–Э–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–µ –і–µ–є—Б—В–≤–Є–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞: —А–Њ–ї—М –≤¬†–њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–µ –≥–Є–њ–Њ–Ї—Б–Є—З–µ—Б–Ї–Є—Е –љ–∞—А—Г—И–µ–љ–Є–є –≤–љ—Г—В—А–Є—Г—В—А–Њ–±–љ–Њ–≥–Њ —А–∞–Ј–≤–Є—В–Є—П –≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞
- –Р–љ–љ–Њ—В–∞—Ж–Є—П
- –°—В–∞—В—М—П
- –°—Б—Л–ї–Ї–Є
- English






–Т–≤–µ–і–µ–љ–Є–µ
–Э–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—Ж–Є—П –Љ–Њ–Ј–≥–∞ –њ–ї–Њ–і–∞ —З—А–µ–Ј–≤—Л—З–∞–є–љ–Њ –≤–∞–ґ–љ–∞ –Ї–∞–Ї –љ–∞¬†—А–∞–љ–љ–Є—Е —Б—А–Њ–Ї–∞—Е –±–µ—А–µ–Љ–µ–љ–љ–Њ—Б—В–Є –і–ї—П –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є –≤—А–Њ–ґ–і–µ–љ–љ—Л—Е –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П –Љ–Њ–Ј–≥–∞, —В–∞–Ї –Є¬†–≤¬†—В–µ—З–µ–љ–Є–µ –≤—Б–µ–є¬†–≥–µ—Б—В–∞—Ж–Є–Є (–Њ—Б–Њ–±–µ–љ–љ–Њ –≤¬†–њ–Њ—Б–ї–µ–і–љ–µ–Љ —В—А–Є–Љ–µ—Б—В—А–µ) –і–ї—П –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є –Є—И–µ–Љ–Є–Є¬†–≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ –њ–ї–Њ–і–∞.
–Р–љ–Њ–Љ–∞–ї–Є–Є —А–∞–Ј–≤–Є—В–Є—П —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –њ–ї–Њ–і–∞ –≤–Њ–Ј–љ–Є–Ї–∞—О—В –≤—Б–ї–µ–і—Б—В–≤–Є–µ –љ–∞—А—Г—И–µ–љ–Є–є –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ —А–∞–Ј–Љ–љ–Њ–ґ–µ–љ–Є—П, –Љ–Є–≥—А–∞—Ж–Є–Є, –і–Є—Д—Д–µ—А–µ–љ—Ж–Є–∞—Ж–Є–Є –Є¬†–њ—А–Њ–≥—А–∞–Љ–Љ–Є—А–Њ–≤–∞–љ–љ–Њ–є¬†–≥–Є–±–µ–ї–Є –Ї–ї–µ—В–Њ–Ї –≤–Њ –≤—А–µ–Љ—П —А–Њ—Б—В–∞ —Н–Љ–±—А–Є–Њ–љ–∞. –Ч–љ–∞—З–Є–Љ—Л–є —Д–∞–Ї—В–Њ—А —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—П –≤—А–Њ–ґ–і–µ–љ–љ—Л—Е –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П –Љ–Њ–Ј–≥–∞¬†вАУ —Б–∞—Е–∞—А–љ—Л–є –і–Є–∞–±–µ—В, –Њ—Б–ї–Њ–ґ–љ—П—О—Й–Є–є —В–µ—З–µ–љ–Є–µ –±–µ—А–µ–Љ–µ–љ–љ–Њ—Б—В–Є –љ–µ¬†–Љ–µ–љ–µ–µ —З–µ–Љ —Г¬†2,4% –ґ–µ–љ—Й–Є–љ¬†[1].
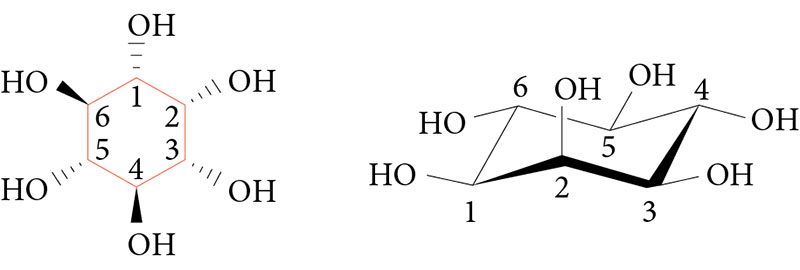
–Я—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–∞ –≤—А–Њ–ґ–і–µ–љ–љ—Л—Е –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П –Љ–Њ–Ј–≥–∞ —Д–Њ–ї–∞—В–∞–Љ–Є –љ–µ—Н—Д—Д–µ–Ї—В–Є–≤–љ–∞ –њ—А–Є —В–∞–Ї –љ–∞–Ј—Л–≤–∞–µ–Љ—Л—Е —Д–Њ–ї–∞—В—А–µ–Ј–Є—Б—В–µ–љ—В–љ—Л—Е –њ–Њ—А–Њ–Ї–∞—Е —А–∞–Ј–≤–Є—В–Є—П, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г —Д–Њ–ї–∞—В—Л¬†вАУ –і–∞–ї–µ–Ї–Њ –љ–µ¬†–µ–і–Є–љ—Б—В–≤–µ–љ–љ—Л–є –Љ–Є–Ї—А–Њ–љ—Г—В—А–Є–µ–љ—В, –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л–є –і–ї—П —А–∞–Ј–≤–Є—В–Є—П —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –њ–ї–Њ–і–∞. –Т¬†—З–Є—Б–ї–Њ¬†–≥–ї–∞–≤–љ–µ–є—И–Є—Е –љ–µ–є—А–Њ–∞–Ї—В–Є–≤–љ—Л—Е –Љ–Є–Ї—А–Њ–љ—Г—В—А–Є–µ–љ—В–Њ–≤, –њ—А–Є–љ–Є–Љ–∞—О—Й–Є—Е –Ї–Њ–Љ–њ–ї–µ–Ї—Б–љ–Њ–µ —Г—З–∞—Б—В–Є–µ –≤¬†—Н–Љ–±—А–Є–Њ–≥–µ–љ–µ–Ј–µ –Є¬†—А–∞–Ј–≤–Є—В–Є–Є –Љ–Њ–Ј–≥–∞ –њ–ї–Њ–і–∞, –≤—Е–Њ–і–Є—В –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї (–≤–Є—В–∞–Љ–Є–љ –Т8), —Б–њ–µ—Ж–Є—Д–Є—З–µ—Б–Ї–∞—П —А–∞–Ј–љ–Њ–≤–Є–і–љ–Њ—Б—В—М —И–µ—Б—В–Є–∞—В–Њ–Љ–љ—Л—Е —Б–њ–Є—А—В–Њ–≤¬†вАУ –Є–љ–Њ–Ј–Є—В–Њ–ї–Њ–≤. –Ш–љ–Њ–Ј–Є—В–Њ–ї—Л (—Ж–Є–Ї–ї–Њ–≥–µ–Ї—Б–∞–љ-1,2,3,4,5,6-–≥–µ–Ї—Б–Њ–ї—Л) –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л –і–µ–≤—П—В—М—О —Б—В–µ—А–µ–Њ–Є–Ј–Њ–Љ–µ—А–∞–Љ–Є, –Є–Ј¬†–Ї–Њ—В–Њ—А—Л—Е –Є–Љ–µ–љ–љ–Њ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї –Є–Љ–µ–µ—В –њ—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ–Њ–µ –Ј–љ–∞—З–µ–љ–Є–µ –і–ї—П —Д—Г–љ–Ї—Ж–Є–Њ–љ–Є—А–Њ–≤–∞–љ–Є—П –≤—Б–µ—Е —В–Є–њ–Њ–≤ –Ї–ї–µ—В–Њ–Ї (—А–Є—Б.¬†1). –Ь–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї –Є¬†–µ–≥–Њ —Д–Њ—Б—Д–∞—В–љ—Л–µ –њ—А–Њ–Є–Ј–≤–Њ–і–љ—Л–µ (–Є–љ–Њ–Ј–Є—В–Њ–ї—Д–Њ—Б—Д–∞—В—Л, —Д–Њ—Б—Д–∞—В–Є–і–Є–ї–Є–љ–Њ–Ј–Є—В–Њ–ї–Њ–≤—Л–µ –ї–Є–њ–Є–і—Л) –≤—Л—Б—В—Г–њ–∞—О—В –≤¬†–Ї–∞—З–µ—Б—В–≤–µ –≤–∞–ґ–љ—Л—Е –њ–µ—А–µ–і–∞—В—З–Є–Ї–Њ–≤ —Б–Є–≥–љ–∞–ї–∞ –≤–Њ –≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ—Л—Е —Б–Є–≥–љ–∞–ї—М–љ—Л—Е –Ї–∞—Б–Ї–∞–і–∞—Е.
–Т¬†—А–µ—Д–µ—А–Є—А—Г–µ–Љ—Л—Е –љ–∞—Г—З–љ—Л—Е –ґ—Г—А–љ–∞–ї–∞—Е –Њ–њ—Г–±–ї–Є–Ї–Њ–≤–∞–љ–Њ –±–Њ–ї–µ–µ 40¬†—В—Л—Б. —А–∞–±–Њ—В –Њ¬†–Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ–Њ-—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Љ–µ—Е–∞–љ–Є–Ј–Љ–∞—Е –і–µ–є—Б—В–≤–Є—П –Љ–Є–Њ¬≠–Є–љ–Њ–Ј–Є—В–Њ–ї–∞, –≤–Ї–ї—О—З–∞—О—Й–Є—Е —В–∞–Ї–ґ–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є. –Т–µ—Б—М —Н—В–Њ—В –Љ–∞—Б—Б–Є–≤ –њ—Г–±–ї–Є–Ї–∞—Ж–Є–є –њ–Њ¬†–Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї—Г –њ–Њ—Б—А–µ–і—Б—В–≤–Њ–Љ —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л—Е –Љ–µ—В–Њ–і–Њ–≤ –Є–љ—В–µ–ї–ї–µ–Ї—В—Г–∞–ї—М–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –і–∞–љ–љ—Л—Е –њ—А–Њ–∞–љ–∞–ї–Є–Ј–Є—А–Њ–≤–∞–ї–Є –Ю.–Р. –У—А–Њ–Љ–Њ–≤–∞ –Є¬†—Б–Њ–∞–≤—В. [2]. –С—Л–ї–Њ —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ –њ—А–Њ–Є–Ј–≤–Њ–і–љ—Л–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ —Г—З–∞—Б—В–≤—Г—О—В –≤¬†–њ–µ—А–µ–і–∞—З–µ —Б–Є–≥–љ–∞–ї–Њ–≤ –Њ—В¬†—А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ —А–Њ—Б—В–Њ–≤—Л—Е —Д–∞–Ї—В–Њ—А–Њ–≤ –Є¬†—А–µ—Ж–µ–њ—В–Њ—А–∞ –Є–љ—Б—Г–ї–Є–љ–∞ [3], —А–∞—Б—Й–µ–њ–ї–µ–љ–Є–Є –ґ–Є—А–Њ–≤ –Є¬†—Б–љ–Є–ґ–µ–љ–Є–Є —Г—А–Њ–≤–љ—П —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –≤¬†–Ї—А–Њ–≤–Є [4], –Љ–Њ–і—Г–ї—П—Ж–Є–Є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –љ–µ–є—А–Њ—В—А–∞–љ—Б–Љ–Є—В—В–µ—А–Њ–≤ [5] –Є¬†–і—А.
–Р–љ–∞–ї–Є–Ј 120 –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–Ј–∞–≤–Є—Б–Є–Љ—Л—Е –±–µ–ї–Ї–Њ–≤ –њ—А–Њ—В–µ–Њ–Љ–∞ —З–µ–ї–Њ–≤–µ–Ї–∞ –њ–Њ–Ї–∞–Ј–∞–ї, —З—В–Њ –±–Њ–ї–µ–µ –њ–Њ–ї–Њ–≤–Є–љ—Л –Є–Ј¬†–љ–Є—Е –≤–Њ–≤–ї–µ—З–µ–љ—Л –≤¬†–њ–Њ–і–і–µ—А–ґ–Ї—Г –ґ–Є–Ј–љ–µ–і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є —Б–µ—А–і–µ—З–љ–Њ-—Б–Њ—Б—Г–і–Є—Б—В–Њ–є —Б–Є—Б—В–µ–Љ—Л, –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞ –Є¬†—Б—В—А—Г–Ї—В—Г—А—Л —Б–Њ–µ–і–Є–љ–Є—В–µ–ї—М–љ–Њ–є —В–Ї–∞–љ–Є (–≤¬†—В–Њ–Љ —З–Є—Б–ї–µ —Б–Њ—Б—В–Њ—П–љ–Є—П –Ї–Њ—Б—В–µ–є, —Е—А—П—Й–∞, –Ї–Њ–ґ–Є –Є¬†–њ—А–Њ—Ж–µ—Б—Б–Њ–≤ –Ј–∞–ґ–Є–≤–ї–µ–љ–Є—П —А–∞–љ). –Э–µ¬†–Љ–µ–љ–µ–µ –≤–∞–ґ–љ–Њ —Г—З–∞—Б—В–Є–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –≤¬†–Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–µ —Б–∞—Е–∞—А–Њ–≤ (–њ—А–µ–ґ–і–µ –≤—Б–µ–≥–Њ —Б–Є–≥–љ–∞–ї—М–љ–Њ–Љ –Ї–∞—Б–Ї–∞–і–µ –Є–љ—Б—Г–ї–Є–љ–∞) –Є¬†–Њ–±–µ—Б–њ–µ—З–µ–љ–Є–Є —Д—Г–љ–Ї—Ж–Є–Њ–љ–Є—А–Њ–≤–∞–љ–Є—П —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л (–≤–Ї–ї—О—З–∞—П –љ–µ–є—А–Њ—В—А–Њ—Д–Є—З–µ—Б–Ї–Є–є –Є¬†–љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–є —Н—Д—Д–µ–Ї—В) (—А–Є—Б.¬†2) [2].
