Генетика сна и суточных биологических ритмов человека: современные представления
- Аннотация
- Статья
- Ссылки
- English
Введение
2 октября 2017 г. Нобелевская премия по физиологии и медицине была присуждена J.C. Hall, M. Rosbash, M.W. Young «за открытие молекулярных механизмов, контролирующих циркадианный ритм». Лауреаты независимо друг от друга при изучении плодовой мушки Drosophila melanogaster выявили основные молекулярно-генетические механизмы работы биологических часов, контролирующих суточные колебания различных физиологических и поведенческих параметров. В последние годы это направление активно развивалось. Были выявлены механизмы циркадианной регуляции у многих организмов, установлены связи с регуляцией физиологических процессов.
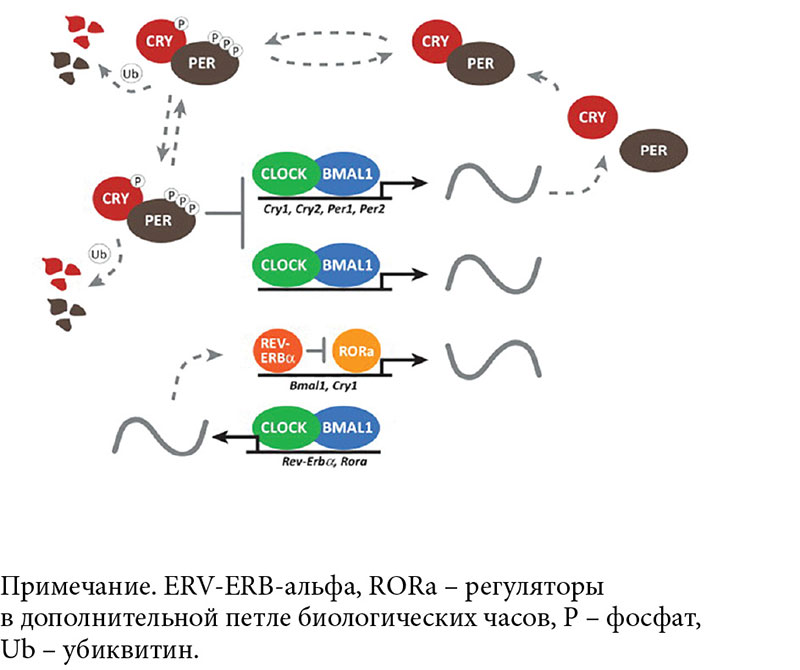
Циркадианный (лат. circa – около, dies – день) процесс проявляется как изменение уровня активности, температуры тела, метаболических и других процессов с периодом около 24 часов. Ведущую роль в формировании суточной ритмики играют биологические часы, имеющие сходный принцип работы у всех млекопитающих. Цикличностью активности и работы систем организма управляют супрахиазматические ядра гипоталамуса (СХЯ), в нейронах которых ритм задается взаимодействием на уровне транскрипции, трансляции и деградации белков – регуляторных факторов. Общий принцип работы системы молекулярных часов основан на взаимодействии систем обратной связи транскрипции и трансляции белков. Этот молекулярный механизм актуален не только для клеток СХЯ, но и для других областей мозга и тканей, но именно осциллятор СХЯ является ведущим в регуляции работы организма (рис. 1).
В основной петле взаимодействуют транскрипционные активаторы CLOCK и BMAL1, которые в виде комплекса активируют транскрипцию генов семейства Period (PER1, PER2, PER3) и криптохромов (CRY1, CRY2). Их белки-транскрипты постепенно накапливаются в цитоплазме в течение дня. Белки PER и CRY попарно связываются, перемещаются в ядро и физически взаимодействуют с комплексом CLOCK/BMAL1, ослабляя собственную транскрипцию. Ночью белки PER и CRY подвергаются убиквитинированию и распадаются в протеосомах. Важную модуляторную роль играет фосфорилирование казеинкиназой 1-эпсилон и 1-дельта. Этот цикл длится приблизительно 24 часа и является ключевым для циркадианных ритмов млекопитающих (см. рис. 1). В дополнительном цикле комплекс CLOCK/BMAL1 активирует транскрипцию ретиноидного орфанного рецептора (RAR-related orphan receptors – ROR – отдельные рецепторы, родственные рецепторам ретиноевой кислоты) и репрессора REV-ERB-альфа, который в свою очередь подавляет транскрипцию BMAL1. Таким образом, CLOCK постоянно экспрессируется в СХЯ, а транскрипция BMAL1 максимальна ночью. Помимо непосредственно элементов молекулярного осциллятора транскрипционные регуляторы молекулярных часов регулируют и многие другие гены, обеспечивая физиологические и поведенческие циркадианные ритмы. Множество эпигенетических, транскрипционных, посттранскрипционных и посттрансляционных механизмов предоставляют возможность регуляции циркадианных колебаний как в мозге, так и в периферических тканях [1].
