–Э–µ–є—А–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Є–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л —А–µ–≥—Г–ї—П—Ж–Є–Є —Б–љ–∞ –Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П: —А–Њ–ї—М –±–ї–Њ–Ї–∞—В–Њ—А–Њ–≤ –≥–Є—Б—В–∞–Љ–Є–љ–Њ–≤—Л—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –≤ –ї–µ—З–µ–љ–Є–Є –Є–љ—Б–Њ–Љ–љ–Є–Є
- –Р–љ–љ–Њ—В–∞—Ж–Є—П
- –°—В–∞—В—М—П
- –°—Б—Л–ї–Ї–Є
- English
–Ю—В–Ї—А—Л—В–Є–µ –Э. Mougoun –Є¬†D.¬†Morruzzi –≤¬†–Ї–Њ–љ—Ж–µ 1940-—Е –≥–≥. —А–µ—В–Є–Ї—Г–ї—П—А–љ–Њ–є —Д–Њ—А–Љ–∞—Ж–Є–Є —Б—В–≤–Њ–ї–∞ –≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ –њ–Њ–і—В–≤–µ—А–і–Є–ї–Њ, —З—В–Њ –љ–Њ—А–Љ–∞–ї—М–љ–Њ–µ —Д—Г–љ–Ї—Ж–Є–Њ–љ–Є—А–Њ–≤–∞–љ–Є–µ —В–∞–ї–∞–Љ–Њ-–Ї–Њ—А—В–Є–Ї–∞–ї—М–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –Љ–Њ–Ј–≥–∞, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—Й–µ–µ –≤–µ—Б—М —Б–њ–µ–Ї—В—А —Б–Њ–Ј–љ–∞—В–µ–ї—М–љ–Њ–є –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є —З–µ–ї–Њ–≤–µ–Ї–∞, –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ —В–Њ–ї—М–Ї–Њ –њ—А–Є –љ–∞–ї–Є—З–Є–Є –Љ–Њ—Й–љ—Л—Е —В–Њ–љ–Є—З–µ—Б–Ї–Є—Е –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–є —Б–Њ —Б—В–Њ—А–Њ–љ—Л –Њ–њ—А–µ–і–µ–ї–µ–љ–љ—Л—Е –њ–Њ–і–Ї–Њ—А–Ї–Њ–≤—Л—Е —Б—В—А—Г–Ї—В—Г—А, –љ–∞–Ј—Л–≤–∞–µ–Љ—Л—Е –∞–Ї—В–Є–≤–Є—А—Г—О—Й–Є–Љ–Є. –Я—А—П–Љ–Њ–µ –Є–Ј—Г—З–µ–љ–Є–µ –љ–µ–є—А–Њ–љ–Њ–≤, –≤–Њ–≤–ї–µ—З–µ–љ–љ—Л—Е –≤¬†—А–µ–≥—Г–ї—П—Ж–Є—О —Ж–Є–Ї–ї–∞ ¬Ђ—Б–Њ–љ¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬ї, –њ—А–Њ–≤–µ–і–µ–љ–љ–Њ–µ –≤–Њ –≤—В–Њ—А–Њ–є –њ–Њ–ї–Њ–≤–Є–љ–µ –Љ–Є–љ—Г–≤—И–µ–≥–Њ —Б—В–Њ–ї–µ—В–Є—П, –њ–Њ–Ї–∞–Ј–∞–ї–Њ, —З—В–Њ –±–ї–∞–≥–Њ–і–∞—А—П —Н—В–Є–Љ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П–Љ –Љ–µ–Љ–±—А–∞–љ–∞ –±–Њ–ї—М—И–Є–љ—Б—В–≤–∞ –Ї–Њ—А—В–Є–Ї–∞–ї—М–љ—Л—Е –љ–µ–є—А–Њ–љ–Њ–≤ –≤¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–Є –і–µ–њ–Њ–ї—П—А–Є–Ј–Њ–≤–∞–љ–∞ –љ–∞ 5вАУ10¬†–Љ–Т –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–њ–Њ—В–µ–љ—Ж–Є–∞–ї–Њ–Љ –њ–Њ–Ї–Њ—П (-68/-70¬†–Љ–Т). –Ґ–Њ–ї—М–Ї–Њ –≤¬†—В–∞–Ї–Њ–Љ —Б–Њ—Б—В–Њ—П–љ–Є–Є —В–Њ–љ–Є—З–µ—Б–Ї–Њ–є –і–µ–њ–Њ–ї—П—А–Є–Ј–∞—Ж–Є–Є —Н—В–Є –љ–µ–є—А–Њ–љ—Л —Б–њ–Њ—Б–Њ–±–љ—Л –Њ–±—А–∞–±–∞—В—Л–≤–∞—В—М –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—О –Є¬†–Њ—В–≤–µ—З–∞—В—М –љ–∞ —Б–Є–≥–љ–∞–ї—Л, –њ—А–Є—Е–Њ–і—П—Й–Є–µ –Ї¬†–љ–Є–Љ –Њ—В –і—А—Г–≥–Є—Е –љ–µ—А–≤–љ—Л—Е –Ї–ї–µ—В–Њ–Ї, –Ї–∞–Ї —А–µ—Ж–µ–њ—В–Њ—А–љ—Л—Е, —В–∞–Ї –Є¬†–≤–љ—Г—В—А–Є–Љ–Њ–Ј–≥–Њ–≤—Л—Е. –Ґ–∞–Ї–Є—Е —Б–Є—Б—В–µ–Љ —В–Њ–љ–Є—З–µ—Б–Ї–Њ–є –і–µ–њ–Њ–ї—П—А–Є–Ј–∞—Ж–Є–Є, –Є–ї–Є –≤–Њ—Б—Е–Њ–і—П—Й–µ–є –∞–Ї—В–Є–≤–∞—Ж–Є–Є –Љ–Њ–Ј–≥–∞ (–Є—Е –Љ–Њ–ґ–љ–Њ —Г—Б–ї–Њ–≤–љ–Њ –љ–∞–Ј–≤–∞—В—М ¬Ђ—Ж–µ–љ—В—А–∞–Љ–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П¬ї), –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ¬†вАУ –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –љ–µ –Љ–µ–љ–µ–µ –і–µ—Б—П—В–Є; –Њ–љ–Є —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ—Л –љ–∞ –≤—Б–µ—Е —Г—А–Њ–≤–љ—П—Е –Љ–Њ–Ј–≥–Њ–≤–Њ–є –Њ—Б–Є –Є¬†–≤—Л–і–µ–ї—П—О—В —А–∞–Ј–ї–Є—З–љ—Л–µ —Е–Є–Љ–Є—З–µ—Б–Ї–Є–µ –Љ–µ–і–Є–∞—В–Њ—А—Л. –Ф–µ—В–∞–ї–Є —Б—В—А–Њ–µ–љ–Є—П –Є¬†—Д—Г–љ–Ї—Ж–Є–Њ–љ–Є—А–Њ–≤–∞–љ–Є—П —Н—В–Є—Е —Б–Є—Б—В–µ–Љ –њ–Њ–і—А–Њ–±–љ–Њ –Є–Ј–ї–Њ–ґ–µ–љ—Л –≤¬†—А—П–і–µ –љ–µ–і–∞–≤–љ–Є—Е —А—Г—Б—Б–Ї–Њ—П–Ј—Л—З–љ—Л—Е –Њ–±–Ј–Њ—А–Њ–≤ [1вАУ5].
–Ю—Б–Њ–±—Г—О —А–Њ–ї—М –≤¬†–њ–Њ–і–і–µ—А–ґ–∞–љ–Є–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є–≥—А–∞–µ—В –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–∞—П —Б–Є—Б—В–µ–Љ–∞, —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–љ–∞—П –≤¬†—В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–Љ (–±—Г–≥–Њ—А–Ї–Њ–≤–Њ-—Б–Њ—Б—Ж–µ–≤–Є–і–љ–Њ–Љ) —П–і—А–µ –Ј–∞–і–љ–µ–≥–Њ –≥–Є–њ–Њ—В–∞–ї–∞–Љ—Г—Б–∞ [6вАУ9]. –Я–Њ –Љ–µ—В–Њ–і–Є—З–µ—Б–Ї–Є–Љ –њ—А–Є—З–Є–љ–∞–Љ —В–Њ—З–љ–∞—П –ї–Њ–Ї–∞–ї–Є–Ј–∞—Ж–Є—П —Н—В–Њ–є —Б–Є—Б—В–µ–Љ—Л –Є¬†–µ–µ –њ—А–Њ–µ–Ї—Ж–Є–є –≤¬†–≥–Њ–ї–Њ–≤–љ–Њ–Љ –Љ–Њ–Ј–≥–µ –Ї—А—Л—Б –±—Л–ї–∞ –Њ–њ–Є—Б–∞–љ–∞ –њ–Њ–Ј–ґ–µ, —З–µ–Љ —Б–Є—Б—В–µ–Љ –і—А—Г–≥–Є—Е –Љ–Њ–Ј–≥–Њ–≤—Л—Е –∞–Љ–Є–љ–Њ–≤,¬†вАУ –ї–Є—И—М –≤¬†1983вАУ1984¬†–≥–≥.¬†вАУ —П–њ–Њ–љ—Б–Ї–Є–Љ–Є –Є¬†–∞–Љ–µ—А–Є–Ї–∞–љ—Б–Ї–Є–Љ–Є –∞–≤—В–Њ—А–∞–Љ–Є. –Р¬†–≤¬†1988¬†–≥. –≤¬†–ї–∞–±–Њ—А–∞—В–Њ—А–Є–Є M. Jouvet –±—Л–ї –Њ–±–љ–∞—А—Г–ґ–µ–љ –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є–є ¬Ђarousal-–Љ–µ—Е–∞–љ–Є–Ј–Љ¬ї –≤¬†–≥–Є–њ–Њ—В–∞–ї–∞–Љ—Г—Б–µ –Ї–Њ—И–Ї–Є [10].
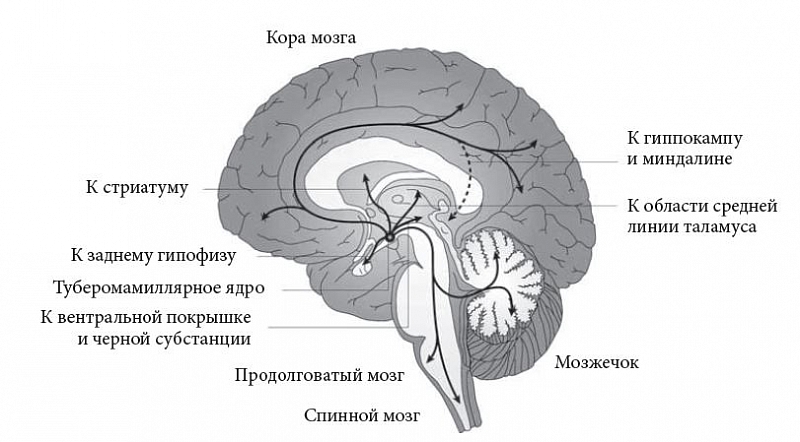
–Ґ—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–µ —П–і—А–Њ¬†вАУ —Н—В–Њ –µ–і–Є–љ—Б—В–≤–µ–љ–љ—Л–є –Є—Б—В–Њ—З–љ–Є–Ї –≥–Є—Б—В–∞–Љ–Є–љ–∞ –≤¬†–≥–Њ–ї–Њ–≤–љ–Њ–Љ –Љ–Њ–Ј–≥–µ –њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е. –Я–Њ–і–Њ–±–љ–Њ –±–Њ–ї—М—И–Є–љ—Б—В–≤—Г –і—А—Г–≥–Є—Е –∞–Ї—В–Є–≤–Є—А—Г—О—Й–Є—Е —Б–Є—Б—В–µ–Љ, –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–∞—П —Б–Є—Б—В–µ–Љ–∞ —В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–≥–Њ —П–і—А–∞ —Г—Б—В—А–Њ–µ–љ–∞ –њ–Њ ¬Ђ–і—А–µ–≤–Њ–≤–Є–і–љ–Њ–Љ—Г¬ї –њ—А–Є–љ—Ж–Є–њ—Г: –Њ—З–µ–љ—М –љ–µ–±–Њ–ї—М—И–Њ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –љ–µ–є—А–Њ–љ–Њ–≤ (–≤ –Љ–Њ–Ј–≥–µ –Ї—А—Л—Б—Л¬†вАУ –ї–Є—И—М 3вАУ4¬†—В—Л—Б., –≤¬†–Љ–Њ–Ј–≥–µ —З–µ–ї–Њ–≤–µ–Ї–∞¬†вАУ 64¬†—В—Л—Б.) –Є–љ–љ–µ—А–≤–Є—А—Г–µ—В –Љ–Є–ї–ї–Є–∞—А–і—Л –Ї–ї–µ—В–Њ–Ї –љ–Њ–≤–Њ–є, –і—А–µ–≤–љ–µ–є –Ї–Њ—А—Л –Є¬†–њ–Њ–і–Ї–Њ—А–Ї–Њ–≤—Л—Е —Б—В—А—Г–Ї—В—Г—А –Ј–∞ —Б—З–µ—В –Ї–Њ–ї–Њ—Б—Б–∞–ї—М–љ–Њ–≥–Њ –≤–µ—В–≤–ї–µ–љ–Є—П —Б–≤–Њ–Є—Е –∞–Ї—Б–Њ–љ–Њ–≤ (–Ї–∞–ґ–і—Л–є –∞–Ї—Б–Њ–љ –Њ–±—А–∞–Ј—Г–µ—В —Б–Њ—В–љ–Є —В—Л—Б—П—З –Њ—В–≤–µ—В–≤–ї–µ–љ–Є–є). –Э–∞–Є–±–Њ–ї–µ–µ –Љ–Њ—Й–љ—Л–µ –≤–Њ—Б—Е–Њ–і—П—Й–Є–µ –њ—А–Њ–µ–Ї—Ж–Є–Є –љ–∞–њ—А–∞–≤–ї—П—О—В—Б—П –≤¬†–љ–µ–є—А–Њ–≥–Є–њ–Њ—Д–Є–Ј, –±–ї–Є–Ј–ї–µ–ґ–∞—Й–Є–µ –і–Њ—Д–∞–Љ–Є–љ—Б–Њ–і–µ—А–ґ–∞—Й–Є–µ –Њ–±–ї–∞—Б—В–Є –≤–µ–љ—В—А–∞–ї—М–љ–Њ–є –њ–Њ–Ї—А—Л—И–Ї–Є —Б—А–µ–і–љ–µ–≥–Њ –Љ–Њ–Ј–≥–∞ –Є¬†–Ї–Њ–Љ–њ–∞–Ї—В–љ–Њ–є —З–∞—Б—В–Є —З–µ—А–љ–Њ–є —Б—Г–±—Б—В–∞–љ—Ж–Є–Є, –±–∞–Ј–∞–ї—М–љ—Г—О –Њ–±–ї–∞—Б—В—М –њ–µ—А–µ–і–љ–µ–≥–Њ –Љ–Њ–Ј–≥–∞ (–Ї—А—Г–њ–љ–Њ–Ї–ї–µ—В–Њ—З–љ—Л–µ —П–і—А–∞ –±–µ–Ј—Л–Љ—П–љ–љ–Њ–є —Б—Г–±—Б—В–∞–љ—Ж–Є–Є, —Б–Њ–і–µ—А–ґ–∞—Й–Є–µ –∞—Ж–µ—В–Є–ї—Е–Њ–ї–Є–љ –Є¬†–≥–∞–Љ–Љ–∞-–∞–Љ–Є–љ–Њ–Љ–∞—Б–ї—П–љ—Г—О –Ї–Є—Б–ї–Њ—В—Г (–У–Р–Ь–Ъ)), —Б—В—А–Є–∞—В—Г–Љ, –љ–µ–Њ–Ї–Њ—А—В–µ–Ї—Б, –≥–Є–њ–њ–Њ–Ї–∞–Љ–њ, –Љ–Є–љ–і–∞–ї–Є–љ—Г –Є¬†—В–∞–ї–∞–Љ–Є—З–µ—Б–Ї–Є–µ —П–і—А–∞ —Б—А–µ–і–љ–µ–є –ї–Є–љ–Є–Є, –∞¬†–љ–Є—Б—Е–Њ–і—П—Й–Є–µ¬†вАУ –≤¬†–Љ–Њ–Ј–ґ–µ—З–Њ–Ї, –њ—А–Њ–і–Њ–ї–≥–Њ–≤–∞—В—Л–є –Є¬†—Б–њ–Є–љ–љ–Њ–є –Љ–Њ–Ј–≥.
–Ґ—Й–∞—В–µ–ї—М–љ—Л–µ –љ–µ–є—А–Њ–Љ–Њ—А—Д–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П, –њ—А–Њ–≤–µ–і–µ–љ–љ—Л–µ –≤¬†–ї–∞–±–Њ—А–∞—В–Њ—А–Є–Є M. Jouvet –Є¬†–≤¬†–і—А—Г–≥–Є—Е –ї–∞–±–Њ—А–∞—В–Њ—А–Є—П—Е –љ–∞ –Љ–Њ–Ј–≥–µ –Ї–Њ—И–Ї–Є –Є¬†–Ї—А—Л—Б—Л, –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є–µ –љ–µ–є—А–Њ–љ—Л —В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–≥–Њ —П–і—А–∞ –њ—А–Њ–µ—Ж–Є—А—Г—О—В—Б—П —В–∞–Ї–ґ–µ –љ–∞ –ї–∞—В–µ—А–Њ–і–Њ—А–Ј–∞–ї—М–љ–Њ–µ –Є¬†–њ–µ–і—Г–љ–Ї—Г–ї–Њ–њ–Њ–љ—В–Є–љ–љ–Њ–µ —П–і—А–∞ –Љ–µ–Ј–Њ–њ–Њ–љ—В–Є–љ–љ–Њ–є –њ–Њ–Ї—А—Л—И–Ї–Є, –≤—Л–і–µ–ї—П—О—Й–Є–µ –∞—Ж–µ—В–Є–ї—Е–Њ–ї–Є–љ, —Б–Є–љ–µ–µ (–≥–Њ–ї—Г–±–Њ–µ) –њ—П—В–љ–Њ, –њ—А–Њ–і—Г—Ж–Є—А—Г—О—Й–µ–µ –љ–Њ—А–∞–і—А–µ–љ–∞–ї–Є–љ, –љ–∞ –і–Њ—А–Ј–∞–ї—М–љ—Л–µ —П–і—А–∞ —И–≤–∞, —Б–Є–љ—В–µ–Ј–Є—А—Г—О—Й–Є–µ —Б–µ—А–Њ—В–Њ–љ–Є–љ (—А–Є—Б.¬†1) [11]. –Т¬†—Б–≤–Њ—О –Њ—З–µ—А–µ–і—М –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є–µ –љ–µ–є—А–Њ–љ—Л —В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–≥–Њ —П–і—А–∞ –њ–Њ–ї—Г—З–∞—О—В –∞—Д—Д–µ—А–µ–љ—В—Л –Њ—В –Є–љ—Д—А–∞–ї–Є–Љ–±–Є—З–µ—Б–Ї–Њ–є –Ї–Њ—А—Л, –ї–∞—В–µ—А–∞–ї—М–љ–Њ–є –Њ–±–ї–∞—Б—В–Є –њ–µ—А–µ–≥–Њ—А–Њ–і–Ї–Є, –њ—А–µ–Њ–њ—В–Є—З–µ—Б–Ї–Њ–є –Њ–±–ї–∞—Б—В–Є –њ–µ—А–µ–і–љ–µ–≥–Њ –≥–Є–њ–Њ—В–∞–ї–∞–Љ—Г—Б–∞, –∞–і—А–µ–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е –Ї–ї–µ—В–Њ–Ї –°1вАУ–°3, –љ–Њ—А–∞–і—А–µ–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е –љ–µ–є—А–Њ–љ–Њ–≤ –Р1вАУ–Р3 –Є¬†—Б–µ—А–Њ—В–Њ–љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е –Ї–ї–µ—В–Њ–Ї –Т5вАУ–Т9. –Ш–љ—В–µ—А–µ—Б–љ–Њ, —З—В–Њ –ї–Є—И—М –Њ–і–Є–љ–Њ—З–љ—Л–µ –≤–Њ–ї–Њ–Ї–љ–∞ –і–Њ—Б—В–Є–≥–∞—О—В —В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–≥–Њ —П–і—А–∞ –Њ—В –љ–Њ—А–∞–і—А–µ–љ–∞–ї–Є–љ—Б–Њ–і–µ—А–ґ–∞—Й–Є—Е –Ї–ї–µ—В–Њ–Ї —Б–Є–љ–µ–≥–Њ –њ—П—В–љ–∞ –Є¬†–і–Њ—Д–∞–Љ–Є–љ—Б–Њ–і–µ—А–ґ–∞—Й–Є—Е –љ–µ–є—А–Њ–љ–Њ–≤ –≤–µ–љ—В—А–∞–ї—М–љ–Њ–є –њ–Њ–Ї—А—Л—И–Ї–Є —Б—А–µ–і–љ–µ–≥–Њ –Љ–Њ–Ј–≥–∞ –Є¬†–Ї–Њ–Љ–њ–∞–Ї—В–љ–Њ–є —З–∞—Б—В–Є —З–µ—А–љ–Њ–є —Б—Г–±—Б—В–∞–љ—Ж–Є–Є [7]. –Ю–і–љ–∞–Ї–Њ –њ—А–Є –±–Њ–ї–µ–Ј–љ–Є –Я–∞—А–Ї–Є–љ—Б–Њ–љ–∞, —Б–≤—П–Ј–∞–љ–љ–Њ–є —Б¬†—А–∞–Ј—А—Г—И–µ–љ–Є–µ–Љ –і–Њ—Д–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Њ–є –њ–µ—А–µ–і–∞—З–Є, –љ–∞–±–ї—О–і–∞–µ—В—Б—П –і–≤—Г–Ї—А–∞—В–љ–Њ–µ –њ–Њ–≤—Л—И–µ–љ–Є–µ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –≥–Є—Б—В–∞–Љ–Є–љ–∞, –њ–Њ—Б—В—Г–њ–∞—О—Й–µ–≥–Њ –Є–Ј —В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–≥–Њ —П–і—А–∞ –≤¬†–Ї–Њ–Љ–њ–∞–Ї—В–љ—Г—О —З–∞—Б—В—М —З–µ—А–љ–Њ–є —Б—Г–±—Б—В–∞–љ—Ж–Є–Є –Є¬†–µ–µ –њ—А–Њ–µ–Ї—Ж–Є—О¬†вАУ –±–ї–µ–і–љ—Л–є —И–∞—А.
–Ш—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –≤–∞–ґ–љ—Л –≤–Ј–∞–Є–Љ–Њ—Б–≤—П–Ј–Є –Љ–µ–ґ–і—Г –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Њ–є –Є¬†–Њ—А–µ–Ї—Б–Є–љ/–≥–Є–њ–Њ–Ї—А–µ—В–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Њ–є —Б–Є—Б—В–µ–Љ–∞–Љ–Є –Љ–Њ–Ј–≥–∞. –Ь–µ–і–Є–∞—В–Њ—А—Л —Н—В–Є—Е –і–≤—Г—Е —Б–Є—Б—В–µ–Љ –і–µ–є—Б—В–≤—Г—О—В —Б–Є–љ–µ—А–≥–Є—З–љ–Њ, –Є–≥—А–∞—П —Г–љ–Є–Ї–∞–ї—М–љ—Г—О —А–Њ–ї—М –≤¬†–њ–Њ–і–і–µ—А–ґ–∞–љ–Є–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П. –Ю—А–µ–Ї—Б–Є–љ/–≥–Є–њ–Њ–Ї—А–µ—В–Є–љ—Б–Њ–і–µ—А–ґ–∞—Й–Є–µ –љ–µ–є—А–Њ–љ—Л —А–∞—Б–њ–Њ–ї–∞–≥–∞—О—В—Б—П –≤¬†–Ј–∞–і–љ–µ–ї–∞—В–µ—А–∞–ї—М–љ–Њ–Љ –≥–Є–њ–Њ—В–∞–ї–∞–Љ—Г—Б–µ –Є¬†–њ–µ—А–Є—Д–Њ—А–љ–Є–Ї–∞–ї—М–љ–Њ–є –Њ–±–ї–∞—Б—В–Є, –≤¬†–љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ–є –±–ї–Є–Ј–Њ—Б—В–Є –Њ—В –≥–Є—Б—В–∞–Љ–Є–љ–Њ–≤—Л—Е –љ–µ–є—А–Њ–љ–Њ–≤ —В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–≥–Њ —П–і—А–∞. –Ю–±–∞ —П–і—А–∞ —З–∞—Б—В–Є—З–љ–Њ –њ–µ—А–µ–Ї—А—Л–≤–∞—О—В—Б—П –Є¬†–Њ–±—А–∞–Ј—Г—О—В —Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–µ –µ–і–Є–љ—Б—В–≤–Њ. –Ю–±–∞ –њ–µ–њ—В–Є–і–∞ –Њ—А–µ–Ї—Б–Є–љ–∞ (–≥–Є–њ–Њ–Ї—А–µ—В–Є–љ–∞) –≤–Њ–Ј–±—Г–ґ–і–∞—О—В –≥–Є—Б—В–∞–Љ–Є–љ–Њ–≤—Л–µ –љ–µ–є—А–Њ–љ—Л —З–µ—А–µ–Ј —Б–≤–Њ–Є —А–µ—Ж–µ–њ—В–Њ—А—Л 2-–≥–Њ —В–Є–њ–∞ –Є¬†–∞–Ї—В–Є–≤–∞—Ж–Є—О –љ–∞—В—А–Є–є-–Ї–∞–ї—М—Ж–Є–µ–≤–Њ–≥–Њ –Є–Њ–љ–љ–Њ–≥–Њ –Њ–±–Љ–µ–љ–∞. –Ю—А–µ–Ї—Б–Є–љ —З–∞—Б—В–Њ —Б–Њ–ї–Њ–Ї–∞–ї–Є–Ј—Г–µ—В—Б—П —Б¬†–і–Є–љ–Њ—А—Д–Є–љ–Њ–Љ, –Ї–Њ—В–Њ—А—Л–є —В–∞–Ї–ґ–µ –Љ–Њ–ґ–µ—В —Г—З–∞—Б—В–≤–Њ–≤–∞—В—М –≤¬†–≤–Њ–Ј–±—Г–ґ–і–µ–љ–Є–Є –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е –љ–µ–є—А–Њ–љ–Њ–≤ –њ—Г—В–µ–Љ –њ–Њ–і–∞–≤–ї–µ–љ–Є—П –У–Р–Ь–Ъ–µ—А–≥–Є—З–µ—Б–Ї–Њ–≥–Њ —В–Њ—А–Љ–Њ–Ј–љ–Њ–≥–Њ –њ—Г—В–Є. –Ю–і–љ–∞–Ї–Њ –≥–Є—Б—В–∞–Љ–Є–љ–Њ–≤—Л–µ –љ–µ–є—А–Њ–љ—Л, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –љ–µ –≤–ї–Є—П—О—В –љ–∞ –≤–Њ–Ј–±—Г–і–Є–Љ–Њ—Б—В—М –Њ—А–µ–Ї—Б–Є–љ–Њ–≤—Л—Е –љ–µ–є—А–Њ–љ–Њ–≤, —В–∞–Ї —З—В–Њ –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є–µ —Н—В–Є—Е –і–≤—Г—Е —Б–Є—Б—В–µ–Љ –љ–Њ—Б–Є—В –Њ–і–љ–Њ—Б—В–Њ—А–Њ–љ–љ–Є–є —Е–∞—А–∞–Ї—В–µ—А [12].
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –Љ–Њ–ґ–љ–Њ —Б–Ї–∞–Ј–∞—В—М, —З—В–Њ –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–∞—П –Є¬†–і—А—Г–≥–Є–µ –∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є–µ —Б–Є—Б—В–µ–Љ—Л –Љ–µ–ґ—Г—В–Њ—З–љ–Њ–≥–Њ, —Б—А–µ–і–љ–µ–≥–Њ –Љ–Њ–Ј–≥–∞ –Є¬†—Б—В–≤–Њ–ї–∞ –Њ–±–ї–∞–і–∞—О—В –≤–µ—Б—М–Љ–∞ –Ј–љ–∞—З–Є—В–µ–ї—М–љ—Л–Љ —Б—Е–Њ–і—Б—В–≤–Њ–Љ –≤¬†—Б–≤–Њ–µ–є –Љ–Њ—А—Д–Њ–ї–Њ–≥–Є–Є, –Ї–ї–µ—В–Њ—З–љ–Њ–є –Є¬†—Б–Є—Б—В–µ–Љ–љ–Њ–є —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є–Є. –Ю–±–ї–∞–і–∞—П –Љ–љ–Њ–ґ–µ—Б—В–≤–µ–љ–љ—Л–Љ–Є –≤–Ј–∞–Є–Љ–љ—Л–Љ–Є —Б–≤—П–Ј—П–Љ–Є, –Њ–љ–Є —Д–Њ—А–Љ–Є—А—Г—О—В —Б–∞–Љ–Њ–Њ—А–≥–∞–љ–Є–Ј—Г—О—Й—Г—О—Б—П —Б–µ—В—М, —Б–≤–Њ–µ–≥–Њ —А–Њ–і–∞ ¬Ђ–Њ—А–Ї–µ—Б—В—А¬ї, –≤¬†–Ї–Њ—В–Њ—А–Њ–Љ –Њ—А–µ–Ї—Б–Є–љ–Њ–≤—Л–µ (–≥–Є–њ–Њ–Ї—А–µ—В–Є–љ–Њ–≤—Л–µ) –љ–µ–є—А–Њ–љ—Л –Є–≥—А–∞—О—В —А–Њ–ї—М –і–Є—А–Є–ґ–µ—А–∞, –∞¬†–≥–Є—Б—В–∞–Љ–Є–љ–Њ–≤—Л–µ¬†вАУ –њ–µ—А–≤–Њ–є —Б–Ї—А–Є–њ–Ї–Є [9].