–Р–љ–Њ–Љ–∞–ї–Є–Є –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ—Л —Б¬†–Ї–Њ–≥–љ–Є—В–Є–≤–љ—Л–Љ–Є –љ–∞—А—Г—И–µ–љ–Є—П–Љ–Є [6], –і–µ–њ—А–µ—Б—Б–Є–µ–є [7], –і–Є–∞–±–µ—В–Є—З–µ—Б–Ї–Њ–є –љ–µ–≤—А–Њ–њ–∞—В–Є–µ–є [8] –Є¬†–і—А. –§—Г–љ–і–∞–Љ–µ–љ—В–∞–ї—М–љ—Л–µ –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї –љ–µ–Њ–±—Е–Њ–і–Є–Љ –і–ї—П –Њ–±–µ—Б–њ–µ—З–µ–љ–Є—П –љ–µ–є—А–Њ–љ–∞–ї—М–љ–Њ–є —Д—Г–љ–Ї—Ж–Є–Є, –≤–Ї–ї—О—З–∞—П —Б–Є–љ–∞–њ—В–Є—З–µ—Б–Ї—Г—О –њ–µ—А–µ–і–∞—З—Г –Є¬†—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ —Н—Д—Д–µ–Ї—В—Л —В–∞–Ї–Є—Е –љ–µ–є—А–Њ—В—А–∞–љ—Б–Љ–Є—В—В–µ—А–Њ–≤, –Ї–∞–Ї —Б–µ—А–Њ—В–Њ–љ–Є–љ, –і–Њ—Д–∞–Љ–Є–љ,¬†–≥–∞–Љ–Љ–∞-–∞–Љ–Є–љ–Њ–Љ–∞—Б–ї—П–љ–∞—П –Ї–Є—Б–ї–Њ—В–∞, –љ–µ–є—А–Њ–Љ–µ–і–Є–љ. –Ь–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї –љ—Г–ґ–µ–љ –і–ї—П –љ–µ–є—А–Њ–≥–µ–љ–µ–Ј–∞ (–Њ–Ї–∞–Ј—Л–≤–∞–µ—В –љ–µ–є—А–Њ—В—А–Њ—Д–Є—З–µ—Б–Ї–Є–є —Н—Д—Д–µ–Ї—В), –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—Ж–Є–Є (–≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –Ј–∞—Й–Є—В—Л –Ї–ї–µ—В–Њ–Ї —Б–µ—В—З–∞—В–Ї–Є¬†–≥–ї–∞–Ј–∞), –Њ—Б—Г—Й–µ—Б—В–≤–ї–µ–љ–Є—П –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ –Ј—А–µ–љ–Є—П, —Б–ї—Г—Е–∞, –≤–Ї—Г—Б–∞ –Є¬†–і–Њ–ї–≥–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–є –њ–Њ—В–µ–љ—Ж–Є–∞—Ж–Є–Є –≤¬†–≥–Є–њ–њ–Њ–Ї–∞–Љ–њ–µ (–њ–Њ–і–і–µ—А–ґ–Ї–∞ –њ–∞–Љ—П—В–Є).
–°—В–Њ–ї—М —И–Є—А–Њ–Ї–Є–є —Б–њ–µ–Ї—В—А –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –∞–Ї—В–Є–≤–љ–Њ—Б—В–µ–є –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –њ–Њ–Ј–≤–Њ–ї—П–µ—В –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ –Њ–љ –Љ–Њ–ґ–µ—В –±–Њ–ї–µ–µ —Б–њ–µ—Ж–Є—Д–Є—З–µ—Б–Ї–Є –≤–Њ–Ј–і–µ–є—Б—В–≤–Њ–≤–∞—В—М –љ–∞¬†—Б–Є–≥–љ–∞–ї—М–љ—Л–µ –Ї–∞—Б–Ї–∞–і—Л –≤—Л–ґ–Є–≤–∞–љ–Є—П –љ–µ–є—А–Њ–љ–Њ–≤ –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е —Б—В—А–µ—Б—Б–∞ (–≥–Є–њ–Њ–Ї—Б–Є–Є, –љ–µ–є—А–Њ—В–Њ–Ї—Б–Є—З–љ–Њ—Б—В–Є¬†–≥–ї—Г—В–∞–Љ–∞—В–∞, —Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–Њ–Љ –і–µ—Д–Є—Ж–Є—В–µ –Є¬†–≥–Є–њ–Њ–≥–ї–Є–Ї–µ–Љ–Є–Є, –і–Є—Б—Д—Г–љ–Ї—Ж–Є–Є –Љ–Є—В–Њ—Е–Њ–љ–і—А–Є–є, –Є–Ј–±—Л—В–Њ—З–љ–Њ–Љ –≤–Њ—Б–њ–∞–ї–µ–љ–Є–Є –Є¬†–і—А.). –Т¬†–љ–∞—Б—В–Њ—П—Й–µ–є —А–∞–±–Њ—В–µ –њ—А–Њ–≤–µ–і–µ–љ–∞ –≤–∞–ї–Є–і–∞—Ж–Є—П –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–≥–Њ –і–µ–є—Б—В–≤–Є—П –Љ–Є–Њ¬≠–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –Љ–µ—В–Њ–і–∞–Љ–Є –љ–µ–є—А–Њ—Ж–Є—В–Њ–ї–Њ–≥–Є–Є, –Є–Ј—Г—З–∞—О—Й–Є–Љ–Є –≤–ї–Є—П–љ–Є–µ –≤–µ—Й–µ—Б—В–≤ –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –љ–∞¬†–љ–µ–є—А–Њ–љ—Л [9вАУ11].
–Э–µ–є—А–Њ—Ж–Є—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –і–∞—О—В –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М —Г—Б—В–∞–љ–Њ–≤–Є—В—М –њ—А—П–Љ–Њ–є –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–є —Н—Д—Д–µ–Ї—В –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –њ—А–Є —А–∞–Ј–љ—Л—Е —Б—В—А–µ—Б—Б–Њ—А–љ—Л—Е –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П—Е. –Э–∞–њ—А–Є–Љ–µ—А, –њ—А–Є –Є—И–µ–Љ–Є–Є¬†–≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ —В–∞–Ї–Њ–≤—Л–Љ–Є —П–≤–ї—П—О—В—Б—П —Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–Є–є –і–µ—Д–Є—Ж–Є—В, –љ–µ–є—А–Њ—В–Њ–Ї—Б–Є—З–љ–Њ—Б—В—М¬†–≥–ї—Г—В–∞–Љ–∞—В–∞, –Њ–Ї–Є—Б–ї–Є—В–µ–ї—М–љ—Л–є —Б—В—А–µ—Б—Б, –і–Є—Б—Д—Г–љ–Ї—Ж–Є—П –Љ–Є—В–Њ—Е–Њ–љ–і—А–Є–є, –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є–є –∞—Ж–Є–і–Њ–Ј [12]. –С–ї–∞–≥–Њ–і–∞—А—П –љ–µ–є—А–Њ—Ж–Є—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П–Љ —В–∞–Ї–ґ–µ –Љ–Њ–ґ–љ–Њ –њ–Њ–Ї–∞–Ј–∞—В—М –≤–ї–Є—П–љ–Є–µ –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ—Л—Е —Б—А–µ–і—Б—В–≤ –љ–∞¬†–Ї–Њ–љ–Ї—А–µ—В–љ—Л–µ —Д–∞–Ї—В–Њ—А—Л —Б—В—А–µ—Б—Б–∞ –Є¬†–і–Њ–Ї–∞–Ј–∞—В—М –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ—Л–є —Н—Д—Д–µ–Ї—В –Є—Б—Б–ї–µ–і—Г–µ–Љ–Њ–≥–Њ –њ—А–µ–њ–∞—А–∞—В–∞ –≤¬†–Њ—В–љ–Њ—И–µ–љ–Є–Є –≤—Л–ґ–Є–≤–∞–љ–Є—П –Є–Љ–µ–љ–љ–Њ –љ–µ–є—А–Њ–љ–Њ–≤ (–∞¬†–љ–µ –і—А—Г–≥–Є—Е —В–Є–њ–Њ–≤ –Ї–ї–µ—В–Њ–Ї) [13, 14].
–Т¬†–љ–∞—Б—В–Њ—П—Й–µ–є —А–∞–±–Њ—В–µ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л —А–µ–Ј—Г–ї—М—В–∞—В—Л —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ–Њ–є –≤–∞–ї–Є–і–∞—Ж–Є–Є –њ—А—П–Љ–Њ–≥–Њ –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–≥–Њ –і–µ–є—Б—В–≤–Є—П –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –≤¬†—Б–Є–љ–µ—А–≥–Є—З–љ–Њ–є –Ї–Њ–Љ–±–Є–љ–∞—Ж–Є–Є —Б¬†—Д–Њ–ї–Є–µ–≤–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є (–њ—А–µ–њ–∞—А–∞—В –§–µ—А—В–Є–љ–∞¬†вАУ 1000¬†–Љ–≥ –Є–љ–Њ–Ј–Є—В–∞ (–Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞), 100¬†–Љ–Ї–≥ —Д–Њ–ї–Є–µ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –≤¬†–Њ–і–љ–Њ–Љ¬†—Б–∞—И–µ). –Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ—А–Њ–≤–Њ–і–Є–ї–Є—Б—М –љ–∞¬†–Ј–µ—А–љ–Є—Б—В—Л—Е –љ–µ–є—А–Њ–љ–∞—Е –Љ–Њ–Ј–ґ–µ—З–Ї–∞ –љ–Њ–≤–Њ—А–Њ–ґ–і–µ–љ–љ—Л—Е –Ї—А—Л—Б, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –≤¬†–Ї—Г–ї—М—В—Г—А–µ –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е¬†–≥–ї—Г—В–∞–Љ–∞—В–љ–Њ–≥–Њ —Б—В—А–µ—Б—Б–∞. –£–љ–Є–Ї–∞–ї—М–љ–Њ—Б—В—М —Н—В–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –Ј–∞–Ї–ї—О—З–∞–µ—В—Б—П –≤¬†—В–Њ–Љ, —З—В–Њ –Њ–љ–Є –њ–Њ–Ј–≤–Њ–ї—П—О—В –њ—А–Њ–і–µ–Љ–Њ–љ—Б—В—А–Є—А–Њ–≤–∞—В—М –њ—А—П–Љ–Њ–µ –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –Љ–Є–Њ¬≠–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –љ–∞¬†–љ–µ–є—А–Њ–љ—Л –Љ–Њ–Ј–≥–∞ –њ–ї–Њ–і–∞. –Т–Њ-–њ–µ—А–≤—Л—Е, –≤–Њ—Б–њ—А–Њ–Є–Ј–≤–µ–і–µ–љ–Є–µ¬†–≥–ї—Г—В–∞–Љ–∞—В–љ–Њ–≥–Њ —Б—В—А–µ—Б—Б–∞ —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є –∞–і–µ–Ї–≤–∞—В–љ–Њ –Љ–Њ–і–µ–ї–Є—А—Г–µ—В —Г—Б–ї–Њ–≤–Є—П —Г–Љ–µ—А–µ–љ–љ–Њ–є –Є—И–µ–Љ–Є–Є –Љ–Њ–Ј–≥–∞, –≤–Њ–Ј–љ–Є–Ї–∞—О—Й–µ–є –њ—А–Є –≤–љ—Г—В—А–Є—Г—В—А–Њ–±–љ–Њ–Љ —А–∞–Ј–≤–Є—В–Є–Є –њ–ї–Њ–і–∞. –Т–Њ-–≤—В–Њ—А—Л—Е, –Є–Ј—Г—З–µ–љ–Є–µ –≤–ї–Є—П–љ–Є—П –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –љ–∞¬†–љ–µ–є—А–Њ–љ—Л, –±–µ–Ј –њ—А–Њ—Е–Њ–ґ–і–µ–љ–Є—П —З–µ—А–µ–Ј —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–µ –Ї—А–Њ–≤–Њ–Њ–±—А–∞—Й–µ–љ–Є–µ, –њ–µ—З–µ–љ—М –Є¬†–і—А—Г–≥–Є–µ —Б–Є—Б—В–µ–Љ—Л –Њ—А–≥–∞–љ–Є–Ј–Љ–∞, –і–∞–µ—В –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –і–Њ–Ї–∞–Ј–∞—В—М, —З—В–Њ –Є–Љ–µ–љ–љ–Њ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї (–∞¬†–љ–µ –Ї–∞–Ї–Є–µ-—В–Њ –і—А—Г–≥–Є–µ –Є–Ј–Љ–µ–љ–µ–љ–Є—П –≤¬†–Њ—А–≥–∞–љ–Є–Ј–Љ–µ, –≤—Л–Ј–≤–∞–љ–љ—Л–µ –њ—А–Є–µ–Љ–Њ–Љ –њ—А–µ–њ–∞—А–∞—В–∞) –њ—А–Њ—П–≤–ї—П–µ—В –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–є —Н—Д—Д–µ–Ї—В.
