Нейрохимические механизмы регуляции сна и бодрствования: роль блокаторов гистаминовых рецепторов в лечении инсомнии
- Аннотация
- Статья
- Ссылки
- English
Открытие Н. Mougoun и D. Morruzzi в конце 1940-х гг. ретикулярной формации ствола головного мозга подтвердило, что нормальное функционирование таламо-кортикальной системы мозга, обеспечивающее весь спектр сознательной деятельности человека, возможно только при наличии мощных тонических воздействий со стороны определенных подкорковых структур, называемых активирующими. Прямое изучение нейронов, вовлеченных в регуляцию цикла «сон – бодрствование», проведенное во второй половине минувшего столетия, показало, что благодаря этим воздействиям мембрана большинства кортикальных нейронов в бодрствовании деполяризована на 5–10 мВ по сравнению с потенциалом покоя (-68/-70 мВ). Только в таком состоянии тонической деполяризации эти нейроны способны обрабатывать информацию и отвечать на сигналы, приходящие к ним от других нервных клеток, как рецепторных, так и внутримозговых. Таких систем тонической деполяризации, или восходящей активации мозга (их можно условно назвать «центрами бодрствования»), несколько – по-видимому, не менее десяти; они расположены на всех уровнях мозговой оси и выделяют различные химические медиаторы. Детали строения и функционирования этих систем подробно изложены в ряде недавних русскоязычных обзоров [1–5].
Особую роль в поддержании бодрствования играет гистаминергическая система, расположенная в туберомамиллярном (бугорково-сосцевидном) ядре заднего гипоталамуса [6–9]. По методическим причинам точная локализация этой системы и ее проекций в головном мозге крыс была описана позже, чем систем других мозговых аминов, – лишь в 1983–1984 гг. – японскими и американскими авторами. А в 1988 г. в лаборатории M. Jouvet был обнаружен гистаминергический «arousal-механизм» в гипоталамусе кошки [10].
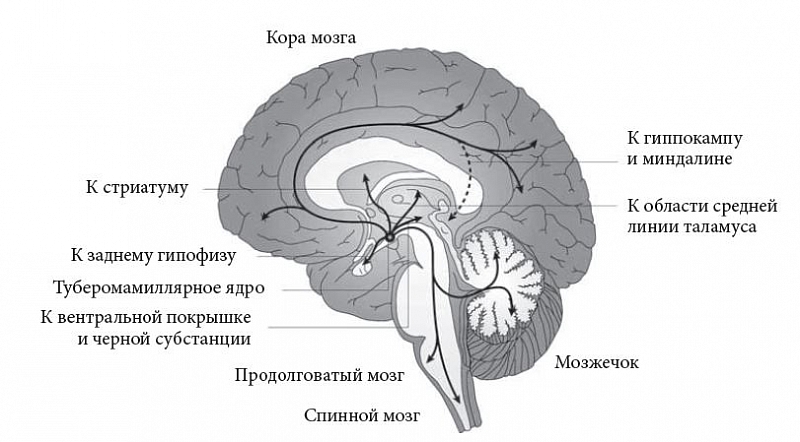
Туберомамиллярное ядро – это единственный источник гистамина в головном мозге позвоночных. Подобно большинству других активирующих систем, гистаминергическая система туберомамиллярного ядра устроена по «древовидному» принципу: очень небольшое количество нейронов (в мозге крысы – лишь 3–4 тыс., в мозге человека – 64 тыс.) иннервирует миллиарды клеток новой, древней коры и подкорковых структур за счет колоссального ветвления своих аксонов (каждый аксон образует сотни тысяч ответвлений). Наиболее мощные восходящие проекции направляются в нейрогипофиз, близлежащие дофаминсодержащие области вентральной покрышки среднего мозга и компактной части черной субстанции, базальную область переднего мозга (крупноклеточные ядра безымянной субстанции, содержащие ацетилхолин и гамма-аминомасляную кислоту (ГАМК)), стриатум, неокортекс, гиппокамп, миндалину и таламические ядра средней линии, а нисходящие – в мозжечок, продолговатый и спинной мозг.