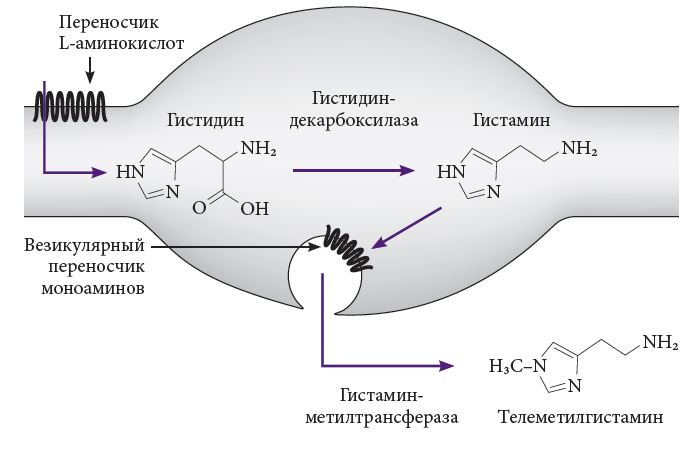
–Ъ–∞–Ї –Є–Ј–≤–µ—Б—В–љ–Њ, –≥–Є—Б—В–∞–Љ–Є–љ –Њ–±—А–∞–Ј—Г–µ—В—Б—П –Є–Ј –∞–Љ–Є–љ–Њ–Ї–Є—Б–ї–Њ—В—Л –≥–Є—Б—В–Є–і–Є–љ–∞, –њ–Њ—Б—В—Г–њ–∞—О—Й–µ–є –≤¬†–Њ—А–≥–∞–љ–Є–Ј–Љ —Б¬†–±–µ–ї–Ї–Њ–≤–Њ–є –њ–Є—Й–µ–є. –Т¬†–Њ—В–ї–Є—З–Є–µ –Њ—В –≥–Є—Б—В–∞–Љ–Є–љ–∞, –≥–Є—Б—В–Є–і–Є–љ –њ—А–Њ—Е–Њ–і–Є—В –≥–µ–Љ–∞—В–Њ—Н–љ—Ж–µ—Д–∞–ї–Є—З–µ—Б–Ї–Є–є –±–∞—А—М–µ—А –Є¬†–Ј–∞—Е–≤–∞—В—Л–≤–∞–µ—В—Б—П –±–µ–ї–Ї–Њ–Љ-—В—А–∞–љ—Б–њ–Њ—А—В–µ—А–Њ–Љ –∞–Љ–Є–љ–Њ–Ї–Є—Б–ї–Њ—В, –њ–µ—А–µ–љ–Њ—Б—П—Й–Є–Љ –µ–≥–Њ –≤–љ—Г—В—А—М —В–µ–ї–∞ –љ–µ–є—А–Њ–љ–∞ –Є–ї–Є –≤–∞—А–Є–Ї–Њ–Ј–љ–Њ–≥–Њ —А–∞—Б—И–Є—А–µ–љ–Є—П –∞–Ї—Б–Њ–љ–∞ (—А–Є—Б.¬†2). –Э–µ–є—А–Њ–љ—Л —В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–≥–Њ —П–і—А–∞ —Н–Ї—Б–њ—А–µ—Б—Б–Є—А—Г—О—В —Д–µ—А–Љ–µ–љ—В –≥–Є—Б—В–Є–і–Є–љ–і–µ–Ї–∞—А–±–Њ–Ї—Б–Є–ї–∞–Ј—Г (–У–Ф–Ъ), –Њ—В—Й–µ–њ–ї—П—О—Й–Є–є –Њ—В –Љ–Њ–ї–µ–Ї—Г–ї—Л –≥–Є—Б—В–Є–і–Є–љ–∞ –Ї–∞—А–±–Њ–Ї—Б–Є–ї –Є¬†–њ—А–µ–≤—А–∞—Й–∞—О—Й–Є–є –µ–µ –≤¬†–≥–Є—Б—В–∞–Љ–Є–љ. –У–Ф–Ъ –љ–∞–Є–±–Њ–ї–µ–µ –∞–Ї—В–Є–≤–µ–љ –≤¬†—Б–Њ–Љ–µ, –љ–Њ –њ—А–Њ—П–≤–ї—П–µ—В—Б—П —В–∞–Ї–ґ–µ –Є¬†–≤ –≤–∞—А–Є–Ї–Њ–Ј–љ—Л—Е —А–∞—Б—И–Є—А–µ–љ–Є—П—Е, –Є¬†–≤ –љ–µ—А–≤–љ—Л—Е –Њ–Ї–Њ–љ—З–∞–љ–Є—П—Е. –§–∞–Ї—В–Њ—А–Њ–Љ –Њ–≥—А–∞–љ–Є—З–µ–љ–Є—П —Б–Ї–Њ—А–Њ—Б—В–Є —Б–Є–љ—В–µ–Ј–∞ –≥–Є—Б—В–∞–Љ–Є–љ–∞ —П–≤–ї—П–µ—В—Б—П —В–Ї–∞–љ–µ–≤–∞—П –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є—П –µ–≥–Њ –њ—А–µ–і—И–µ—Б—В–≤–µ–љ–љ–Є–Ї–∞, –≥–Є—Б—В–Є–і–Є–љ–∞. –У–Є—Б—В–∞–Љ–Є–љ –њ–µ—А–µ–љ–Њ—Б–Є—В—Б—П –≤¬†–≤–µ–Ј–Є–Ї—Г–ї—Л —Б¬†–њ–Њ–Љ–Њ—Й—М—О –Њ—Б–Њ–±–Њ–≥–Њ –±–µ–ї–Ї–∞, –љ–∞–Ј—Л–≤–∞–µ–Љ–Њ–≥–Њ –≤–µ–Ј–Є–Ї—Г–ї—П—А–љ—Л–Љ –Љ–Њ–љ–Њ–∞–Љ–Є–љ–љ—Л–Љ —В—А–∞–љ—Б–њ–Њ—А—В–µ—А–Њ–Љ 2-–≥–Њ —В–Є–њ–∞ (–Т–Ь–Р–Ґ-2), –Є¬†—В–∞–Љ –љ–∞–Ї–∞–њ–ї–Є–≤–∞–µ—В—Б—П. –≠—В–Є –≤–µ–Ј–Є–Ї—Г–ї—Л —А–∞—Б–њ–Њ–ї–∞–≥–∞—О—В—Б—П –љ–µ —В–Њ–ї—М–Ї–Њ –≤¬†–Ї–ї–µ—В–Њ—З–љ—Л—Е —В–µ–ї–∞—Е, –љ–Њ –Є¬†–≤ –≤–∞—А–Є–Ї–Њ–Ј–љ—Л—Е —А–∞—Б—И–Є—А–µ–љ–Є—П—Е –∞–Ї—Б–Њ–љ–Њ–≤. –Я—А–Є –≤–Њ–Ј–љ–Є–Ї–љ–Њ–≤–µ–љ–Є–Є –њ–Њ—В–µ–љ—Ж–Є–∞–ї–∞ –і–µ–є—Б—В–≤–Є—П –≥–Є—Б—В–∞–Љ–Є–љ –≤—Л–і–µ–ї—П–µ—В—Б—П Ca++-–Ј–∞–≤–Є—Б–Є–Љ—Л–Љ –њ—Г—В–µ–Љ. –Т—Л–і–µ–ї–Є–≤—И–Є–є—Б—П –≤¬†—Б–Є–љ–∞–њ—В–Є—З–µ—Б–Ї—Г—О —Й–µ–ї—М –Є–ї–Є –Љ–µ–ґ–Ї–ї–µ—В–Њ—З–љ–Њ–µ –њ—А–Њ—Б—В—А–∞–љ—Б—В–≤–Њ –≥–Є—Б—В–∞–Љ–Є–љ, –љ–µ —Б–≤—П–Ј–∞–≤—И–Є–є—Б—П —Б¬†—А–µ—Ж–µ–њ—В–Њ—А–Њ–Љ, –Є–љ–∞–Ї—В–Є–≤–Є—А—Г–µ—В—Б—П –њ—Г—В–µ–Љ –Љ–µ—В–Є–ї–Є—А–Њ–≤–∞–љ–Є—П —Б¬†–њ–Њ–Љ–Њ—Й—М—О —Д–µ—А–Љ–µ–љ—В–∞ –≥–Є—Б—В–∞–Љ–Є–љ–Љ–µ—В–Є–ї—В—А–∞–љ—Б—Д–µ—А–∞–Ј—Л (—Б–Є–љ—В–µ–Ј–Є—А—Г–µ–Љ–Њ–≥–Њ –њ–Њ—Б—В—Б–Є–љ–∞–њ—В–Є—З–µ—Б–Ї–Є –Є–ї–Є –≤¬†–≥–ї–Є–Є), –њ—А–µ–≤—А–∞—Й–∞—О—Й–µ–є –µ–≥–Њ –≤¬†—В–µ–ї–µ–Љ–µ—В–Є–ї–≥–Є—Б—В–∞–Љ–Є–љ. –Я–Њ—Б–ї–µ–і–љ–Є–є –њ–Њ–і–≤–µ—А–≥–∞–µ—В—Б—П –Њ–Ї–Є—Б–ї–Є—В–µ–ї—М–љ–Њ–Љ—Г –і–µ–∞–Љ–Є–љ–Є—А–Њ–≤–∞–љ–Є—О —Б¬†–њ–Њ–Љ–Њ—Й—М—О —Д–µ—А–Љ–µ–љ—В–∞ –Љ–Њ–љ–Њ–∞–Љ–Є–љ–Њ–Њ–Ї—Б–Є–і–∞–Ј—Л —В–Є–њ–∞ –С (–Ь–Р–Ю-–С), –њ—А–µ–≤—А–∞—Й–∞—П—Б—М –≤¬†t-–Љ–µ—В–Є–ї-–Є–Љ–Є–і–∞–Ј–Њ–ї—Г–Ї—Б—Г—Б–љ—Г—О –Ї–Є—Б–ї–Њ—В—Г. –Ь–µ—Е–∞–љ–Є–Ј–Љ–∞ –Њ–±—А–∞—В–љ–Њ–≥–Њ –Ј–∞—Е–≤–∞—В–∞ –і–ї—П –≥–Є—Б—В–∞–Љ–Є–љ–∞ –љ–µ —Б—Г—Й–µ—Б—В–≤—Г–µ—В. –Ю–±—Л—З–љ–Њ –њ–µ—А–Є–Њ–і –њ–Њ–ї—Г–ґ–Є–Ј–љ–Є –љ–µ–є—А–Њ–љ–∞–ї—М–љ–Њ–≥–Њ –≥–Є—Б—В–∞–Љ–Є–љ–∞ —Б–Њ—Б—В–∞–≤–ї—П–µ—В –Њ–Ї–Њ–ї–Њ –њ–Њ–ї—Г—З–∞—Б–∞, –љ–Њ –Њ–љ –Љ–Њ–ґ–µ—В —А–µ–Ј–Ї–Њ —Г–Ї–Њ—А–∞—З–Є–≤–∞—В—М—Б—П –њ–Њ–і –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ–Љ –≤–љ–µ—И–љ–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤, –љ–∞–њ—А–Є–Љ–µ—А —Б—В—А–µ—Б—Б–∞ [7, 13].
