Характеристики ЭЭГ при возбуждениях мозга во время второй стадии дневного сна, сопровождающихся самопроизвольными микропробуждениями и минимальной поведенческой активностью в ситуации выполнения психомоторного теста
- Аннотация
- Статья
- Ссылки
- English


Введение
Изучение возбуждений мозга в состоянии сна, которые заканчиваются двигательной активностью и пробуждениями, важно для оценки нестабильности этого состояния в условиях влияния на него социальных и экологических факторов [1], при исследовании расстройств возбуждений во время сна, которые приводят к непроизвольной двигательной активности различной длительности и сложности [2]. Оно также может быть полезно специалистам при оценке рисков, которые вызывают сознательные реакции субъекта (в том числе сновидения) во время анестезии и после операции [3].
В исследованиях внутримозговых записей биоэлектрической активности пациентов с эпилепсией в период ремиссии показано, что переходы коры от сна к бодрствованию носят продолжительный характер и пробуждения можно рассматривать как разные состояния мозга. Таламическая активность единообразна и соответствует состоянию между бодрствованием и сном. При этом активация коры неоднородна и обусловлена разными факторами регуляции. Ее спектральный состав зависит от модальности пробуждения (спонтанная или индуцированная), гомеостаза, стадии сна и области отведения [4, 5]. Эти состояния сопровождаются таким явлением, как локальное бодрствование, когда на фоне активности, похожей на сон, наблюдаемой в большинстве корковых областей мозга, появляются паттерны, соответствующие состоянию пробуждения [6]. В этих паттернах отмечается более выраженное присутствие высокочастотных колебаний электроэнцефалограммы (ЭЭГ), которые либо замещают низкочастотные, либо накладываются на них [7]. Это явление может отражать сенсомоторную обработку внешних раздражителей и готовить мозг к пробуждению [8] или же сопровождать двигательную активность без ее осознавания [2, 9, 10].
В исследованиях ЭЭГ во время эпизодов спонтанных пробуждений во время ночного сна, которые испытуемые подтверждали нажатиями на кнопку, было показано, что в большинстве случаев этим нажатиям предшествовал период появления в биоэлектрической активности альфа-ритма [11]. Особенность нашей работы состоит в том, что мы исследуем активации мозга с последующим коротким эпизодом самопроизвольного пробуждения, сопровождающегося восстановлением выполнения психомоторного теста, прерываемого сном. Мы предполагаем, что различия в поведении (в числе нажатий на кнопку) и ЭЭГ, которая предшествует и сопровождает это поведение, позволят оценить состояние субъекта и изменения уровня сознания в этих обстоятельствах [12]. Ранее нами были приведены доказательства, которые позволяют предположить, что даже при небольшом числе нажатий психомоторная активность в этот период не является неосознаваемой, а сопровождается сниженным уровнем сознания [13]. Мы решили продолжить нашу работу: исследовать ЭЭГ субъектов во время микропробуждений, которые сопровождаются единичными нажатиями, и сопоставить ее с ЭЭГ тех же субъектов в ситуации с коротким паттерном нажатий (2–6).
Материал и методы
Исследовали 19 взрослых здоровых испытуемых (трое мужчин и 16 женщин, студенты, правши, средний возраст – 21,26 ± 0,89 года) без диагностированных неврологических, психиатрических заболеваний и нарушений сна. Всех испытуемых просили не употреблять алкоголь в течение суток, а также чай и кофе в течение шести часов перед опытом. Исследование было одобрено этической комиссией Института высшей нервной деятельности и нейрофизиологии РАН (протокол № 2 от 3 июня 2019 г.) и соответствовало этическим нормам Хельсинкской декларации Всемирной медицинской ассоциации «Этические принципы проведения научных медицинских исследований с участием человека» с поправками 2000 г. Состояние перед опытом оценивалось с помощью дневника сна [14] и опросника уровня сонливости KSS. Субъекты спали в ночь перед сном в среднем М = 6,46 ± 0,46 часа. Качество сна и самочувствие при пробуждении по пятибалльной шкале (5 – «отличное»): М = 4,0 ± 0,19 и М = 3,32 ± 0,27 соответственно. Отметим высокий уровень сонливости: М = 5,42 ± 0,41 (по семибалльной шкале, 7 – «очень высокий»).
