–†–į–Ī–ĺ—ā–į –Ņ–ĺ–ī–ī–Ķ—Ä–∂–į–Ĺ–į –≥—Ä–į–Ĺ—ā–į–ľ–ł –†–§–§–ė ‚ĄĖ¬†13-04-00941 –ł¬†13-04-00741.
–ú–Ķ–ī–ł—Ü–ł–Ĺ—Ā–ļ–ł–Ļ –Ņ–ĺ—Ä—ā–į–Ľ –ī–Ľ—Ź –≤—Ä–į—á–Ķ–Ļ
–ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–ĺ —Ā—ā–į—ā–Ķ–Ļ
7497
–ó–į–≥—Ä—É–∑–ļ–į...
–ü–ĺ–∂–į–Ľ—É–Ļ—Ā—ā–į, –į–≤—ā–ĺ—Ä–ł–∑—É–Ļ—ā–Ķ—Ā—Ć:
–ě–Ī–∑–ĺ—Ä—č
–ź—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–Ĺ–ĺ–Ķ —Ä–į–∑–≤–ł—ā–ł–Ķ —Ā–Ĺ–į –ļ–į–ļ –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–į—Ź –Ņ—Ä–ł—á–ł–Ĺ–į —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł—Ź –ļ–ĺ–≥–Ĺ–ł—ā–ł–≤–Ĺ—č—Ö —Ą—É–Ĺ–ļ—Ü–ł–Ļ –ł –≤–ĺ–∑–Ĺ–ł–ļ–Ĺ–ĺ–≤–Ķ–Ĺ–ł—Ź —Ä—Ź–ī–į –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ļ, —Ā–≤—Ź–∑–į–Ĺ–Ĺ—č—Ö —Ā —Ü–ł–ļ–Ľ–ĺ–ľ ¬ę—Ā–ĺ–Ĺ ‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ
"–≠–§–§–ē–ö–Ę–ė–í–Ě–ź–Į –§–ź–†–ú–ź–ö–ě–Ę–ē–†–ź–ü–ė–Į. –Ě–Ķ–≤—Ä–ĺ–Ľ–ĺ–≥–ł—Ź —Ā–Ņ–Ķ—Ü–≤—č–Ņ—É—Ā–ļ (22)
- –ź–Ĺ–Ĺ–ĺ—ā–į—Ü–ł—Ź
- –°—ā–į—ā—Ć—Ź
- –°—Ā—č–Ľ–ļ–ł
- English
–í —Ā—ā–į—ā—Ć–Ķ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ –ļ—Ä–į—ā–ļ–ł–Ļ –ĺ–Ī–∑–ĺ—Ä —ā–Ķ–ĺ—Ä–ł–Ļ, —Ä–į—Ā—Ā–ľ–į—ā—Ä–ł–≤–į—é—Č–ł—Ö —Ā–ĺ–Ĺ –ļ–į–ļ –Ņ–Ķ—Ä–ł–ĺ–ī –≤–ĺ—Ā—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ł—Ź —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł —Ä–į–Ī–ĺ—ā—č –ľ–ĺ–∑–≥–į –Ņ–ĺ—Ā–Ľ–Ķ –Ņ–Ķ—Ä–ł–ĺ–ī–į –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź, –ł —Ä–į–Ī–ĺ—ā, –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–į—é—Č–ł—Ö —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł–Ķ —É—Ä–ĺ–≤–Ĺ—Ź –ļ–ĺ–≥–Ĺ–ł—ā–ł–≤–Ĺ–ĺ–Ļ –ī–Ķ—Ź—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł –Ĺ–į —Ą–ĺ–Ĺ–Ķ –≤–ĺ–∑—Ä–ĺ—Ā—ą–Ķ–Ļ —Ā–ĺ–Ĺ–Ľ–ł–≤–ĺ—Ā—ā–ł. –ü—Ä–Ķ–ī–Ľ–ĺ–∂–Ķ–Ĺ–į –į–Ľ—Ć—ā–Ķ—Ä–Ĺ–į—ā–ł–≤–Ĺ–į—Ź –≥–ł–Ņ–ĺ—ā–Ķ–∑–į, –ĺ–Ī—ä—Ź—Ā–Ĺ—Ź—é—Č–į—Ź —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł–Ķ –ļ–ĺ–≥–Ĺ–ł—ā–ł–≤–Ĺ—č—Ö —Ą—É–Ĺ–ļ—Ü–ł–Ļ –Ĺ–Ķ —É—Ö—É–ī—ą–Ķ–Ĺ–ł–Ķ–ľ —Ä–į–Ī–ĺ—ā–ĺ—Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ–ĺ—Ā—ā–ł –ľ–ĺ–∑–≥–į, –į –ĺ—ā–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ–ľ –ĺ—ā–ī–Ķ–Ľ—Ć–Ĺ—č—Ö –ļ–ĺ—Ä–ļ–ĺ–≤—č—Ö –∑–ĺ–Ĺ –ĺ—ā —É–Ņ—Ä–į–≤–Ľ–Ķ–Ĺ–ł—Ź –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ–ľ –ł–∑-–∑–į —Ä–į–∑–≤–ł—ā–ł—Ź –≤ –Ĺ–ł—Ö –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į. –Ē–į–Ĺ –ĺ–Ī–∑–ĺ—Ä –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ –į—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–Ĺ–ĺ–≥–ĺ —Ä–į–∑–≤–ł—ā–ł—Ź —Ā–Ĺ–į –≤ –ļ–ĺ—Ä–ļ–ĺ–≤—č—Ö –∑–ĺ–Ĺ–į—Ö. –ü—Ä–ł—á–ł–Ĺ—č –ł —É—Ā–Ľ–ĺ–≤–ł—Ź —Ä–į–∑–≤–ł—ā–ł—Ź –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į —Ä–į—Ā—Ā–ľ–ĺ—ā—Ä–Ķ–Ĺ—č –≤ –ļ–ĺ–Ĺ—ā–Ķ–ļ—Ā—ā–Ķ —Ä–į–∑—Ä–į–Ī–į—ā—č–≤–į–Ķ–ľ–ĺ–Ļ –į–≤—ā–ĺ—Ä–į–ľ–ł –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —ā–Ķ–ĺ—Ä–ł–ł —Ā–Ĺ–į. –í –ĺ—Ā–Ĺ–ĺ–≤–Ķ —ć—ā–ĺ–Ļ —ā–Ķ–ĺ—Ä–ł–ł –Ľ–Ķ–∂–ł—ā –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–ł–Ķ, —á—ā–ĺ –≤–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į —Ü–Ķ–Ĺ—ā—Ä–į–Ľ—Ć–Ĺ—č–Ķ –ĺ—ā–ī–Ķ–Ľ—č –Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č, –≤–ļ–Ľ—é—á–į—Ź –ļ–ĺ—Ä—É –ľ–ĺ–∑–≥–į, –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–į—é—ā—Ā—Ź —Ā –į–Ĺ–į–Ľ–ł–∑–į –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł, –Ņ–ĺ—Ā—ā—É–Ņ–į—é—Č–Ķ–Ļ –ł–∑ –≤–Ĺ–Ķ—ą–Ĺ–Ķ–Ļ —Ā—Ä–Ķ–ī—č, –Ĺ–į –į–Ĺ–į–Ľ–ł–∑ –Ņ–ĺ—ā–ĺ–ļ–į —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤ –ĺ—ā –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ā–ł—Ā—ā–Ķ–ľ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į. –ü—Ä–ł–≤–Ķ–ī–Ķ–Ĺ–į –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–į–≥–į–Ķ–ľ–į—Ź —Ā—Ö–Ķ–ľ–į –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ļ –ĺ—Ā–Ĺ–ĺ–≤–Ĺ—č—Ö –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č—Ö –Ņ–ĺ—ā–ĺ–ļ–ĺ–≤ –≤ —Ü–ł–ļ–Ľ–Ķ ¬ę—Ā–ĺ–Ĺ ‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ. –ü–ĺ–ļ–į–∑–į–Ĺ–ĺ, —á—ā–ĺ –į—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–Ĺ–ĺ—Ā—ā—Ć —ć—ā–ł—Ö –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ļ –ľ–ĺ–∂–Ķ—ā –ĺ–Ī—ä—Ź—Ā–Ĺ–ł—ā—Ć –≤–ĺ–∑–Ĺ–ł–ļ–Ĺ–ĺ–≤–Ķ–Ĺ–ł–Ķ —Ä—Ź–ī–į –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ļ, —Ā–≤—Ź–∑–į–Ĺ–Ĺ—č—Ö —Ā —Ü–ł–ļ–Ľ–ĺ–ľ ¬ę—Ā–ĺ–Ĺ ‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ (–Ņ—Ä–ĺ—Ā–ĺ–Ĺ–ĺ—á–Ĺ—č–Ķ –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—Ü–ł–ł, –≤–∑–ī—Ä–į–≥–ł–≤–į–Ĺ–ł—Ź –≤–ĺ —Ā–Ĺ–Ķ, —Ā–Ĺ–ĺ–≤–ł–ī–Ķ–Ĺ–ł—Ź –ł –Ĺ–ĺ—á–Ĺ—č–Ķ –ļ–ĺ—ą–ľ–į—Ä—č, —Ā–ĺ–ľ–Ĺ–į–ľ–Ī—É–Ľ–ł–∑–ľ –ł –Ņ–į—Ä–į–Ľ–ł—á —Ā–Ĺ–į). –ü—Ä–Ķ–ī–Ľ–ĺ–∂–Ķ–Ĺ–Ĺ–į—Ź —ā–Ķ–ĺ—Ä–ł—Ź –Ņ—Ä–Ķ–ī—Ā–ļ–į–∑—č–≤–į–Ķ—ā –Ņ–ĺ—Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ ¬ę–≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—Ü–ł–Ļ¬Ľ, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ľ–ĺ–≥—É—ā –Ľ–Ķ–∂–į—ā—Ć –≤ –ĺ—Ā–Ĺ–ĺ–≤–Ķ —ā–į–ļ–ł—Ö —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ļ, –ļ–į–ļ —É–ļ–į—á–ł–≤–į–Ĺ–ł–Ķ, –Ī–ĺ–Ľ–Ķ–∑–Ĺ—Ć –ī–≤–ł–∂–Ķ–Ĺ–ł–Ļ, –ł —Ä—Ź–ī–į –ī—Ä—É–≥–ł—Ö –Ņ—Ā–ł—Ö–ĺ—Ā–ĺ–ľ–į—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł–Ļ.
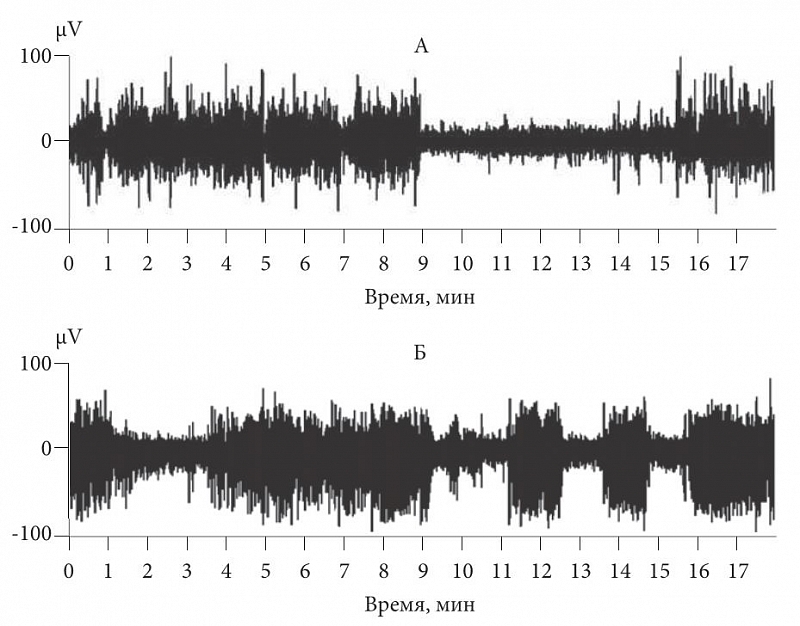
–†–ł—Ā. 1. –≠–Ľ–Ķ–ļ—ā—Ä–ĺ—ć–Ĺ—Ü–Ķ—Ą–į–Ľ–ĺ–≥—Ä–į–ľ–ľ–į –ļ—Ä–ĺ–Ľ–ł–ļ–į: –ź ‚Äď –Ņ–Ķ—Ä–ł–ĺ–ī—č –ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į –ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź; –Ď ‚Äď –ł–ľ–ł—ā–į—Ü–ł—Ź –ī–Ķ–Ľ—Ć—ā–į-–≤–ĺ–Ľ–Ĺ —Ā–Ĺ–į –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ —Ā—ā–ł–ľ—É–Ľ—Ź—Ü–ł–Ķ–Ļ –≤ —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –į–ļ—ā–ł–≤–Ĺ–ĺ–≥–ĺ –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź
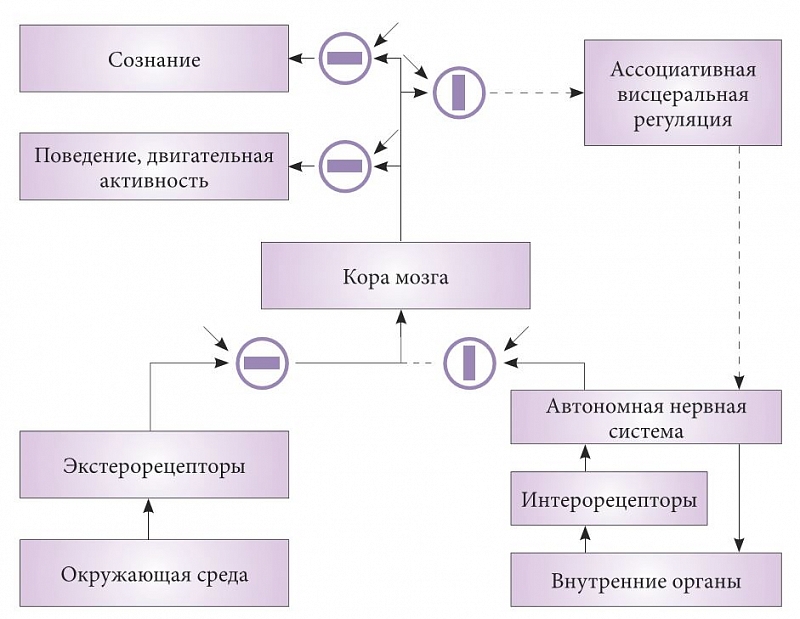
–†–ł—Ā. 2. –°—Ö–Ķ–ľ–į —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł—Ź –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č—Ö –Ņ–ĺ—ā–ĺ–ļ–ĺ–≤ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į –≤ –Ņ–Ķ—Ä–ł–ĺ–ī –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź
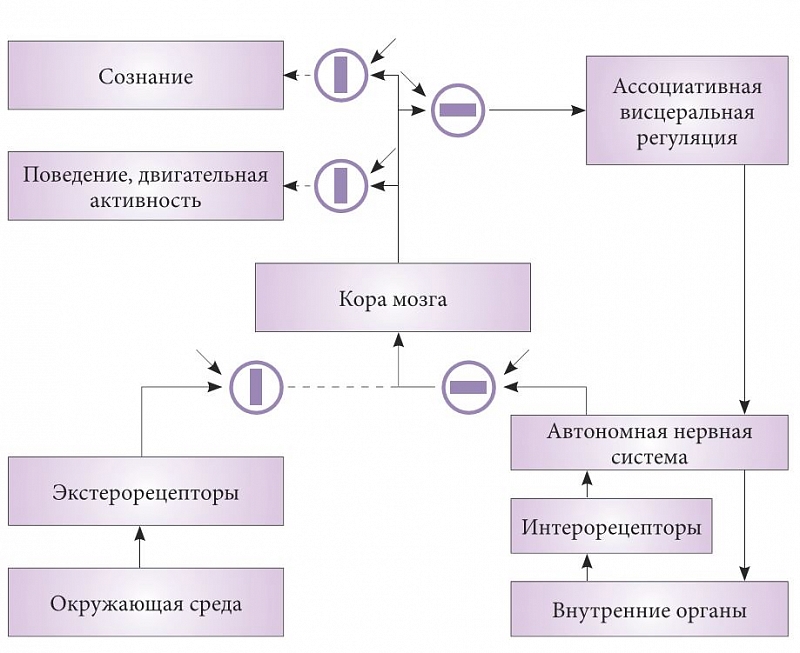
–†–ł—Ā. 3. –°—Ö–Ķ–ľ–į —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł—Ź –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č—Ö –Ņ–ĺ—ā–ĺ–ļ–ĺ–≤ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į –≤ –Ņ–Ķ—Ä–ł–ĺ–ī —Ā–Ĺ–į
–í–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ
–Ē–į–≤–Ĺ–ĺ –∑–į–ľ–Ķ—á–Ķ–Ĺ–Ĺ–į—Ź —Ā–≤—Ź–∑—Ć —ā–Ķ—Ö–Ĺ–ĺ–≥–Ķ–Ĺ–Ĺ—č—Ö –ļ–į—ā–į—Ā—ā—Ä–ĺ—Ą, –ī–ĺ—Ä–ĺ–∂–Ĺ—č—Ö –Ņ—Ä–ĺ–ł—Ā—ą–Ķ—Ā—ā–≤–ł–Ļ –ł¬†–ľ–Ķ–ī–ł—Ü–ł–Ĺ—Ā–ļ–ł—Ö –ĺ—ą–ł–Ī–ĺ–ļ —Ā¬†–Ĺ–į—Ā—ā—É–Ņ–Ľ–Ķ–Ĺ–ł–Ķ–ľ —ā–Ķ–ľ–Ĺ–ĺ–≥–ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā—É—ā–ĺ–ļ —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤—É–Ķ—ā –嬆—Ā–Ĺ–ł–∂–Ķ–Ĺ–ł–ł —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł —Ä–į–Ī–ĺ—ā—č –ľ–ĺ–∑–≥–į –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī—č –Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ–ĺ–Ļ —Ā–ĺ–Ĺ–Ľ–ł–≤–ĺ—Ā—ā–ł [1‚Äď3]. –Ē–į–Ĺ–Ĺ—č–Ķ –Ĺ–į–Ī–Ľ—é–ī–Ķ–Ĺ–ł—Ź –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–į–Ľ–ł—Ā—Ć –ł¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–į–ľ–ł —Ā–Ņ–Ķ—Ü–ł–į–Ľ—Ć–Ĺ—č—Ö –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ, –≤¬†–ļ–ĺ—ā–ĺ—Ä—č—Ö –Ī—č–Ľ–ĺ –Ņ–ĺ–ļ–į–∑–į–Ĺ–ĺ, —á—ā–ĺ –ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł—Ź —Ā–Ĺ–į –∑–į—ā—Ä—É–ī–Ĺ—Ź–Ķ—ā —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ–ĺ—Ā—ā—Ć –ļ¬†–ĺ–Ī—É—á–Ķ–Ĺ–ł—é –ł¬†–∑–į–Ņ–ĺ–ľ–ł–Ĺ–į–Ĺ–ł–Ķ –ľ–į—ā–Ķ—Ä–ł–į–Ľ–į [4, 5]. –ě–ī–Ĺ–ł –į–≤—ā–ĺ—Ä—č —Ā–≤—Ź–∑—č–≤–į–Ľ–ł –ļ–ĺ–Ĺ—Ā–ĺ–Ľ–ł–ī–į—Ü–ł—é –Ņ–į–ľ—Ź—ā–ł –ł, —Ā–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –ĺ–Ī—É—á–į–Ķ–ľ–ĺ—Ā—ā—Ć —Ā¬†–ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–≤–ĺ–Ľ–Ĺ–ĺ–≤–ĺ–Ļ —Ą–į–∑–ĺ–Ļ —Ā–Ĺ–į, —ā–ĺ–≥–ī–į –ļ–į–ļ –ī—Ä—É–≥–ł–Ķ¬†‚Äď —Ā¬†—Ą–į–∑–ĺ–Ļ –Ī—č—Ā—ā—Ä–ĺ–≥–ĺ —Ā–Ĺ–į. –ü—Ä–ł —ć—ā–ĺ–ľ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ–ĺ–Ķ –≤–Ľ–ł—Ź–Ĺ–ł–Ķ —Ā–Ĺ–į –Ĺ–į –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č –ļ–ĺ–Ĺ—Ā–ĺ–Ľ–ł–ī–į—Ü–ł–ł –Ņ–į–ľ—Ź—ā–ł –Ĺ–ł —ɬ†–ļ–ĺ–≥–ĺ –Ĺ–Ķ –≤—č–∑—č–≤–į–Ľ–ĺ —Ā–ĺ–ľ–Ĺ–Ķ–Ĺ–ł—Ź.