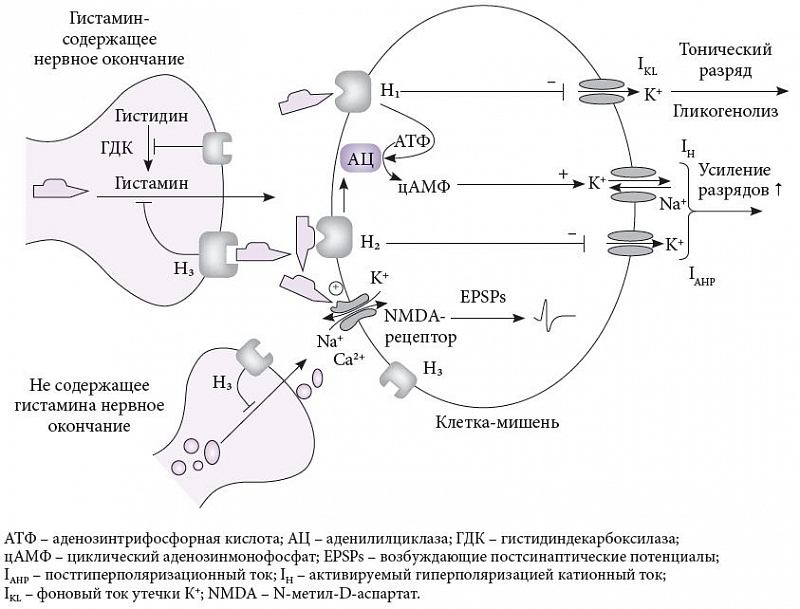
–Ш–Ј–≤–µ—Б—В–љ–Њ 4¬†—В–Є–њ–∞ –Љ–µ—В–∞–±–Њ—В—А–Њ–њ–љ—Л—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –≥–Є—Б—В–∞–Љ–Є–љ–∞, —Б–≤—П–Ј–∞–љ–љ—Л—Е —Б¬†G-–±–µ–ї–Ї–∞–Љ–Є: –і–≤–∞ –≤–Њ–Ј–±—Г–і–Є—В–µ–ї—М–љ—Л—Е (–ЭвВБ –Є¬†–ЭвВВ) –Є¬†–і–≤–∞ —В–Њ—А–Љ–Њ–Ј–љ—Л—Е (–ЭвВГ –Є¬†–ЭвВД). –†–µ—Ж–µ–њ—В–Њ—А—Л –ЭвВБвАУ–ЭвВГ –њ–Њ–≤—Б–µ–Љ–µ—Б—В–љ–Њ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л –≤¬†–≥–Њ–ї–Њ–≤–љ–Њ–Љ –Љ–Њ–Ј–≥–µ, –∞¬†—А–µ—Ж–µ–њ—В–Њ—А –ЭвВД¬†вАУ –≥–ї–∞–≤–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ –≤¬†—Б–њ–Є–љ–љ–Њ–Љ. –Р–Ї—В–Є–≤–∞—Ж–Є—П –њ–Њ—Б—В—Б–Є–љ–∞–њ—В–Є—З–µ—Б–Ї–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –ЭвВБ –Є–ї–Є –ЭвВВ, —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–љ—Л—Е –љ–∞ —Б–Њ–Љ–µ –љ–µ–є—А–Њ–љ–Њ–≤-–Љ–Є—И–µ–љ–µ–є, –Ј–∞–њ—Г—Б–Ї–∞–µ—В –≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ—Л–µ –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–µ –Ї–∞—Б–Ї–∞–і—Л, —Б–≤—П–Ј–∞–љ–љ—Л–µ —Б¬†–∞–і–µ–љ–Њ–Ј–Є–љ—В—А–Є—Д–Њ—Б—Д–∞—В–Њ–Љ, –∞–і–µ–љ–Є–ї–Є–ї—Ж–Є–Ї–ї–∞–Ј–Њ–є –Є¬†—Ж–Є–Ї–ї–Є—З–µ—Б–Ї–Є–Љ –∞–і–µ–љ–Њ–Ј–Є–љ–Љ–Њ–љ–Њ—Д–Њ—Б—Д–∞—В–Њ–Љ, –њ–Њ–≤—Л—И–∞–µ—В –Ї–ї–µ—В–Њ—З–љ—Г—О –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –Є¬†–≤–Њ–Ј–±—Г–і–Є–Љ–Њ—Б—В—М, –ї–Є–±–Њ —Б–љ–Є–ґ–∞—П —Д–Њ–љ–Њ–≤—Л–є —В–Њ–Ї —Г—В–µ—З–Ї–Є –Ъ+¬†(IKL) –Є–ї–Є –њ–Њ—Б—В–≥–Є–њ–µ—А–њ–Њ–ї—П—А–Є–Ј–∞—Ж–Є–Њ–љ–љ—Л–є —В–Њ–Ї (IAHP), –ї–Є–±–Њ –њ–Њ–≤—Л—И–∞—П –∞–Ї—В–Є–≤–Є—А—Г–µ–Љ—Л–є –≥–Є–њ–µ—А–њ–Њ–ї—П—А–Є–Ј–∞—Ж–Є–µ–є –Ї–∞—В–Є–Њ–љ–љ—Л–є —В–Њ–Ї (IH). –У–Є—Б—В–∞–Љ–Є–љ —В–∞–Ї–ґ–µ –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤—Г–µ—В —Б¬†–њ–Њ–ї–Є–∞–Љ–Є–љ–љ–Њ–є –Њ–±–ї–∞—Б—В—М—О NMDA (N-–Љ–µ—В–Є–ї-D-–∞—Б–њ–∞—А—В–∞—В–љ–Њ–≥–Њ) —А–µ—Ж–µ–њ—В–Њ—А–∞, –Љ–Њ–і—Г–ї–Є—А—Г—П –≤–Њ–Ј–±—Г–і–Є—В–µ–ї—М–љ—Л–µ –њ–Њ—Б—В—Б–Є–љ–∞–њ—В–Є—З–µ—Б–Ї–Є–µ –њ–Њ—В–µ–љ—Ж–Є–∞–ї—Л (excitatory post-synaptic potential, EPSPs) (—А–Є—Б.¬†3) [14]. –Я—А–µ—Б–Є–љ–∞–њ—В–Є—З–µ—Б–Ї–Є–µ –∞—Г—В–Њ- –Є¬†–≥–µ—В–µ—А–Њ—А–µ—Ж–µ–њ—В–Њ—А—Л —В–Є–њ–∞ –ЭвВГ –Љ–Њ–≥—Г—В —А–∞—Б–њ–Њ–ї–∞–≥–∞—В—М—Б—П –љ–∞ —Б–Њ–Љ–µ, –∞–Ї—Б–Њ–љ–∞—Е –Є¬†–і–µ–љ–і—А–Є—В–∞—Е, —В–Њ—А–Љ–Њ–Ј—П —Б–Є–љ—В–µ–Ј –Є¬†–≤—Л–і–µ–ї–µ–љ–Є–µ –≥–Є—Б—В–∞–Љ–Є–љ–∞ –Є¬†–і—А—Г–≥–Є—Е –њ–µ—А–µ–і–∞—В—З–Є–Ї–Њ–≤. –І—В–Њ –Ї–∞—Б–∞–µ—В—Б—П –ЭвВГ –њ–Њ—Б—В—Б–Є–љ–∞–њ—В–Є—З–µ—Б–Ї–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤, –љ–∞—Е–Њ–і—П—Й–Є—Е—Б—П –љ–∞ —Б–Њ–Љ–µ –Ї–ї–µ—В–Њ–Ї-–Љ–Є—И–µ–љ–µ–є, —В–Њ, –љ–∞–њ—А–Є–Љ–µ—А, –≤¬†—Б—В—А–Є–∞—В—Г–Љ–µ –Њ–љ–Є –≤—Б—В—А–µ—З–∞—О—В—Б—П —З–∞—Б—В–Њ –≤¬†–њ–∞—А–µ —Б¬†–і–Њ—Д–∞–Љ–Є–љ–Њ–≤—Л–Љ–Є —А–µ—Ж–µ–њ—В–Њ—А–∞–Љ–Є DвВВ, –њ–Њ–љ–Є–ґ–∞—П –Є—Е —Б—А–Њ–і—Б—В–≤–Њ —Б¬†–ї–Є–≥–∞–љ–і–∞–Љ–Є. –Ш–љ—В–µ—А–µ—Б–љ—Л–Љ —Б–≤–Њ–є—Б—В–≤–Њ–Љ –ЭвВГ-—А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ —П–≤–ї—П–µ—В—Б—П –Є—Е –≤—Л—Б–Њ–Ї–∞—П –Ї–Њ–љ—Б—В–Є—В—Г—В–Є–≤–љ–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М, —В–Њ –µ—Б—В—М —Б–њ–Њ–љ—В–∞–љ–љ–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –≤¬†–Њ—В—Б—Г—В—Б—В–≤–Є–µ –≥–Є—Б—В–∞–Љ–Є–љ–∞, –Ї–Њ—В–Њ—А–∞—П –Є–≥—А–∞–µ—В –≤–∞–ґ–љ—Г—О —А–Њ–ї—М –≤¬†–≥–Њ–ї–Њ–≤–љ–Њ–Љ –Љ–Њ–Ј–≥–µ. –ЭвВГ-—А–µ—Ж–µ–њ—В–Њ—А—Л —Г—З–∞—Б—В–≤—Г—О—В –≤¬†—А–µ–≥—Г–ї—П—Ж–Є–Є —Б–љ–∞¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†–Ї–Њ–≥–љ–Є—В–Є–≤–љ—Л—Е —Д—Г–љ–Ї—Ж–Є–є –њ—Г—В–µ–Љ –Љ–Њ–і—Г–ї—П—Ж–Є–Є –≤—Л–±—А–Њ—Б–∞ –Є–ї–Є —Б–Є–љ—В–µ–Ј–∞ –≥–Є—Б—В–∞–Љ–Є–љ–∞ –Є¬†–і—А—Г–≥–Є—Е –љ–µ–є—А–Њ–њ–µ—А–µ–і–∞—В—З–Є–Ї–Њ–≤. –£–ґ–µ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –Њ–±—А–∞—В–Є–Љ—Л—Е –∞–≥–Њ–љ–Є—Б—В–Њ–≤ –ЭвВГ-—А–µ—Ж–µ–њ—В–Њ—А–Њ–≤, —Б–њ–Њ—Б–Њ–±–љ—Л—Е –±–ї–Њ–Ї–Є—А–Њ–≤–∞—В—М –Є—Е —Б–њ–Њ–љ—В–∞–љ–љ—Г—О –∞–Ї—В–Є–≤–љ–Њ—Б—В—М, –њ—А–Њ—Е–Њ–і—П—В –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–µ –Є—Б–њ—Л—В–∞–љ–Є—П —Г¬†–±–Њ–ї—М–љ—Л—Е —И–Є–Ј–Њ—Д—А–µ–љ–Є–µ–є, —Н–њ–Є–ї–µ–њ—Б–Є–µ–є, –љ–∞—А–Ї–Њ–ї–µ–њ—Б–Є–µ–є, –Њ–ґ–Є—А–µ–љ–Є–µ–Љ –Є¬†–±–Њ–ї–µ–Ј–љ—М—О –Р–ї—М—Ж–≥–µ–є–Љ–µ—А–∞. –Ф—А—Г–≥–Є–Љ –Њ—В–ї–Є—З–Є—В–µ–ї—М–љ—Л–Љ —Б–≤–Њ–є—Б—В–≤–Њ–Љ —П–≤–ї—П–µ—В—Б—П –Љ–љ–Њ–ґ–µ—Б—В–≤–µ–љ–љ–Њ—Б—В—М –Є–Ј–Њ—Д–Њ—А–Љ —Н—В–Њ–≥–Њ —А–µ—Ж–µ–њ—В–Њ—А–∞, –њ—А–Њ–Є—Б—Е–Њ–і—П—Й–Є—Е –Є–Ј –Њ–±—Й–µ–≥–Њ –≥–µ–љ–∞ –Є¬†–Њ–±—А–∞–Ј—Г—О—Й–Є—Е—Б—П –Ј–∞ —Б—З–µ—В –∞–ї—М—В–µ—А–љ–∞—В–Є–≤–љ–Њ–≥–Њ —Б–њ–ї–∞–є—Б–Є–љ–≥–∞ [13].
–Ш–љ—В–µ—А–µ—Б–љ–Њ, —З—В–Њ –Ї—А–Њ–Љ–µ –љ–µ–є—А–Њ–љ–∞–ї—М–љ—Л—Е, —В—Г—З–љ—Л—Е –Є¬†–Љ–Є–Ї—А–Њ–≥–ї–Є–∞–ї—М–љ—Л—Е –Ї–ї–µ—В–Њ–Ї –≥–Є—Б—В–∞–Љ–Є–љ–і–µ–Ї–∞—А–±–Њ–Ї—Б–Є–ї–∞–Ј—Г —Н–Ї—Б–њ—А–µ—Б—Б–Є—А—Г—О—В –Ї–ї–µ—В–Ї–Є —Н–њ–µ–љ–і–Є–Љ—Л –≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞. –≠—В–Њ—В –≥–Є—Б—В–∞–Љ–Є–љ –Љ–Њ–ґ–µ—В –±—Л—В—М –≤–Њ–≤–ї–µ—З–µ–љ –≤¬†—А–µ–≥—Г–ї—П—Ж–Є—О –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П —Б—В–≤–Њ–ї–Њ–≤—Л—Е –Ї–ї–µ—В–Њ–Ї, —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–љ—Л—Е –њ–Њ–і —Н–њ–µ–љ–і–Є–Љ–∞–ї—М–љ—Л–Љ —Б–ї–Њ–µ–Љ. –Э–µ–є—А–Њ–љ–∞–ї—М–љ—Л–µ —Б—В–≤–Њ–ї–Њ–≤—Л–µ –Ї–ї–µ—В–Ї–Є in vitro —А–µ–∞–≥–Є—А—Г—О—В –љ–∞ –ї–Є–≥–∞–љ–і—Л —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –ЭвВБ¬†–Є¬†–ЭвВВ [13].