Тщательные нейроморфологические исследования, проведенные в лаборатории M. Jouvet и в других лабораториях на мозге кошки и крысы, показали, что гистаминергические нейроны туберомамиллярного ядра проецируются также на латеродорзальное и педункулопонтинное ядра мезопонтинной покрышки, выделяющие ацетилхолин, синее (голубое) пятно, продуцирующее норадреналин, на дорзальные ядра шва, синтезирующие серотонин (рис. 1) [11]. В свою очередь гистаминергические нейроны туберомамиллярного ядра получают афференты от инфралимбической коры, латеральной области перегородки, преоптической области переднего гипоталамуса, адренергических клеток С1–С3, норадренергических нейронов А1–А3 и серотонинергических клеток В5–В9. Интересно, что лишь одиночные волокна достигают туберомамиллярного ядра от норадреналинсодержащих клеток синего пятна и дофаминсодержащих нейронов вентральной покрышки среднего мозга и компактной части черной субстанции [7]. Однако при болезни Паркинсона, связанной с разрушением дофаминергической передачи, наблюдается двукратное повышение концентрации гистамина, поступающего из туберомамиллярного ядра в компактную часть черной субстанции и ее проекцию – бледный шар.
Исключительно важны взаимосвязи между гистаминергической и орексин/гипокретинергической системами мозга. Медиаторы этих двух систем действуют синергично, играя уникальную роль в поддержании бодрствования. Орексин/гипокретинсодержащие нейроны располагаются в заднелатеральном гипоталамусе и перифорникальной области, в непосредственной близости от гистаминовых нейронов туберомамиллярного ядра. Оба ядра частично перекрываются и образуют функциональное единство. Оба пептида орексина (гипокретина) возбуждают гистаминовые нейроны через свои рецепторы 2-го типа и активацию натрий-кальциевого ионного обмена. Орексин часто солокализуется с динорфином, который также может участвовать в возбуждении гистаминергических нейронов путем подавления ГАМКергического тормозного пути. Однако гистаминовые нейроны, по-видимому, не влияют на возбудимость орексиновых нейронов, так что взаимодействие этих двух систем носит односторонний характер [12].
Таким образом, можно сказать, что гистаминергическая и другие аминергические системы межуточного, среднего мозга и ствола обладают весьма значительным сходством в своей морфологии, клеточной и системной физиологии. Обладая множественными взаимными связями, они формируют самоорганизующуюся сеть, своего рода «оркестр», в котором орексиновые (гипокретиновые) нейроны играют роль дирижера, а гистаминовые – первой скрипки [9].
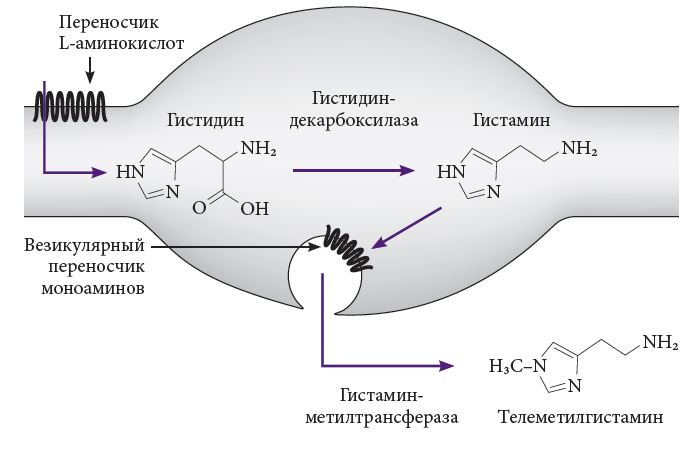
Как известно, гистамин образуется из аминокислоты гистидина, поступающей в организм с белковой пищей. В отличие от гистамина, гистидин проходит гематоэнцефалический барьер и захватывается белком-транспортером аминокислот, переносящим его внутрь тела нейрона или варикозного расширения аксона (рис. 2). Нейроны туберомамиллярного ядра экспрессируют фермент гистидиндекарбоксилазу (ГДК), отщепляющий от молекулы гистидина карбоксил и превращающий ее в гистамин. ГДК наиболее активен в соме, но проявляется также и в варикозных расширениях, и в нервных окончаниях. Фактором ограничения скорости синтеза гистамина является тканевая концентрация его предшественника, гистидина. Гистамин переносится в везикулы с помощью особого белка, называемого везикулярным моноаминным транспортером 2-го типа (ВМАТ-2), и там накапливается. Эти везикулы располагаются не только в клеточных телах, но и в варикозных расширениях аксонов. При возникновении потенциала действия гистамин выделяется Ca++-зависимым путем. Выделившийся в синаптическую щель или межклеточное пространство гистамин, не связавшийся с рецептором, инактивируется путем метилирования с помощью фермента гистаминметилтрансферазы (синтезируемого постсинаптически или в глии), превращающей его в телеметилгистамин. Последний подвергается окислительному деаминированию с помощью фермента моноаминооксидазы типа Б (МАО-Б), превращаясь в t-метил-имидазолуксусную кислоту. Механизма обратного захвата для гистамина не существует. Обычно период полужизни нейронального гистамина составляет около получаса, но он может резко укорачиваться под воздействием внешних факторов, например стресса [7, 13].