–ě–ī–Ĺ–į–ļ–ĺ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č —ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–ĺ–≤ –Ņ–ĺ –ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł–ł —Ā–Ĺ–į –Ī—č–Ľ–ł –≤–Ķ—Ā—Ć–ľ–į –Ņ—Ä–ĺ—ā–ł–≤–ĺ—Ä–Ķ—á–ł–≤—č –ł¬†–∑–į–≤–ł—Ā–Ķ–Ľ–ł –Ĺ–Ķ —ā–ĺ–Ľ—Ć–ļ–ĺ –ĺ—ā —ā–ł–Ņ–į –Ņ—Ä–Ķ–ī—ä—Ź–≤–Ľ—Ź–Ķ–ľ–ĺ–Ļ –∑–į–ī–į—á–ł, –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į, –Ķ–≥–ĺ —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–Ļ —Ą–į–∑—č, –≤¬†–ļ–ĺ—ā–ĺ—Ä–ĺ–Ļ —ā–Ķ—Ā—ā–ł—Ä–ĺ–≤–į–Ľ–ĺ—Ā—Ć –≤—č–Ņ–ĺ–Ľ–Ĺ–Ķ–Ĺ–ł–Ķ –∑–į–ī–į—á–ł, –Ĺ–ĺ –ł¬†–ĺ—ā –ł–Ĺ–ī–ł–≤–ł–ī—É–į–Ľ—Ć–Ĺ—č—Ö –ĺ—Ā–ĺ–Ī–Ķ–Ĺ–Ĺ–ĺ—Ā—ā–Ķ–Ļ –ł—Ā–Ņ—č—ā—É–Ķ–ľ—č—Ö. –í¬†–Ņ–ĺ–ī—Ä–ĺ–Ī–Ĺ–ĺ–ľ –ĺ–Ī–∑–ĺ—Ä–Ķ –Ľ–ł—ā–Ķ—Ä–į—ā—É—Ä—č –ĺ¬†–≤–Ľ–ł—Ź–Ĺ–ł–ł –ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł–ł —Ā–Ĺ–į –Ĺ–į –ĺ–Ī—É—á–Ķ–Ĺ–ł–Ķ –ł¬†–Ņ–į–ľ—Ź—ā—Ć –Ī—č–Ľ —Ā–ī–Ķ–Ľ–į–Ĺ –≤—č–≤–ĺ–ī: –≤¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –Ĺ–į—Ä—É—ą–Ķ–Ĺ–ł—Ź —Ā–Ĺ–į —É—Ö—É–ī—ą–į–Ķ—ā—Ā—Ź –≤—č–Ņ–ĺ–Ľ–Ĺ–Ķ–Ĺ–ł–Ķ —Ā–Ľ–ĺ–∂–Ĺ—č—Ö –∑–į–ī–į—á, —ā—Ä–Ķ–Ī—É—é—Č–ł—Ö –Ņ—Ä–ł–Ĺ—Ź—ā–ł—Ź —Ä–Ķ—ą–Ķ–Ĺ–ł—Ź, –≥–ł–Ī–ļ–ĺ—Ā—ā–ł, –ł–Ĺ–Ĺ–ĺ–≤–į—Ü–ł–ł –ł–Ľ–ł –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł—Ź –Ņ–Ľ–į–Ĺ–į –ī–Ķ–Ļ—Ā—ā–≤–ł–Ļ [6]. –ě—ā–ľ–Ķ—á–į–Ķ–ľ–ĺ–Ķ —É—Ö—É–ī—ą–Ķ–Ĺ–ł–Ķ –ļ—Ä–į—ā–ļ–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ–Ļ –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –Ņ–į–ľ—Ź—ā–ł –Ņ–ĺ—Ā–Ľ–Ķ –ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł–ł —Ā–Ĺ–į –ľ–ĺ–∂–Ĺ–ĺ –ĺ–Ī—ä—Ź—Ā–Ĺ–ł—ā—Ć –Ĺ–Ķ —Ā—ā–ĺ–Ľ—Ć–ļ–ĺ –∑–į—ā—Ä—É–ī–Ĺ–Ķ–Ĺ–ł–Ķ–ľ –ļ–ĺ–Ĺ—Ā–ĺ–Ľ–ł–ī–į—Ü–ł–ł –Ņ–į–ľ—Ź—ā–ł, —Ā–ļ–ĺ–Ľ—Ć–ļ–ĺ —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł–Ķ–ľ –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ –≤–Ĺ–ł–ľ–į–Ĺ–ł—Ź –ł/–ł–Ľ–ł –∑–į—ā—Ä—É–ī–Ĺ–Ķ–Ĺ–ł–Ķ–ľ –ĺ–Ī—Ä–į–Ī–ĺ—ā–ļ–ł –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł [7]. –°–ļ–Ľ–į–ī—č–≤–į–Ķ—ā—Ā—Ź –≤–Ņ–Ķ—á–į—ā–Ľ–Ķ–Ĺ–ł–Ķ, —á—ā–ĺ –ī–į–∂–Ķ –Ĺ–Ķ–Ī–ĺ–Ľ—Ć—ą–ĺ–Ķ —Ä–į–∑–Ľ–ł—á–ł–Ķ –≤¬†–Ņ—Ä–ĺ—ā–ĺ–ļ–ĺ–Ľ–Ķ —ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–ĺ–≤ –Ņ—Ä–ł–≤–ĺ–ī–ł–Ľ–ĺ –ļ¬†—Ā—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–ľ—É —Ä–į—Ā—Ö–ĺ–∂–ī–Ķ–Ĺ–ł—é –≤—č–≤–ĺ–ī–ĺ–≤ –ĺ—ā–Ĺ–ĺ—Ā–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –∑–į–≤–ł—Ā–ł–ľ–ĺ—Ā—ā–ł –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤ –Ņ–į–ľ—Ź—ā–ł –ĺ—ā —Ö–į—Ä–į–ļ—ā–Ķ—Ä–ł—Ā—ā–ł–ļ —Ā–Ĺ–į. –í¬†–Ņ–ĺ–Ľ—Ć–∑—É —ć—ā–ĺ–≥–ĺ –∑–į–ļ–Ľ—é—á–Ķ–Ĺ–ł—Ź —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤—É–Ķ—ā –ł¬†—ā–ĺ, —á—ā–ĺ –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č–Ķ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –Ī—č–Ľ–ł –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ—č —ā–ĺ–Ľ—Ć–ļ–ĺ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ—č–ľ–ł –≥—Ä—É–Ņ–Ņ–į–ľ–ł –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ–Ķ–Ļ [8].
–ě–ī–Ĺ–į–ļ–ĺ –Ĺ–į–Ī–Ľ—é–ī–į–Ķ–ľ–ĺ–Ķ –≤–Ľ–ł—Ź–Ĺ–ł–Ķ —Ā–Ĺ–į –Ĺ–į –∑–į–Ņ–ĺ–ľ–ł–Ĺ–į–Ĺ–ł–Ķ –Ī—č–Ľ–ĺ –Ĺ–Ķ —Ā—ā–ĺ–Ľ—Ć –∑–Ĺ–į—á–ł—ā–Ķ–Ľ—Ć–Ĺ—č–ľ, –ł¬†–ļ–į–∑–į–Ľ–ĺ—Ā—Ć –ľ–į–Ľ–ĺ–≤–Ķ—Ä–ĺ—Ź—ā–Ĺ—č–ľ, —á—ā–ĺ –ī–Ľ—Ź –Ĺ–Ķ–Ī–ĺ–Ľ—Ć—ą–ł—Ö —É–Ľ—É—á—ą–Ķ–Ĺ–ł–Ļ —ā—Ä–Ķ–Ī–ĺ–≤–į–Ľ–ł—Ā—Ć —Ā—ā–ĺ–Ľ—Ć –≥–Ľ–ĺ–Ī–į–Ľ—Ć–Ĺ—č–Ķ –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł—Ź –≤¬†—Ä–į–Ī–ĺ—ā–Ķ –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į, –ī–į –ł¬†–≤—Ā–Ķ–≥–ĺ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī—Ź—ā –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī —Ā–Ĺ–į.