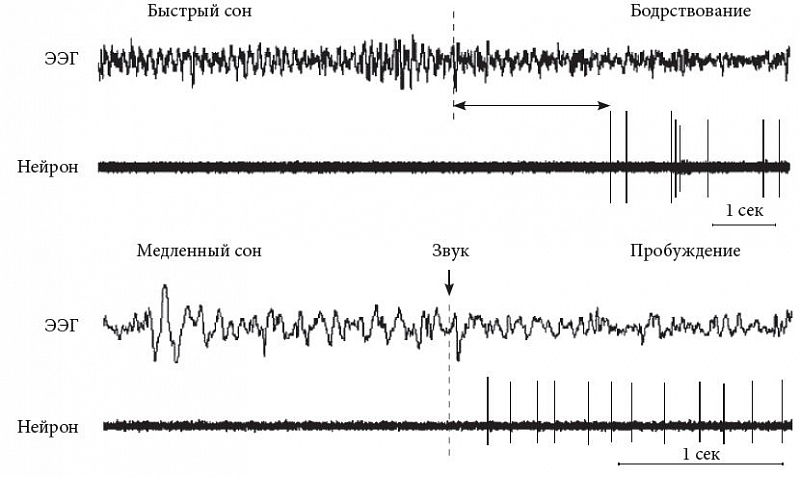
–У–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є–µ –љ–µ–є—А–Њ–љ—Л —П–≤–ї—П—О—В—Б—П —А–Є—В–Љ–Њ–≤–Њ–і–Є—В–µ–ї—П–Љ–Є –Є¬†–і–µ–Љ–Њ–љ—Б—В—А–Є—А—Г—О—В —А–µ–≥—Г–ї—П—А–љ—Л–µ —Б–њ–Њ–љ—В–∞–љ–љ—Л–µ –љ–Є–Ј–Ї–Њ—З–∞—Б—В–Њ—В–љ—Л–µ —А–∞–Ј—А—П–і—Л (1вАУ4¬†–У—Ж). –Я—А–Є –њ—А–Њ–±—Г–ґ–і–µ–љ–Є–Є –Є¬†–њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–∞—Ж–Є–Є –Є—Е —З–∞—Б—В–Њ—В–∞ –≤–Њ–Ј—А–∞—Б—В–∞–µ—В, –њ—А–Є –Ј–∞—Б—Л–њ–∞–љ–Є–Є –Є¬†–Љ–µ–і–ї–µ–љ–љ–Њ–Љ —Б–љ–µ¬†вАУ —Б–љ–Є–ґ–∞–µ—В—Б—П, –њ—А–Є –±—Л—Б—В—А–Њ–Љ —Б–љ–µ¬†вАУ –Є—Б—З–µ–Ј–∞–µ—В (—А–Є—Б.¬†4). –Ґ–Њ—А–Љ–Њ–ґ–µ–љ–Є–µ –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е –љ–µ–є—А–Њ–љ–Њ–≤ –≤–Њ —Б–љ–µ –Њ–њ–Њ—Б—А–µ–і—Г–µ—В—Б—П –У–Р–Ь–Ъ–µ—А–≥–Є—З–µ—Б–Ї–Є–Љ–Є –љ–µ–є—А–Њ–љ–∞–Љ–Є ¬Ђ—Ж–µ–љ—В—А–∞ —Б–љ–∞¬ї –≤¬†–≤–µ–љ—В—А–Њ-–ї–∞—В–µ—А–∞–ї—М–љ–Њ–є –њ—А–µ–Њ–њ—В–Є—З–µ—Б–Ї–Њ–є –Њ–±–ї–∞—Б—В–Є (ventrolateral preoptic nucleus, VLPO) [13]. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –љ–∞ –љ–µ–є—А–Њ–љ—Л —В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–≥–Њ —П–і—А–∞ –Њ–Ї–∞–Ј—Л–≤–∞—О—В –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ —В–Њ—А–Љ–Њ–Ј–љ—Л–µ –љ–µ–є—А–Њ–њ–µ–њ—В–Є–і—Л¬†вАУ –≥–∞–ї–∞–љ–Є–љ –Є¬†—Н–љ–і–Њ–Љ–Њ—А—Д–Є–љ. –У–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –љ–∞ –Ї–ї–µ—В–Ї–∞—Е VLPO –љ–µ—В, —В–∞–Ї —З—В–Њ –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є–µ —Н—В–Є—Е –і–≤—Г—Е —Б–Є—Б—В–µ–Љ¬†вАУ –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Њ–≥–Њ ¬Ђ—Ж–µ–љ—В—А–∞ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П¬ї —В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–≥–Њ —П–і—А–∞ –Є¬†–У–Р–Ь–Ъ–µ—А–≥–Є—З–µ—Б–Ї–Њ–≥–Њ ¬Ђ—Ж–µ–љ—В—А–∞ —Б–љ–∞¬ї –≤–µ–љ—В—А–Њ-–ї–∞—В–µ—А–∞–ї—М–љ–Њ–є –њ—А–µ–Њ–њ—В–Є—З–µ—Б–Ї–Њ–є –Њ–±–ї–∞—Б—В–Є¬†вАУ –љ–Њ—Б–Є—В –Њ–і–љ–Њ—Б—В–Њ—А–Њ–љ–љ–Є–є —Е–∞—А–∞–Ї—В–µ—А. –°—З–Є—В–∞–µ—В—Б—П, —З—В–Њ —В–∞–Ї–Њ–µ –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є–µ –≥–Є—Б—В–∞–Љ–Є–љ–Њ–≤–Њ–є —Б–Є—Б—В–µ–Љ—Л —Б¬†–∞–Ї—В–Є–≤–Є—А—Г—О—Й–µ–є (–Њ—А–µ–Ї—Б–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Њ–є) –Є¬†—В–Њ—А–Љ–Њ–Ј–љ–Њ–є (–У–Р–Ь–Ъ–µ—А–≥–Є—З–µ—Б–Ї–Њ–є) –њ—А–Є–і–∞–µ—В –і–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ—Г—О —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В—М –≤—Б–µ–Љ—Г –Љ–µ—Е–∞–љ–Є–Ј–Љ—Г.
–Э–µ–є—А–Њ–љ–∞–ї—М–љ—Л–є –≥–Є—Б—В–∞–Љ–Є–љ —Г—З–∞—Б—В–≤—Г–µ—В –≤–Њ –Љ–љ–Њ–ґ–µ—Б—В–≤–µ —Д—Г–љ–Ї—Ж–Є–є –Љ–Њ–Ј–≥–∞: –њ–Њ–і–і–µ—А–ґ–∞–љ–Є–Є –≥–Њ–Љ–µ–Њ—Б—В–∞–Ј–∞ –Љ–Њ–Ј–≥–Њ–≤–Њ–є —В–Ї–∞–љ–Є, —А–µ–≥—Г–ї—П—Ж–Є–Є –љ–µ–Ї–Њ—В–Њ—А—Л—Е –љ–µ–є—А–Њ—Н–љ–і–Њ–Ї—А–Є–љ–љ—Л—Е —Д—Г–љ–Ї—Ж–Є–є, –њ–Њ–≤–µ–і–µ–љ–Є—П, –±–Є–Њ—А–Є—В–Љ–Њ–≤, —А–µ–њ—А–Њ–і—Г–Ї—Ж–Є–Є, —В–µ–Љ–њ–µ—А–∞—В—Г—А—Л –Є¬†–Љ–∞—Б—Б—Л —В–µ–ї–∞, —Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–Њ–≥–Њ –Њ–±–Љ–µ–љ–∞ –Є¬†–≤–Њ–і–љ–Њ–≥–Њ –±–∞–ї–∞–љ—Б–∞, –≤¬†—А–µ–∞–Ї—Ж–Є–Є –љ–∞ —Б—В—А–µ—Б—Б. –Ъ—А–Њ–Љ–µ –њ–Њ–і–і–µ—А–ґ–∞–љ–Є—П –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –Љ–Њ–Ј–≥–Њ–≤–Њ–є –≥–Є—Б—В–∞–Љ–Є–љ —Г—З–∞—Б—В–≤—Г–µ—В –≤¬†—Б–µ–љ—Б–Њ—А–љ—Л—Е –Є¬†–Љ–Њ—В–Њ—А–љ—Л—Е —А–µ–∞–Ї—Ж–Є—П—Е, —А–µ–≥—Г–ї—П—Ж–Є–Є —Н–Љ–Њ—Ж–Є–Њ–љ–∞–ї—М–љ–Њ—Б—В–Є, –Њ–±—Г—З–µ–љ–Є–Є –Є¬†–њ–∞–Љ—П—В–Є [13].
–Т–њ–µ—А–≤—Л–µ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–µ–љ–Є—П –Њ¬†—В–Њ–Љ, —З—В–Њ –≥–Є—Б—В–∞–Љ–Є–љ —П–≤–ї—П–µ—В—Б—П ¬Ђ–≥–Њ—А–Љ–Њ–љ–Њ–Љ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П¬ї, –њ–Њ—П–≤–Є–ї–Є—Б—М –њ–Њ—Б–ї–µ —В–Њ–≥–Њ, –Ї–∞–Ї –±—Л–ї–Є –Њ–±–љ–∞—А—Г–ґ–µ–љ—Л –њ–Њ–±–Њ—З–љ—Л–µ —Б–љ–Њ—В–≤–Њ—А–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л –њ–µ—А–≤–Њ–≥–Њ –њ–Њ–Ї–Њ–ї–µ–љ–Є—П –∞–љ—В–Є–≥–Є—Б—В–∞–Љ–Є–љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤, –њ—А–Њ—Е–Њ–і—П—Й–Є—Е –≥–µ–Љ–∞—В–Њ—Н–љ—Ж–µ—Д–∞–ї–Є—З–µ—Б–Ї–Є–є –±–∞—А—М–µ—А (–і–Є—Д–µ–љ–≥–Є–і—А–∞–Љ–Є–љ). –Ф–∞–ї–µ–µ –±—Л–ї–Њ –Њ–±–љ–∞—А—Г–ґ–µ–љ–Њ, —З—В–Њ –љ–µ–є—А–Њ–љ—Л —В—Г–±–µ—А–Њ–Љ–∞–Љ–Є–ї–ї—П—А–љ–Њ–≥–Њ —П–і—А–∞ –∞–Ї—В–Є–≤–љ—Л —В–Њ–ї—М–Ї–Њ –≤¬†—Б–Њ—Б—В–Њ—П–љ–Є–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –љ–Њ –љ–µ —Б–љ–∞. –Э–∞–Ї–Њ–љ–µ—Ж, –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ –љ–∞ –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї—Г—О —Б–Є—Б—В–µ–Љ—Г –њ—Г—В–µ–Љ –ї–Є–±–Њ –≤–≤–µ–і–µ–љ–Є—П –∞–љ—В–∞–≥–Њ–љ–Є—Б—В–Њ–≤ –ЭвВГ-—А–µ—Ж–µ–њ—В–Њ—А–Њ–≤, –Ї–Њ—В–Њ—А—Л–µ –µ–µ –∞–Ї—В–Є–≤–Є—А—Г—О—В, –ї–Є–±–Њ –≤–≤–µ–і–µ–љ–Є—П –∞–ї—М—Д–∞-—Д—В–Њ—А–Љ–µ—В–Є–ї–≥–Є—Б—В–Є–і–Є–љ–∞, –±–ї–Њ–Ї–Є—А—Г—О—Й–µ–≥–Њ —Б–Є–љ—В–µ–Ј –≥–Є—Б—В–∞–Љ–Є–љ–∞, –ї–Є–±–Њ —Г–і–∞–ї–µ–љ–Є—П –≥–µ–љ–∞ –У–Ф–Ъ –љ–∞—А—Г—И–∞–µ—В —Ж–Є–Ї–ї –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П¬†вАУ —Б–љ–∞ —Г¬†–љ–Њ–Ї–∞—Г—В–љ—Л—Е –Љ—Л—И–µ–є [13]. –Ш–Ј –≤—Б–µ—Е –љ—Л–љ–µ –Є–Ј–≤–µ—Б—В–љ—Л—Е –љ–µ–є—А–Њ–љ–љ—Л—Е —Б–Є—Б—В–µ–Љ –Є–Љ–µ–љ–љ–Њ –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–∞—П –љ–∞–Є–±–Њ–ї–µ–µ —З—Г–≤—Б—В–≤–Є—В–µ–ї—М–љ–∞ –Ї¬†–Є–Ј–Љ–µ–љ–µ–љ–Є—О —Г—А–Њ–≤–љ—П –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П¬†
[8, 15].
–£ –Љ—Л—И–µ–є, –љ–Њ–Ї–∞—Г—В–љ—Л—Е –њ–Њ –≥–µ–љ—Г –У–Ф–Ъ, –Њ—В–Љ–µ—З–∞–µ—В—Б—П —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –і–Њ–ї–Є –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–љ–Њ—Б—В–Є –±—Л—Б—В—А–Њ–≥–Њ —Б–љ–∞, —Б–љ–Є–ґ–µ–љ–Є–µ –Љ–Њ—Й–љ–Њ—Б—В–Є —В–µ—В–∞-—А–Є—В–Љ–∞ –≤¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–Є –Є¬†–і–µ–ї—М—В–∞-—А–Є—В–Љ–∞¬†вАУ –≤¬†–Љ–µ–і–ї–µ–љ–љ–Њ–Љ —Б–љ–µ, —Б–љ–Є–ґ–µ–љ–Є–µ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–љ–Њ—Б—В–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –≤¬†—В–µ–Љ–љ–Њ–µ –≤—А–µ–Љ—П —Б—Г—В–Њ–Ї –Є¬†–њ–Њ–≤—Л—И–µ–љ–љ–∞—П —Б–Њ–љ–ї–Є–≤–Њ—Б—В—М. –Я–Њ—Б–ї–µ–і–љ—П—П –њ—А–Њ—П–≤–ї—П–µ—В—Б—П —Б–љ–Є–ґ–µ–љ–Є–µ–Љ –Ї–∞–Ї —А–µ–∞–Ї—В–Є–≤–љ–Њ—Б—В–Є (–ї–∞—В–µ–љ—В–љ–Њ—Б—В–Є –Ї–Њ —Б–љ—Г –њ–Њ—Б–ї–µ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –љ–∞ –ґ–Є–≤–Њ—В–љ–Њ–µ –њ—А–Њ–±—Г–ґ–і–∞—О—Й–Є–Љ–Є —Б—В–Є–Љ—Г–ї–∞–Љ–Є: –≤—Л–Ї–ї—О—З–µ–љ–Є–µ–Љ —Б–≤–µ—В–∞ –Є–ї–Є –њ–Њ–Љ–µ—Й–µ–љ–Є–µ–Љ –Љ—Л—И–Є –≤¬†–љ–Њ–≤—Г—О –Ї–ї–µ—В–Ї—Г), —В–∞–Ї –Є¬†–∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –≤¬†—В–µ–Љ–љ—Л–є –њ–µ—А–Є–Њ–і —Б—Г—В–Њ–Ї –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–Ї–Њ–љ—В—А–Њ–ї—М–љ—Л–Љ–Є –Љ—Л—И–∞–Љ–Є (wild-type). –Ь—Л—И–Є, –љ–Њ–Ї–∞—Г—В–љ—Л–µ –њ–Њ –≥–µ–љ—Г –У–Ф–Ъ –Є–ї–Є –њ–Њ –≥–µ–љ—Г –ЭвВБ-—А–µ—Ж–µ–њ—В–Њ—А–∞, –±–Њ–ї–µ–µ –∞–Ї—В–Є–≤–љ—Л –≤¬†–і–љ–µ–≤–љ–Њ–µ –≤—А–µ–Љ—П, —З–µ–Љ –Љ—Л—И–Є –Є–Ј –≥—А—Г–њ–њ—Л –Ї–Њ–љ—В—А–Њ–ї—П. –Т–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤—Г—П —Б¬†–У–Р–Ь–Ъ–µ—А–≥–Є—З–µ—Б–Ї–Њ–є, –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–∞—П —Б–Є—Б—В–µ–Љ–∞ —В–Њ—А–Љ–Њ–Ј–Є—В –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–µ –њ—А–Њ—П–≤–ї–µ–љ–Є—П —Б–µ–љ—Б–Є—В–Є–Ј–∞—Ж–Є–Є (–њ–Њ–љ—П—В–Є–µ, –Њ–±—А–∞—В–љ–Њ–µ —В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В–Є), –≤—Л–Ј–≤–∞–љ–љ–Њ–є —Е—А–Њ–љ–Є—З–µ—Б–Ї–Є–Љ –≤–≤–µ–і–µ–љ–Є–µ–Љ –Љ–µ—В–∞–Љ—Д–µ—В–∞–Љ–Є–љ–∞ [16, 17].