Генетические исследования на человеке
Большинство молекулярно-генетических исследований на человеке основано на сравнении выраженности того или иного признака у носителей различных полиморфизмов (вариантов) исследуемого гена. Чтобы какой-то вариант гена считался полиморфизмом, он должен быть относительно распространенным в популяции [2]. Большинство исследований сфокусировано на поиске и анализе эффектов однонуклеотидных полиморфизмов (ОНП), при которых различия между аллельными вариантами гена заключаются в одном нуклеотиде. Кроме того, распространен и активно изучается такой тип полиморфизма, как варьирующее число тандемных повторов (Variable Number of Tandem Repeats – VNTR), при котором определенная последовательность нуклеотидов в разных аллелях повторяется разное число раз. Существуют и другие варианты различий между аллелями.
Традиционным для психогенетических исследований был и остается близнецовый метод, который позволяет оценить общую наследуемость признака с помощью сравнения его вариабельности в парах монозиготных и дизиготных близнецов. В исследованиях, особенно в генетической эпидемиологии, применяется подход «случай – контроль», который подразумевает сравнение частоты встречаемости полиморфизмов в общепопуляционной выборке и группе с каким-либо заболеванием. Этот метод используется и в общих психогенетических исследованиях при сравнении групп с выраженными различиями в исследуемых особенностях.
В процессе поиска генетических основ индивидуальных особенностей человека широко применяется подход «ген-кандидат». Определение нейрональных сетей и систем, вовлеченных в обеспечение психических функций, дает возможность отбирать связанные с их развитием и функционированием гены для поиска генетических ассоциаций индивидуальной вариабельности [3].
Благодаря развитию и удешевлению техники для генетического анализа, а также методов биоинформатики появились и новые подходы. Среди них – полногеномные исследования ассоциаций, направленные на анализ ОНП во всем геноме, что позволяет обнаружить новые гены-кандидаты для более узконаправленных исследований. Поскольку полногеномное исследование ассоциаций в анализе не опирается на гипотезы или теории, интерпретация полученных ассоциаций иногда может вызывать затруднения [4].
Среди наиболее вероятных генов-мишеней можно рассматривать те, которые относятся к метаболическим или регуляторным сетям. При обнаружении связи таких мутаций с популяционной вариабельностью интересующих исследователя параметров возможное объяснение механизма действия мутации будет наиболее прямолинейно. Кроме того, исследования мутаций и модификаций этих генов-мишеней на животных моделях позволяют эффективно и подробно описывать результирующие фенотипы, изменения в физиологии и рассматривать молекулярные механизмы.
Для циркадианной регуляции ключевыми объектами являются, во-первых, гены системы молекулярных часов и сами элементы системы. Во-вторых, ферменты метаболизма, отвечающие за модификацию и распад основных элементов. В-третьих, регуляторы транскрипции и трансляции, осуществляющие тонкую настройку системы и принимающие сигнал от нее. В случае исследования параметров сна круг поиска значительно расширяется. Поскольку в регуляцию системы сна и бодрствования входит целый ряд медиаторных систем и структур мозга, то в поле зрения исследователей попадают гены рецепторов и транспортеров медиаторов, синтезирующих и расщепляющих их ферментов, модуляторов возбуждения и передачи сигнала. Важную роль играют полиморфизмы в регуляторных областях ДНК, расположенные вне генов и способные повлиять на их экспрессию. Но и этот спектр мишеней для поиска не описывает все получаемые результаты. При полногеномном поиске ассоциаций обнаруживаются связи с участками, не относящимися ни к одной из этих групп, механизм действия которых пока не удается объяснить.