–í —ā–ĺ –∂–Ķ –≤—Ä–Ķ–ľ—Ź —Ä–Ķ–ī–ļ–ł–Ķ, –Ĺ–ĺ –ī—Ä–į–ľ–į—ā–ł—á–Ķ—Ā–ļ–ł–Ķ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź –į–Ĺ—ā—Ä–ĺ–Ņ–ĺ–≥–Ķ–Ĺ–Ĺ—č—Ö –ļ–į—ā–į—Ā—ā—Ä–ĺ—Ą –ł¬†–į–≤–į—Ä–ł–Ļ –Ĺ–į –ī–ĺ—Ä–ĺ–≥–į—Ö, –ļ–ĺ—ā–ĺ—Ä—č–Ķ —Ź–≤–Ľ—Ź—é—ā—Ā—Ź —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–ĺ–ľ —Ā–ĺ–Ĺ–Ľ–ł–≤–ĺ—Ā—ā–ł, –į¬†—ā–į–ļ–∂–Ķ —Ą–į–Ĺ—ā–į—Ā—ā–ł—á–Ķ—Ā–ļ–ł–Ķ –ļ–į—Ä—ā–ł–Ĺ—č —Ā–Ĺ–ĺ–≤–ł–ī–Ķ–Ĺ–ł–Ļ –Ņ–ĺ–ī–ī–Ķ—Ä–∂–ł–≤–į–Ľ–ł –ł–ī–Ķ—é –ĺ¬†—ā–ĺ–ľ, —á—ā–ĺ —Ā–ĺ–Ĺ –≤–į–∂–Ķ–Ĺ –Ņ—Ä–Ķ–∂–ī–Ķ –≤—Ā–Ķ–≥–ĺ –ī–Ľ—Ź —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ–≥–ĺ —Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –ľ–ĺ–∑–≥–į. –ź–≤—ā–ĺ—Ä—č –Ī–ĺ–Ľ—Ć—ą–ł–Ĺ—Ā—ā–≤–į —ā–Ķ–ĺ—Ä–ł–Ļ —Ā–Ĺ–į –Ņ—Ä–Ķ–ī–Ľ–į–≥–į–Ľ–ł —Ä–į–∑–Ľ–ł—á–Ĺ—č–Ķ –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ—č, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ľ–ĺ–≥–Ľ–ł –Ī—č—ā—Ć –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ—č –ľ–ĺ–∑–≥–ĺ–ľ –≤¬†—ć—ā–ł—Ö —Ü–Ķ–Ľ—Ź—Ö: —ā–Ķ–ĺ—Ä–ł—Ź –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–Ĺ—č—Ö –≥—Ä—É–Ņ–Ņ [9] –ł–Ľ–ł —ā–Ķ–ĺ—Ä–ł—Ź —Ā–ł–Ĺ–į–Ņ—ā–ł—á–Ķ—Ā–ļ–ĺ–≥–ĺ –≥–ĺ–ľ–Ķ–ĺ—Ā—ā–į–∑–į [10]. –ė—Ā—Ö–ĺ–ī—Ź –ł–∑ —ć—ā–ł—Ö –≥–ł–Ņ–ĺ—ā–Ķ–∑, –Ī—č–Ľ–ĺ –Ī—č –Ľ–ĺ–≥–ł—á–Ĺ–ĺ –ĺ–∂–ł–ī–į—ā—Ć, —á—ā–ĺ –≤¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł–ł —Ā–Ĺ–į –≤¬†–Ņ–Ķ—Ä–≤—É—é –ĺ—á–Ķ—Ä–Ķ–ī—Ć –ī–ĺ–Ľ–∂–Ķ–Ĺ —Ā—ā—Ä–į–ī–į—ā—Ć –ł–ľ–Ķ–Ĺ–Ĺ–ĺ –ľ–ĺ–∑–≥.
–ě–ī–Ĺ–į–ļ–ĺ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź A. ¬†Rechtschaffen –ł¬†—Ā–ĺ–į–≤—ā. [11‚Äď13] –Ņ–ĺ–ļ–į–∑–į–Ľ–ł, —á—ā–ĺ —ā–ĺ—ā–į–Ľ—Ć–Ĺ–į—Ź –ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł—Ź —Ā–Ĺ–į –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –Ņ—Ä–Ķ–∂–ī–Ķ –≤—Ā–Ķ–≥–ĺ –ļ¬†–Ĺ–į—Ä—É—ą–Ķ–Ĺ–ł—Ź–ľ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č (–≤—č–Ņ–į–ī–Ķ–Ĺ–ł–Ķ –≤–ĺ–Ľ–ĺ—Ā, –Ņ–ĺ—Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ —Ź–∑–≤ –Ĺ–į –ļ–ĺ–∂–Ķ –ł–Ľ–ł —Ā–Ľ–ł–∑–ł—Ā—ā–ĺ–Ļ –∂–Ķ–Ľ—É–ī–ļ–į) –ł¬†–ļ¬†–Ĺ–Ķ–ľ–ł–Ĺ—É–Ķ–ľ–ĺ–Ļ –≥–ł–Ī–Ķ–Ľ–ł –∂–ł–≤–ĺ—ā–Ĺ—č—Ö. –ü–ĺ—Ä–į–∂–į–Ľ–ĺ, —á—ā–ĺ —É¬†–ļ—Ä—č—Ā, –Ņ–ĺ–≥–ł–Ī—ą–ł—Ö –≤¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł–ł —Ā–Ĺ–į, –Ķ–ī–ł–Ĺ—Ā—ā–≤–Ķ–Ĺ–Ĺ—č–ľ –ĺ—Ä–≥–į–Ĺ–ĺ–ľ, –Ĺ–Ķ –ł–ľ–Ķ–≤—ą–ł–ľ —Ź—Ä–ļ–ł—Ö –ī–Ķ–≥–Ķ–Ĺ–Ķ—Ä–į—ā–ł–≤–Ĺ—č—Ö –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł–Ļ, –Ī—č–Ľ –ł–ľ–Ķ–Ĺ–Ĺ–ĺ –ľ–ĺ–∑–≥ [12]. –≠—ā–ĺ –Ĺ–į–Ī–Ľ—é–ī–Ķ–Ĺ–ł–Ķ –Ī—č–Ľ–ĺ —É–ī–ł–≤–ł—ā–Ķ–Ľ—Ć–Ĺ—č–ľ, –Ņ–ĺ—Ā–ļ–ĺ–Ľ—Ć–ļ—É –Ĺ–Ķ–≥–į—ā–ł–≤–Ĺ–ĺ–Ķ –≤–ĺ–∑–ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ –ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł–ł —Ā–Ĺ–į –Ĺ–į —Ä–į–Ī–ĺ—ā—É –ľ–ĺ–∑–≥–į —Ö–ĺ—Ä–ĺ—ą–ĺ –ł–∑–≤–Ķ—Ā—ā–Ĺ–ĺ.