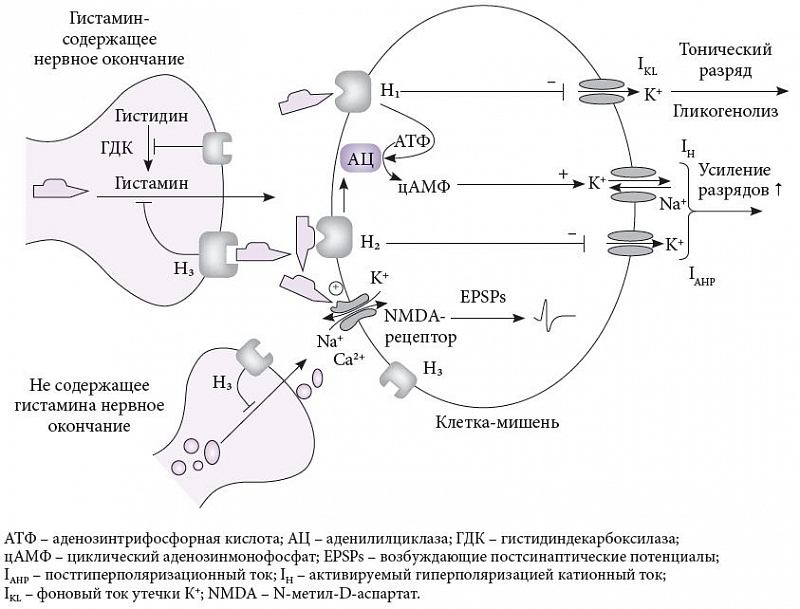
Известно 4 типа метаботропных рецепторов гистамина, связанных с G-белками: два возбудительных (Н₁ и Н₂) и два тормозных (Н₃ и Н₄). Рецепторы Н₁–Н₃ повсеместно представлены в головном мозге, а рецептор Н₄ – главным образом в спинном. Активация постсинаптических рецепторов Н₁ или Н₂, расположенных на соме нейронов-мишеней, запускает внутриклеточные молекулярные каскады, связанные с аденозинтрифосфатом, аденилилциклазой и циклическим аденозинмонофосфатом, повышает клеточную активность и возбудимость, либо снижая фоновый ток утечки К+ (IKL) или постгиперполяризационный ток (IAHP), либо повышая активируемый гиперполяризацией катионный ток (IH). Гистамин также взаимодействует с полиаминной областью NMDA (N-метил-D-аспартатного) рецептора, модулируя возбудительные постсинаптические потенциалы (excitatory post-synaptic potential, EPSPs) (рис. 3) [14]. Пресинаптические ауто- и гетерорецепторы типа Н₃ могут располагаться на соме, аксонах и дендритах, тормозя синтез и выделение гистамина и других передатчиков. Что касается Н₃ постсинаптических рецепторов, находящихся на соме клеток-мишеней, то, например, в стриатуме они встречаются часто в паре с дофаминовыми рецепторами D₂, понижая их сродство с лигандами. Интересным свойством Н₃-рецепторов является их высокая конститутивная активность, то есть спонтанная активность в отсутствие гистамина, которая играет важную роль в головном мозге. Н₃-рецепторы участвуют в регуляции сна – бодрствования и когнитивных функций путем модуляции выброса или синтеза гистамина и других нейропередатчиков. Уже несколько обратимых агонистов Н₃-рецепторов, способных блокировать их спонтанную активность, проходят клинические испытания у больных шизофренией, эпилепсией, нарколепсией, ожирением и болезнью Альцгеймера. Другим отличительным свойством является множественность изоформ этого рецептора, происходящих из общего гена и образующихся за счет альтернативного сплайсинга [13].
Интересно, что кроме нейрональных, тучных и микроглиальных клеток гистаминдекарбоксилазу экспрессируют клетки эпендимы головного мозга. Этот гистамин может быть вовлечен в регуляцию образования стволовых клеток, расположенных под эпендимальным слоем. Нейрональные стволовые клетки in vitro реагируют на лиганды рецепторов Н₁ и Н₂ [13].