–У–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–∞—П —Б–Є—Б—В–µ–Љ–∞ –Є–≥—А–∞–µ—В –≤–∞–ґ–љ—Г—О —А–Њ–ї—М –≤¬†—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–Є –љ–∞—А–Ї–Њ–ї–µ–њ—В–Є—З–µ—Б–Ї–Њ–≥–Њ —Д–µ–љ–Њ—В–Є–њ–∞. –•–Њ—В—П —Б–∞–Љ–Њ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–µ —Б–≤—П–Ј–∞–љ–Њ —Б¬†–љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ—Б—В—М—О –Њ—А–µ–Ї—Б–Є–љ–Њ–≤–Њ–є –њ–µ—А–µ–і–∞—З–Є, –≤¬†–Њ–њ—Л—В–∞—Е –љ–∞ —Б–Њ–±–∞–Ї–∞—Е-¬Ђ–љ–∞—А–Ї–Њ–ї–µ–њ—В–Є–Ї–∞—Е¬ї –±—Л–ї–Њ –њ–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ –≤–Њ –≤—А–µ–Љ—П –Ї–∞—В–∞–њ–ї–µ–Ї—Б–Є—З–µ—Б–Ї–Є—Е –њ—А–Є—Б—В—Г–њ–Њ–≤ –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е –љ–µ–є—А–Њ–љ–Њ–≤ —Б–Њ—Е—А–∞–љ—П–µ—В—Б—П, –≤¬†—В–Њ –≤—А–µ–Љ—П –Ї–∞–Ї —Б–µ—А–Њ—В–Њ–љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е¬†вАУ —А–µ–Ј–Ї–Њ —Б–љ–Є–ґ–∞–µ—В—Б—П, –∞¬†–љ–Њ—А–∞–і—А–µ–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е¬†вАУ –≤–Њ–≤—Б–µ –њ—А–µ–Ї—А–∞—Й–∞–µ—В—Б—П [18]. –Я—А–Є —Н—В–Њ–Љ –∞–љ—В–∞–≥–Њ–љ–Є—Б—В—Л –ЭвВГ-—А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ —Б–љ–Є–ґ–∞—О—В –Є–Ј–±—Л—В–Њ—З–љ—Г—О —Б–Њ–љ–ї–Є–≤–Њ—Б—В—М –Є¬†—З–∞—Б—В–Њ—В—Г –Ї–∞—В–∞–њ–ї–µ–Ї—Б–Є—З–µ—Б–Ї–Є—Е –њ—А–Є—Б—В—Г–њ–Њ–≤, –±–ї–Њ–Ї–Є—А—Г—П, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, —В–Њ—А–Љ–Њ–Ј–љ—Л–µ –∞—Г—В–Њ—А–µ—Ж–µ–њ—В–Њ—А—Л, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—Й–Є–µ –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Г—О –Њ–±—А–∞—В–љ—Г—О —Б–≤—П–Ј—М, —З—В–Њ –њ—А–Є–≤–Њ–і–Є—В –Ї¬†—Г–≤–µ–ї–Є—З–µ–љ–Є—О –≤—Л–±—А–Њ—Б–∞ –≥–Є—Б—В–∞–Љ–Є–љ–∞ –≤¬†—Б–Є–љ–∞–њ—В–Є—З–µ—Б–Ї–Є–µ —Й–µ–ї–Є. –£–ґ–µ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –≤–µ—Й–µ—Б—В–≤ —В–∞–Ї–Њ–≥–Њ —А–Њ–і–∞ –њ—А–Њ—Е–Њ–і—П—В –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–µ –Є—Б–њ—Л—В–∞–љ–Є—П –≤¬†–Ї–∞—З–µ—Б—В–≤–µ –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ—Л—Е —Б—А–µ–і—Б—В–≤ –і–ї—П –ї–µ—З–µ–љ–Є—П –љ–∞—А–Ї–Њ–ї–µ–њ—Б–Є–Є [14].
–Ъ—А–Њ–Љ–µ —Н—В–Њ–≥–Њ, –Љ–Њ–і—Г–ї—П—Ж–Є—П –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Њ–є —Б–Є—Б—В–µ–Љ—Л –Љ–Њ–ґ–µ—В –±—Л—В—М –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–∞ –і–ї—П –ї–µ—З–µ–љ–Є—П –Є¬†–і—А—Г–≥–Є—Е –љ–∞—А—Г—И–µ–љ–Є–є —Ж–Є–Ї–ї–∞ ¬Ђ—Б–Њ–љ¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬ї. –Ґ–∞–Ї, —В—А–Є—Ж–Є–Ї–ї–Є—З–µ—Б–Ї–Є–є –∞–љ—В–Є–і–µ–њ—А–µ—Б—Б–∞–љ—В –і–Њ–Ї—Б–µ–њ–Є–љ –љ–µ —В–Њ–ї—М–Ї–Њ —В–Њ—А–Љ–Њ–Ј–Є—В –Њ–±—А–∞—В–љ—Л–є –Ј–∞—Е–≤–∞—В –љ–Њ—А–∞–і—А–µ–љ–∞–ї–Є–љ–∞ –Є¬†—Б–µ—А–Њ—В–Њ–љ–Є–љ–∞, –љ–Њ –Є¬†—П–≤–ї—П–µ—В—Б—П –∞–љ—В–∞–≥–Њ–љ–Є—Б—В–Њ–Љ —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –ЭвВБ –Є¬†–ЭвВВ –Є, –≤—Б–ї–µ–і—Б—В–≤–Є–µ —Н—В–Њ–≥–Њ, —Б¬†—Г—Б–њ–µ—Е–Њ–Љ –њ—А–Є–Љ–µ–љ—П–µ—В—Б—П –і–ї—П –ї–µ—З–µ–љ–Є—П –Є–љ—Б–Њ–Љ–љ–Є–Є —Г¬†–њ–Њ–ґ–Є–ї—Л—Е –±–Њ–ї—М–љ—Л—Е. –Э–∞–Њ–±–Њ—А–Њ—В, –Є–Ј–±—Л—В–Њ—З–љ–∞—П —Б–Њ–љ–ї–Є–≤–Њ—Б—В—М –Љ–Њ–ґ–µ—В –±—Л—В—М –њ–Њ–і–∞–≤–ї–µ–љ–∞ –≤–≤–µ–і–µ–љ–Є–µ–Љ –∞–љ—В–∞–≥–Њ–љ–Є—Б—В–Њ–≤ –ЭвВГ-—А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ [14].
–Ш–љ—Б–Њ–Љ–љ–Є—П —П–≤–ї—П–µ—В—Б—П —Б–∞–Љ—Л–Љ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–љ—Л–Љ –љ–∞—А—Г—И–µ–љ–Є–µ–Љ —Б–љ–∞ –Є, –њ–Њ —А–∞–Ј–љ—Л–Љ –і–∞–љ–љ—Л–Љ, –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В —Б–µ—А—М–µ–Ј–љ—Г—О –њ—А–Њ–±–ї–µ–Љ—Г –і–ї—П 12вАУ22% –љ–∞—Б–µ–ї–µ–љ–Є—П. –§–∞—А–Љ–∞–Ї–Њ—В–µ—А–∞–њ–Є—П –љ–∞—А—Г—И–µ–љ–Є–є —Б–љ–∞, –љ–µ—Б–Љ–Њ—В—А—П –љ–∞ –±–Њ–ї—М—И–Њ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ —Б–љ–Њ—В–≤–Њ—А–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤, –Њ—Б—В–∞–µ—В—Б—П —Б–ї–Њ–ґ–љ–Њ–є –Ј–∞–і–∞—З–µ–є. –Т¬†—Ж–µ–ї–Њ–Љ –≥–Є–њ–љ–Њ—В–Є–Ї–Є –Љ–Њ–ґ–љ–Њ —А–∞–Ј–і–µ–ї–Є—В—М –љ–∞ pro S (pro sleep)¬†вАУ –і–ї—П —Б–љ–∞¬†вАУ –Є¬†anti W (anti wake)¬†вАУ –њ—А–Њ—В–Є–≤ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П [19, 20]. –Т¬†–љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П –±–Њ–ї—М—И–Є–љ—Б—В–≤–Њ —Б–љ–Њ—В–≤–Њ—А–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Њ—В–љ–Њ—Б—П—В –Ї¬†pro sleep –≥—А—Г–њ–њ–µ. –Ю–љ–Є –≤¬†–Њ—Б–љ–Њ–≤–љ–Њ–Љ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л —Б–љ–Њ—В–≤–Њ—А–љ—Л–Љ–Є —Б—А–µ–і—Б—В–≤–∞–Љ–Є, –≤–Њ–Ј–і–µ–є—Б—В–≤—Г—О—Й–Є–Љ–Є –љ–∞ –њ–Њ—Б—В—Б–Є–љ–∞–њ—В–Є—З–µ—Б–Ї–Є–є –У–Р–Ь–Ъ–µ—А–≥–Є—З–µ—Б–Ї–Є–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б. –°—О–і–∞ –≤—Е–Њ–і—П—В –±–∞—А–±–Є—В—Г—А–∞—В—Л, –±–µ–љ–Ј–Њ–і–Є–∞–Ј–µ–њ–Є–љ—Л –Є¬†Z-–њ—А–µ–њ–∞—А–∞—В—Л (–Ј–Њ–њ–Є–Ї–ї–Њ–љ, –Ј–Њ–ї–њ–Є–і–µ–Љ –Є¬†–Ј–∞–ї–µ–њ–ї–Њ–љ). –Т¬†—Н—В–Њ–є —Б–≤—П–Ј–Є –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В—Б—П –Є–љ—В–µ—А–µ—Б–љ—Л–Љ –њ—А–Њ–∞–љ–∞–ї–Є–Ј–Є—А–Њ–≤–∞—В—М —Б—Г—Й–µ—Б—В–≤—Г—О—Й–Є–є –Њ–њ—Л—В –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –≤¬†–Ї–∞—З–µ—Б—В–≤–µ –≥–Є–њ–љ–Њ—В–Є–Ї–∞ —В–Є–њ–Є—З–љ–Њ–≥–Њ anti W –њ—А–µ–њ–∞—А–∞—В–∞¬†вАУ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ —Б—Г–Ї—Ж–Є–љ–∞—В–∞ (–Ф–Њ–љ–Њ—А–Љ–Є–ї).
–Ф–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ —Б—Г–Ї—Ж–Є–љ–∞—В —П–≤–ї—П–µ—В—Б—П –∞–љ—В–∞–≥–Њ–љ–Є—Б—В–Њ–Љ –ЭвВБ-–≥–Є—Б—В–∞–Љ–Є–љ–Њ–≤—Л—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤, —Б¬†–≤—А–µ–Љ–µ–љ–µ–Љ –і–Њ—Б—В–Є–ґ–µ–љ–Є—П –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ–є –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –≤¬†–њ–ї–∞–Ј–Љ–µ, —А–∞–≤–љ—Л–Љ 2¬†—З–∞—Б–∞–Љ, –Є¬†–њ–µ—А–Є–Њ–і–Њ–Љ –њ–Њ–ї—Г–≤—Л–≤–µ–і–µ–љ–Є—П 10¬†—З–∞—Б–Њ–≤. –Я—А–Є–Љ–µ—А–љ–Њ 60% –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ –≤—Л–≤–Њ–і–Є—В—Б—П —Б¬†–Љ–Њ—З–Њ–є –≤¬†–љ–µ–Є–Ј–Љ–µ–љ–µ–љ–љ–Њ–Љ –≤–Є–і–µ, –∞¬†–µ–≥–Њ –Љ–µ—В–∞–±–Њ–ї–Є—В—Л –љ–µ–∞–Ї—В–Є–≤–љ—Л.
–С—Л–ї –њ—А–Њ–≤–µ–і–µ–љ —А—П–і –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –њ–Њ –њ—А–Є–Љ–µ–љ–µ–љ–Є—О —Н—В–Њ–≥–Њ –њ—А–µ–њ–∞—А–∞—В–∞ –≤¬†–Ї–∞—З–µ—Б—В–≤–µ —Б–љ–Њ—В–≤–Њ—А–љ–Њ–≥–Њ —Б—А–µ–і—Б—В–≤–∞. –Т¬†–Њ–і–љ–Њ–Љ –Є–Ј –љ–Є—Е, —А–∞–љ–і–Њ–Љ–Є–Ј–Є—А–Њ–≤–∞–љ–љ–Њ–Љ –і–≤–Њ–є–љ–Њ–Љ —Б–ї–µ–њ–Њ–Љ —Б¬†3¬†–њ–∞—А–∞–ї–ї–µ–ї—М–љ—Л–Љ–Є –≥—А—Г–њ–њ–∞–Љ–Є, —Б—А–∞–≤–љ–Є–≤–∞–ї–∞—Б—М —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М –Є¬†–њ–µ—А–µ–љ–Њ—Б–Є–Љ–Њ—Б—В—М –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ —Б—Г–Ї—Ж–Є–љ–∞—В–∞ (15¬†–Љ–≥) —Б¬†–Ј–Њ–ї–њ–Є–і–µ–Љ–Њ–Љ (10¬†–Љ–≥) –Є¬†–њ–ї–∞—Ж–µ–±–Њ –њ—А–Є –ї–µ—З–µ–љ–Є–Є –Є–љ—Б–Њ–Љ–љ–Є–Є (—Б –Є–Ј—Г—З–µ–љ–Є–µ–Љ —Б–Є–љ–і—А–Њ–Љ–∞ –Њ—В–Љ–µ–љ—Л) [21]. –Т¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є –њ—А–Є–љ—П–ї–Є —Г—З–∞—Б—В–Є–µ 338¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ –≤¬†–≤–Њ–Ј—А–∞—Б—В–µ –Њ—В 18 –і–Њ 73¬†–ї–µ—В. –°¬†–Њ–і–љ–Њ–є —Б—В–Њ—А–Њ–љ—Л, —А–µ–Ј—Г–ї—М—В–∞—В—Л –њ–Њ–і—В–≤–µ—А–і–Є–ї–Є –њ—А–µ–≤–Њ—Б—Е–Њ–і—Б—В–≤–Њ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ –љ–∞–і –њ–ї–∞—Ж–µ–±–Њ –≤¬†–Њ—В–љ–Њ—И–µ–љ–Є–Є —Б–љ–Њ—В–≤–Њ—А–љ–Њ–≥–Њ –і–µ–є—Б—В–≤–Є—П, —Б¬†–і—А—Г–≥–Њ–є¬†вАУ –њ–Њ–Ї–∞–Ј–∞–ї–Є —Б–Њ–њ–Њ—Б—В–∞–≤–Є–Љ—Г—О —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ –Є¬†–Ј–Њ–ї–њ–Є–і–µ–Љ–∞, –њ–Њ–і—В–≤–µ—А–і–Є–ї–Є —Е–Њ—А–Њ—И—Г—О –њ–µ—А–µ–љ–Њ—Б–Є–Љ–Њ—Б—В—М –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ –Є¬†–Ј–Њ–ї–њ–Є–і–µ–Љ–∞ –Є¬†–љ–µ –≤—Л—П–≤–Є–ї–Є —Б–Є–љ–і—А–Њ–Љ–∞ –Њ—В–Љ–µ–љ—Л –њ—А–Є –њ—А–µ–Ї—А–∞—Й–µ–љ–Є–Є –њ—А–Є–µ–Љ–∞ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞.