–§–Ķ–Ĺ–ĺ–ľ–Ķ–Ĺ –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į –ĺ—ā–ī–Ķ–Ľ—Ć–Ĺ—č—Ö –∑–ĺ–Ĺ –ļ–ĺ—Ä—č –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į
–í —Ö–ĺ–ī–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤ –ļ–ĺ—Ä—č –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į —ɬ†–Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ—á–Ķ—Ā–ļ–ł –Ī–ĺ–ī—Ä—Ā—ā–≤—É—é—Č–ł—Ö –∂–ł–≤–ĺ—ā–Ĺ—č—Ö –Ī—č–Ľ–į –≤—č—Ź–≤–Ľ–Ķ–Ĺ–į –Ņ—Ä–ł—á–ł–Ĺ–į —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł—Ź —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł—Ź, —Ā–≤—Ź–∑–į–Ĺ–Ĺ–į—Ź —Ā–ĺ —Ā–Ĺ–ĺ–ľ, –Ĺ–ĺ –Ņ—Ä–ĺ—Ź–≤–Ľ—Ź—é—Č–į—Ź—Ā—Ź –Ņ—Ä–ł –Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ä–į–Ī–ĺ—ā–Ķ –ľ–ĺ–∑–≥–į. –Ď—č–Ľ–ĺ –Ņ–ĺ–ļ–į–∑–į–Ĺ–ĺ, —á—ā–ĺ –≤¬†—É—Ā–Ľ–ĺ–≤–ł—Ź—Ö –ĺ–Ī—Č–Ķ–≥–ĺ –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –Ņ—Ä–ł –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ—č—Ö —É—Ā–Ľ–ĺ–≤–ł—Ź—Ö —Ā–ĺ–Ĺ –ľ–ĺ–∂–Ķ—ā —Ä–į–∑–≤–ł–≤–į—ā—Ć—Ā—Ź –Ľ–ĺ–ļ–į–Ľ—ƖŖ嬆‚Äď —ā–ĺ–Ľ—Ć–ļ–ĺ –≤¬†–Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö –ĺ–Ī–Ľ–į—Ā—ā—Ź—Ö –ļ–ĺ—Ä—č. –õ–ĺ–ļ–į–Ľ—Ć–Ĺ—č–Ļ —Ā–ĺ–Ĺ –ĺ–Ņ–į—Ā–Ķ–Ĺ —ā–Ķ–ľ, —á—ā–ĺ –Ķ–≥–ĺ –Ņ–ĺ—Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ –ł¬†—Ä–į–∑–≤–ł—ā–ł–Ķ –Ĺ–Ķ –∑–į–ľ–Ķ—á–į—é—ā –Ĺ–ł —Ā–į–ľ —á–Ķ–Ľ–ĺ–≤–Ķ–ļ, –Ĺ–ł –ĺ–ļ—Ä—É–∂–į—é—Č–ł–Ķ –Ķ–≥–ĺ –Ľ—é–ī–ł.
–§–Ķ–Ĺ–ĺ–ľ–Ķ–Ĺ –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ (–ł–Ľ–ł —á–į—Ā—ā–ł—á–Ĺ–ĺ–≥–ĺ) —Ā–Ĺ–į –Ī—č–Ľ –∑–į–ľ–Ķ—á–Ķ–Ĺ –ī–į–≤–Ĺ–ĺ, –Ņ—Ä–ł –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–ł –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –∑–ĺ–Ĺ –Ľ–ĺ–Ī–Ĺ–ĺ–Ļ –ļ–ĺ—Ä—č –ļ–ĺ—ą–Ķ–ļ [14]. –Ě–Ķ–Ļ—Ä–ĺ–Ĺ—č —ć—ā–ĺ–Ļ —á–į—Ā—ā–ł –ľ–ĺ–∑–≥–į –į–ļ—ā–ł–≤–Ĺ–ĺ —Ä–Ķ–į–≥–ł—Ä–ĺ–≤–į–Ľ–ł –Ĺ–į –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ—č–Ķ —Ā—ā–ł–ľ—É–Ľ—č –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī—č –Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ–ĺ–Ļ –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ—á–Ķ—Ā–ļ–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł, –Ĺ–ĺ –ł—Ö –ĺ—ā–≤–Ķ—ā—č —Ā–Ĺ–ł–∂–į–Ľ–ł—Ā—Ć –ł¬†–Ņ—Ä–ĺ–Ņ–į–ī–į–Ľ–ł –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī—č —Ā–Ņ–ĺ–ļ–ĺ–Ļ–Ĺ–ĺ–≥–ĺ –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź. –Ě–į–Ī–Ľ—é–ī–į–Ķ–ľ–į—Ź –ļ–į—Ä—ā–ł–Ĺ–į –ł–ľ–Ņ—É–Ľ—Ć—Ā–į—Ü–ł–ł –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤ –Ī—č–Ľ–į —ā–ł–Ņ–ł—á–Ĺ–ĺ–Ļ –ī–Ľ—Ź –Ņ–Ķ—Ä–ł–ĺ–ī–į —Ā–Ĺ–į. –Ē–į–Ĺ–Ĺ–ĺ–Ķ —Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ –ľ—č –Ņ–ĺ—Ā—ā–ĺ—Ź–Ĺ–Ĺ–ĺ –ĺ—ā–ľ–Ķ—á–į–Ľ–ł –≤¬†—Ö–ĺ–ī–Ķ –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ –Ņ–ĺ –ł–∑—É—á–Ķ–Ĺ–ł—é –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –∑–ĺ–Ĺ —ā–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ–Ļ –ł¬†–≤–ł—Ā–ĺ—á–Ĺ–ĺ–Ļ –ļ–ĺ—Ä—č –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į, –į¬†—ā–į–ļ–∂–Ķ –į—Ā—Ā–ĺ—Ü–ł–į—ā–ł–≤–Ĺ–ĺ–Ļ —Ā–ĺ–ľ–į—ā–ĺ—Ā–Ķ–Ĺ—Ā–ĺ—Ä–Ĺ–ĺ–Ļ –ļ–ĺ—Ä—č –ļ–ĺ—ą–Ķ–ļ.