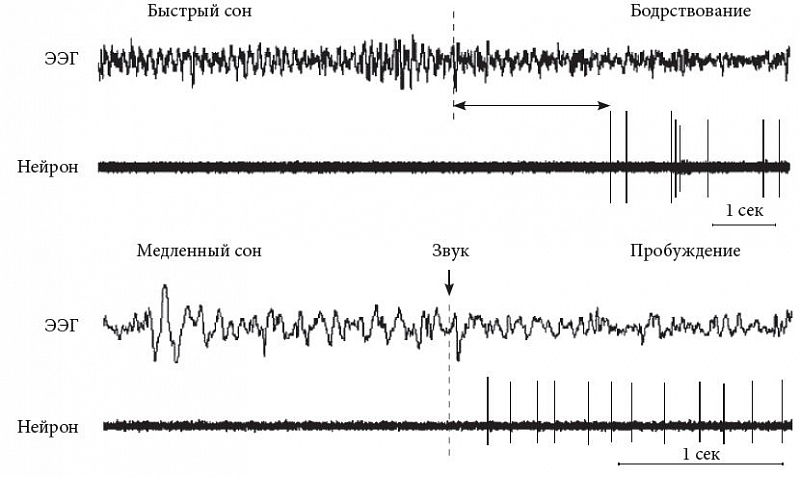
Гистаминергические нейроны являются ритмоводителями и демонстрируют регулярные спонтанные низкочастотные разряды (1–4 Гц). При пробуждении и поведенческой активации их частота возрастает, при засыпании и медленном сне – снижается, при быстром сне – исчезает (рис. 4). Торможение гистаминергических нейронов во сне опосредуется ГАМКергическими нейронами «центра сна» в вентро-латеральной преоптической области (ventrolateral preoptic nucleus, VLPO) [13]. Кроме того, на нейроны туберомамиллярного ядра оказывают воздействие тормозные нейропептиды – галанин и эндоморфин. Гистаминергических рецепторов на клетках VLPO нет, так что взаимодействие этих двух систем – гистаминергического «центра бодрствования» туберомамиллярного ядра и ГАМКергического «центра сна» вентро-латеральной преоптической области – носит односторонний характер. Считается, что такое взаимодействие гистаминовой системы с активирующей (орексинергической) и тормозной (ГАМКергической) придает дополнительную устойчивость всему механизму.
Нейрональный гистамин участвует во множестве функций мозга: поддержании гомеостаза мозговой ткани, регуляции некоторых нейроэндокринных функций, поведения, биоритмов, репродукции, температуры и массы тела, энергетического обмена и водного баланса, в реакции на стресс. Кроме поддержания бодрствования, мозговой гистамин участвует в сенсорных и моторных реакциях, регуляции эмоциональности, обучении и памяти [13].
Впервые предположения о том, что гистамин является «гормоном бодрствования», появились после того, как были обнаружены побочные снотворные эффекты первого поколения антигистаминных препаратов, проходящих гематоэнцефалический барьер (дифенгидрамин). Далее было обнаружено, что нейроны туберомамиллярного ядра активны только в состоянии бодрствования, но не сна. Наконец, воздействие на гистаминергическую систему путем либо введения антагонистов Н₃-рецепторов, которые ее активируют, либо введения альфа-фторметилгистидина, блокирующего синтез гистамина, либо удаления гена ГДК нарушает цикл бодрствования – сна у нокаутных мышей [13]. Из всех ныне известных нейронных систем именно гистаминергическая наиболее чувствительна к изменению уровня бодрствования
[8, 15].
У мышей, нокаутных по гену ГДК, отмечается увеличение доли представленности быстрого сна, снижение мощности тета-ритма в бодрствовании и дельта-ритма – в медленном сне, снижение представленности бодрствования в темное время суток и повышенная сонливость. Последняя проявляется снижением как реактивности (латентности ко сну после воздействия на животное пробуждающими стимулами: выключением света или помещением мыши в новую клетку), так и активности в темный период суток по сравнению с контрольными мышами (wild-type). Мыши, нокаутные по гену ГДК или по гену Н₁-рецептора, более активны в дневное время, чем мыши из группы контроля. Взаимодействуя с ГАМКергической, гистаминергическая система тормозит поведенческие проявления сенситизации (понятие, обратное толерантности), вызванной хроническим введением метамфетамина [16, 17].
