых наряду со временем сна приведены результаты измерения циркадианной фазы на примере ритмов температуры тела или мелатонина.
Например, в лонгитюдном исследовании S. Crowley и соавт. (2014) фаза начала синтеза мелатонина оставалась неизменной при переходе от младшего к старшему подростковому возрасту, хотя при этом наблюдалась типичная для такого перехода картина запаздывающего сдвига времени сна в выходные [10]. К этому можно добавить два уточнения. Во-первых, обратный (опережающий) сдвиг времени сна по мере дальнейшего взросления и старения начинается раньше, чем обнаруживаются какие-либо признаки опережающего сдвига фазы ритмов-маркеров циркадианного ритма, а опережающий сдвиг этой фазы, наблюдаемый в глубокой старости, может быть объяснен другими причинами (более подробно это рассмотрено в работе A. Putilov 2016 г. [11]). Во-вторых, циркадианный период не меняется с возрастом, что было обнаружено как при сравнении подростков с взрослыми, так и при сравнении взрослых с пожилыми людьми [4, 5, 11].
К большому удивлению, публикации, в которых циркадианная фаза измерялась бы сначала в будни, а затем в выходные, практически отсутствуют. По крайней мере, единожды это было сделано в диссертационной работе G. Zerbini (2017), в числе руководителей которой оказались авторы статьи по социальному десинхронозу [12]. Автор обнаружил сдвиг фазы начала секреции мелатонина у подростков с поздним хронотипом в выходные дни, причем и зимой, и летом, чего не наблюдалось у подростков с ранним и промежуточным хронотипом. Поэтому приведенное выше объяснение различий между эмпирическим и предсказанным временем подъема в выходные дни у людей с поздним хронотипом может быть вполне правдоподобным.
Выводы
Из сказанного следует, что представления о хронобиологических механизмах, лежащих в основе явления социального десинхроноза, нуждаются в основательном пересмотре. По сути, для большинства людей это не десинхроноз, а не более чем безвозвратное укорочение привычного времени сна. Каких-либо последствий такого укорочения для циркадианного и гомеостатического компонентов процесса регуляции цикла «сон – бодрствование» возникнуть просто не может (в частности, в форме накопления в течение рабочей/учебной недели долга сна с его дальнейшей, пусть даже частичной, компенсацией в выходные).
Исследование выполнено при финансовой поддержке РФФИ («Оценка различий между четырьмя крайними хронотипами в параметрах колебаний объективных показателей сонливости и работоспособности при пролонгированном бодрствовании», № 19-013-00424-а).
Автор признателен к.б.н. Евгению Георгиевичу Веревкину за выполнение в программе Microsoft Excel симуляций для данной статьи.
A.A. Putilov, DBSci, PhD
Institute of Higher Nervous Activity and Neurophysiology of the Russian Academy of Sciences, Moscow
Federal Research Centre for Fundamental and Translational Medicine, Novosibirsk
Contact person: Arcady A. Putilov, putilov@ngs.ru
Purpose. People believe that, by extending sleep on the weekend, they can compensate its reduction during the preceding weekdays. This belief, however, has never been supported with simulations based on a model of sleep-wake regulation. Therefore, such a model was applied to debunk the myth of catchup weekend sleep along with other myths around the phenomenon of ‘social jet lag’ (a misalignment between work hours and preferred sleep times governed by the body clocks entrained to the 24-hour light-dark cycle rather than to any social time cue, e.g., work hours).
Material and methods. To simulate sleep times reported in the literature for 190 samples (mean ages vary from 0.5 to 60 years), A. Putilov (1995) version of the classical two-process model of sleep-wake regulation S. Daan et al. (1984) was used, and additional 117 samples were collected from the most recent publications to confirm the results.
Results. The empirical analysis of sleep times and their simulations suggested that sleep debt cannot be accumulated after early morning awakenings on weekdays, and sleep lost on weekdays due to such scheduled early wakeups cannot be repaid; in general, clock times for weekend sleep cannot be solely used to distinguish between ‘larks’ and ‘owls’, and ‘social jet lag’ cannot be more prominent in ‘owls’ than ‘larks’ the circadian phase cannot delay in adolescents following the delay of their sleep times, and it cannot advance back despite advancing sleep times in middle-aged adults; a sleep phase determined by level of electroencephalogram slow wave activity in the morning hours remains rather stable throughout the week, while the circadian phase in ‘owls’ can be shifted back-and-forth relative to natural light regimen throughout the week.
Conclusion. It seems that the chronobiologial mechanisms underlying ‘social jet lag’ require rethinking.