–Ď–ĺ–Ľ–Ķ–Ķ –ī–Ķ—ā–į–Ľ—Ć–Ĺ–ĺ–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ —Ą–Ķ–Ĺ–ĺ–ľ–Ķ–Ĺ–į –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į –Ī—č–Ľ–ĺ –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–ĺ –Ņ—Ä–ł –ł–∑—É—á–Ķ–Ĺ–ł–ł —Ā–≤–ĺ–Ļ—Ā—ā–≤ –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤ –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –į—Ā—Ā–ĺ—Ü–ł–į—ā–ł–≤–Ĺ–ĺ–Ļ –∑–ĺ–Ĺ—č V4 –ĺ–Ī–Ķ–∑—Ć—Ź–Ĺ—č [15]. –í¬†–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–Ķ –ī–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ —ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į –Ņ—Ä–ł –≤—č–Ņ–ĺ–Ľ–Ĺ–Ķ–Ĺ–ł–ł –ĺ–Ī–Ķ–∑—Ć—Ź–Ĺ–ĺ–Ļ –ľ–ĺ–Ĺ–ĺ—ā–ĺ–Ĺ–Ĺ–ĺ–Ļ –∑–į–ī–į—á–ł¬†‚Äď —Ā—Ä–į–≤–Ĺ–Ķ–Ĺ–ł—Ź –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö —Ā—ā–ł–ľ—É–Ľ–ĺ–≤, –Ņ–ĺ—Ź–≤–Ľ—Ź—é—Č–ł—Ö—Ā—Ź –Ĺ–į —ć–ļ—Ä–į–Ĺ–Ķ –ľ–ĺ–Ĺ–ł—ā–ĺ—Ä–į,¬†‚Äď —á–į—Ā—ā–ĺ –∑–į–ľ–Ķ–ī–Ľ—Ź–Ľ–į—Ā—Ć —Ä–Ķ–į–ļ—Ü–ł—Ź –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤ –≤¬†–∑–ĺ–Ĺ–Ķ V4 –Ĺ–į –ĺ–ī–ł–Ĺ –ł¬†—ā–ĺ—ā –∂–Ķ –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ—č–Ļ —Ā—ā–ł–ľ—É–Ľ. –°–ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ķ–ľ –ĺ—ā–≤–Ķ—ā—č –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–į –ľ–ĺ–≥–Ľ–ł –Ņ–ĺ–Ľ–Ĺ–ĺ—Ā—ā—Ć—é –Ņ—Ä–Ķ–ļ—Ä–į—ā–ł—ā—Ć—Ā—Ź, —Ö–ĺ—ā—Ź –ĺ–Ī–Ķ–∑—Ć—Ź–Ĺ–į –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–į–Ľ–į –≤—č–Ņ–ĺ–Ľ–Ĺ—Ź—ā—Ć –Ņ–ĺ—Ā—ā–į–≤–Ľ–Ķ–Ĺ–Ĺ—É—é –∑–į–ī–į—á—É. –ö–į—á–Ķ—Ā—ā–≤–ĺ —Ä–Ķ—ą–Ķ–Ĺ–ł—Ź –∑–į–ī–į—á–ł –Ņ—Ä–ł —ć—ā–ĺ–ľ –ĺ—Ā—ā–į–≤–į–Ľ–ĺ—Ā—Ć –≤—č—Ā–ĺ–ļ–ł–ľ –ł¬†–Ľ–ł—ą—Ć –ł–Ĺ–ĺ–≥–ī–į –Ĺ–Ķ—Ā–ļ–ĺ–Ľ—Ć–ļ–ĺ —Ā–Ĺ–ł–∂–į–Ľ–ĺ—Ā—Ć –Ņ–ĺ —Ā—Ä–į–≤–Ĺ–Ķ–Ĺ–ł—é —Ā¬†—ā–į–ļ–ĺ–≤—č–ľ –≤¬†–Ĺ–į—á–į–Ľ–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź. –°–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –Ņ–ĺ –ļ—Ä–į–Ļ–Ĺ–Ķ–Ļ –ľ–Ķ—Ä–Ķ –Ņ–Ķ—Ä–≤–ł—á–Ĺ–į—Ź –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–į—Ź –ļ–ĺ—Ä–į –ľ–ĺ–∑–≥–į –ĺ–Ī–Ķ–∑—Ć—Ź–Ĺ—č –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–į–Ľ–į —Ä–į–Ī–ĺ—ā–į—ā—Ć.
–§–ĺ–Ĺ–ĺ–≤–į—Ź –ł–ľ–Ņ—É–Ľ—Ć—Ā–į—Ü–ł—Ź –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤ –≤¬†–∑–ĺ–Ĺ–Ķ V4 –≤¬†—ā–į–ļ–ł–Ķ –Ņ–Ķ—Ä–ł–ĺ–ī—č –Ĺ–į–Ņ–ĺ–ľ–ł–Ĺ–į–Ľ–į —ā–ł–Ņ–ł—á–Ĺ—É—é –ļ–į—Ä—ā–ł–Ĺ—É –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤ –≤¬†–Ķ—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–ľ —Ā–Ĺ–Ķ. –ö–ĺ–≥–ī–į –∑–į–ī–į—á—É –Ņ—Ä–Ķ—Ä—č–≤–į–Ľ–ł, –ĺ–Ī–Ķ–∑—Ć—Ź–Ĺ–į —Ā—Ä–į–∑—É –≥–Ľ—É–Ī–ĺ–ļ–ĺ –∑–į—Ā—č–Ņ–į–Ľ–į. –ü–ĺ—Ā–Ľ–Ķ –ļ–ĺ—Ä–ĺ—ā–ļ–ĺ–≥–ĺ —Ā–Ĺ–į —Ä–Ķ–į–ļ—Ü–ł—Ź –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤ –Ĺ–į –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ—č–Ķ —Ā—ā–ł–ľ—É–Ľ—č –ĺ–Ī—č—á–Ĺ–ĺ –≤–ĺ—Ā—Ā—ā–į–Ĺ–į–≤–Ľ–ł–≤–į–Ľ–ł—Ā—Ć. –í¬†—ā–ĺ–ľ –∂–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–ł –Ī—č–Ľ–ĺ –ĺ—ā–ľ–Ķ—á–Ķ–Ĺ–ĺ, —á—ā–ĺ –ī–į–∂–Ķ –≤¬†–Ņ—Ä–Ķ–ī–Ķ–Ľ–į—Ö –ĺ–ī–Ĺ–ĺ–Ļ –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –∑–ĺ–Ĺ—č V4 –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ—č–Ļ —Ā–ĺ–Ĺ —Ä–į–∑–≤–ł–≤–į–Ľ—Ā—Ź –Ĺ–Ķ –ĺ–ī–Ĺ–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ. –ě–Ĺ —Ā–Ĺ–į—á–į–Ľ–į –Ņ–ĺ—Ź–≤–Ľ—Ź–Ľ—Ā—Ź –≤¬†—É—á–į—Ā—ā–ļ–į—Ö –Ņ—Ä–ĺ–Ķ–ļ—Ü–ł–ł –Ĺ–į –ļ–ĺ—Ä—É –Ņ–Ķ—Ä–ł—Ą–Ķ—Ä–ł–ł –Ņ–ĺ–Ľ—Ź –∑—Ä–Ķ–Ĺ–ł—Ź, –į¬†–Ņ–ĺ—ā–ĺ–ľ —Ä–į—Ā–Ņ—Ä–ĺ—Ā—ā—Ä–į–Ĺ—Ź–Ľ—Ā—Ź –ļ¬†–∑–ĺ–Ĺ–Ķ –Ņ—Ä–ĺ–Ķ–ļ—Ü–ł–ł —Ą–ĺ–≤–Ķ–į (—Ü–Ķ–Ĺ—ā—Ä–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —É–≥–Ľ—É–Ī–Ľ–Ķ–Ĺ–ł—Ź –≤¬†—Ü–Ķ–Ĺ—ā—Ä–Ķ –∂–Ķ–Ľ—ā–ĺ–≥–ĺ –Ņ—Ź—ā–Ĺ–į —Ā–Ķ—ā—á–į—ā–ĺ–Ļ –ĺ–Ī–ĺ–Ľ–ĺ—á–ļ–ł –≥–Ľ–į–∑–į).