Хронотип
Уровень активности метаболических и физиологических процессов находится под контролем биологических часов. Для человека нормальны дневная активность и ночной сон, однако режим у разных людей неодинаков. Индивидуальные предпочтения времени сна и активности в течение суток описываются в рамках понятия диурнальных предпочтений, или хронотипа человека. Традиционно выделяют ранний («жаворонки», рано встают и ложатся спать), промежуточный и поздний («совы», поздно встают и ложатся спать) хронотип. Среди мужчин чаще встречается поздний хронотип, но эта разница сглаживается с возрастом [5–7].
Согласно близнецовым и семейным исследованиям, вклад наследственности в хронотип составляет, по разным оценкам, 12–42% [8–10], но генетические основы вариабельности еще не поняты. Мутации в генах компонентов молекулярных часов и их регуляторов в теории могут приводить к изменению или сдвигу ритмов активности и, соответственно, хронотипа. Сейчас уже обнаружен ряд генетических вариантов, способствующих формированию того или иного хронотипа и даже развитию нарушений ритма сна – бодрствования. Нарушения сна, связанные с циркадианным ритмом, определяются постоянными или периодическими нарушениями ритма сна – бодрствования и делятся на три подтипа: синдром задержки фазы сна, синдром опережающей фазы сна (или синдром семейного опережения фазы сна) и несинхронизированный тип. При синдроме задержки фазы сна цикл сна – бодрствования удлинен, люди значительно позже, чем в норме, засыпают и просыпаются. При синдроме семейного опережения фазы сна основной эпизод сна смещен на более раннее время по отношению к желаемому. При несинхронизированном типе каждые сутки время отхода ко сну сдвигается на 30–60 минут вперед.
Полногеномные исследования уже затрагивали вопрос ассоциаций ОНП и параметров хронотипа. В крупном исследовании, включавшем более 89 000 человек и самоотчет о диурнальных предпочтениях, было обнаружено 15 полиморфизмов-кандидатов, ассоциированных с хронотипом. Семь были расположены близко к генам, связанным с циркадианной регуляцией: rs12736689 возле гена RGS16, rs9479402 возле VIP, rs55694368 возле PER2, rs35833281 возле HCRTR2, rs11545787 возле RASD1, rs11121022 возле PER3, rs9565309 возле FBXL3. RGS16 – регулятор проведения сигнала рецепторов, сопряженных с G-белками, вовлеченный в работу биологических часов. Вазоинтестинальный пептид VIP играет ключевую роль в проведении в СХЯ. PER2 – элемент биологических часов, полиморфизм расположен в регуляторной области ДНК. HCRTR2 – ген рецептора орексина типа 2, мутации которого связывают с нарколепсией. RASD1 – ген активатора проведения сигнала G-белками, его удаление приводит к ослаблению реакции биологических часов на свет у мышей, а сам полиморфизм расположен в регуляторной области. Генетические варианты в области PER3 ассоциируют с хронотипом, чувствительностью циркадианной системы к свету. Продукт гена FBXL3 убиквитинирует и таким образом участвует в деградации белков биологических часов CRY1 и CRY2. Еще четыре полиморфизма ассоциированы с генами, потенциально имеющими отношение к циркадианной системе: PLCL1 – rs1595824, APH1A (CA14) – rs34714364, FBXL13 (FAM185A) – rs3972456, NOL4 – rs12965577. Последние четыре обнаруженных ОНП-кандидата в настоящее время не имеют известных ассоциаций близлежащих генов с регуляцией суточного цикла: TOX3 – rs12927162, AK5 – rs10493596, DLX5 (SHFM1) – rs2948276, ALG10B – rs6582618. В целом в результатах исследования было обнаружено значительное количество элементов циркадианной регуляции и зрительной фототрансдукции. Кроме того, по результатам исследования, утреннему типу испытуемых субъективно требовалось менее восьми часов, чтобы выспаться, у них был снижен риск бессонницы и апноэ сна [11].