–Ь–∞—В–µ—А–Є–∞–ї –Є¬†–Љ–µ—В–Њ–і—Л
–Т¬†—А–∞–±–Њ—В–µ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–Є—Б—М —Б–µ–Љ–Є-–≤–Њ—Б—М–Љ–Є—Б—Г—В–Њ—З–љ—Л–µ –Ї—Г–ї—М—В—Г—А—Л, –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ –Љ–µ—В–Њ–і–Њ–Љ —Д–µ—А–Љ–µ–љ—В–љ–Њ-–Љ–µ—Е–∞–љ–Є—З–µ—Б–Ї–Њ–є –і–Є—Б—Б–Њ—Ж–Є–∞—Ж–Є–Є –Ї–ї–µ—В–Њ–Ї –Љ–Њ–Ј–ґ–µ—З–Ї–∞ —Б–µ–Љ–Є–і–љ–µ–≤–љ—Л—Е –Ї—А—Л—Б –њ–Њ¬†–Њ–±—Й–µ–њ—А–Є–љ—П—В–Њ–є –Љ–µ—В–Њ–і–Є–Ї–µ. –Ц–Є–≤–Њ—В–љ—Л—Е —Г–Љ–µ—А—Й–≤–ї—П–ї–Є –ї–µ—В–∞–ї—М–љ–Њ–є –і–Њ–Ј–Њ–є —Н—Д–Є—А–љ–Њ–≥–Њ –љ–∞—А–Ї–Њ–Ј–∞, –њ–Њ—Б–ї–µ —З–µ–≥–Њ –њ—П—В—М –Љ–Є–љ—Г—В —Б—В–µ—А–Є–ї–Є–Ј–Њ–≤–∞–ї–Є 70%-–љ—Л–Љ —Б–њ–Є—А—В–Њ–Љ. –Ф–∞–ї–µ–µ –Є–Ј–≤–ї–µ–Ї–∞–ї–Є –Љ–Њ–Ј–ґ–µ—З–Њ–Ї –Є¬†–њ–µ—А–µ–љ–Њ—Б–Є–ї–Є –µ–≥–Њ –≤¬†–њ–ї–∞—Б—В–Є–Ї–Њ–≤—Г—О —З–∞—И–Ї—Г –Я–µ—В—А–Є, –Ї–Њ—В–Њ—А–∞—П –±—Л–ї–∞ –Ј–∞–њ–Њ–ї–љ–µ–љ–∞ —Д–Њ—Б—Д–∞—В–љ—Л–Љ –±—Г—Д–µ—А–Њ–Љ, –ї–Є—И–µ–љ–љ—Л–Љ –Є–Њ–љ–Њ–≤ –Ї–∞–ї—М—Ж–Є—П –Є¬†–Љ–∞–≥–љ–Є—П. –§—А–∞–≥–Љ–µ–љ—В—Л —В–Ї–∞–љ–Є –Є–љ–Ї—Г–±–Є—А–Њ–≤–∞–ї–Є 15 –Љ–Є–љ—Г—В –њ—А–Є 37¬†–Њ–° –≤¬†—Д–Њ—Б—Д–∞—В–љ–Њ–Љ –±—Г—Д–µ—А–µ, —Б–Њ–і–µ—А–ґ–∞—Й–µ–Љ 0,05% —В—А–Є–њ—Б–Є–љ–∞, 0,02% —Н—В–Є–ї–µ–љ–і–Є–∞–Љ–Є–љ—В–µ—В—А–∞—Г–Ї—Б—Г—Б–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Є¬†0,8%¬†–≥–ї—О–Ї–Њ–Ј—Л. –Я–Њ—Б–ї–µ –Є–љ–Ї—Г–±–∞—Ж–Є–Є —В–Ї–∞–љ—М –њ—А–Њ–Љ—Л–≤–∞–ї–Є –≤¬†–і–≤—Г—Е —Б–Љ–µ–љ–∞—Е —Д–Њ—Б—Д–∞—В–љ–Њ–≥–Њ –±—Г—Д–µ—А–∞ –Є¬†–Њ–і–Є–љ —А–∞–Ј –≤¬†—Б—А–µ–і–µ –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–Є—П, –Ј–∞—В–µ–Љ –њ–Њ–і–≤–µ—А–≥–∞–ї–Є –Љ–µ—Е–∞–љ–Є—З–µ—Б–Ї–Њ–є –і–Є—Б—Б–Њ—Ж–Є–∞—Ж–Є–Є –≤¬†–њ–Є—В–∞—В–µ–ї—М–љ–Њ–є —Б—А–µ–і–µ –і–ї—П –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–Є—П. –Т¬†—Б–Њ—Б—В–∞–≤ –њ–Є—В–∞—В–µ–ї—М–љ–Њ–є —Б—А–µ–і—Л –≤—Е–Њ–і–Є–ї–Є 90% –Љ–Є–љ–Є–Љ–∞–ї—М–љ–Њ–є —Б—А–µ–і—Л ¬Ђ–Ш–≥–ї–∞¬ї, 10% —Н–Љ–±—А–Є–Њ–љ–∞–ї—М–љ–Њ–є —В–µ–ї—П—З—М–µ–є —Б—Л–≤–Њ—А–Њ—В–Ї–Є, 2 –Љ–Ь¬†–≥–ї—Г—В–∞–Љ–Є–љ–∞, 5 –Љ–Ь KCl –Є¬†10 –Љ–Ь –±—Г—Д–µ—А–∞ –Э–Х–†–ХS, pH¬†7,2вАУ7,4. –°—Г—Б–њ–µ–љ–Ј–Є—О –Ї–ї–µ—В–Њ–Ї —Ж–µ–љ—В—А–Є—Д—Г–≥–Є—А–Њ–≤–∞–ї–Є –≤¬†—В–µ—З–µ–љ–Є–µ –Љ–Є–љ—Г—В—Л –њ—А–Є 1000 –Њ–±/–Љ–Є–љ, —Б—Г–њ–µ—А–љ–∞—В–∞–љ—В —Г–і–∞–ї—П–ї–Є, –∞¬†–Њ—Б–∞–і–Њ–Ї —А–µ—Б—Г—Б–њ–µ–љ–і–Є—А–Њ–≤–∞–ї–Є –≤¬†–њ–Є—В–∞—В–µ–ї—М–љ–Њ–є —Б—А–µ–і–µ.
–Ъ—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–Є–µ –љ–µ–є—А–Њ–љ–Њ–≤ –Њ—Б—Г—Й–µ—Б—В–≤–ї—П–ї–Є –≤¬†96-–ї—Г–љ–Њ—З–љ—Л—Е –њ–ї–∞—Б—В–Є–Ї–Њ–≤—Л—Е –њ–ї–∞–љ—И–µ—В–∞—Е, –њ–Њ–Ї—А—Л—В—Л—Е –њ–Њ–ї–Є—Н—В–Є–ї–µ–љ–Є–Љ–Є–љ–Њ–Љ –Є–ї–Є –њ–Њ–ї–Є–ї–Є–Ј–Є–љ–Њ–Љ (25 –Љ–Ь —Е–ї–Њ—А–Є–і–∞ –Ї–∞–ї–Є—П). –Т¬†–Ї–∞–ґ–і—Г—О —П—З–µ–є–Ї—Г –њ–ї–∞–љ—И–µ—В–∞ –і–Њ–±–∞–≤–ї—П–ї–Є –њ–Њ¬†0,1¬†–Љ–ї —Б—Г—Б–њ–µ–љ–Ј–Є–Є –Ї–ї–µ—В–Њ–Ї. –Ъ—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–Є–µ –њ—А–Њ–≤–Њ–і–Є–ї–Є –≤¬†—В–µ—З–µ–љ–Є–µ —Б–µ–Љ–Є-–≤–Њ—Б—М–Љ–Є —Б—Г—В–Њ–Ї –≤¬†–°–Ю2-–Є–љ–Ї—Г–±–∞—В–Њ—А–µ, –Ј–∞–њ–Њ–ї–љ–µ–љ–љ–Њ–Љ¬†–≥–∞–Ј–Њ–≤–Њ–є —Б–Љ–µ—Б—М—О (95% –≤–Њ–Ј–і—Г—Е–∞ + 5% –°–Ю2), –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ 35,5 –Њ–° –Є¬†–Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ–є –≤–ї–∞–ґ–љ–Њ—Б—В–Є 98%. –Ъ¬†—Н—В–Њ–Љ—Г —Б—А–Њ–Ї—Г –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–љ—Л–µ –Ј–µ—А–љ–Є—Б—В—Л–µ –љ–µ–є—А–Њ–љ—Л –і–Њ—Б—В–Є–≥–∞–ї–Є –Љ–Њ—А—Д–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є –Є¬†–љ–µ–є—А–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Њ–є –Ј—А–µ–ї–Њ—Б—В–Є. –°–Њ—Б—В–Њ—П–љ–Є–µ –Ї—Г–ї—М—В—Г—А –Ї–Њ–љ—В—А–Њ–ї–Є—А–Њ–≤–∞–ї–Є –µ–ґ–µ–і–љ–µ–≤–љ–Њ –Є¬†–љ–∞ –Ї–∞–ґ–і–Њ–Љ —Н—В–∞–њ–µ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞ –њ—Г—В–µ–Љ –≤–Є–Ј—Г–∞–ї—М–љ–Њ–≥–Њ –њ—А–Њ—Б–Љ–Њ—В—А–∞ –≤¬†–Є–љ–≤–µ—А—В–Є—А–Њ–≤–∞–љ–љ–Њ–Љ –Љ–Є–Ї—А–Њ—Б–Ї–Њ–њ–µ –њ—А–Є —Д–∞–Ј–Њ–≤–Њ–Љ –Ї–Њ–љ—В—А–∞—Б—В–µ. –Т–µ—Й–µ—Б—В–≤–∞ –і–Њ–±–∞–≤–ї—П–ї–Є –≤¬†—Б—А–µ–і—Г –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–Є—П –љ–∞¬†–≤—В–Њ—А—Л–µ —Б—Г—В–Ї–Є in¬†vitro –љ–∞¬†–≤–µ—Б—М —Б—А–Њ–Ї –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–Є—П (–і–Њ —Б–µ–Љ–Є —Б—Г—В–Њ–Ї). –Ш—Б—Е–Њ–і—П –Є–Ј¬†–Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –≤–µ—Й–µ—Б—В–≤–∞ –≤¬†–њ—А–Њ–±–Є—А–Ї–µ (10 –Љ–Ь) –µ–≥–Њ –Љ–Є–љ–Є–Љ–∞–ї—М–љ–Њ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ–µ —А–∞–Ј–≤–µ–і–µ–љ–Є–µ –і–ї—П –і–Њ–±–∞–≤–ї–µ–љ–Є—П –Ї¬†–Ї—Г–ї—М—В—Г—А–∞–Љ, –њ–Њ–Ј–≤–Њ–ї—П—О—Й–µ–µ —Б–Њ—Е—А–∞–љ–Є—В—М –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л–µ —Б–≤–Њ–є—Б—В–≤–∞ –њ–Є—В–∞—В–µ–ї—М–љ–Њ–є —Б—А–µ–і—Л, —Б–Њ—Б—В–∞–≤–ї—П–ї–Њ 1:10, —В–Њ –µ—Б—В—М 1 –Љ–Ь. –С–ї–∞–≥–Њ–і–∞—А—П –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—О 96-–ї—Г–љ–Њ—З–љ—Л—Е –њ–ї–∞—Б—В–Є–Ї–Њ–≤—Л—Е –њ–ї–∞–љ—И–µ—В–Њ–≤ –Љ–Њ–ґ–љ–Њ –±—Л–ї–Њ —В–µ—Б—В–Є—А–Њ–≤–∞—В—М —Б—А–∞–Ј—Г —З–µ—В—Л—А–µ —А–∞–Ј–ї–Є—З–љ—Л–µ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –Њ–±—А–∞–Ј—Ж–Њ–≤. –С—Л–ї–Є –≤—Л–±—А–∞–љ—Л —Б–ї–µ–і—Г—О—Й–Є–µ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є: 0,1, 0,2, 0,5 –Є¬†1 –Љ–Ь.
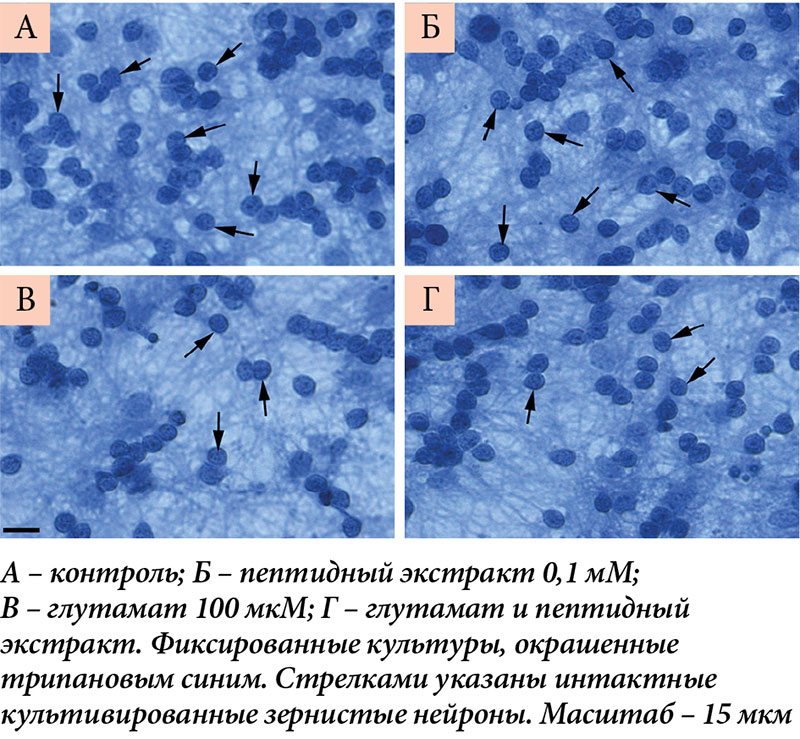
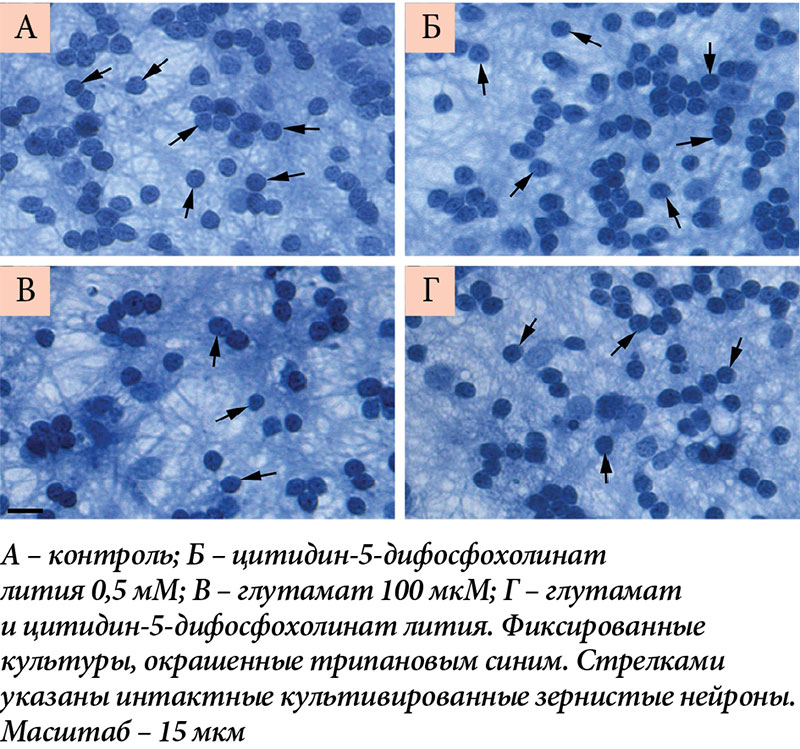
–Ъ–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ—Г—О –Њ—Ж–µ–љ–Ї—Г –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В–Є –Ї–ї–µ—В–Њ–Ї –њ—А–Њ–≤–Њ–і–Є–ї–Є —Б¬†–њ–Њ–Љ–Њ—Й—М—О –њ—А—П–Љ–Њ–≥–Њ –њ–Њ–і—Б—З–µ—В–∞ –ґ–Є–≤—Л—Е –љ–µ–є—А–Њ–љ–Њ–≤. –Ъ–ї–µ—В–Ї–Є-–Ј–µ—А–љ–∞ –ї–µ–≥–Ї–Њ –Є–і–µ–љ—В–Є—Д–Є—Ж–Є—А–Њ–≤–∞—В—М –њ—А–Є–ґ–Є–Ј–љ–µ–љ–љ–Њ –Ї–∞–Ї –љ–µ–±–Њ–ї—М—И–Є–µ (7вАУ10 –Љ–Ї–Љ –≤¬†–і–Є–∞–Љ–µ—В—А–µ) –Њ–Ї—А—Г–≥–ї—Л–µ –Є–ї–Є –Њ–≤–∞–ї—М–љ—Л–µ –љ–µ–є—А–Њ–љ—Л. –Я—А–Є –Њ–Ї—А–∞—Б–Ї–µ —Д–Є–Ї—Б–Є—А–Њ–≤–∞–љ–љ—Л—Е –Ї—Г–ї—М—В—Г—А —В—А–Є–њ–∞–љ–Њ–≤—Л–Љ —Б–Є–љ–Є–Љ —Е–Њ—А–Њ—И–Њ –≤–Є–і–љ—Л —П–і—А–∞ –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–љ—Л—Е –Ј–µ—А–љ–Є—Б—В—Л—Е –љ–µ–є—А–Њ–љ–Њ–≤, –Ј–∞–љ–Є–Љ–∞—О—Й–Є–µ –±–Њ–ї—М—И—Г—О —З–∞—Б—В—М —В–µ–ї –љ–µ–є—А–Њ–љ–Њ–≤ –Є¬†–Њ–Ї—А—Г–ґ–µ–љ–љ—Л–µ —В–Њ–љ–Ї–Є–Љ –Њ–±–Њ–і–Ї–Њ–Љ —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ—Л (—А–Є—Б.¬†3).