Эксперимент начинали в период 12:00–13:00 часов, длительность его – 1,5–2 часа. В течение всего опыта испытуемый находился в затемненном звукозаглушающем помещении, на кушетке, с закрытыми глазами. С помощью психомоторного теста [12] в его унимануальной модификации получали ряд последовательных периодов засыпания – пробуждения. Тест состоит из двух чередующихся фаз. Испытуемые считали про себя от 1 до 10, одновременно при каждом отсчете они нажимали на кнопку большим пальцем правой руки, зафиксированную на указательном пальце той же руки (первая фаза). Далее они продолжали считать про себя от 1 до 10, но уже без нажатий (вторая фаза). Чередование этих фаз длилось до тех пор, пока испытуемые не засыпали. При самопроизвольных пробуждениях они возобновляли выполнение теста с первой фазы.
Регистрировали ЭЭГ от 17 отведений в соответствии со схемой 10–20, электроокулограмму, электромиограмму короткой мышцы, отводящей большой палец кисти правой руки, и механограмму нажатий на кнопку. Регистрацию проводили с помощью системы Neocortex-Pro (Neurobotics). Частота дискретизации – 250 Гц, полоса пропускания частот: 0,5–70 Гц. Использовали шлем и хлорсеребряные электроды, сопротивление – меньше 5 кОм.
Для анализа выбирали безартефактные отрезки полисомнограмм, соответствующие эпизодам самопроизвольного короткого пробуждения из второй стадии ортодоксального сна (N2) с частичным выполнением первой фазы психомоторного теста. Длительность N2 должна была быть не менее 1,5 минуты, оценка принадлежности полисомнограмм к N2 проводилась визуально по критериям AASM (2023 г.). У каждого испытуемого выбирали по два отрезка полисомнограмм – с единичным нажатием и с паттерном (2–6 нажатий). Если отрезков с тем или иным числом нажатий было несколько, то предпочтение отдавалось парам, наиболее близким по времени. Пример полисомнограммы с пробуждением и частичным выполнением психомоторного теста приведен в работе [13].
Анализировали 40-секундные отрезки записи ЭЭГ, на середину которых приходились нажатия на кнопку: по 20 секунд до и после начала нажатий. Для оценки мощностных характеристик биоэлектрической активности коры проводили непрерывное вейвлет-преобразование на основе материнского комплексного Morlet-вейвлета (Matlab 78.01). Карты распределения значений модуля коэффициента вейвлет-преобразования (КВП) строили в полосе 0,5–40 Гц с шагом 0,5 Гц и разрешением по времени 0,01 секунды. Проводили усреднение КВП по частоте в диапазонах 0,5–3,5, 4–7,5, 8–10,5 и 11–14,5, 15–20,5 и 21–40 Гц (дельта-, тета-, низкочастотный и высокочастотный альфа-ритмы, бета- и низкочастотный гамма-ритм), далее усредняли по времени, в двухсекундных интервалах.
Полученные амплитудно-мощностные характеристики ЭЭГ анализировали с помощью дисперсионного анализа (ANOVA RM). Рассматривали влияние на них факторов: «число нажатий» – два уровня (единичное нажатие и паттерн нажатий), «поведенческая активность» – два уровня (периоды – до начала нажатий на кнопку и после) и «время» (10 уровней – 10 двухсекундных интервалов времени). ANOVA RM проводили как суммарно (то есть по усредненным мощностным характеристикам всех отведений ЭЭГ для каждого отсчета), так и по отдельности для каждого отведения ЭЭГ. Статистические оценки проводили с учетом поправки Гринхауза – Гессера. С помощью парного критерия Стьюдента полученные в результате усреднений по всем отведениям ЭЭГ амплитудно-мощностные характеристики сравнивали между ситуациями с единичным нажатием и паттерном нажатий – по каждому выделенному двухсекундному интервалу и в каждом из спектральных диапазонов. По результатам этого сравнения выделяли временные интервалы с различающимися характеристиками. Внутри этих интервалов для каждого отведения проводили аналогичное сравнение – также по парному критерию Стьюдента. Все вычисления проводили с помощью пакета программ SPSS 13.0.
Результаты
Показано влияние факторов «число нажатий» и «время» на суммарные (усредненные по всем отведениям) спектральные характеристики ЭЭГ дельта-колебаний (F (1; 18) = 8,01, p < 0,011 и F (9; 10) = 3,13, p < 0,05 соответственно) и тета-ритма (F (1; 18) = 8,9, p < 0,01 и F (9; 10) = 3,89, p < 0,03 соответственно), а также факторов «поведенческая активность» и «время» на альфа1-ритм (F (1; 18) = 5,63, p < 0,03 и F (9; 10) = 9,35, p < 0,001 соответственно).