–Э–∞–Љ–Є –±—Л–ї–Њ –њ—А–Њ–≤–µ–і–µ–љ–Њ –Њ—В–Ї—А—Л—В–Њ–µ –љ–µ—Б—А–∞–≤–љ–Є—В–µ–ї—М–љ–Њ–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –њ—А–µ–њ–∞—А–∞—В–∞ –Ф–Њ–љ–Њ—А–Љ–Є–ї —Б¬†—Ж–µ–ї—М—О –Њ—Ж–µ–љ–Ї–Є –µ–≥–Њ —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є –Є¬†–±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В–Є —Г¬†–±–Њ–ї—М–љ—Л—Е –Є–љ—Б–Њ–Љ–љ–Є–µ–є [22]. –°¬†–њ–Њ–Љ–Њ—Й—М—О –∞–љ–Ї–µ—В–љ–Њ–≥–Њ –Љ–µ—В–Њ–і–∞, –њ—Б–Є—Е–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —В–µ—Б—В–Њ–≤, –њ–Њ–ї–Є—Б–Њ–Љ–љ–Њ–≥—А–∞—Д–Є–Є –±—Л–ї–Њ –Њ–±—Б–ї–µ–і–Њ–≤–∞–љ–Њ 50¬†–±–Њ–ї—М–љ—Л—Е –Є–љ—Б–Њ–Љ–љ–Є–µ–є –≤¬†–≤–Њ–Ј—А–∞—Б—В–µ –Њ—В¬†24 –і–Њ 60¬†–ї–µ—В (—Б—А–µ–і–љ–Є–є –≤–Њ–Ј—А–∞—Б—В 56,6¬†–ї–µ—В) –і–Њ –Є¬†–њ–Њ—Б–ї–µ 10-–і–љ–µ–≤–љ–Њ–≥–Њ –Ї—Г—А—Б–∞ –ї–µ—З–µ–љ–Є—П –Ф–Њ–љ–Њ—А–Љ–Є–ї–Њ–Љ (15¬†–Љ–≥ –љ–∞ –љ–Њ—З—М). –†–µ–Ј—Г–ї—М—В–∞—В—Л –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ –њ–Њ—Б–ї–µ —В–µ—А–∞–њ–Є–Є –Ф–Њ–љ–Њ—А–Љ–Є–ї–Њ–Љ —Г–ї—Г—З—И–Є–ї–Є—Б—М —В–∞–Ї–Є–µ —Б—Г–±—К–µ–Ї—В–Є–≤–љ—Л–µ —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є —Б–љ–∞, –Ї–∞–Ї –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М —Б–љ–∞, –Ї–∞—З–µ—Б—В–≤–Њ —Б–љ–∞ –Є¬†—Г—В—А–µ–љ–љ–µ–≥–Њ –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П, —Б–љ–Є–Ј–Є–ї–Њ—Б—М –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –љ–Њ—З–љ—Л—Е –њ—А–Њ–±—Г–ґ–і–µ–љ–Є–є –Є¬†–і–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М –Ј–∞—Б—Л–њ–∞–љ–Є—П. –Ю–±—К–µ–Ї—В–Є–≤–љ—Л–µ –њ–Њ–ї–Є—Б–Њ–Љ–љ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Є–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ–і—В–≤–µ—А–і–Є–ї–Є ¬Ђ—Б—Г–±—К–µ–Ї—В–Є–≤–љ—Г—О¬ї —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М –Ф–Њ–љ–Њ—А–Љ–Є–ї–∞, –Њ¬†—З–µ–Љ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О—В: —Г–Љ–µ–љ—М—И–µ–љ–Є–µ –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В–Є –Ј–∞—Б—Л–њ–∞–љ–Є—П; —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Б–љ–∞; —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –≤—А–µ–Љ–µ–љ–Є —Д–∞–Ј—Л –±—Л—Б—В—А–Њ–≥–Њ —Б–љ–∞; —Г–ї—Г—З—И–µ–љ–Є–µ –Є–љ–і–µ–Ї—Б–∞ –Ї–∞—З–µ—Б—В–≤–∞ —Б–љ–∞. –Я–µ—А–µ–љ–Њ—Б–Є–Љ–Њ—Б—В—М —В–µ—А–∞–њ–Є–Є –±—Л–ї–∞ —Е–Њ—А–Њ—И–µ–є¬†вАУ –љ–Є –Њ–і–Є–љ –њ–∞—Ж–Є–µ–љ—В –љ–µ –Њ—В–Љ–µ—В–Є–ї –Ї–∞–Ї–Є—Е-–ї–Є–±–Њ –њ–Њ–±–Њ—З–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤, –≤—Б–µ –њ–∞—Ж–Є–µ–љ—В—Л –њ–Њ–ї–љ–Њ—Б—В—М—О –њ—А–Њ—И–ї–Є –њ–ї–∞–љ–Є—А—Г–µ–Љ—Л–є –Ї—Г—А—Б –ї–µ—З–µ–љ–Є—П. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –љ–∞ —Д–Њ–љ–µ –ї–µ—З–µ–љ–Є—П –Ф–Њ–љ–Њ—А–Љ–Є–ї–Њ–Љ –љ–µ –±—Л–ї–Њ –Њ—В–Љ–µ—З–µ–љ–Њ —Г—Е—Г–і—И–µ–љ–Є—П —В–µ—З–µ–љ–Є—П —Б–Њ–њ—Г—В—Б—В–≤—Г—О—Й–Є—Е —Б–Њ–Љ–∞—В–Є—З–µ—Б–Ї–Є—Е –Є¬†–љ–µ–≤—А–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є.
–Т —А–∞–љ–і–Њ–Љ–Є–Ј–Є—А–Њ–≤–∞–љ–љ–Њ–Љ –і–≤–Њ–є–љ–Њ–Љ —Б–ї–µ–њ–Њ–Љ –њ–µ—А–µ–Ї—А–µ—Б—В–љ–Њ–Љ –њ–ї–∞—Ж–µ–±–Њ–Ї–Њ–љ—В—А–Њ–ї–Є—А—Г–µ–Љ–Њ–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є –Є–Ј—Г—З–∞–ї–Њ—Б—М –≤–ї–Є—П–љ–Є–µ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ –љ–∞ —Б—В—А—Г–Ї—В—Г—А—Г —Б–љ–∞ –Є¬†—Б–Њ—Б—В–Њ—П–љ–Є–µ –Ї–Њ–≥–љ–Є—В–Є–≤–љ—Л—Е —Д—Г–љ–Ї—Ж–Є–є, –њ–∞–Љ—П—В–Є –Є¬†—Б–Ї–Њ—А–Њ—Б—В–Є —А–µ–∞–Ї—Ж–Є–є –њ—А–Є –Њ–і–љ–Њ–Ї—А–∞—В–љ–Њ–Љ –њ—А–Є–µ–Љ–µ 15¬†–Љ–≥ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ —Б—Г–Ї—Ж–Є–љ–∞—В–∞ –Є–ї–Є –њ–ї–∞—Ж–µ–±–Њ —Г¬†–Ј–і–Њ—А–Њ–≤—Л—Е –і–Њ–±—А–Њ–≤–Њ–ї—М—Ж–µ–≤ [23]. –Т¬†—Е–Њ–і–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –±—Л–ї–Њ –≤—Л—П–≤–ї–µ–љ–Њ, —З—В–Њ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є –Њ–±—Й–µ–є –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Б–љ–∞, —З–Є—Б–ї–∞ –њ—А–Њ–±—Г–ґ–і–µ–љ–Є–є –≤–Њ –≤—А–µ–Љ—П —Б–љ–∞, –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В–Є —З–µ—В–≤–µ—А—В–Њ–є —Д–∞–Ј—Л –њ–µ—А–≤–Њ–≥–Њ —Ж–Є–Ї–ї–∞ —Б–љ–∞ –Є¬†–њ–∞—А–∞–і–Њ–Ї—Б–∞–ї—М–љ–Њ–є —Д–∞–Ј—Л –њ–µ—А–≤–Њ–≥–Њ —Ж–Є–Ї–ї–∞ —Б–љ–∞, –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ —Ж–Є–Ї–ї–Њ–≤ —Б–љ–∞ (—Б–Љ–µ–љ—Л —Д–∞–Ј –≥–ї—Г–±–Њ–Ї–Њ–≥–Њ –Є¬†–њ–∞—А–∞–і–Њ–Ї—Б–∞–ї—М–љ–Њ–≥–Њ —Б–љ–∞) –≤¬†–≥—А—Г–њ–њ–µ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ –Є¬†–њ–ї–∞—Ж–µ–±–Њ –љ–µ –Њ—В–ї–Є—З–∞–ї–Є—Б—М. –Я–Њ—Б–ї–µ –њ—А–Є–µ–Љ–∞ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ —Б–љ–Є–ґ–∞–ї–∞—Б—М –Њ–±—Й–∞—П –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М –њ—А–Њ–±—Г–ґ–і–µ–љ–Є–є –≤–Њ –≤—А–µ–Љ—П —Б–љ–∞. –Я—А–Є–µ–Љ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ –њ—А–Є–≤–Њ–і–Є–ї –Ї¬†–Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–Љ—Г —Г–Ї–Њ—А–Њ—З–µ–љ–Є—О –њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–Є –Є¬†—Г–і–ї–Є–љ–µ–љ–Є—О –≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–Є. –Я—А–Є —Н—В–Њ–Љ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ –љ–µ –≤–ї–Є—П–ї –љ–∞ –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М —В—А–µ—В—М–µ–є –Є¬†—З–µ—В–≤–µ—А—В–Њ–є —Б—В–∞–і–Є–є –Є¬†—Д–∞–Ј—Л –±—Л—Б—В—А–Њ–≥–Њ —Б–љ–∞. –С–Њ–ї–µ–µ –њ–Њ–і—А–Њ–±–љ—Л–є –∞–љ–∞–ї–Є–Ј –њ–Њ–Ї–∞–Ј–∞–ї, —З—В–Њ –њ–Њ—Б–ї–µ –њ—А–Є–µ–Љ–∞ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ –≤¬†—Б—А–∞–≤–љ–µ–љ–Є–Є —Б¬†–њ–ї–∞—Ж–µ–±–Њ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ —Г–ї—Г—З—И–∞–ї–Њ—Б—М –Ї–∞—З–µ—Б—В–≤–Њ –Є¬†—Г–≤–µ–ї–Є—З–Є–≤–∞–ї–∞—Б—М –≥–ї—Г–±–Є–љ–∞ —Б–љ–∞, —В–Њ–≥–і–∞ –Ї–∞–Ї —П—Б–љ–Њ—Б—В—М —Б–Њ–Ј–љ–∞–љ–Є—П –Є¬†—Б–Њ—Б—В–Њ—П–љ–Є–µ –њ—А–Є –њ—А–Њ–±—Г–ґ–і–µ–љ–Є–Є –њ—А–Є –њ—А–Є–µ–Љ–µ –њ—А–µ–њ–∞—А–∞—В–∞ –Є¬†–њ–ї–∞—Ж–µ–±–Њ –љ–µ –Њ—В–ї–Є—З–∞–ї–Є—Б—М. –Я—А–Є –њ—А–Є–µ–Љ–µ –і–Њ–Ї—Б–Є–ї–∞–Љ–Є–љ–∞ –љ–Є —Г¬†–Њ–і–љ–Њ–≥–Њ –Є–Ј 18¬†–Є—Б–њ—Л—В—Г–µ–Љ—Л—Е –љ–µ –Њ—В–Љ–µ—З–µ–љ–Њ –Ї–∞–Ї–Є—Е-–ї–Є–±–Њ –Є–Ј–Љ–µ–љ–µ–љ–Є–є –Ї—А–∞—В–Ї–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–є –њ–∞–Љ—П—В–Є –Є¬†—Б–Ї–Њ—А–Њ—Б—В–Є —А–µ–∞–Ї—Ж–Є–є. –Я—А–Є –њ—А–Њ–≤–µ–і–µ–љ–Є–Є –і–∞–љ–љ–Њ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –љ–µ –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М –Ї–∞–Ї–Є—Е-–ї–Є–±–Њ –љ–µ–ґ–µ–ї–∞—В–µ–ї—М–љ—Л—Е —П–≤–ї–µ–љ–Є–є.