–Ф–ї—П –Ї–∞–ґ–і–Њ–≥–Њ –≤–µ—Й–µ—Б—В–≤–∞ –±—Л–ї–Њ –≤—Л–њ–Њ–ї–љ–µ–љ–Њ –Ї–∞–Ї –Љ–Є–љ–Є–Љ—Г–Љ —В—А–Є —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞, –њ—А–Є—З–µ–Љ –і–ї—П –≤—Б–µ—Е —В–Њ—З–µ–Ї –±—А–∞–ї–Є –њ–Њ¬†—В—А–Є –Ї—Г–ї—М—В—Г—А—Л, –≤¬†–Ї–∞–ґ–і–Њ–є –Є–Ј¬†–Ї–Њ—В–Њ—А—Л—Е —Д–Њ—В–Њ–≥—А–∞—Д–Є—А–Њ–≤–∞–ї–Є –Є¬†–њ—А–Њ—Б—З–Є—В—Л–≤–∞–ї–Є –њ–Њ¬†–њ—П—В—М –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ—Л—Е –њ–Њ–ї–µ–є –Ј—А–µ–љ–Є—П (–Ї–∞–Ї –Љ–Є–љ–Є–Љ—Г–Љ 45 –њ–Њ–ї–µ–є –Ј—А–µ–љ–Є—П –Є–Ј¬†–і–µ–≤—П—В–Є –Ї—Г–ї—М—В—Г—А —В—А–µ—Е –љ–µ–Ј–∞–≤–Є—Б–Є–Љ—Л—Е —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–Њ–≤). –Ъ–Њ–ї–Є—З–µ—Б—В–≤–Њ –љ–µ–є—А–Њ–љ–Њ–≤ —Б¬†–љ–µ–Є–Ј–Љ–µ–љ–µ–љ–љ–Њ–є –Љ–Њ—А—Д–Њ–ї–Њ–≥–Є–µ–є –≤¬†–Ї–Њ–љ—В—А–Њ–ї—М–љ—Л—Е –Ї—Г–ї—М—В—Г—А–∞—Е –њ—А–Є–љ–Є–Љ–∞–ї–Є –Ј–∞ 100%-–љ—Г—О –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М. –Ф–ї—П —Б—В–∞—В–Є—Б—В–Є—З–µ—Б–Ї–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–Є —В–µ—Б—В ANOVA —Б¬†–њ–Њ–њ—А–∞–≤–Ї–∞–Љ–Є –С–Њ–љ—Д–µ—А—А–Њ–љ–Є –Є¬†–Ф–∞–љ–љ–µ—В–∞. –Ю—В–ї–Є—З–Є—П –Љ–µ–ґ–і—Г¬†–≥—А—Г–њ–њ–∞–Љ–Є —Б—З–Є—В–∞–ї–Є –і–Њ—Б—В–Њ–≤–µ—А–љ—Л–Љ–Є –њ—А–Є p < 0,05. –†–µ–Ј—Г–ї—М—В–∞—В—Л –≤—Л—А–∞–ґ–∞–ї–Є –Ї–∞–Ї —Б—А–µ–і–љ–µ–µ ¬± SEM.
–†–µ–Ј—Г–ї—М—В–∞—В—Л
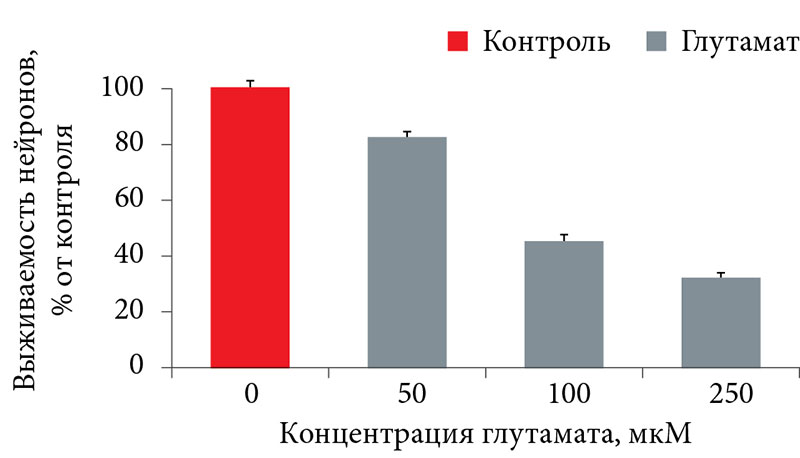
–Я–Њ¬†–і–∞–љ–љ—Л–Љ –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–≥–Њ –Љ–Њ–і–µ–ї–Є—А–Њ–≤–∞–љ–Є—П –њ–Њ–≤—А–µ–ґ–і–µ–љ–Є—П –Ї—Г–ї—М—В—Г—А –љ–µ–є—А–Њ–љ–Њ–≤¬†–≥–ї—Г—В–∞–Љ–∞—В–Њ–Љ, –±—Л–ї–Њ –≤—Л—П–≤–ї–µ–љ–Њ –і–Њ–Ј–Њ–Ј–∞–≤–Є—Б–Є–Љ–Њ–µ —В–Њ–Ї—Б–Є—З–µ—Б–Ї–Њ–µ –≤–ї–Є—П–љ–Є–µ¬†–≥–ї—Г—В–∞–Љ–∞—В–∞ –љ–∞¬†–≤—Л–ґ–Є–≤–∞–љ–Є–µ –љ–µ–є—А–Њ–љ–Њ–≤ (—А–Є—Б.¬†4). –Т—Л–±–Њ—А –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є¬†–≥–ї—Г—В–∞–Љ–∞—В–∞ –≤¬†–Ї–∞–ґ–і–Њ–Љ –Њ–њ—Л—В–µ –Њ—Б—Г—Й–µ—Б—В–≤–ї—П–ї—Б—П —В–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, —З—В–Њ–±—Л –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–љ—Л—Е –Ј–µ—А–љ–Є—Б—В—Л—Е –љ–µ–є—А–Њ–љ–Њ–≤ —Б–Њ—Б—В–∞–≤–ї—П–ї–∞ 30вАУ80% –Њ—В¬†–Є–љ—В–∞–Ї—В–љ–Њ–≥–Њ –Ї–Њ–љ—В—А–Њ–ї—П (—З—В–Њ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г–µ—В —Г–Љ–µ—А–µ–љ–љ–Њ–Љ—Г¬†–≥–ї—Г—В–∞–Љ–∞—В–љ–Њ–Љ—Г —Б—В—А–µ—Б—Б—Г). –Я—А–Є –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В–Є –љ–µ–є—А–Њ–љ–Њ–≤ –Љ–µ–љ–µ–µ 30% (—Б–Є–ї—М–љ—Л–є¬†–≥–ї—Г—В–∞–Љ–∞—В–љ—Л–є —Б—В—А–µ—Б—Б) –Є–ї–Є –±–Њ–ї–µ–µ 80% (—Б–ї–∞–±—Л–є¬†–≥–ї—Г—В–∞–Љ–∞—В–љ—Л–є —Б—В—А–µ—Б—Б) –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–µ —Б–≤–Њ–є—Б—В–≤–∞ –≤–µ—Й–µ—Б—В–≤ –љ–µ¬†—В–∞–Ї –љ–∞–≥–ї—П–і–љ—Л.
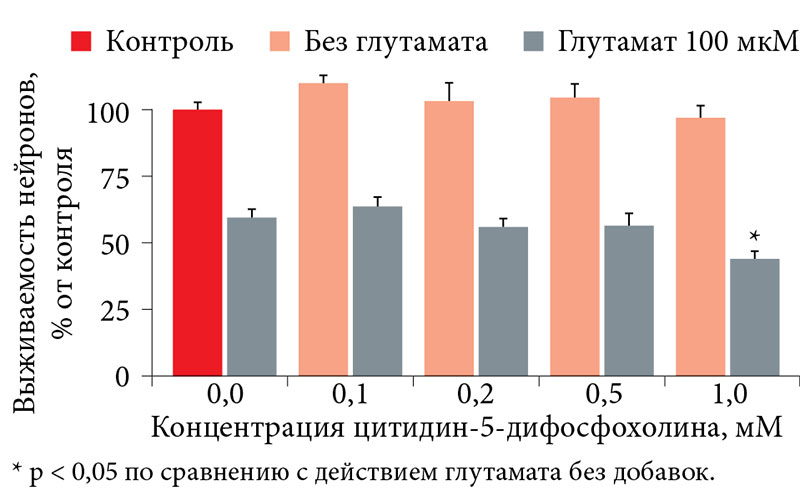
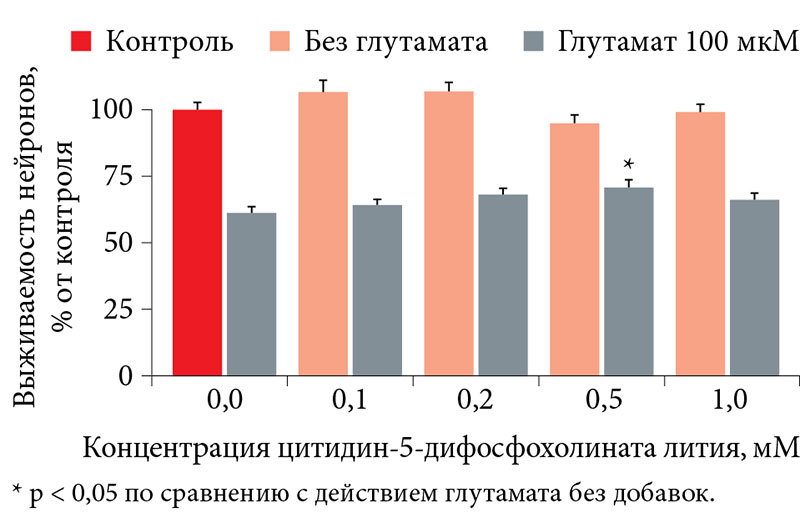
–Ш–Ј–≤–µ—Б—В–љ–Њ, —З—В–Њ –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–µ —Б–≤–Њ–є—Б—В–≤–∞ –≤—Л—П–≤–ї—П—О—В—Б—П, –Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ, –њ—А–Є –і–Њ—Б—В–∞—В–Њ—З–љ–Њ –і–ї–Є—В–µ–ї—М–љ–Њ–Љ –њ—А–Є–Љ–µ–љ–µ–љ–Є–Є. –Я–Њ—Н—В–Њ–Љ—Г –≤¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–∞—Б—М –Њ—В—А–∞–±–Њ—В–∞–љ–љ–∞—П —А–∞–љ–µ–µ ¬Ђ–њ—А–Њ—Д–Є–ї–∞–Ї—В–Є—З–µ—Б–Ї–∞—П¬ї —Б—Е–µ–Љ–∞ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞: –≤–µ—Й–µ—Б—В–≤–Њ –≤–љ–Њ—Б–Є–ї–Њ—Б—М –≤¬†—Б—А–µ–і—Г –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–Є—П –љ–∞¬†–≤—В–Њ—А—Л–µ —Б—Г—В–Ї–Є –Є¬†–Њ—Б—В–∞–≤–∞–ї–Њ—Б—М —В–∞–Љ –і–Њ —Б–µ–і—М–Љ—Л—Е —Б—Г—В–Њ–Ї. –Ч–∞—В–µ–Љ –њ—А–Њ–≤–Њ–і–Є–ї–∞—Б—М –Њ–±—А–∞–±–Њ—В–Ї–∞ –Ї–ї–µ—В–Њ–Ї¬†–≥–ї—Г—В–∞–Љ–∞—В–Њ–Љ –Є¬†–њ–Њ–і—Б—З–Є—В—Л–≤–∞–ї–Њ—Б—М —З–Є—Б–ї–Њ –≤—Л–ґ–Є–≤—И–Є—Е –љ–µ–є—А–Њ–љ–Њ–≤ [9]. –Э–µ–є—А–Њ—Ж–Є—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В—Л –±—Л–ї–Є –њ—А–Њ–≤–µ–і–µ–љ—Л –і–ї—П –Љ–Є–Њ¬≠–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –Є¬†—В—А–µ—Е –і—А—Г–≥–Є—Е –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л—Е —Б—А–µ–і—Б—В–≤: –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞ –њ–µ–њ—В–Є–і–Њ–≤, –њ–Њ–ї—Г—З–µ–љ–љ—Л—Е –Є–Ј¬†–≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ —Б–≤–Є–љ—М–Є, —Ж–Є—В–Є–і–Є–љ-5-–і–Є—Д–Њ—Б—Д–Њ—Е–Њ–ї–Є–љ–∞, —Ж–Є—В–Є–і–Є–љ-5-–і–Є—Д–Њ—Б—Д–Њ—Е–Њ–ї–Є–љ–∞—В–∞ –ї–Є—В–Є—П. –Т¬†—Е–Њ–і–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –±—Л–ї–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Њ 1920 –Ї—Г–ї—М—В—Г—А –Є¬†–≤—Л–њ–Њ–ї–љ–µ–љ—Л –њ–Њ–і—Б—З–µ—В—Л –±–Њ–ї–µ–µ —З–µ–Љ –і–ї—П 200 —В—Л—Б. –љ–µ–є—А–Њ–љ–Њ–≤.