Мощность дельта- и тета-активности в целом была больше во время пробуждения при единичном нажатии, чем при серии нажатий. Низкочастотный альфа-ритм был больше после начала нажатий, чем в период до них. Значимые различия между ситуациями с единичным нажатием и серией нажатий по временным отрезкам были показаны для дельта-колебаний и альфа1-ритма за 13–14 секунд до начала нажатий и для дельта-колебаний в первые четыре секунды психомоторной деятельности (рис. 1).
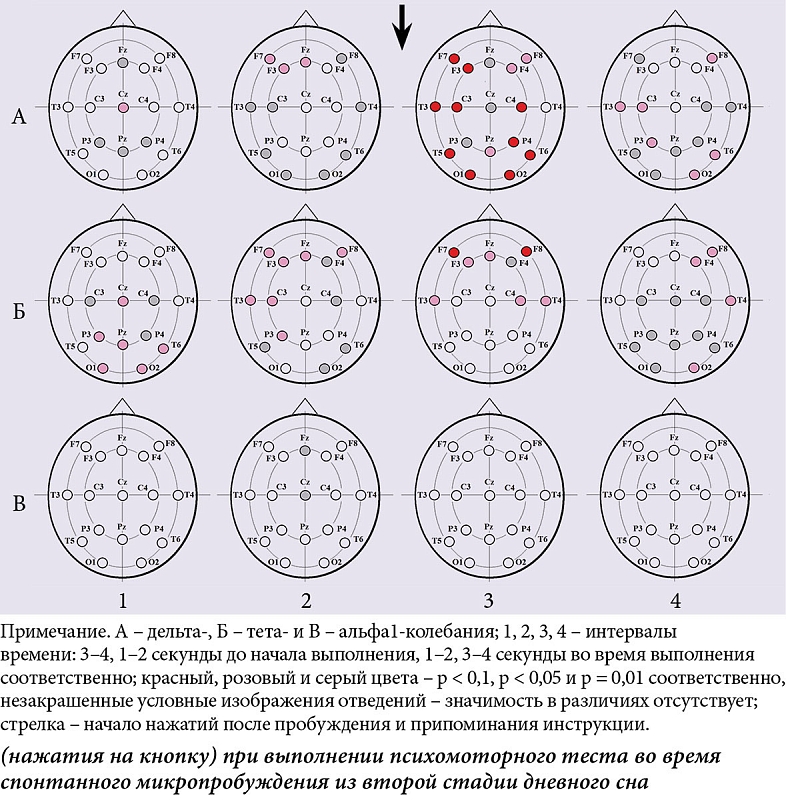
Показано генерализованное влияние по отведениям ЭЭГ фактора «число нажатий» на мощность дельта-колебаний и тета-ритма (рис. 2А.1 и рис. 2Б.1) и фактора «поведенческая активность» на мощность альфа1-ритма в каудальных областях коры (рис. 2В.2).