Еще одно полногеномное исследование хронотипа включало 100 420 человек из когорты UK Biobank и оценивало хронотип по результатам самоотчета по одной шкале из четырех категорий. Поиск ассоциаций дал 12 значимых областей в геноме. Была установлена ассоциация с ОНП рядом с геном PER2, а также тремя другими генами, связанными с циркадианной регуляцией (APH1A, RGS16 и FBXL13). Обнаруженные 12 локусов (rs141175086, rs2050122, rs76681500, rs10157197, rs1144566, rs11895698, rs11708779, rs148750727, rs372229746, rs17311976, rs542675489, rs4821940) объясняли 4,3% вариабельности хронотипа. Всего в этих локусах располагается 72 гена-кандидата, среди которых преобладают гены циркадианной регуляции, связанные с психическими расстройствами, нарушениями сна, сплайсосомой и болезнью Альцгеймера. В целом в данной выборке вечерний тип ассоциировался с более молодым возрастом, мужским полом, более длительным сном и большим риском депрессии. Это исследование частично воспроизвело результаты описанной выше работы (воспроизвелись восемь из 15 локусов, и все 15 демонстрировали влияние в одинаковом направлении), что увеличивает достоверность полученных ассоциаций. При совместном анализе обнаружились еще три значимых локуса около генов PER3, VIP и TOX [7]. Для той же когорты, но расширенной выборки в 128 266 человек был проведен еще один полногеномный анализ. Было обнаружено 16 локусов, ассоциированных с хронотипом. Полученные ассоциации проверяли и валидировали на двух других выборках. Оказалось, что варианты расположены поблизости генов, для которых уже была установлена ассоциация с фоторецепцией и циркадианными ритмами. Наиболее сильные ассоциации с rs516134 возле гена RGS16, связанного с циркадианной регуляцией, и rs75804782 возле PER2 [12].
В другом исследовании (n = 749) оценивались сонливость, время отхода ко сну и длительность сна. Единственный расположенный в кодирующей области полиморфизм гена рецептора 1 нейропептида S NPSR1 rs324981 в минорном варианте проявляется в более позднем времени отхода ко сну. Это может объясняться ролью нейропептида в регуляции сна и бодрствования. Минорный аллель приводит к замене аминокислоты в рецепторе и повышает чувствительность к нейропептиду S [8]. На животных моделях введение нейропептида S снижает количество сна и стимулирует бодрствование [13].
Полногеномные исследования позволяют охватить огромный диапазон генов, но механизм обнаруженных связей не всегда удается объяснить, как видно на примере приведенных работ. Поиск имеющих фенотипические проявления мутаций в системе биологических часов и ее регуляторов дает возможность более эффективно устанавливать механизмы действия полиморфизмов. Пожалуй, наибольшее внимание исследователей генетических основ хронотипа сосредоточено на одном из генов биологических часов – PER3. Сначала была обнаружена связь двух миссенс-мутаций (G647 и R1158) этого гена с синдромом задержки фазы сна [14]. Последующие работы выявили полиморфизм, для которого получены наиболее полные сведения о влиянии на хронотип. В кодирующей области гена PER3 содержится локус с VNTR. В нем последовательность, кодирующая 18 аминокислот, может повторяться четыре (PER34) или пять (PER35) раз. Данный полиморфизм связан с общими диурнальными предпочтениями. Примерно 10% популяции – гомозиготы по длинному аллелю [15]. В европейской и японской популяциях длинный аллель связывают с утренним типом, а короткий ‒ с вечерним [14, 16]. Неоднозначны ассоциации с нарушениями циркадианной ритмики. В проведенном в Лондоне исследовании короткий аллель ассоциировался с вечерним типом и синдромом задержки фазы сна [16]. В бразильской когорте в Сан-Паолу был получен иной результат: длинный аллель ассоциировался с синдромом задержки фазы сна, возможно, из-за разницы в широте городов и различий в ответе на сезонные колебания [17]. Однако этот полиморфизм не оказывает влияния на чувствительность к хроническому ограничению сна: наблюдаемые нейроповеденческие изменения одинаковы для обоих аллелей [18, 19]. В наибольшей степени связь между VNTR и диурнальными предпочтениями проявляется у молодых людей 18–29 лет, с возрастом эта ассоциация ослабевает [20].