Гистаминергическая система играет важную роль в формировании нарколептического фенотипа. Хотя само заболевание связано с недостаточностью орексиновой передачи, в опытах на собаках-«нарколептиках» было показано, что во время катаплексических приступов активность гистаминергических нейронов сохраняется, в то время как серотонинергических – резко снижается, а норадренергических – вовсе прекращается [18]. При этом антагонисты Н₃-рецепторов снижают избыточную сонливость и частоту катаплексических приступов, блокируя, по-видимому, тормозные ауторецепторы, обеспечивающие отрицательную обратную связь, что приводит к увеличению выброса гистамина в синаптические щели. Уже несколько веществ такого рода проходят клинические испытания в качестве лекарственных средств для лечения нарколепсии [14].
Кроме этого, модуляция гистаминергической системы может быть использована для лечения и других нарушений цикла «сон – бодрствование». Так, трициклический антидепрессант доксепин не только тормозит обратный захват норадреналина и серотонина, но и является антагонистом рецепторов Н₁ и Н₂ и, вследствие этого, с успехом применяется для лечения инсомнии у пожилых больных. Наоборот, избыточная сонливость может быть подавлена введением антагонистов Н₃-рецепторов [14].
Инсомния является самым распространенным нарушением сна и, по разным данным, представляет серьезную проблему для 12–22% населения. Фармакотерапия нарушений сна, несмотря на большое количество снотворных препаратов, остается сложной задачей. В целом гипнотики можно разделить на pro S (pro sleep) – для сна – и anti W (anti wake) – против бодрствования [19, 20]. В настоящее время большинство снотворных препаратов относят к pro sleep группе. Они в основном представлены снотворными средствами, воздействующими на постсинаптический ГАМКергический комплекс. Сюда входят барбитураты, бензодиазепины и Z-препараты (зопиклон, золпидем и залеплон). В этой связи представляется интересным проанализировать существующий опыт использования в качестве гипнотика типичного anti W препарата – доксиламина сукцината (Донормил).
Доксиламина сукцинат является антагонистом Н₁-гистаминовых рецепторов, с временем достижения максимальной концентрации в плазме, равным 2 часам, и периодом полувыведения 10 часов. Примерно 60% доксиламина выводится с мочой в неизмененном виде, а его метаболиты неактивны.
Был проведен ряд исследований по применению этого препарата в качестве снотворного средства. В одном из них, рандомизированном двойном слепом с 3 параллельными группами, сравнивалась эффективность и переносимость доксиламина сукцината (15 мг) с золпидемом (10 мг) и плацебо при лечении инсомнии (с изучением синдрома отмены) [21]. В исследовании приняли участие 338 пациентов в возрасте от 18 до 73 лет. С одной стороны, результаты подтвердили превосходство доксиламина над плацебо в отношении снотворного действия, с другой – показали сопоставимую эффективность доксиламина и золпидема, подтвердили хорошую переносимость доксиламина и золпидема и не выявили синдрома отмены при прекращении приема доксиламина.
Нами было проведено открытое несравнительное исследование препарата Донормил с целью оценки его эффективности и безопасности у больных инсомнией [22]. С помощью анкетного метода, психологических тестов, полисомнографии было обследовано 50 больных инсомнией в возрасте от 24 до 60 лет (средний возраст 56,6 лет) до и после 10-дневного курса лечения Донормилом (15 мг на ночь). Результаты исследования показали, что после терапии Донормилом улучшились такие субъективные характеристики сна, как продолжительность сна, качество сна и утреннего пробуждения, снизилось количество ночных пробуждений и длительность засыпания. Объективные полисомнографические исследования подтвердили «субъективную» эффективность Донормила, о чем свидетельствуют: уменьшение длительности засыпания; увеличение длительности сна; увеличение времени фазы быстрого сна; улучшение индекса качества сна. Переносимость терапии была хорошей – ни один пациент не отметил каких-либо побочных эффектов, все пациенты полностью прошли планируемый курс лечения. Кроме того, на фоне лечения Донормилом не было отмечено ухудшения течения сопутствующих соматических и неврологических заболеваний.