–†–µ–Ј—Г–ї—М—В–∞—В—Л –њ—А–Њ–≤–µ–і–µ–љ–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О—В –Њ–± —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є –Є¬†–±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В–Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Ф–Њ–љ–Њ—А–Љ–Є–ї–∞ –њ—А–Є –љ–∞—А—Г—И–µ–љ–Є—П—Е —Б–љ–∞ –Є¬†—И–Є—А–Њ–Ї–Є—Е –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—П—Е –µ–≥–Њ –њ—А–Є–Љ–µ–љ–µ–љ–Є—П. –Ю—В—Б—Г—В—Б—В–≤–Є–µ —Н—Д—Д–µ–Ї—В–∞ –њ–Њ—Б–ї–µ–і–µ–є—Б—В–≤–Є—П –Є, —З—В–Њ –љ–µ–Љ–∞–ї–Њ–≤–∞–ґ–љ–Њ, –і–Њ—Б—В—Г–њ–љ–Њ—Б—В—М –њ—А–µ–њ–∞—А–∞—В–∞ –њ–Њ–Ј–≤–Њ–ї—П—О—В –њ—А–Є–Љ–µ–љ—П—В—М –Ф–Њ–љ–Њ—А–Љ–Є–ї –і–ї—П —Б–Є—В—Г–∞—Ж–Є–Њ–љ–љ–Њ–≥–Њ –њ—А–Є–µ–Љ–∞ –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е —Б—В—А–µ—Б—Б–∞ –њ—А–Є —А–∞–Ј–≤–Є—В–Є–Є –∞–і–∞–њ—В–∞—Ж–Є–Њ–љ–љ–Њ–є —Д–Њ—А–Љ—Л –Є–љ—Б–Њ–Љ–љ–Є–Є. –Я–Њ –і–∞–љ–љ—Л–Љ –Р–Љ–µ—А–Є–Ї–∞–љ—Б–Ї–Њ–є –∞—Б—Б–Њ—Ж–Є–∞—Ж–Є–Є –Љ–µ–і–Є—Ж–Є–љ—Л —Б–љ–∞ (American Academy of Sleep Medicine), 20% –љ–∞—Б–µ–ї–µ–љ–Є—П –Є—Б–њ—Л—В—Л–≤–∞—О—В –Њ—Б—В—А—Г—О –Є–љ—Б–Њ–Љ–љ–Є—О –њ–Њ –Ї—А–∞–є–љ–µ–є –Љ–µ—А–µ —А–∞–Ј –≤¬†–≥–Њ–і, –∞¬†90%¬†вАУ –њ–Њ –Ї—А–∞–є–љ–µ–є –Љ–µ—А–µ —А–∞–Ј –≤¬†–ґ–Є–Ј–љ–Є. –Я–µ—А—Б–Є—Б—В–Є—А–Њ–≤–∞–љ–Є–µ —Б—В—А–µ—Б—Б–∞ –Є¬†—Б–≤—П–Ј–∞–љ–љ—Л–µ —Б¬†–љ–Є–Љ –љ–∞—А—Г—И–µ–љ–Є—П —Б–љ–∞ —П–≤–ї—П—О—В—Б—П –≤–µ–і—Г—Й–Є–Љ —Д–∞–Ї—В–Њ—А–Њ–Љ —Е—А–Њ–љ–Є—Д–Є–Ї–∞—Ж–Є–Є –Є–љ—Б–Њ–Љ–љ–Є–Є. –Я—А–Њ–≤–µ–і–µ–љ–љ—Л–µ –љ–∞–Љ–Є –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ —А–∞–Ј–Њ–≤—Л–є –њ—А–Є–µ–Љ —Б–љ–Њ—В–≤–Њ—А–љ—Л—Е —Б—А–µ–і—Б—В–≤ –њ–Њ—Б–ї–µ –Њ—Б—В—А–Њ–≥–Њ —Н–Љ–Њ—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–≥–Њ —Б—В—А–µ—Б—Б–∞ –њ–Њ–ї–љ–Њ—Б—В—М—О –љ–Є–≤–µ–ї–Є—А–Њ–≤–∞–ї –љ–µ–≥–∞—В–Є–≤–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –њ–Њ—Б–ї–µ–і–љ–µ–≥–Њ –љ–∞ —Б—В—А—Г–Ї—В—Г—А—Г —Б–љ–∞, –∞¬†—В–∞–Ї–ґ–µ –Њ—В–Љ–µ—З–µ–љ–љ–Њ–µ –њ–Њ—Б–ї–µ —Б—В—А–µ—Б—Б–∞ —Г–≤–µ–ї–Є—З–µ–љ–Є–µ —З–∞—Б—В–Њ—В—Л —Б–µ—А–і–µ—З–љ—Л—Е —Б–Њ–Ї—А–∞—Й–µ–љ–Є–є –Є¬†–≤–∞—А–Є–∞–±–µ–ї—М–љ–Њ—Б—В–Є —Б–µ—А–і–µ—З–љ–Њ–≥–Њ —А–Є—В–Љ–∞. –С–Њ–ї–µ–µ –Ї–∞—З–µ—Б—В–≤–µ–љ–љ—Л–є —Б–Њ–љ –њ—А–Є–≤–Њ–і–Є–ї –Ї¬†—Г–ї—Г—З—И–µ–љ–Є—О –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–µ–є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П (–њ–Њ —А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ —В–µ—Б—В–Њ–≤ –і–ї—П –Њ—Ж–µ–љ–Ї–Є –≤–љ–Є–Љ–∞–љ–Є—П, –њ–∞–Љ—П—В–Є, —В–Њ–љ–Ї–Њ–є –Љ–Њ—В–Њ—А–Є–Ї–Є —А—Г–Ї) [24]. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –Ї–Њ—А—А–µ–Ї—Ж–Є—П –њ–∞—А–∞–Љ–µ—В—А–Њ–≤ –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞ –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е —Б—В—А–µ—Б—Б–∞ –љ–µ–є—В—А–∞–ї–Є–Ј—Г–µ—В –љ–µ–≥–∞—В–Є–≤–љ—Л–µ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П —Б—В—А–µ—Б—Б–∞, –њ–Њ–≤—Л—И–∞–µ—В –∞–і–∞–њ—В–Є–≤–љ—Л–µ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–Є —З–µ–ї–Њ–≤–µ–Ї–∞ –Є¬†—П–≤–ї—П–µ—В—Б—П –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Њ–є —А–∞–Ј–≤–Є—В–Є—П —Е—А–Њ–љ–Є—З–µ—Б–Ї–Њ–є –Є–љ—Б–Њ–Љ–љ–Є–Є.
–Х—Й–µ –Њ–і–љ–Є–Љ –≤–∞–ґ–љ—Л–Љ –њ–Њ–Ї–∞–Ј–∞–љ–Є–µ–Љ –Ї¬†–њ—А–Є–Љ–µ–љ–µ–љ–Є—О –Ф–Њ–љ–Њ—А–Љ–Є–ї–∞ –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В—Б—П –Є–љ—Б–Њ–Љ–љ–Є—П –љ–∞ —Д–Њ–љ–µ –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ–Њ–є –Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є. –Э–∞–Є–±–Њ–ї–µ–µ —З–∞—Б—В–Њ–є –њ—А–Є—З–Є–љ–Њ–є –≤–Њ–Ј–љ–Є–Ї–љ–Њ–≤–µ–љ–Є—П –і–∞–љ–љ–Њ–є —Д–Њ—А–Љ—Л –Є–љ—Б–Њ–Љ–љ–Є–Є —П–≤–ї—П–µ—В—Б—П –і–ї–Є—В–µ–ї—М–љ—Л–є –њ—А–Є–µ–Љ —Б–љ–Њ—В–≤–Њ—А–љ—Л—Е —Б—А–µ–і—Б—В–≤, –≤–Њ–Ј–і–µ–є—Б—В–≤—Г—О—Й–Є—Е –љ–∞ –њ–Њ—Б—В—Б–Є–љ–∞–њ—В–Є—З–µ—Б–Ї–Є–є –У–Р–Ь–Ъ–µ—А–≥–Є—З–µ—Б–Ї–Є–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б, –≤¬†–±–Њ–ї—М—И–Є–љ—Б—В–≤–µ —Б–ї—Г—З–∞–µ–≤ –±–µ–љ–Ј–Њ–і–Є–∞–Ј–µ–њ–Є–љ–Њ–≤. –Я—А–Є –і–∞–љ–љ–Њ–Љ —А–∞—Б—Б—В—А–Њ–є—Б—В–≤–µ –Њ–і–Є–љ –Є–Ј —Н—В–∞–њ–Њ–≤ –ї–µ—З–µ–љ–Є—П¬†вАУ –њ–µ—А–µ–≤–Њ–і –њ–∞—Ж–Є–µ–љ—В–∞ –љ–∞ —Б–љ–Њ—В–≤–Њ—А–љ—Л–є –њ—А–µ–њ–∞—А–∞—В –і—А—Г–≥–Њ–є —Е–Є–Љ–Є—З–µ—Б–Ї–Њ–є –≥—А—Г–њ–њ—Л, –≤¬†–Ї–∞—З–µ—Б—В–≤–µ –Ї–Њ—В–Њ—А–Њ–≥–Њ —Б¬†—Г—Б–њ–µ—Е–Њ–Љ –Љ–Њ–ґ–µ—В –±—Л—В—М –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ –Ф–Њ–љ–Њ—А–Љ–Є–ї, —В–∞–Ї –Ї–∞–Ї –Њ–љ —П–≤–ї—П–µ—В—Б—П –∞–љ—В–∞–≥–Њ–љ–Є—Б—В–Њ–Љ —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –і—А—Г–≥–Њ–≥–Њ –љ–µ–є—А–Њ–Љ–µ–і–Є–∞—В–Њ—А–∞. –°–ї–µ–і—Г–µ—В –Њ—В–Љ–µ—В–Є—В—М, —З—В–Њ –Ф–Њ–љ–Њ—А–Љ–Є–ї –Љ–Њ–ґ–љ–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞—В—М –≤–Њ –≤—А–µ–Љ—П –±–µ—А–µ–Љ–µ–љ–љ–Њ—Б—В–Є, –њ–Њ —А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ –∞–і–µ–Ї–≤–∞—В–љ—Л—Е –Є —Е–Њ—А–Њ—И–Њ –Ї–Њ–љ—В—А–Њ–ї–Є—А—Г–µ–Љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є [25].
–Ь–µ–ґ–і—Г —В–µ–Љ –Ф–Њ–љ–Њ—А–Љ–Є–ї, –љ–µ –Њ–±–ї–∞–і–∞—О—Й–Є–є –Љ–Є–Њ—А–µ–ї–∞–Ї—Б–Є—А—Г—О—Й–Є–Љ –і–µ–є—Б—В–≤–Є–µ–Љ, —Б¬†–Њ—Б—В–Њ—А–Њ–ґ–љ–Њ—Б—В—М—О –і–Њ–ї–ґ–µ–љ –њ—А–Є–Љ–µ–љ—П—В—М—Б—П –≤¬†—Б–ї—Г—З–∞—П—Е –њ–Њ–і–Њ–Ј—А–µ–љ–Є—П –љ–∞ –љ–∞–ї–Є—З–Є–µ —Б–Є–љ–і—А–Њ–Љ–∞ –Њ–±—Б—В—А—Г–Ї—В–Є–≤–љ–Њ–≥–Њ –∞–њ–љ–Њ—Н —Б–љ–∞, —З—В–Њ –Њ—Б–Њ–±–µ–љ–љ–Њ –∞–Ї—В—Г–∞–ї—М–љ–Њ —Г¬†–њ–Њ–ґ–Є–ї—Л—Е –±–Њ–ї—М–љ—Л—Е, —В–∞–Ї –Ї–∞–Ї —Г¬†–і–∞–љ–љ–Њ–є –Ї–∞—В–µ–≥–Њ—А–Є–Є –њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Н—В–∞ –±–Њ–ї–µ–Ј–љ—М —З–∞—Б—В–Њ –њ—А–Њ—В–µ–Ї–∞–µ—В —Б—Г–±–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є –Є¬†–Љ–Њ–ґ–µ—В –±—Л—В—М –і–Є–∞–≥–љ–Њ—Б—В–Є—А–Њ–≤–∞–љ–∞ —В–Њ–ї—М–Ї–Њ –њ—А–Є –њ—А–Њ–≤–µ–і–µ–љ–Є–Є –њ–Њ–ї–Є—Б–Њ–Љ–љ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є [25].
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –Љ–љ–Њ–≥–Њ–Њ–±—А–∞–Ј–Є–µ —Б–Є—Б—В–µ–Љ, —Г—З–∞—Б—В–≤—Г—О—Й–Є—Е –≤¬†–Њ—А–≥–∞–љ–Є–Ј–∞—Ж–Є–Є —Б–Њ—Б—В–Њ—П–љ–Є–є —Б–љ–∞ –Є¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –њ–Њ–Ј–≤–Њ–ї—П–µ—В —Г—Б–њ–µ—И–љ–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞—В—М –і–ї—П –Ї–Њ—А—А–µ–Ї—Ж–Є–Є –Є–љ—Б–Њ–Љ–љ–Є–Є –њ—А–µ–њ–∞—А–∞—В—Л, –≤–Њ–Ј–і–µ–є—Б—В–≤—Г—О—Й–Є–µ –љ–∞ —А–∞–Ј–ї–Є—З–љ—Л–µ —А–µ—Ж–µ–њ—В–Њ—А–љ—Л–µ –Ї–Њ–Љ–њ–ї–µ–Ї—Б—Л, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –≤—Е–Њ–і—П—Й–Є–µ –≤¬†—Б–Њ—Б—В–∞–≤ –Њ–і–љ–Њ–є –Є–Ј –љ–∞–Є–±–Њ–ї–µ–µ –≤–∞–ґ–љ—Л—Е –∞–Ї—В–Є–≤–Є—А—Г—О—Й–Є—Е —Б–Є—Б—В–µ–Љ –≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞¬†вАУ –≥–Є—Б—В–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Њ–є.
Neurochemical mechanisms of the regulation of the sleep-wake cycle and wakefulness: role of histamine receptors antihistamines in the treatment of insomnia
V.M. Kovalzon1, K.N. Strygin2
1 Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences
2 I.M. Sechenov First Moscow State Medical University
Contact person: Vladimir Matveyevich Kovalzon, kovalzon@sevin.ru
The authors review structure, morphological and neurochemical chains of the brain histaminergic system, one of the main wakefulness promoting systems. Biochemistry of the histamine turnover and characteristics of histamine receptors are summarized. The authors focuse on the interactions between the histaminergic and orexinergic (hypocretinergic) systems. Examples of effects of experimental modulation of the histaminergic system on the sleep-wake cycle are presented. The authors deduce that orexine neurons coordinate wakefulness promoting system primarily through their effects on histamine neurons. Uses of central HвВБ-antihistamines for the correction of varied forms of insomnia were discussed.
–£–≤–∞–ґ–∞–µ–Љ—Л–є –њ–Њ—Б–µ—В–Є—В–µ–ї—М uMEDp!
–£–≤–µ–і–Њ–Љ–ї—П–µ–Љ –Т–∞—Б –Њ —В–Њ–Љ, —З—В–Њ –Ј–і–µ—Б—М —Б–Њ–і–µ—А–ґ–Є—В—Б—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П, –њ—А–µ–і–љ–∞–Ј–љ–∞—З–µ–љ–љ–∞—П –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –і–ї—П —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П.
–Х—Б–ї–Є –Т—Л –љ–µ —П–≤–ї—П–µ—В–µ—Б—М —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–Љ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П, –∞–і–Љ–Є–љ–Є—Б—В—А–∞—Ж–Є—П –љ–µ –љ–µ—Б–µ—В –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ—Б—В–Є –Ј–∞ –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ –њ–Њ—Б–ї–µ–і—Б—В–≤–Є—П, –≤–Њ–Ј–љ–Є–Ї—И–Є–µ –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–∞–Љ–Њ—Б—В–Њ—П—В–µ–ї—М–љ–Њ–≥–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Т–∞–Љ–Є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є —Б –њ–Њ—А—В–∞–ї–∞ –±–µ–Ј –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є –Ї–Њ–љ—Б—Г–ї—М—В–∞—Ж–Є–Є —Б –≤—А–∞—З–Њ–Љ.
–Э–∞–ґ–Є–Љ–∞—П –љ–∞ –Ї–љ–Њ–њ–Ї—Г ¬Ђ–Т–Њ–є—В–Є¬ї, –Т—Л –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В–µ, —З—В–Њ —П–≤–ї—П–µ—В–µ—Б—М –≤—А–∞—З–Њ–Љ –Є–ї–Є —Б—В—Г–і–µ–љ—В–Њ–Љ –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–≥–Њ –≤—Г–Ј–∞.