Вклад вариабельности генов молекулярных часов, в особенности регуляции ритмов сна и бодрствования, не ограничивается описанным VNTR. В колумбийском исследовании полиморфизм rs2640909 гена PER3 оказался связан с подшкалой «утренний тип» Комплексного опросника утреннего типа. Кроме того, подшкала «утренняя активация» была связана с полиморфизмом другого часового гена – PER2 rs934945 [21]. Одну из связей удалось подтвердить и на российской выборке: с полиморфизмом rs2640909 ассоциирован параметр утренней активации [22]. Другой ОНП PER3, rs228697, оказался ассоциирован с хронотипом и несинхронизированным типом [23]. На хронотип влияют и другие полиморфизмы генов биологических часов: ОНП PER3 (rs10462020), ARNTL2 (rs922270), а также в меньшей степени редкие варианты в FBXL3 (825679097) и GNβ3 (rs5443) [24]. Аллель гена CRY1 c.1657+3A>C, приводящий у его продукта к усиленному ингибированию транскрипции, удлиняет период биологических часов до 24,5 часа с частотой до 0,6%. На поведенческом уровне это проявляется как наследуемый по доминантному типу синдром задержки фазы сна [25].
Многие обнаруженные в настоящее время мутации укорачивают период молекулярных часов и сдвигают фазы сна – бодрствования вперед. При этом затрагиваются механизмы регуляции стабильности и активности белков-компонентов биологических часов [26–28]. С синдромом семейного опережения фазы сна связывают приводящие к снижению стабильности или ускорению деградации мутации в генах элементов биологических часов CRY2 (A260T), PER2 (S662G) и PER3 (P415A/H417R) и регулирующей активность транскрипции и стабильность продукта PER2 казеинкиназы 1-дельта CK1δ (T44A) [27, 29]. Этот же вариант PER3 ассоциирован с более высокими оценками депрессии и сезонного аффективного расстройства. В сочетании со сходными результатами исследований на животных можно предположить, что PER3 может быть узлом связи биологических часов и эмоциональной регуляции [29].
Исследование полиморфизмов генов-кандидатов, не имеющих отношения к системе биологических часов, также может дополнить картину генетических влияний на хронотип. Например, минорные аллели полиморфизмов генов NPSR1 (rs324981) и транспортера дофамина SLC6A3 (rs6347) связаны с более поздним хронотипом в российской выборке [30]. Эффект ОНП NPSR1 воспроизводит результат полногеномного исследования.
Роль мелатониновой регуляции
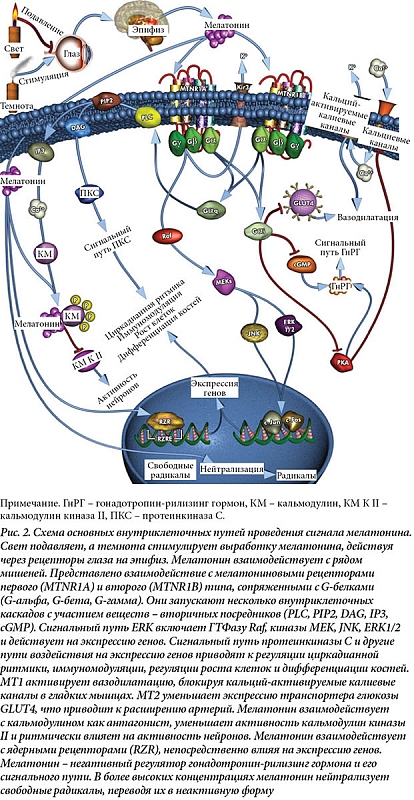
Мелатонин – гормон эпифиза, суточные колебания его концентрации хорошо известны и даже используются как маркер оценки циркадианных процессов. Его выработка находится под контролем биологических часов в СХЯ и освещенности (также через СХЯ). У млекопитающих днем уровень мелатонина низкий, но нарастает примерно за два часа до отхода ко сну и достигает максимальных значений ночью. Ближе к пробуждению начинается спад, который завершается после пробуждения. Свет способен блокировать ночную выработку мелатонина и нарушить обычный ритм (рис. 2) [31].
Мелатонин имеет широкий спектр мишеней как в мозге, так и в периферических тканях (см. рис. 2). Его эффекты зависят от преобладающих в тканях рецепторов. Наиболее изучены два связанных с G-белками рецептора: МТ1 и МТ2, которые активируют целый ряд внутриклеточных сигнальных каскадов. Активация МТ1 приводит к снижению уровня продукции клеточного мессенджера циклического аденозинмонофосфата и активности протеинкиназ. В результате меняется активность транскрипционного фактора CREB. Кроме того, следствием этого становятся активация фосфолипазы и изменение направления ионных токов через мембрану клетки. Активация МТ2 снижает уровни циклического аденозинмонофосфата и гуанозинмонофосфата, действует на протеинкиназу, влияющую на ионные токи. Экспрессия рецепторов мелатонина и степень его связывания с ними также демонстрируют суточные колебания. Мелатонин в больших концентрациях способен уменьшать число своих рецепторов [32, 33].