В рандомизированном двойном слепом перекрестном плацебоконтролируемом исследовании изучалось влияние доксиламина на структуру сна и состояние когнитивных функций, памяти и скорости реакций при однократном приеме 15 мг доксиламина сукцината или плацебо у здоровых добровольцев [23]. В ходе исследования было выявлено, что показатели общей продолжительности сна, числа пробуждений во время сна, длительности четвертой фазы первого цикла сна и парадоксальной фазы первого цикла сна, количество циклов сна (смены фаз глубокого и парадоксального сна) в группе доксиламина и плацебо не отличались. После приема доксиламина значительно снижалась общая продолжительность пробуждений во время сна. Прием доксиламина приводил к значительному укорочению первой стадии и удлинению второй стадии. При этом доксиламин не влиял на продолжительность третьей и четвертой стадий и фазы быстрого сна. Более подробный анализ показал, что после приема доксиламина в сравнении с плацебо значительно улучшалось качество и увеличивалась глубина сна, тогда как ясность сознания и состояние при пробуждении при приеме препарата и плацебо не отличались. При приеме доксиламина ни у одного из 18 испытуемых не отмечено каких-либо изменений кратковременной памяти и скорости реакций. При проведении данного исследования не наблюдалось каких-либо нежелательных явлений.
Результаты проведенных исследований свидетельствуют об эффективности и безопасности использования Донормила при нарушениях сна и широких возможностях его применения. Отсутствие эффекта последействия и, что немаловажно, доступность препарата позволяют применять Донормил для ситуационного приема в условиях стресса при развитии адаптационной формы инсомнии. По данным Американской ассоциации медицины сна (American Academy of Sleep Medicine), 20% населения испытывают острую инсомнию по крайней мере раз в год, а 90% – по крайней мере раз в жизни. Персистирование стресса и связанные с ним нарушения сна являются ведущим фактором хронификации инсомнии. Проведенные нами исследования показали, что разовый прием снотворных средств после острого эмоционального стресса полностью нивелировал негативное влияние последнего на структуру сна, а также отмеченное после стресса увеличение частоты сердечных сокращений и вариабельности сердечного ритма. Более качественный сон приводил к улучшению показателей бодрствования (по результатам тестов для оценки внимания, памяти, тонкой моторики рук) [24]. Таким образом, коррекция параметров ночного сна в условиях стресса нейтрализует негативные воздействия стресса, повышает адаптивные возможности человека и является профилактикой развития хронической инсомнии.
Еще одним важным показанием к применению Донормила представляется инсомния на фоне лекарственной зависимости. Наиболее частой причиной возникновения данной формы инсомнии является длительный прием снотворных средств, воздействующих на постсинаптический ГАМКергический комплекс, в большинстве случаев бензодиазепинов. При данном расстройстве один из этапов лечения – перевод пациента на снотворный препарат другой химической группы, в качестве которого с успехом может быть использован Донормил, так как он является антагонистом рецепторов другого нейромедиатора. Следует отметить, что Донормил можно использовать во время беременности, по результатам адекватных и хорошо контролируемых исследований [25].
Между тем Донормил, не обладающий миорелаксирующим действием, с осторожностью должен применяться в случаях подозрения на наличие синдрома обструктивного апноэ сна, что особенно актуально у пожилых больных, так как у данной категории пациентов эта болезнь часто протекает субклинически и может быть диагностирована только при проведении полисомнографических исследований [25].
Таким образом, многообразие систем, участвующих в организации состояний сна и бодрствования, позволяет успешно использовать для коррекции инсомнии препараты, воздействующие на различные рецепторные комплексы, в том числе входящие в состав одной из наиболее важных активирующих систем головного мозга – гистаминергической.
Neurochemical mechanisms of the regulation of the sleep-wake cycle and wakefulness: role of histamine receptors antihistamines in the treatment of insomnia
V.M. Kovalzon1, K.N. Strygin2
1 Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences
2 I.M. Sechenov First Moscow State Medical University
Contact person: Vladimir Matveyevich Kovalzon, kovalzon@sevin.ru
The authors review structure, morphological and neurochemical chains of the brain histaminergic system, one of the main wakefulness promoting systems. Biochemistry of the histamine turnover and characteristics of histamine receptors are summarized. The authors focuse on the interactions between the histaminergic and orexinergic (hypocretinergic) systems. Examples of effects of experimental modulation of the histaminergic system on the sleep-wake cycle are presented. The authors deduce that orexine neurons coordinate wakefulness promoting system primarily through their effects on histamine neurons. Uses of central H₁-antihistamines for the correction of varied forms of insomnia were discussed.
Уважаемый посетитель uMEDp!
Уведомляем Вас о том, что здесь содержится информация, предназначенная исключительно для специалистов здравоохранения.
Если Вы не являетесь специалистом здравоохранения, администрация не несет ответственности за возможные отрицательные последствия, возникшие в результате самостоятельного использования Вами информации с портала без предварительной консультации с врачом.
Нажимая на кнопку «Войти», Вы подтверждаете, что являетесь врачом или студентом медицинского вуза.