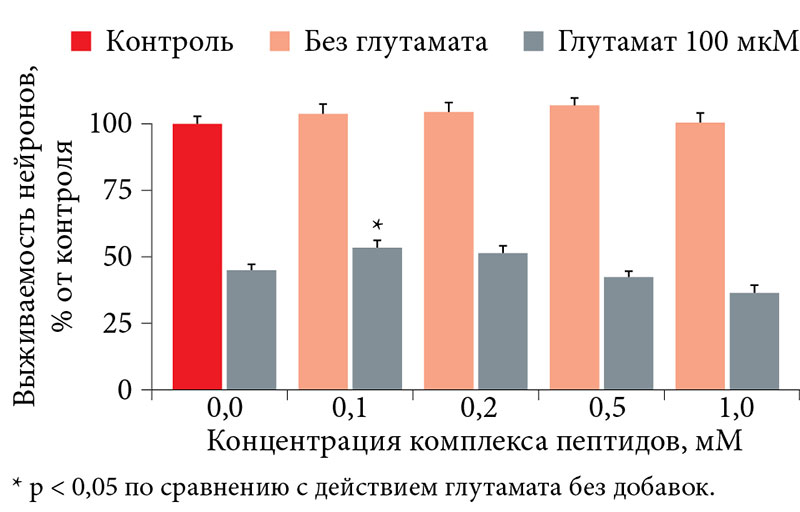
–Ь–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї –љ–µ¬†–њ—А–Њ—П–≤–ї—П–ї —В–Њ–Ї—Б–Є—З–µ—Б–Ї–Є—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –Є¬†–љ–µ –≤–ї–Є—П–ї –љ–∞¬†–≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–љ—Л—Е –Ј–µ—А–љ–Є—Б—В—Л—Е –љ–µ–є—А–Њ–љ–Њ–≤ –≤¬†–Ї–Њ–љ—В—А–Њ–ї–µ, —В–Њ –µ—Б—В—М –≤¬†¬Ђ—Е–Њ–ї–Њ—Б—В–Њ–Љ¬ї —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–µ (–±–µ–Ј –і–Њ–±–∞–≤–ї–µ–љ–Є—П¬†–≥–ї—Г—В–∞–Љ–∞—В–∞). –Я—А–Є –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–Є¬†–≥–ї—Г—В–∞–Љ–∞—В–∞ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї –≤¬†–Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є 0,2вАУ0,5 –Љ–Ь –і–Њ—Б—В–Њ–≤–µ—А–љ–Њ –њ–Њ–≤—Л—И–∞–ї –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –љ–µ–є—А–Њ–љ–Њ–≤ –љ–∞¬†12вАУ17%. –Ґ–∞–Ї, –њ—А–Є –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ 0,2¬†–Љ–Ь –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –љ–µ–є—А–Њ–љ–Њ–≤ —Б–Њ—Б—В–∞–≤–Є–ї–∞ 54,9 ¬± 2,6%, –њ—А–Є 0,5¬†–Љ–Ь¬†вАУ 59,1 ¬± 2,9% (—А–µ–Ј—Г–ї—М—В–∞—В—Л¬† –Ї–Њ–љ—В—А–Њ–ї—П¬†вАУ 42,6 ¬± 2,2%, —А–Є—Б.¬†5). –Ш–Ј–Њ–±—А–∞–ґ–µ–љ–Є—П –Њ–±—Б—З–Є—В–∞–љ–љ—Л—Е –Ї—Г–ї—М—В—Г—А –љ–µ–є—А–Њ–љ–Њ–≤ –њ—А–Є–≤–µ–і–µ–љ—Л –љ–∞¬†—А–Є—Б. 6.
–Ф—А—Г–≥–Є–µ —Б—А–µ–і—Б—В–≤–∞ –њ—А–Њ–і–µ–Љ–Њ–љ—Б—В—А–Є—А–Њ–≤–∞–ї–Є¬†–≥–Њ—А–∞–Ј–і–Њ –Љ–µ–љ–µ–µ –≤—Л—А–∞–ґ–µ–љ–љ—Л–µ –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–µ —Б–≤–Њ–є—Б—В–≤–∞. –Ъ–Њ–Љ–њ–ї–µ–Ї—Б –њ–µ–њ—В–Є–і–Њ–≤, –њ–Њ–ї—Г—З–µ–љ–љ—Л—Е –Є–Ј¬†–≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ —Б–≤–Є–љ—М–Є, –њ–Њ–Ї–∞–Ј–∞–ї —Б–ї–∞–±—Л–є, –љ–Њ¬†–і–Њ—Б—В–Њ–≤–µ—А–љ—Л–є –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–є —Н—Д—Д–µ–Ї—В¬†вАУ —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В–Є –љ–∞¬†5вАУ8%. –Ґ–∞–Ї, –Њ–±—Й–∞—П –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –њ—А–Є –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є 0,1¬†–Љ–Ь —Б–Њ—Б—В–∞–≤–Є–ї–∞ 53¬†¬± 3%, 0,2¬†–Љ–Ь¬†вАУ 52 ¬± 3% (–≤ –Ї–Њ–љ—В—А–Њ–ї–µ¬†вАУ 45 ¬± 2%, —А < 0,05). –С–Њ–ї–µ–µ —В–Њ–≥–Њ, –њ—А–Є –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞ –њ–µ–њ—В–Є–і–Њ–≤, —А–∞–≤–љ–Њ–є 1,0 –Љ–Ь, –±—Л–ї –Њ–±–љ–∞—А—Г–ґ–µ–љ —Б–ї–∞–±—Л–є —В–Њ–Ї—Б–Є—З–µ—Б–Ї–Є–є —Н—Д—Д–µ–Ї—В¬†вАУ –≤—Л–ґ–Є–≤–∞–ї–Њ –≤—Б–µ–≥–Њ 36,3 ¬± 2,6% –љ–µ–є—А–Њ–љ–Њ–≤ (—А–Є—Б.¬†7). –§–Њ—В–Њ–≥—А–∞—Д–Є–Є –Ї—Г–ї—М—В—Г—А –Є–Ј¬†–њ—А–Њ—Б—З–Є—В–∞–љ–љ—Л—Е –њ–Њ–ї–µ–є –Ј—А–µ–љ–Є—П –њ—А–Є–≤–µ–і–µ–љ—Л –љ–∞¬†—А–Є—Б. 8.
–£¬†–Њ—Б—В–∞–ї—М–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ—Л—Е –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л—Е —Б—А–µ–і—Б—В–≤ –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ–µ –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –љ–∞¬†–≤—Л–ґ–Є–≤–∞–љ–Є–µ –љ–µ–є—А–Њ–љ–Њ–≤ –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –Њ—В—Б—Г—В—Б—В–≤–Њ–≤–∞–ї–Њ. –Э–∞–њ—А–Є–Љ–µ—А, –њ—А–Є –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–Є —Ж–Є—В–Є–і–Є–љ-5-–і–Є—Д–Њ—Б—Д–Њ—Е–Њ–ї–Є–љ–∞ –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –≤¬†–Ї–Њ–љ—В—А–Њ–ї–µ –њ—А–Є –і–µ–є—Б—В–≤–Є–Є¬†–≥–ї—Г—В–∞–Љ–∞—В–∞ —Б–Њ—Б—В–∞–≤–Є–ї–∞ 59,3 ¬± 3,0%, –∞¬†–њ—А–Є –і–Њ–±–∞–≤–ї–µ–љ–Є–Є —Ж–Є—В–Є–і–Є–љ-5-–і–Є—Д–Њ—Б—Д–Њ—Е–Њ–ї–Є–љ–∞¬†вАУ –љ–µ¬†–њ—А–µ–≤—Л—И–∞–ї–∞ 65% (0,1¬†–Љ–Ь¬†вАУ 63,6 ¬± 3,6%, –љ–µ—В –і–Њ—Б—В–Њ–≤–µ—А–љ—Л—Е —А–∞–Ј–ї–Є—З–Є–є, p > 0,05). –Я—А–Є —Н—В–Њ–Љ –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–∞—П –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є—П —Ж–Є—В–Є–і–Є–љ-5-–і–Є—Д–Њ—Б—Д–Њ—Е–Њ–ї–Є–љ–∞ (1 –Љ–Ь) –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —Г–Љ–µ–љ—М—И–∞–ї–∞ –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –Ї—Г–ї—М—В–Є–≤–Є—А–Њ–≤–∞–љ–љ—Л—Е –Ј–µ—А–љ–Є—Б—В—Л—Е –љ–µ–є—А–Њ–љ–Њ–≤¬†вАУ 43,6 ¬± 2,4% (—А–Є—Б.¬†9 –Є¬†10). –°—Е–Њ–ґ–∞—П —Б–Є—В—Г–∞—Ж–Є—П –љ–∞–±–ї—О–і–∞–ї–∞—Б—М –Є¬†–њ—А–Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–Є –њ–Њ—В–µ–љ—Ж–Є–∞–ї—М–љ–Њ–≥–Њ –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Њ—А–∞ —Ж–Є—В–Є–і–Є–љ-5-–і–Є—Д–Њ—Б—Д–Њ—Е–Њ–ї–Є–љ–∞—В–∞ –ї–Є—В–Є—П (—А–Є—Б.¬†11 –Є¬†12).
–Ю–±—Б—Г–ґ–і–µ–љ–Є–µ —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤
–†–µ–Ј—Г–ї—М—В–∞—В—Л –њ—А–Њ–≤–µ–і–µ–љ–љ–Њ–≥–Њ –љ–µ–є—А–Њ—Ж–Є—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –≤–∞–ґ–љ—Л —Б¬†—В–Њ—З–Ї–Є –Ј—А–µ–љ–Є—П –Њ—Ж–µ–љ–Ї–Є –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –Ї–∞–Ї –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞, —В–∞–Ї –Є¬†¬Ђ–њ—А–Є–Ј–љ–∞–љ–љ—Л—Е¬ї –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Њ—А–Њ–≤. –Ъ–∞–Ї –±—Л–ї–Њ –Њ—В–Љ–µ—З–µ–љ–Њ –≤—Л—И–µ, –љ–µ–є—А–Њ—Ж–Є—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П —Г–љ–Є–Ї–∞–ї—М–љ—Л, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г –њ–Њ–Ј–≤–Њ–ї—П—О—В –њ–Њ–і—В–≤–µ—А–і–Є—В—М –њ—А—П–Љ–Њ–µ –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –Є–Ј—Г—З–∞–µ–Љ—Л—Е –≤–µ—Й–µ—Б—В–≤ –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –љ–∞¬†—А–∞—Б—В—Г—Й–Є–µ –љ–µ–є—А–Њ–љ—Л –њ–ї–Њ–і–∞. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П —Н—Д—Д–µ–Ї—В –і—А—Г–≥–Є—Е –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Њ—А–Њ–≤ –Љ–Њ–ґ–µ—В –±—Л—В—М –Њ–њ–Њ—Б—А–µ–і–Њ–≤–∞–љ–љ—Л–Љ: —Д–Њ—Б—Д–Њ—Е–Њ–ї–Є–љ, –љ–∞–њ—А–Є–Љ–µ—А, –Љ–Њ–ґ–µ—В –Њ–Ї–∞–Ј—Л–≤–∞—В—М –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–µ –і–µ–є—Б—В–≤–Є–µ —З–µ—А–µ–Ј –њ–Њ–і–і–µ—А–ґ–Ї—Г —Д—Г–љ–Ї—Ж–Є–Є –њ–µ—З–µ–љ–Є [15].
–Я–Њ¬†—А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ –њ—А–Њ–≤–µ–і–µ–љ–Є—П –љ–∞—Б—В–Њ—П—Й–µ–є —Б–µ—А–Є–Є —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–Њ–≤ —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ ¬Ђ–њ—А–Њ—Д–Є–ї–∞–Ї—В–Є—З–µ—Б–Ї–Њ–µ¬ї –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ (–Ј–∞ –њ—П—В—М —Б—Г—В–Њ–Ї –і–Њ —Б–Њ–Ј–і–∞–љ–Є—П¬†–≥–ї—Г—В–∞–Љ–∞—В–љ–Њ–≥–Њ —Б—В—А–µ—Б—Б–∞) –і–Њ—Б—В–Њ–≤–µ—А–љ–Њ –Є¬†—Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ (–≤¬†—Б—А–µ–і–љ–µ–Љ –љ–∞¬†12вАУ17%, —А = 0,01) –њ–Њ–≤—Л—И–∞–µ—В –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –љ–µ–є—А–Њ–љ–Њ–≤ –≤¬†–Ї—Г–ї—М—В—Г—А–µ. –°–Њ–Ј–і–∞–љ–Є–µ¬†–≥–ї—Г—В–∞–Љ–∞—В–љ–Њ–≥–Њ —Б—В—А–µ—Б—Б–∞ –≤¬†–Ї—Г–ї—М—В—Г—А–µ –љ–µ–є—А–Њ–љ–Њ–≤ –Љ–Њ–і–µ–ї–Є—А—Г–µ—В –Є—И–µ–Љ–Є—О –Љ–Њ–Ј–≥–∞ –њ–ї–Њ–і–∞, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –њ—А–Є –њ–∞—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —А–Њ–і–∞—Е. –Ю–±–Њ–≥–∞—Й–µ–љ–Є–µ –Ї–ї–µ—В–Њ—З–љ–Њ–є —Б—А–µ–і—Л –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–Њ–Љ –і–Њ —Б–Њ–Ј–і–∞–љ–Є—П¬†–≥–ї—Г—В–∞–Љ–∞—В–љ–Њ–≥–Њ —Б—В—А–µ—Б—Б–∞ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г–µ—В –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є—З–µ—Б–Ї–Є–Љ –і–Њ—В–∞—Ж–Є—П–Љ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –≤–Њ –≤—А–µ–Љ—П –±–µ—А–µ–Љ–µ–љ–љ–Њ—Б—В–Є. –Я–Њ—Н—В–Њ–Љ—Г –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л –љ–µ–є—А–Њ—Ж–Є—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –Є–Љ–µ—О—В –Ј–љ–∞—З–µ–љ–Є–µ –і–ї—П –∞–љ—В–Є–Є—И–µ–Љ–Є—З–µ—Б–Ї–Њ–є –Ј–∞—Й–Є—В—Л –Љ–Њ–Ј–≥–∞ –њ–ї–Њ–і–∞ –Є¬†–≤ —А–∞–љ–љ–Є–µ, –Є¬†–≤ –њ–Њ–Ј–і–љ–Є–µ —Б—А–Њ–Ї–Є¬†–≥–µ—Б—В–∞—Ж–Є–Є.