Связывание с мелатониновыми рецепторами – не единственный путь действия мелатонина, который непосредственно связывается и с внутриклеточными белками, например кальмодулином, кальретикулином и тубулином. Мелатонин препятствует связыванию кальмодулина с кальцием, нарушая кальмодулиновый путь активации транскрипции. Кроме того, он способен действовать на ретиноидные орфанные рецепторы (RZR/ROR). Эти пути обеспечивают его антипролиферативные и иммуномодулирующие свойства. Еще один рецептор мелатонина, иногда обозначаемый МТ3, – фермент хинон редуктаза 2 обеспечивает защиту от окислительного стресса в периферических тканях [34].
Рецепторы МТ2 играют роль в регуляции сна: у мышей без этих рецепторов снижается доля медленноволнового сна, мощность дельта- и сигма-ритмов электроэнцефалограммы (ЭЭГ) сна. Они также связаны с патофизиологией тревожных расстройств, депрессии, болезни Альцгеймера [35]. Мелатониновый ритм находится в петле обратной связи с биологическими часами. Действуя через MT1, мелатонин уменьшает активность нейронов СХЯ, в особенности днем. Мелатонин и его агонисты могут сдвигать фазу биологических часов. Направление сдвига зависит от времени приема относительно фазы циркадианного ритма (так называемые фазовые кривые ответа). В целом физиологические эффекты мелатонина могут быть сцеплены с фазой суточного цикла. Мелатонин или синтетические агонисты МТ1 и МТ2 могут стабилизировать убегающие биологические ритмы и нормализовать нарушения циркадианной ритмики. В настоящее время мелатонин и агонисты МТ1 и МТ2 используются при нарушениях сна и циркадианных расстройствах. Это перспективные препараты для лечения заболеваний, связанных и с иными функциями мелатонина [33, 36]. Генетические исследования пока не подтверждают ассоциации между генами, ассоциированными с мелатониновой регуляцией и параметрами сна или хронотипа. Однако мелатонин, помимо прочих функций, – важный элемент регуляции и интеграции своей системы суточных осцилляций, и обнаружение таких связей возможно в дальнейшем.
В настоящее время свойство синтетических препаратов мелатонина влиять на циркадианный компонент системы регуляции сна используется для лечения инсомнии и различных форм нарушений биологических ритмов. В рандомизированном плацебоконтролируемом исследовании эффективности препарата мелатонина с замедленным высвобождением Циркадин (в России распространяется фирмой «Бофур Ипсен») продемонстрировано, что при первичной инсомнии прием препарата приводил к уменьшению на 50% времени засыпания и на 50% времени бодрствования в период сна. Эти эффекты достоверно превышали эффекты плацебо [37]. В другом исследовании, основанном на субъективной оценке, было показано улучшение как качества сна, так и утреннего бодрствования при применении Циркадина по сравнению с плацебо [38]. В третьем исследовании на фоне длительной терапии Циркадином (в течение шести месяцев) первичный положительный эффект в виде сокращения времени засыпания и улучшения качества сна по сравнению с плацебо сохранялся и через три месяца лечения, при этом не отмечено негативных побочных эффектов, в том числе привыкания [39]. Наибольший эффект препарата Циркадин достигался при меньшем уровне секреции собственного мелатонина и у людей старше 55 лет, что позволило Британской ассоциации психофармакологии в 2010 г. рекомендовать препарат мелатонина с пролонгированным высвобождением для лечения больных инсомнией в этой возрастной группе [40].
Препараты мелатонина – единственные лекарственные средства, которые применяются в лечении расстройств суточного ритма сна – бодрствования. Согласно рекомендациям Американской академии медицины сна по лечению этих расстройств, опубликованным в 2015 г., мелатонин рекомендован при синдроме задержки фазы сна (как у взрослых, так и у детей), свободно выбранном ритме сна – бодрствования у слепых людей и нерегулярном цикле у детей с неврологическими расстройствами [41]. Что касается именно Циркадина, то положительный эффект этого препарата мелатонина при расстройствах цикла «сон – бодрствование» был подтвержден в исследованиях с участием детей с неврологическими расстройствами и слепых людей [42, 43].