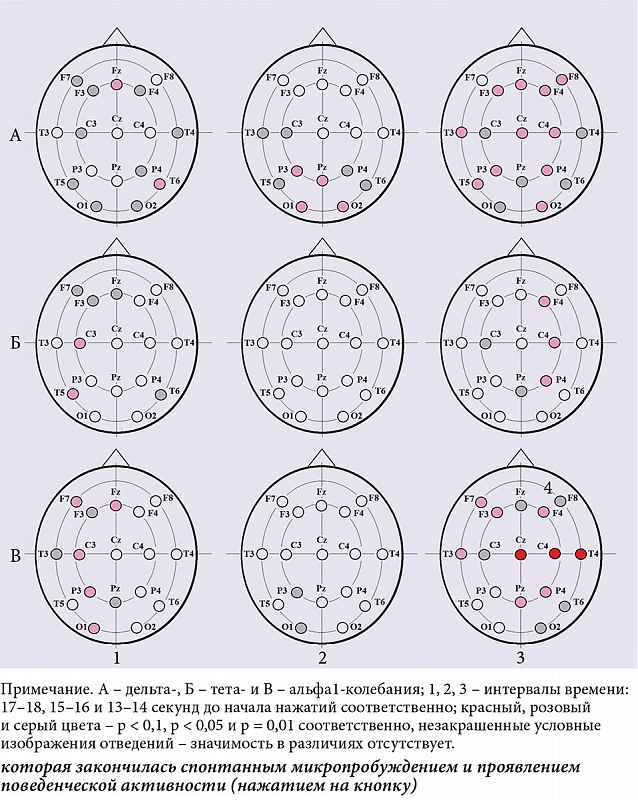
В интервале 3–4 секунды до начала деятельности между ситуацией с единичным нажатием и ситуацией с серией нажатий показаны значимые различия в мощности тета-ритма в каудальных областях коры (рис. 3Б.1). В интервале 1–2 секунды, непосредственно перед деятельностью, – различия в мощности дельта-колебаний в левой лобной и лобно-височной коре (рис. 3А.2), в мощности тета-ритма в лобной, лобно-височной, передневисочной, центральной и теменной областях левого и лобно-височной области правого полушария (рис. 3Б.2). В интервале 1–2 секунды, непосредственно после начала нажатий, показаны различия в мощности дельта-колебаний практически во всех отведениях ЭЭГ (рис. 3А.3) и в мощности тета-ритма в передних и височных областях обоих полушарий (рис. 3Б.3). Во всех случаях величины спектральных характеристик были больше в ситуации с единичными нажатиями.
В начале активации, которая закончилась микропробуждением и эпизодом поведенческой активности в интервале 13–14 секунд до начала нажатий между ситуацией с единичным нажатием и ситуацией с серией нажатий, показаны значимые различия в мощности дельта-колебаний практически во всей области отведения ЭЭГ (рис. 4А.3) и в мощности альфа1-ритма в передних областях левого и генерализованно – в правом полушарии (рис. 4В.3).
Обсуждение
Периоды активации, которые заканчиваются микропробуждениями с единичным нажатием на кнопку, обусловленным необходимостью выполнения психомоторного теста, сопровождаются ЭЭГ с более выраженными дельта-колебаниями и тета-ритмом большей мощности, чем в ситуации с серией нажатий. Этот факт позволяет предположить, что более короткий период моторной активности происходит при пробуждении из более выраженной второй стадии ортодоксального сна. В этом случае более высокие амплитуды К-комплексов, которые непосредственно предшествуют единичному нажатию, можно рассматривать как проявление активационных процессов, требующихся для окончательного выведения субъекта из более глубокого сонного состояния. Роль К-комплексов в активациях мозга, которые приводят к пробуждению, показана в работах [8, 15]. Также были получены доказательства, что мощность К-комплексов должна достигать определенного уровня, чтобы запустить этот процесс и подвести субъекта к деятельности [16]. Вместе с тем более глубокое сонное состояние, предшествующее деятельности, предполагает более сильное влияние на нее такого явления, как инерция сна [17]. На более выраженное влияние инерции сна, которое, как мы предполагаем, сократило поведение до единичного нажатия на кнопку, указывают более высокие характеристики дельта- и тета-активности во время и после единичного нажатия, чем в ситуации серии нажатий. Отметим, что в других обстоятельствах, а именно на начальной стадии исследуемой нами продолжительной активации мозга (за 13–14 секунд до единичного нажатия), появление К-комплексов существенно меньшей амплитуды не приводило к пробуждению и двигательной активности.
Более выраженная мощность тета-ритма при единичных нажатиях, как мы уже упоминали, связывается с процессами инерции сна. Вместе с тем тета-ритм, особенно во фронтальной коре, связывают с процессами когнитивного контроля [18]. При этом тета-ритм увеличивается не только во время выполнения заданий психомоторного теста, но и в процессе подготовки к нему. Иначе говоря, функция контроля включается раньше начала деятельности и носит упреждающий характер. Наблюдаемые нами бóльшие величины мощности тета-ритма в ситуации единичного нажатия по сравнению с серией нажатий во фронтальных областях отведения ЭЭГ можно рассматривать как усиление функции когнитивного контроля, который необходим при психомоторной деятельности в условиях преодоления более выраженного влияния ортодоксального сна.