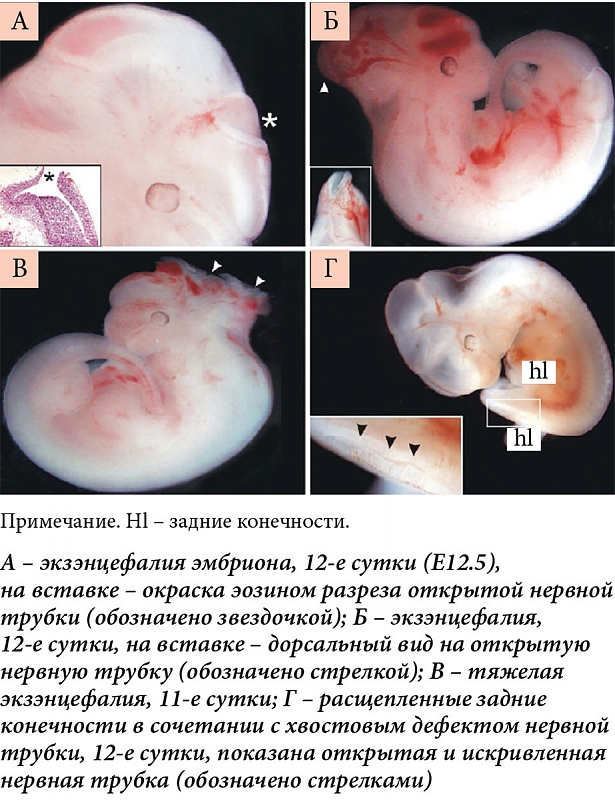
–Ш—И–µ–Љ–Є—З–µ—Б–Ї–Є–є —Б—В—А–µ—Б—Б, –њ–µ—А–µ–љ–µ—Б–µ–љ–љ—Л–є —Н–Љ–±—А–Є–Њ–љ–Њ–Љ –≤¬†—А–∞–љ–љ–Є–µ —Б—А–Њ–Ї–Є –±–µ—А–µ–Љ–µ–љ–љ–Њ—Б—В–Є, —Б—В–Є–Љ—Г–ї–Є—А—Г–µ—В —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–µ –≤—А–Њ–ґ–і–µ–љ–љ—Л—Е –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П –Љ–Њ–Ј–≥–∞. –Ю–љ–Є –≤–Њ–Ј–љ–Є–Ї–∞—О—В –њ–Њ–і –і–µ–є—Б—В–≤–Є–µ–Љ —А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–љ—Л—Е —Д–∞–Ї—В–Њ—А–Њ–≤ —В–µ—А–∞—В–Њ–≥–µ–љ–µ–Ј–∞: –∞–ї–Ї–Њ–≥–Њ–ї—П, –Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є, –њ–µ—А–µ–і–∞—О—Й–Є—Е—Б—П –Њ—В¬†–Љ–∞—В–µ—А–Є –њ–ї–Њ–і—Г, –Є–Њ–љ–Є–Ј–Є—А—Г—О—Й–µ–≥–Њ –Є–Ј–ї—Г—З–µ–љ–Є—П, —Д–∞—А–Љ–∞—Ж–µ–≤—В–Є—З–µ—Б–Ї–Є—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤, –љ–Є–Ї–Њ—В–Є–љ–∞, –≤–і—Л—Е–∞–µ–Љ–Њ–≥–Њ —Б¬†—Б–Є–≥–∞—А–µ—В–љ—Л–Љ –і—Л–Љ–Њ–Љ, –Є¬†–і—А. –§–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–µ –≤—А–Њ–ґ–і–µ–љ–љ—Л—Е –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П –Љ–Њ–Ј–≥–∞ —В–∞–Ї–ґ–µ –Љ–Њ–ґ–µ—В –±—Л—В—М —Б–ї–µ–і—Б—В–≤–Є–µ–Љ –і–Є—Б–±–∞–ї–∞–љ—Б–∞ (–Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ, –љ–µ–і–Њ—Б—В–∞—В–Ї–∞) —Д–∞–Ї—В–Њ—А–Њ–≤ —А–Њ—Б—В–∞, –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л—Е –і–ї—П —А–Њ—Б—В–∞ —Н–Љ–±—А–Є–Њ–љ–∞: –≤–Є—В–∞–Љ–Є–љ–∞ –Р, —Д–Њ–ї–∞—В–Њ–≤ (–≤–Є—В–∞–Љ–Є–љ–∞ –Т9) –Є¬†—Б–Є–љ–µ—А–≥–Є—З–љ—Л—Е —Б¬†–љ–Є–Љ–Є –њ–Є—А–Є–і–Њ–Ї—Б–Є–љ–∞ (–≤–Є—В–∞–Љ–Є–љ–∞ –Т6), —Ж–Є–љ–Ї–∞, –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ (–≤–Є—В–∞–Љ–Є–љ–∞ –Т8) –Є¬†–і—А—Г–≥–Є—Е —Н—Б—Б–µ–љ—Ж–Є–∞–ї—М–љ—Л—Е –Љ–Є–Ї—А–Њ–љ—Г—В—А–Є–µ–љ—В–Њ–≤ [16].
–С–Њ–ї—М—И–Є–љ—Б—В–≤–Њ (–±–Њ–ї–µ–µ 70%) –≤—А–Њ–ґ–і–µ–љ–љ—Л—Е –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П –Љ–Њ–Ј–≥–∞ (–≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –і–µ—Д–µ–Ї—В—Л –љ–µ—А–≤–љ–Њ–є —В—А—Г–±–Ї–Є, —А–∞—Б—Й–µ–ї–Є–љ—Л –љ–µ–±–∞) —Д–Њ–ї–∞—В—З—Г–≤—Б—В–≤–Є—В–µ–ї—М–љ—Л, –Є¬†–Є—Е —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–µ –Љ–Њ–ґ–љ–Њ –њ—А–µ–і—Г–њ—А–µ–і–Є—В—М –і–Њ—В–∞—Ж–Є—П–Љ–Є —Д–Њ–ї–∞—В–Њ–≤ (–њ—А–µ–і–њ–Њ—З—В–Є—В–µ–ї—М–љ–Њ –≤¬†—Б–Њ—Б—В–∞–≤–µ –≤–Є—В–∞–Љ–Є–љ–љ–Њ-–Љ–Є–љ–µ—А–∞–ї—М–љ—Л—Е –Ї–Њ–Љ–њ–ї–µ–Ї—Б–Њ–≤) –≤¬†—А–∞–љ–љ–Є–µ —Б—А–Њ–Ї–Є –±–µ—А–µ–Љ–µ–љ–љ–Њ—Б—В–Є. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–µ –≤—А–Њ–ґ–і–µ–љ–љ—Л—Е –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П –Љ–Њ–Ј–≥–∞, –љ–µ—Б–Љ–Њ—В—А—П –љ–∞¬†–і–Њ—В–∞—Ж–Є–Є —Д–Њ–ї–∞—В–Њ–≤, —Г–Ї–∞–Ј—Л–≤–∞–µ—В –љ–∞¬†—Б—Г—Й–µ—Б—В–≤–Њ–≤–∞–љ–Є–µ —Д–Њ–ї–∞—В—А–µ–Ј–Є—Б—В–µ–љ—В–љ—Л—Е –≤—А–Њ–ґ–і–µ–љ–љ—Л—Е –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П –Љ–Њ–Ј–≥–∞ (30%). –†–Є—Б–Ї —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—П —В–∞–Ї–Є—Е –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П –Љ–Њ–ґ–љ–Њ –Ј–љ–∞—З–Є–Љ–Њ —Б–љ–Є–Ј–Є—В—М –њ—Г—В–µ–Љ –њ—А–Є–µ–Љ–∞ –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ [17], –њ–Њ–і–і–µ—А–ґ–Є–≤–∞—О—Й–µ–≥–Њ —Н–Љ–±—А–Є–Њ–≥–µ–љ–µ–Ј –Є¬†—А–∞–Ј–≤–Є—В–Є–µ –њ–ї–Њ–і–∞ [18].
–£—Б—В–∞–љ–Њ–≤–ї–µ–љ–љ—Л–є –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ—Л–є –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–є —Н—Д—Д–µ–Ї—В –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞¬†вАУ –≤–∞–ґ–љ–∞—П —Б–Њ—Б—В–∞–≤–ї—П—О—Й–∞—П —Н–Љ–±—А–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–≥–Њ –і–µ–є—Б—В–≤–Є—П –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞. –Т¬†—З–∞—Б—В–љ–Њ—Б—В–Є, —В—А—Г–і–љ–Њ –њ–µ—А–µ–Њ—Ж–µ–љ–Є—В—М —А–Њ–ї—М –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –≤¬†–њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–µ –≤—А–Њ–ґ–і–µ–љ–љ—Л—Е –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П, —Б–≤—П–Ј–∞–љ–љ—Л—Е —Б¬†–Є–љ—Б—Г–ї–Є–љ–Њ—А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В—М—О, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г –њ—А–Њ–Є–Ј–≤–Њ–і–љ—Л–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ —Г—З–∞—Б—В–≤—Г—О—В –≤¬†–њ—А–Њ—Ж–µ—Б—Б–∞—Е –њ–µ—А–µ–і–∞—З–Є —Б–Є–≥–љ–∞–ї–∞ –Њ—В¬†–Є–љ—Б—Г–ї–Є–љ–Њ–≤–Њ–≥–Њ —А–µ—Ж–µ–њ—В–Њ—А–∞ [19]. –Ш—Б—В–Њ—Й–µ–љ–Є–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –≤¬†—Н–Љ–±—А–Є–Њ–љ–∞–ї—М–љ–Њ–є —В–Ї–∞–љ–Є –љ–∞¬†—Н—В–∞–њ–µ –Њ—А–≥–∞–љ–Њ–≥–µ–љ–µ–Ј–∞ –Є–≥—А–∞–µ—В, –њ–Њ¬†–≤—Б–µ–є –≤–Є–і–Є–Љ–Њ—Б—В–Є, –≤–µ–і—Г—Й—Г—О —А–Њ–ї—М –≤¬†–Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–љ–Є–Є —Н–Љ–±—А–Є–Њ–њ–∞—В–Є–є, –≤—Л–Ј—Л–≤–∞–µ–Љ—Л—Е¬†–≥–Є–њ–µ—А–≥–ї–Є–Ї–µ–Љ–Є–µ–є. –Т¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–µ —Б¬†–Љ–Њ–і–µ–ї—П–Љ–Є —Б—В—А–µ–њ—В–Њ–Ј–Њ—В–Њ—Ж–Є–љ–Њ–≤–Њ–≥–Њ –і–Є–∞–±–µ—В–∞ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –≤¬†—Н–Љ–±—А–Є–Њ–љ–∞—Е –±—Л–ї–Њ –љ–Є–ґ–µ –љ–∞¬†36% (p = 0,01) –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–Ї–Њ–љ—В—А–Њ–ї–µ–Љ, –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–ї–Њ—Б—М —Б¬†–њ–Њ–≤—Л—И–µ–љ–љ–Њ–є —З–∞—Б—В–Њ—В–Њ–є –љ–µ–є—А–Њ–љ–љ—Л—Е –њ–Њ–≤—А–µ–ґ–і–µ–љ–Є–є (17,6% –≤¬†–Њ—Б–љ–Њ–≤–љ–Њ–є¬†–≥—А—Г–њ–њ–µ –Є¬†1,9% –≤¬†–≥—А—Г–њ–њ–µ –Ї–Њ–љ—В—А–Њ–ї—П, —А < 0,001) –Є¬†–Ј–∞–і–µ—А–ґ–Ї–Њ–є —А–∞–Ј–≤–Є—В–Є—П. –Ґ–∞–Ї, –і–ї–Є–љ–∞ —Н–Љ–±—А–Є–Њ–љ–∞ –≤¬†–Њ—Б–љ–Њ–≤–љ–Њ–є¬†–≥—А—Г–њ–њ–µ —Б–Њ—Б—В–∞–≤–Є–ї–∞ 3,37 ¬± 0,04 –Љ–Љ –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†3,87 ¬± 0,03 –Љ–Љ –≤¬†–≥—А—Г–њ–њ–µ –Ї–Њ–љ—В—А–Њ–ї—П (p = 0,01), –∞¬†—З–Є—Б–ї–Њ —Б–Њ–Љ–Є—В–Њ–≤¬†вАУ 27,5¬†¬±¬†0,2 –Є¬†29,1 ¬± 0,2 —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ (p = 0,01) [20].
–Ь–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В —Г–Љ–µ–љ—М—И–µ–љ–Є—О –Є–љ—Б—Г–ї–Є–љ–Њ—А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є –Є¬†–Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ –љ–µ–Њ–±—Е–Њ–і–Є–Љ –і–ї—П –њ—А–µ–Њ–і–Њ–ї–µ–љ–Є—П –љ–µ–≥–∞—В–Є–≤–љ–Њ–≥–Њ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –љ–∞¬†–љ–µ–є—А–Њ–љ—Л –њ–Њ–≤—Л—И–µ–љ–љ—Л—Е —Г—А–Њ–≤–љ–µ–є¬†–≥–ї—О–Ї–Њ–Ј—Л. –Т¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–µ –њ—А–Є–µ–Љ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –њ—А–Є–≤–Њ–і–Є–ї –Ї¬†–≤—Л—А–∞–ґ–µ–љ–љ–Њ–Љ—Г —Б–љ–Є–ґ–µ–љ–Є—О —З–∞—Б—В–Њ—В—Л —А–∞–Ј–≤–Є—В–Є—П –і–µ—Д–µ–Ї—В–Њ–≤ –љ–µ—А–≤–љ–Њ–є —В—А—Г–±–Ї–Є –≤¬†–Љ–Њ–і–µ–ї–Є —Б—В—А–µ–њ—В–Њ–Ј–Њ—В–Њ—Ж–Є–љ–Њ–≤–Њ–≥–Њ –і–Є–∞–±–µ—В–∞, –Ї–Њ—В–Њ—А–∞—П —Б–Њ—Б—В–∞–≤–Є–ї–∞ 9,5% –≤¬†–≥—А—Г–њ–њ–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –Є¬†20,4% –≤¬†–≥—А—Г–њ–њ–µ –Ї–Њ–љ—В—А–Њ–ї—П (—А < 0,05) [21].