Циркадин назначается в неделимой таблетке в дозе 2 мг после приема пищи за один-два часа перед сном. Противопоказан при врожденной непереносимости галактозы, синдроме глюкозо-галактозной мальабсорбции, врожденном дефиците лактазы, аутоиммунных заболеваниях, печеночной недостаточности, детском и подростковом возрасте до 18 лет. Курс лечения может составлять до 13 недель [44].
Длительность и качество сна
Предпочтительная длительность, а также типичная структура и глубина сна – индивидуальные черты, частично находящиеся под влиянием наследственности. Полногеномные исследования на больших выборках указывают на возможные гены-кандидаты, варианты которых ассоциированы с параметрами длительности или качества сна. Исследования конкретных генетических вариантов или полиморфизмов позволяют воспроизвести эти ассоциации, обнаружить новые и работать с редкими нарушениями или вариантами нормы, для которых полногеномный подход невозможен.
Финское исследование, сочетавшее подход полногеномного анализа, оценки экспрессии и экспериментальной верификации, указывает на два возможных варианта: ОНП rs2031573 возле KLF6 и rs1037079 между PCDH7-CENTD1 [45]. Обнаружено, что аллели, связанные с более коротким сном, ассоциируются и с большей экспрессией этих генов, но механизм их участия в регуляции сна пока неясен. В британском полногеномном исследовании удалось воспроизвести обнаруженные в более ранних полногеномных исследованиях ассоциации ОНП гена кальциевого канала L-типа CACNA1C (rs16929277) с качеством сна и гена аденозинтрифосфатзависимого калиевого канала ABCC9 (rs11046209) с длительностью сна [46]. Ген ABCC9, а также и ранее связанные с длительностью сна (TSHZ2, FBXO15) и нарколепсией (NFATC2, SALL4), и новые гены SORCS1 и ELOVL2 оказались ассоциированы с длительностью сна в более позднем американском полногеномном исследовании [47]. К сожалению, для этих генетических вариантов точные молекулярные механизмы работы пока неизвестны.
В полногеномных исследованиях обнаруживаются и не ассоциированные напрямую с каким-либо геном варианты, которые тем не менее связаны с длительностью сна. На базе 18 когорт европеоидов обнаружена и подтверждена на афроамериканской группе ассоциация длительности сна и вариаций в межгенной области во второй хромосоме перед геном PAX8 и особенно однонуклеотидным вариантом rs1823125. Причем носители минорного варианта спали дольше, имели более безопасный метаболический профиль и меньший риск синдрома гиперактивности с дефицитом внимания [48]. В полногеномном исследовании 750 человек было обнаружено четыре области, связанных с временем отхода ко сну, и одна – с длительностью сна, хотя эти ассоциации не были ярко выражены. Наиболее отчетливая связь времени отхода ко сну и области на 16-й хромосоме обнаружена рядом с геном казеинкиназы 2-альфа-2 (CSNK2A2) – элементом биологических часов у дрозофилы. Пик связи длительности сна включает область на третьей хромосоме возле гена прокинетицина 2 (PROK2), который, предположительно, проводит сигнал от СХЯ [8].
Исследования влияния полиморфизмов генов-кандидатов также указывают на влияние наследственности на параметры сна. Например, нейромодулятор аденозин тесно связан с регуляцией гомеостаза сна. Количественный генетический анализ животных моделей и здоровых людей показал, что ряд мутаций генов аденозинергической системы может не приводить к значительным нарушениям, но влиять на параметры сна. Функциональный полиморфизм гена, регулирующего уровень аденозина фермента аденозиндезаминазы ADA 22 G-A (минорный вариант А приводит к снижению активности фермента), проявляется неодинаковой чувствительностью к кофеину, а также различиями в архитектуре и ЭЭГ сна. Носители G/A-генотипа реже просыпались ночью, имели более глубокий и длительный медленноволновой сон, более выраженные дельта- и тета-ритмы в медленноволновом сне. Для полиморфизма гена аденозинового рецептора А2А ADORA2A 1976T-C функциональное проявление не было ограничено сном: у носителей С/С-генотипа мощность ЭЭГ в диапазоне 7,5–10 Гц больше, чем у носителей Т/Т-генотипа, и во сне, и в бодрствовании [49, 50].