Усиление альфа-ритма в период микропробуждения непосредственно перед началом нажатий мы связываем в наших исследованиях с припоминанием инструкции и подготовкой к воспроизведению психомоторного теста [13, 16]. Увеличение его мощностных характеристик в период после начала нажатий, особенно в теменно-затылочных областях, отражает поддержание активации при выполнении этой психомоторной деятельности, а далее, когда нажатия прекращаются, вероятно, ее следовые процессы. Теменно-затылочный альфа-ритм необходим для поддержания сознания [19]. Необходимость присутствия этой ритмической составляющей в ЭЭГ для осуществления сознательной деятельности при коротких пробуждениях показана в работе [11]. Ранее нами отмечались существенная роль этого ритма при выполнении психомоторного теста и его связь с уровнем сознания во время более длительных пробуждений [13]. На этом основании можно предположить, что не только паттерн из 2–6 нажатий, но и единичные нажатия являются сознательными актами, однако уровень сознания, вероятно, значительно снижен по сравнению с выполнением данной деятельности в состоянии бодрствования. При этом отсутствие различий в мощностных характеристиках альфа-ритма, сопровождающих обсуждаемую кратковременную деятельность разной длительности, свидетельствует, что в обоих случаях воспроизведение психомоторного теста в условиях микропробуждения происходит на сходном уровне.
Заключение
Длительность выполнения психомоторного теста в условиях кратковременного пробуждения из второй стадии ортодоксального дневного сна связана с глубиной этой стадии, которая характеризуется разной величиной мощностных характеристик дельта-колебаний и тета-ритма. По сравнению с несколькими нажатиями единичным нажатиям предшествует период более глубокого сонного состояния, который завершается К-комплексами с более высокими амплитудами. К-комплексы в этих обстоятельствах рассматриваются как проявление активационных процессов, требующихся для подготовки к выполнению нажатий на кнопку. Отсутствие различий в мощностных характеристиках альфа-ритма при деятельности разной длительности свидетельствует, что в обоих случаях воспроизведение психомоторного теста в условиях микропробуждения происходит на сходном функциональном и сознательном уровне. Полученные результаты будут полезны в клинической практике при исследовании расстройств возбуждений во время сна, которые приводят к непроизвольной двигательной активности различной длительности и сложности.
Работа выполнена в рамках государственного задания Министерства образования и науки Российской Федерации на 2021–2023 гг.
Конфликт интересов отсутствует.
E.A. Cheremushkin, PhD, N.E. Petrenko, V.B. Dorokhov
Institute of Higher Nervous Activity and Neurophysiology RAS
Contact person: Evgeny Al. Cheremushkin, khton@mail.ru
The recovery of different durations cognitive activity under the conditions of performing a psychomotor test during micro-awakening from stage 2 orthodox daytime sleep was investigated in 19 healthy young adults, students. We analyzed electroencephalogram (EEG) during the period of brain arousal that ended with an episode of awakening with different numbers of button presses, as well as the period of time that followed behavioral activity. The number of clicks was related to the depth of this sleep stage, which was characterized by different magnitudes of the power characteristics of delta oscillations and theta rhythm. Compared to the activity pattern (2–6 taps), the single-tap situation is preceded by a period of deeper sleep state, which is completed by K-complexes with higher amplitudes. The K-complexes in these circumstances are considered to be a manifestation of the activation processes required to prepare for the execution of button presses. The higher power values of the theta rhythm in the situation of a single press compared to the pattern of presses in the frontal regions of the EEG can be considered as a strengthening of the function of cognitive control, which is necessary for psychomotor activity in conditions of overcoming the more pronounced influence of orthodox sleep. At the same time, the theta rhythm increases not only during the performance of psychomotor test tasks, but also in the process of preparation for it. In other words, the control function is switched on before the beginning of activity and has an anticipatory character. The absence of differences in the power characteristics of the alpha rhythm during the period of instruction recall and during activities of different duration indicates that in both cases the reproduction of the psychomotor test under conditions of micro-awakening occurs at a similar functional and conscious level. The results obtained will be useful in clinical practice in the study of disorders of arousal during sleep, which are accompanied by involuntary motor activity of varying duration and complexity.
Уважаемый посетитель uMEDp!
Уведомляем Вас о том, что здесь содержится информация, предназначенная исключительно для специалистов здравоохранения.
Если Вы не являетесь специалистом здравоохранения, администрация не несет ответственности за возможные отрицательные последствия, возникшие в результате самостоятельного использования Вами информации с портала без предварительной консультации с врачом.
Нажимая на кнопку «Войти», Вы подтверждаете, что являетесь врачом или студентом медицинского вуза.