–Р–љ–∞–ї–Є–Ј —Д–Њ–ї–∞—В—З—Г–≤—Б—В–≤–Є—В–µ–ї—М–љ—Л—Е –Є¬†—Д–Њ–ї–∞—В—А–µ–Ј–Є—Б—В–µ–љ—В–љ—Л—Е –Љ–Њ–і–µ–ї–µ–є –і–µ—Д–µ–Ї—В–Њ–≤ –љ–µ—А–≤–љ–Њ–є —В—А—Г–±–Ї–Є [22] –≤¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞—Е –њ–Њ¬†–і–µ–ї–µ—Ж–Є–Є¬†–≥–µ–љ–Њ–≤ –њ–Њ–Ј–≤–Њ–ї–Є–ї —Г—Б—В–∞–љ–Њ–≤–Є—В—М –±–Њ–ї–µ–µ 60¬†–≥–µ–љ–Њ–≤, –Є–љ–∞–Ї—В–Є–≤–∞—Ж–Є—П –Ї–Њ—В–Њ—А—Л—Е –њ—А–Є–≤–Њ–і–Є—В –Ї¬†–њ–Њ—П–≤–ї–µ–љ–Є—О –ї–Є–љ–Є–є –Љ—Л—И–µ–є —Б¬†–і–µ—Д–µ–Ї—В–∞–Љ–Є –љ–µ—А–≤–љ–Њ–є —В—А—Г–±–Ї–Є [23]. –Э–µ¬†–Љ–µ–љ–µ–µ 22 –Є–Ј¬†—Н—В–Є—Е 60¬†–≥–µ–љ–Њ–≤ –Ї–Њ–і–Є—А—Г—О—В –±–µ–ї–Ї–Є –Є¬†—Д–µ—А–Љ–µ–љ—В—Л, –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –Є–ї–Є —Г—А–Њ–≤–љ–Є –Ї–Њ—В–Њ—А—Л—Е —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –Ј–∞–≤–Є—Б—П—В –Њ—В¬†–Њ–њ—А–µ–і–µ–ї–µ–љ–љ—Л—Е –Љ–Є–Ї—А–Њ–љ—Г—В—А–Є–µ–љ—В–љ—Л—Е –Ї–Њ—Д–∞–Ї—В–Њ—А–Њ–≤ (—В–∞–±–ї–Є—Ж–∞).
–Т–Њ–Ј–і–µ–є—Б—В–≤–Є–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ (–≤–Є—В–∞–Љ–Є–љ–∞ –Т8) –љ–∞¬†–њ—А–Њ—Ж–µ—Б—Б—Л —А–Њ—Б—В–∞ —Н–Љ–±—А–Є–Њ–љ–∞ –љ–µ—А–∞–Ј—А—Л–≤–љ–Њ —Б–≤—П–Ј–∞–љ–Њ —Б¬†–∞–Ї—В–Є–≤–љ–Њ—Б—В—М—О —Б–Є–≥–љ–∞–ї—М–љ–Њ–≥–Њ –±–µ–ї–Ї–∞ –њ—А–Њ—В–µ–Є–љ–Ї–Є–љ–∞–Ј—Л –°,¬†–Ї–Њ—В–Њ—А–∞—П –њ–Њ–і–і–µ—А¬≠–ґ–Є–≤–∞–µ—В –њ–µ—А–µ–і–∞—З—Г —Б–Є–≥–љ–∞–ї–∞ –Њ—В¬†–±–µ–ї–Ї–Њ–≤—Л—Е —Д–∞–Ї—В–Њ—А–Њ–≤ —А–Њ—Б—В–∞,¬†–≥–Њ—А–Љ–Њ–љ–Њ–≤ –Є¬†–љ–µ–є—А–Њ—В—А–∞–љ—Б–Љ–Є—В—В–µ—А–Њ–≤ (–њ—А–Њ—Б—В–∞–≥–ї–∞–љ–і–Є–љ–Њ–≤, –∞–і—А–µ–љ–∞–ї–Є–љ–∞, –∞—Ж–µ—В–Є–ї—Е–Њ–ї–Є–љ–∞, —Б–µ—А–Њ—В–Њ–љ–Є–љ–∞, –∞–љ–≥–Є–Њ—В–µ–љ–Ј–Є–љ–∞ –Є¬†–і—А.), —А–µ–≥—Г–ї–Є—А—Г–µ—В –≤–∞–Ј–Њ–і–Є–ї–∞—В–∞—Ж–Є—О –Є¬†–≥–ї–Є–Ї–Њ–ї–Є–Ј –Є¬†–њ—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ–Њ –≤–∞–ґ–љ–∞ –і–ї—П –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ —А–Њ—Б—В–∞ —Н–Љ–±—А–Є–Њ–љ–∞. –Т¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–µ, –њ—А–Њ–≤–µ–і–µ–љ–љ–Њ–Љ –≤–Њ –≤—А–µ–Љ—П –љ–µ–є—А—Г–ї—П—Ж–Є–Є, –±—Л–ї–Њ —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ –њ—А–Њ—В–Є–≤–Њ–і–µ–є—Б—В–≤–Є–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—О –і–µ—Д–µ–Ї—В–Њ–≤ –љ–µ—А–≤–љ–Њ–є —В—А—Г–±–Ї–Є —Б–≤—П–Ј–∞–љ–Њ —Б¬†–∞–Ї—В–Є–≤–љ–Њ—Б—В—М—О –њ—А–Њ—В–µ–Є–љ–Ї–Є–љ–∞–Ј C –±–µ—В–∞ 1 –Є¬†–≥–∞–Љ–Љ–∞ [24].
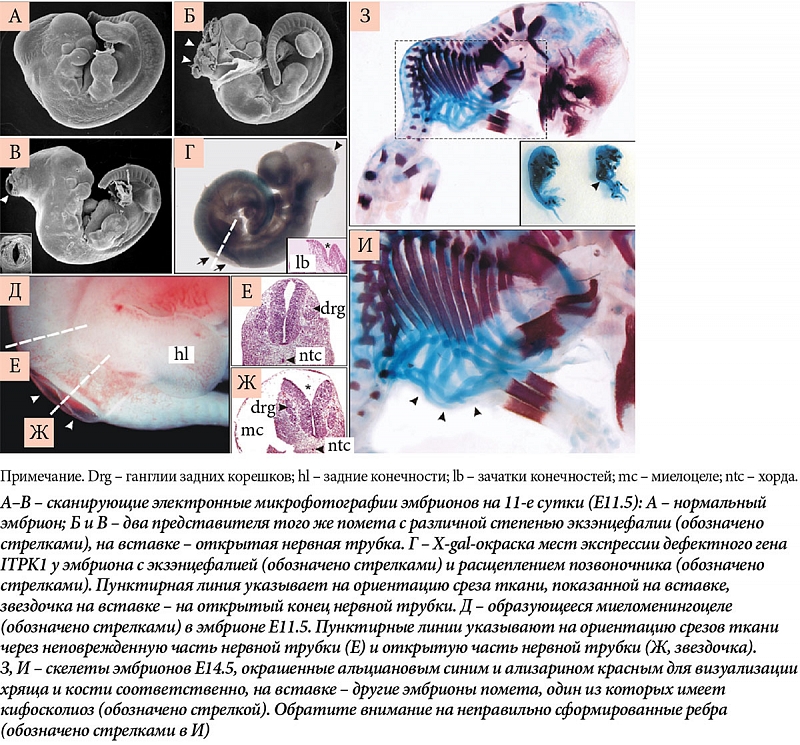
–†–∞—Б—Б–Љ–Њ—В—А–Є–Љ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ¬†–≥–µ–љ–Њ–≤/–±–µ–ї–Ї–Њ–≤, –њ–µ—А–µ—З–Є—Б–ї–µ–љ–љ—Л—Е –≤¬†—В–∞–±–ї–Є—Ж–µ. –Т¬†—З–∞—Б—В–љ–Њ—Б—В–Є, —Д–µ—А–Љ–µ–љ—В –Є–љ–Њ–Ј–Є—В–Њ–ї-1,3,4-—В—А–Є—Д–Њ—Б—Д–∞—В-5/6-–Ї–Є–љ–∞–Ј–∞ (–≥–µ–љ Itpk1)¬†вАУ –Ї–ї—О—З–µ–≤–Њ–є —А–µ–≥—Г–ї—П—В–Њ—А–љ—Л–є —Д–µ—А–Љ–µ–љ—В —Б–Є–љ—В–µ–Ј–∞ —Б–Є–≥–љ–∞–ї—М–љ–Њ–є –Љ–Њ–ї–µ–Ї—Г–ї—Л –Є–љ–Њ–Ј–Є—В–Њ–ї–≥–µ–Ї—Б–∞–Ї–Є—Б—Д–Њ—Б—Д–∞—В–∞ (IP6)¬†вАУ –≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ–Њ–є —Б–Є–≥–љ–∞–ї—М–љ–Њ–є –Љ–Њ–ї–µ–Ї—Г–ї—Л, —Г—З–∞—Б—В–≤—Г—О—Й–µ–є –≤¬†—А–µ–≥—Г–ї—П—Ж–Є–Є –Є–Њ–љ–љ—Л—Е –Ї–∞–љ–∞–ї–Њ–≤, —В—А–∞–љ—Б–њ–Њ—А—В–µ –љ—Г—В—А–Є–µ–љ—В–Њ–≤ –Є¬†—Б—В—А–Њ–Є—В–µ–ї—М–љ—Л—Е –Љ–∞—В–µ—А–Є–∞–ї–Њ–≤ —З–µ—А–µ–Ј –Ї–ї–µ—В–Њ—З–љ—Г—О –Љ–µ–Љ–±—А–∞–љ—Г (—Н–љ–і–Њ—Ж–Є—В–Њ–Ј–µ, —Н–Ї–Ј–Њ—Ж–Є—В–Њ–Ј–µ), —В—А–∞–љ—Б–Ї—А–Є–њ—Ж–Є–Є –Є¬†—А–µ–њ–∞—А–∞—Ж–Є–Є –Ф–Э–Ъ [25]. –Ц–Є–≤–Њ—В–љ—Л–µ —Б¬†–і–µ–ї–µ—Ж–Є–µ–є/–Є–љ–∞–Ї—В–Є–≤–∞—Ж–Є–µ–є¬†–≥–µ–љ–∞ –ґ–Є–Ј–љ–µ—Б–њ–Њ—Б–Њ–±–љ—Л, —Д–µ—А—В–Є–ї—М–љ—Л, –љ–Њ¬†—Г –Є—Е —Н–Љ–±—А–Є–Њ–љ–Њ–≤ —З–∞—Б—В–Њ –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞–ї–Є—Б—М –і–µ—Д–µ–Ї—В—Л –љ–µ—А–≤–љ–Њ–є —В—А—Г–±–Ї–Є, –Њ—Б–µ–≤—Л–µ –і–µ—Д–µ–Ї—В—Л —Б–Ї–µ–ї–µ—В–∞, –Ј–∞–Љ–µ–і–ї–µ–љ–Є–µ —А–Њ—Б—В–∞ –Є¬†–њ–Њ–≤—Л—И–µ–љ–љ–∞—П¬†–≥–Є–±–µ–ї—М –љ–µ–є—А–Њ–љ–Њ–≤ (—А–Є—Б.¬†13) [26]. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–Ј–∞–≤–Є—Б–Є–Љ—Л–є —Д–µ—А–Љ–µ–љ—В Itpk1 –љ–µ–Њ–±—Е–Њ–і–Є–Љ –і–ї—П –∞–і–µ–Ї–≤–∞—В–љ–Њ–≥–Њ —А–∞–Ј–≤–Є—В–Є—П –љ–µ—А–≤–љ–Њ–є —В—А—Г–±–Ї–Є –Є¬†–њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є –і–µ—Д–µ–Ї—В–Њ–≤ –љ–µ—А–≤–љ–Њ–є —В—А—Г–±–Ї–Є (—А–Є—Б.¬†14)¬†[25].
–§–µ—А–Љ–µ–љ—В —Д–Њ—Б—Д–∞—В–Є–і–Є–ї–Є–љ–Њ–Ј–Є—В–Њ–ї-4-—Д–Њ—Б—Д–∞—В-5-–Ї–Є–љ–∞–Ј–∞ (PIP5K,¬†–≥–µ–љ PIP5K1–°) –Ї–∞—В–∞–ї–Є–Ј–Є—А—Г–µ—В —Б–Є–љ—В–µ–Ј –Њ–і–љ–Њ–є –Є–Ј¬†–Њ—Б–љ–Њ–≤–љ—Л—Е –≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ—Л—Е —Б–Є–≥–љ–∞–ї—М–љ—Л—Е –Љ–Њ–ї–µ–Ї—Г–ї¬†вАУ —Д–Њ—Б—Д–∞—В–Є–і–Є–ї–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –і–Є—Д–Њ—Б—Д–∞—В–∞ (PIP2). –Ш–Ј–Њ—Д–Њ—А–Љ–∞ —Д–µ—А–Љ–µ–љ—В–∞ PIP5K-–≥–∞–Љ–Љ–∞ –љ–µ–Њ–±—Е–Њ–і–Є–Љ–∞ –і–ї—П —А–∞–Ј–≤–Є—В–Є—П —Б–µ—А–і–µ—З–љ–Њ-—Б–Њ—Б—Г–і–Є—Б—В–Њ–є –Є¬†—Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ. –¶–µ–ї–µ–љ–∞–њ—А–∞–≤–ї–µ–љ–љ–∞—П –Є–љ–∞–Ї—В–Є–≤–∞—Ж–Є—П¬†–≥–∞–Љ–Љ–∞-–Є–Ј–Њ—Д–µ—А–Љ–µ–љ—В–∞ PIP5K –≤¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–µ –≤—Л–Ј—Л–≤–∞–ї–∞ –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л–µ –љ–∞—А—Г—И–µ–љ–Є—П —А–Њ—Б—В–∞ –Ї–ї–µ—В–Њ–Ї –Є¬†—А–∞–Ј–≤–Є—В–Є—П —В–Ї–∞–љ–µ–є, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –њ—А–Є–≤–Њ–і—П—Й–Є–µ –Ї¬†—Б–µ—А–і–µ—З–љ–Њ–є –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ—Б—В–Є, —Г—Б–Є–ї–µ–љ–Є—О¬†–≥–Є–±–µ–ї–Є –љ–µ–є—А–Њ–љ–Њ–≤ –Є¬†–і–µ—Д–µ–Ї—В–∞–Љ –љ–µ—А–≤–љ–Њ–є —В—А—Г–±–Ї–Є¬†[27].