ОНП гена NPSR1 rs324981 связан не только с хронотипом, но и с качеством сна. В актиграфическом исследовании параметров сна минорный аллель ассоциировался с меньшей длительностью сна и отдыха, но не временем начала отдыха или засыпанием [51]. Кроме того, он был связан с худшим субъективным качеством сна у мужчин [52]. Полученный результат не полностью сочетается с обнаруженной связью с поздним хронотипом, и данный ОНП требует дальнейшего изучения.
Полиморфизмы генов биологических часов могут влиять не только на хронотип, но и на параметры сна. ОНП rs5443 гена GNβ3 влиял на общую оценку сна по Питтсбургской шкале качества сна, ОНП FBXL3 (825679097) – на качество сна, а редкий полиморфизм гена PER2 (rs2304672) – как на качество, так и на общую продолжительность сна [24].
В популяции можно выделить подгруппу людей, для которых естествен короткий сон. Они спят значительно меньше средних значений, но при этом здоровы и не страдают от недостатка сна. Уже обнаружены мутации, ассоциированные с семейными формами такого фенотипа. Они найдены для гена basic helix-loop-helix family member e41 (BHLHE41 или DEC2), который относится к регуляторам элементов молекулярных часов CLOCK/BMAL1 и NPAS2/BMAL1. В двух различных когортах замены Y362H и P385R в гене DEC2 приводили к снижению активности белка и появлению малоспящего фенотипа [53, 54].
Один из недостатков многих исследований – использование субъективных опросных данных для получения таких параметров, как время отхода ко сну, длительность и качество сна. Психологическое состояние, образ жизни, возраст и пол, принимаемые препараты и даже время года могут влиять на сон. Эти факторы сложно проконтролировать при определении фенотипа. Обычно исследования генетических ассоциаций только указывают на возможную связь какого-то нарушения или параметра нормальной вариабельности, на самом деле определяющими будут средовые факторы или особенности выборки. Особую ценность приобретают генетические исследования, в которых помимо субъективной оценки используются и другие методы фенотипирования: актиграфия, регистрация полисомнограммы. Важны воспроизведение результатов в независимых исследованиях и поиск механизмов действия мутации. Проверка на животных моделях – эффективный подход подтверждения полученных результатов.
Человеческая популяция генетически значительно разнообразнее, чем модельные животные, и это создает дополнительный уровень сложности. Исследования последних лет в основном фокусируются на независимых влияниях отдельных полиморфизмов. Не для всех обнаруженных ассоциаций даже известен точно тип наследования фенотипа (доминантный или рецессивный). Если рассматриваются моногенные влияния, то следует помнить, что мы наблюдаем их эффект на фоне разнообразной наследственности. Например, люди с нарушениями циркадианной ритмики могут рассматриваться как крайние варианты распределения в популяции, их особенность может быть обусловлена одной мутацией или суммарным воздействием множества полиморфизмов. Вопросы взаимодействия генов крайне сложны и требуют еще более масштабных исследований, чем те, что проводятся сейчас.
Работа выполнена при поддержке Российского фонда фундаментальных исследований № 16-06-01054 /17-ОГОН.
A.N. Puchkova
Institute of Higher Nervous Activity and Neurophysiology of RAS, Moscow
Pushkin State Russian Language Institute, Moscow
Contact person: Aleksandra Nikolayevna Puchkova, puchkovaan@gmail.com
In the recent decades, many studies have been dedicated to factors that affect the individual variability of sleep-wake rhythms and sleep characteristics. Genetic research has made a significant contribution to this field, since many chronobiological features and sleep parameters reveal a hereditary component. This review covers recent genetic studies of the chronotype and sleep of the parameters in humans. The focus of the article is on normal variability of sleep and chronotype parameters, as well as on circadian rhythm disorders. Particular attention is paid to the biological clock genes and the effects of their variability in the population. In addition, the article highlights the main approaches in genetic research on humans and points to the role of melatonin in the regulation of circadian rhythm.
Уважаемый посетитель uMEDp!
Уведомляем Вас о том, что здесь содержится информация, предназначенная исключительно для специалистов здравоохранения.
Если Вы не являетесь специалистом здравоохранения, администрация не несет ответственности за возможные отрицательные последствия, возникшие в результате самостоятельного использования Вами информации с портала без предварительной консультации с врачом.
Нажимая на кнопку «Войти», Вы подтверждаете, что являетесь врачом или студентом медицинского вуза.