–Ч–∞–Ї–ї—О—З–µ–љ–Є–µ
–Ш—И–µ–Љ–Є—З–µ—Б–Ї–Є–µ –њ–Њ–≤—А–µ–ґ–і–µ–љ–Є—П —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –њ–ї–Њ–і–∞¬†вАУ –Њ—Б–љ–Њ–≤–љ–∞—П –њ—А–Є—З–Є–љ–∞ –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л —Г¬†–љ–Њ–≤–Њ—А–Њ–ґ–і–µ–љ–љ—Л—Е. –Э–∞¬†—А–∞–љ–љ–Є—Е —Б—А–Њ–Ї–∞—Е –±–µ—А–µ–Љ–µ–љ–љ–Њ—Б—В–Є —Е—А–Њ–љ–Є—З–µ—Б–Ї–∞—П –Є—И–µ–Љ–Є—П —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –њ—А–Є–≤–Њ–і–Є—В –Ї¬†–њ–Њ—А–Њ–Ї–∞–Љ —А–∞–Ј–≤–Є—В–Є—П –Љ–Њ–Ј–≥–∞, –љ–∞¬†–њ–Њ–Ј–і–љ–Є—Е —Б—А–Њ–Ї–∞—Е –Њ–љ–∞ –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ–∞ —Б¬†–≤—Л—Б–Њ–Ї–Є–Љ —А–Є—Б–Ї–Њ–Љ –∞—Б—Д–Є–Ї—Б–Є–Є –≤¬†—А–Њ–і–∞—Е, –і–Є—Б–Ї–Њ–Њ—А–і–Є–љ–∞—Ж–Є–µ–є —А–Њ–і–Њ–≤–Њ–є –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є, –њ–Њ–≤—Л—И–µ–љ–љ—Л–Љ —А–Є—Б–Ї–Њ–Љ –њ–Њ—Б—В–≥–Є–њ–Њ–Ї—Б–Є—З–µ—Б–Ї–Њ–є —Н–љ—Ж–µ—Д–∞–ї–Њ–њ–∞—В–Є–Є¬†вАУ —В–∞–Ї –љ–∞–Ј—Л–≤–∞–µ–Љ–Њ–є –Љ–Є–љ–Є–Љ–∞–ї—М–љ–Њ–є –Љ–Њ–Ј–≥–Њ–≤–Њ–є –і–Є—Б—Д—Г–љ–Ї—Ж–Є–Є –Є¬†–і–µ—В—Б–Ї–Њ–≥–Њ —Ж–µ—А–µ–±—А–∞–ї—М–љ–Њ–≥–Њ –њ–∞—А–∞–ї–Є—З–∞. –•—А–Њ–љ–Є—З–µ—Б–Ї–∞—П –Є—И–µ–Љ–Є—П –Љ–Њ–Ј–≥–∞ –њ–ї–Њ–і–∞ —Г—Б—Г–≥—Г–±–ї—П–µ—В—Б—П –љ–∞¬†—Д–Њ–љ–µ –Є–љ—Б—Г–ї–Є–љ–Њ—А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є –Є¬†–≥–ї—О–Ї–Њ–Ј–Њ—В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В–Є.
–Ь–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї –љ–µ–Њ–±—Е–Њ–і–Є–Љ –і–ї—П —Б–Є–љ—В–µ–Ј–∞ –Є–љ–Њ–Ј–Є—В–Њ–ї—Д–Њ—Б—Д–∞—В–Њ–≤ –Є¬†—Д–Њ—Б—Д–∞—В–Є–і–Є–ї–Є–љ–Њ–Ј–Є—В–Њ–ї–Њ–≤—Л—Е –ї–Є–њ–Є–і–Њ–≤, –Ї–Њ—В–Њ—А—Л–µ –Њ–њ–Њ—Б—А–µ–і—Г—О—В –њ–µ—А–µ–і–∞—З—Г —Б–Є–≥–љ–∞–ї–∞ –Њ—В¬†—А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ —Д–∞–Ї—В–Њ—А–Њ–≤ —А–Њ—Б—В–∞ –Є¬†–љ–µ–є—А–Њ—В—А–∞–љ—Б–Љ–Є—В—В–µ—А–Њ–≤ –≤–љ—Г—В—А—М –Ї–ї–µ—В–Ї–Є. –≠—В–Є –њ—А–Њ–Є–Ј–≤–Њ–і–љ—Л–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –Ї—А–∞–є–љ–µ –≤–∞–ґ–љ—Л –і–ї—П —А–∞–Ј–≤–Є–≤–∞—О—Й–µ–≥–Њ—Б—П –Љ–Њ–Ј–≥–∞, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—В –∞–і–µ–Ї–≤–∞—В–љ—Г—О –Ї–Њ–Љ–Љ—Г–љ–Є–Ї–∞—Ж–Є—О –Љ–µ–ґ–і—Г –љ–µ–є—А–Њ–љ–∞–Љ–Є, —Б–љ–Є–ґ–∞—О—В —Е—А–Њ–љ–Є—З–µ—Б–Ї—Г—О –Є—И–µ–Љ–Є—О –љ–µ–є—А–Њ–љ–Њ–≤ –Є¬†–Ј–∞—Й–Є—Й–∞—О—В –Њ—В¬†–љ–µ–≥–∞—В–Є–≤–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤¬†–≥–ї—О–Ї–Њ–Ј–Њ—В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В–Є.
–Я—А–µ–і—Б—В–∞–≤–ї–µ–љ–љ—Л–µ –≤¬†—А–∞–±–Њ—В–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л –њ–Њ–і—В–≤–µ—А–ґ–і–∞—О—В –≤—Л—А–∞–ґ–µ–љ–љ–Њ–µ –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –љ–∞¬†—А–Њ—Б—В –љ–µ–є—А–Њ–љ–Њ–≤ –≤¬†–Ї—Г–ї—М—В—Г—А–µ –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е¬†–≥–ї—Г—В–∞–Љ–∞—В–љ–Њ–≥–Њ —Б—В—А–µ—Б—Б–∞ (–њ–Њ–≤—Л—И–µ–љ–Є–µ –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В–Є –љ–µ–є—А–Њ–љ–Њ–≤ –≤¬†—Б—А–µ–і–љ–µ–Љ –љ–∞¬†17%). –Я—А—П–Љ–Њ–µ –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–µ –і–µ–є—Б—В–≤–Є–µ –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ —Г–Ї–∞–Ј—Л–≤–∞–µ—В –љ–∞¬†–Ј–љ–∞—З–Є–Љ–Њ—Б—В—М –µ–≥–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Ї–∞–Ї –і–ї—П –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є –њ–Њ—А–Њ–Ї–Њ–≤ —А–∞–Ј–≤–Є—В–Є—П, –≤–Њ–Ј–љ–Є–Ї–∞—О—Й–Є—Е –љ–∞¬†—А–∞–љ–љ–Є—Е —Б—А–Њ–Ї–∞—Е, —В–∞–Ї –Є¬†–і–ї—П –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—Ж–Є–Є –Љ–Њ–Ј–≥–∞ –њ–ї–Њ–і–∞ –љ–∞¬†–њ–Њ–Ј–і–љ–Є—Е —Б—А–Њ–Ї–∞—Е¬†–≥–µ—Б—В–∞—Ж–Є–Є. –Я–Њ–і—З–µ—А–Ї–љ–µ–Љ, —З—В–Њ –≤¬†–і–Њ—В–∞—Ж–Є—П—Е –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ –Њ—Б–Њ–±–µ–љ–љ–Њ –љ—Г–ґ–і–∞—О—В—Б—П –±–µ—А–µ–Љ–µ–љ–љ—Л–µ, —А–∞—Ж–Є–Њ–љ –Ї–Њ—В–Њ—А—Л—Е –њ–µ—А–µ–≥—А—Г–ґ–µ–љ —Г–≥–ї–µ–≤–Њ–і–∞–Љ–Є, –ґ–µ–љ—Й–Є–љ—Л —Б¬†–і–Є–∞–±–µ—В–Њ–Љ (–≤¬†—В–Њ–Љ —З–Є—Б–ї–µ¬†–≥–µ—Б—В–∞—Ж–Є–Њ–љ–љ—Л–Љ), –ґ–µ–љ—Й–Є–љ—Л, —А–∞–љ–µ–µ —А–Њ–і–Є–≤—И–Є–µ —А–µ–±–µ–љ–Ї–∞ —Б¬†–њ–Њ—А–Њ–Ї–Њ–Љ —А–∞–Ј–≤–Є—В–Є—П.
–°—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ–µ –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–Њ –Љ–Є–Ї—А–Њ–љ—Г—В—А–Є–µ–љ—В–љ–Њ–є –њ–Њ–і–і–µ—А–ґ–Ї–Є –±–µ—А–µ–Љ–µ–љ–љ–Њ—Б—В–Є –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–Њ–Љ¬†вАУ –≤—Л—Б–Њ–Ї–∞—П –±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В—М –µ–≥–Њ –њ—А–Є–Љ–µ–љ–µ–љ–Є—П. –Ф–∞–ґ–µ –Ї—А–∞–є–љ–µ –≤—Л—Б–Њ–Ї–Є–µ –і–Њ–Ј—Л –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї–∞ (12¬†–≥/—Б—Г—В) –≤—Л–Ј—Л–≤–∞—О—В —В–Њ–ї—М–Ї–Њ –ї–µ–≥–Ї–Є–µ –њ–Њ–±–Њ—З–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л —Б–Њ¬†—Б—В–Њ—А–Њ–љ—Л –ґ–µ–ї—Г–і–Њ—З–љ–Њ-–Ї–Є—И–µ—З–љ–Њ–≥–Њ —В—А–∞–Ї—В–∞ (—В–Њ—И–љ–Њ—В—Г, –Љ–µ—В–µ–Њ—А–Є–Ј–Љ, –і–Є–∞—А–µ—О) [28], –≤¬†—В–Њ –≤—А–µ–Љ—П –Ї–∞–Ї –≤¬†—А–µ–∞–ї—М–љ–Њ–є –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Њ–є –њ—А–∞–Ї—В–Є–Ї–µ –і–∞–ґ–µ –≤¬†–≥–Њ—А–∞–Ј–і–Њ –Љ–µ–љ—М—И–Є—Е –і–Њ–Ј–∞—Е (0,5вАУ4¬†–≥/—Б—Г—В), —Н—Д—Д–µ–Ї—В–Є–≤–љ—Л—Е –Є¬†–њ–Њ–ї–љ–Њ—Б—В—М—О –±–µ–Ј–Њ–њ–∞—Б–љ—Л—Е, –Љ–Є–Њ–Є–љ–Њ–Ј–Є—В–Њ–ї –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ –≤–ї–Є—П–µ—В –љ–∞¬†—А–∞–Ј–≤–Є—В–Є–µ –Љ–Њ–Ј–≥–∞ –њ–ї–Њ–і–∞.
–†–∞–±–Њ—В–∞ –≤—Л–њ–Њ–ї–љ–µ–љ–∞ –њ—А–Є –њ–Њ–і–і–µ—А–ґ–Ї–µ –†–§–§–Ш (–≥—А–∞–љ—В—Л 19-07-00356, 17-07-01419)
I.Yu. Torshin, PhD, O.A. Gromova, MD, PhD, Prof., A.G. Kalacheva, PhD, N.K. Tetruashvili, MD, PhD, E.V. Stelmashook, DBSci, PhD, L.G. Khaspekov, DBSci, PhD
Federal Research Center вАШComputer Science and ControlвАЩ of the Russian Academy of Sciences, Moscow
Ivanovo State Medical Academy
National Medical Research Center for Obstetrics, Gynecology and Perinatology named after Academician V.I. Kulakov, Moscow
Scientific Center of Neurology, Moscow
Contact person: Olga A. Gromova, unesco.gromova@gmail.com
Myo-inositol is the basis for the synthesis of an important group of signal molecules, inositolphosphates, which mediate signal transmission from receptors of growth factors and neurotransmitters. Grants myo-inositol promotes the prevention of folate-resistant defects and neuroprotection of the fetal brain ischemia. The paper presents the results of a study of the effects of myo-inositol on the growth of cerebellar neurons in culture under glutamate stress. It is shown that the effects of myo-inositol on the survival of neurons (17%) exceed the effects of drugs that are usually used for neuroprotection (peptide extracts вАУ 10%, choline preparations вАУ no more than 3%). Confirmed in the present work, a direct neuroprotective effect of myo-inositol indicates the importance of the use of myo-inositol during pregnancy with the aim of neuroprotection of the fetal brain.
–£–≤–∞–ґ–∞–µ–Љ—Л–є –њ–Њ—Б–µ—В–Є—В–µ–ї—М uMEDp!
–£–≤–µ–і–Њ–Љ–ї—П–µ–Љ –Т–∞—Б –Њ —В–Њ–Љ, —З—В–Њ –Ј–і–µ—Б—М —Б–Њ–і–µ—А–ґ–Є—В—Б—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П, –њ—А–µ–і–љ–∞–Ј–љ–∞—З–µ–љ–љ–∞—П –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –і–ї—П —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П.
–Х—Б–ї–Є –Т—Л –љ–µ —П–≤–ї—П–µ—В–µ—Б—М —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–Љ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П, –∞–і–Љ–Є–љ–Є—Б—В—А–∞—Ж–Є—П –љ–µ –љ–µ—Б–µ—В –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ—Б—В–Є –Ј–∞ –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ –њ–Њ—Б–ї–µ–і—Б—В–≤–Є—П, –≤–Њ–Ј–љ–Є–Ї—И–Є–µ –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–∞–Љ–Њ—Б—В–Њ—П—В–µ–ї—М–љ–Њ–≥–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Т–∞–Љ–Є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є —Б –њ–Њ—А—В–∞–ї–∞ –±–µ–Ј –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є –Ї–Њ–љ—Б—Г–ї—М—В–∞—Ж–Є–Є —Б –≤—А–∞—З–Њ–Љ.
–Э–∞–ґ–Є–Љ–∞—П –љ–∞ –Ї–љ–Њ–њ–Ї—Г ¬Ђ–Т–Њ–є—В–Є¬ї, –Т—Л –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В–µ, —З—В–Њ —П–≤–ї—П–µ—В–µ—Б—М –≤—А–∞—З–Њ–Љ –Є–ї–Є —Б—В—Г–і–µ–љ—В–Њ–Љ –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–≥–Њ –≤—Г–Ј–∞.