–í –ĺ–Ņ–ł—Ā–į–Ĺ–Ĺ–ĺ–ľ —Ā–Ľ—É—á–į–Ķ, –ĺ—á–Ķ–≤–ł–ī–Ĺ–ĺ, —Ä–į—Ā–Ņ—Ä–ĺ—Ā—ā—Ä–į–Ĺ–Ķ–Ĺ–ł–Ķ –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į –Ĺ–į—á–ł–Ĺ–į–Ķ—ā—Ā—Ź –ĺ—ā —Ā–Ķ–Ĺ—Ā–ĺ—Ä–Ĺ—č—Ö –ĺ–Ī–Ľ–į—Ā—ā–Ķ–Ļ ¬ę–≤—č—Ā—ą–Ķ–≥–ĺ –Ņ–ĺ—Ä—Ź–ī–ļ–į¬Ľ, –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –≤–į–∂–Ĺ—č—Ö –ī–Ľ—Ź —Ā–Ľ–ĺ–∂–Ĺ—č—Ö –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ—á–Ķ—Ā–ļ–ł—Ö —Ā–ł—ā—É–į—Ü–ł–Ļ. –ė–ľ–Ķ–Ĺ–Ĺ–ĺ –Ņ–ĺ—ć—ā–ĺ–ľ—É —Ā–Ľ–Ķ–ī–ĺ–≤–į–Ľ–ĺ –ĺ–∂–ł–ī–į—ā—Ć –Ņ–ĺ—Ź–≤–Ľ–Ķ–Ĺ–ł—Ź –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į —ɬ†–∂–ł–≤–ĺ—ā–Ĺ—č—Ö —Ā¬†–Ī–ĺ–Ľ–Ķ–Ķ –≤—č—Ä–į–∂–Ķ–Ĺ–Ĺ—č–ľ–ł –ľ–Ĺ–ĺ–∂–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ—č–ľ–ł —Ā–Ķ–Ĺ—Ā–ĺ—Ä–Ĺ—č–ľ–ł –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–ł—ā–Ķ–Ľ—Ć—Ā—ā–≤–į–ľ–ł, –Ņ—Ä–Ķ–∂–ī–Ķ –≤—Ā–Ķ–≥–ĺ —É¬†–Ņ—Ä–ł–ľ–į—ā–ĺ–≤. –ė¬†–ī–Ķ–Ļ—Ā—ā–≤–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –Ľ–ĺ–Ī–Ĺ–ĺ-–∑–į—ā—č–Ľ–ĺ—á–Ĺ—č–Ļ —ā—Ä–Ķ–Ĺ–ī —Ä–į–∑–≤–ł—ā–ł—Ź —Ā–Ĺ–į –Ī—č–Ľ –ĺ–Ī–Ĺ–į—Ä—É–∂–Ķ–Ĺ A. Borbely –ł¬†—Ā–ĺ–į–≤—ā. –≤¬†–ļ–ĺ—Ä–Ķ –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į —á–Ķ–Ľ–ĺ–≤–Ķ–ļ–į [16]. –ė—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź –ī–ł–Ĺ–į–ľ–ł–ļ–ł —Ä–į—Ā–Ņ—Ä–ĺ—Ā—ā—Ä–į–Ĺ–Ķ–Ĺ–ł—Ź —Ā–Ĺ–į –≤¬†–ľ–ĺ–∑–≥–Ķ —á–Ķ–Ľ–ĺ–≤–Ķ–ļ–į –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ—č –ł¬†–Ņ—Ä–ĺ–į–Ĺ–į–Ľ–ł–∑–ł—Ä–ĺ–≤–į–Ĺ—č –≤¬†–ĺ–Ī–∑–ĺ—Ä–Ķ 2011 –≥. [17].
–í–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ—Ā—ā—Ć –ĺ–Ī–Ĺ–į—Ä—É–∂–ł—ā—Ć –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ—č–Ļ —Ā–ĺ–Ĺ —ɬ†–∂–ł–≤–ĺ—ā–Ĺ—č—Ö —Ā¬†–Ī–ĺ–Ľ–Ķ–Ķ –Ņ—Ä–ĺ—Ā—ā–ĺ–Ļ –ĺ—Ä–≥–į–Ĺ–ł–∑–į—Ü–ł–Ķ–Ļ –ļ–ĺ—Ä—č –ł¬†–ĺ–≥—Ä–į–Ĺ–ł—á–Ķ–Ĺ–Ĺ—č–ľ –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–ĺ–ľ —Ā–Ķ–Ĺ—Ā–ĺ—Ä–Ĺ—č—Ö –∑–ĺ–Ĺ –ļ–į–∑–į–Ľ–į—Ā—Ć –ľ–į–Ľ–ĺ–≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ–Ļ. –ě–ī–Ĺ–į–ļ–ĺ –Ĺ–Ķ–ī–į–≤–Ĺ–ĺ –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ—č–Ļ —Ā–ĺ–Ĺ –Ī—č–Ľ –∑–į—Ą–ł–ļ—Ā–ł—Ä–ĺ–≤–į–Ĺ –≤¬†–Ľ–ĺ–Ī–Ĺ–ĺ–Ļ –ĺ–Ī–Ľ–į—Ā—ā–ł –ľ–ĺ–∑–≥–į –ļ—Ä—č—Ā, –≤—č–Ņ–ĺ–Ľ–Ĺ—Ź—é—Č–ł—Ö –ī–≤–ł–≥–į—ā–Ķ–Ľ—Ć–Ĺ—É—é –∑–į–ī–į—á—É [18].
–ü—Ä–ł–Ĺ–ł–ľ–į—Ź –≤–ĺ –≤–Ĺ–ł–ľ–į–Ĺ–ł–Ķ –≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ—Ā—ā—Ć —Ä–į–∑–≤–ł—ā–ł—Ź –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į, —É—Ö—É–ī—ą–Ķ–Ĺ–ł–Ķ –ļ–ĺ–≥–Ĺ–ł—ā–ł–≤–Ĺ—č—Ö —Ą—É–Ĺ–ļ—Ü–ł–Ļ –≤¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł–ł —Ā–Ĺ–į –ľ–ĺ–∂–Ĺ–ĺ –ĺ–Ī—ä—Ź—Ā–Ĺ–ł—ā—Ć –Ĺ–Ķ —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł–Ķ–ľ —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł —Ä–į–Ī–ĺ—ā—č –ľ–ĺ–∑–≥–į, –į¬†–ĺ—ā–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ–ľ –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö –∑–ĺ–Ĺ –ļ–ĺ—Ä—č –Ī–ĺ–Ľ—Ć—ą–ł—Ö –Ņ–ĺ–Ľ—É—ą–į—Ä–ł–Ļ –ĺ—ā –≤—č–Ņ–ĺ–Ľ–Ĺ–Ķ–Ĺ–ł—Ź —Ą—É–Ĺ–ļ—Ü–ł–Ļ, –∑–į –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ĺ–Ĺ–ł –ĺ—ā–≤–Ķ—á–į—é—ā –≤¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź.
–Ě–į –Ņ–Ķ—Ä–≤—č–Ļ –≤–∑–≥–Ľ—Ź–ī —ć—ā–į –≥–ł–Ņ–ĺ—ā–Ķ–∑–į –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–į–Ķ—ā –ľ–Ĺ–Ķ–Ĺ–ł–Ķ –嬆–Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ–ĺ—Ā—ā–ł –≤–ĺ—Ā—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ł—Ź —Ä–į–Ī–ĺ—ā–ĺ—Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ–ĺ—Ā—ā–ł –ľ–ĺ–∑–≥–į –Ņ–ĺ—Ā–Ľ–Ķ –ī–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ –Ņ–Ķ—Ä–ł–ĺ–ī–į –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź, —á—ā–ĺ –ł¬†–Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā –≤–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į. –ü–ĺ—Ā–Ľ–Ķ –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ī–Ľ—Ź –≤–ĺ—Ā—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ł—Ź —Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –≤—Ā–Ķ –ĺ—ā–ī–Ķ–Ľ—č –ľ–ĺ–∑–≥–į –ľ–ĺ–≥—É—ā –∑–į—Ā–Ĺ—É—ā—Ć –ĺ–ī–Ĺ–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ –ł–Ľ–ł –∂–Ķ –ļ–į–ļ–ł–Ķ-—ā–ĺ —É—á–į—Ā—ā–ļ–ł –ľ–ĺ–≥—É—ā —ć—ā–ĺ —Ā–ī–Ķ–Ľ–į—ā—Ć –≤¬†–Ņ–Ķ—Ä–≤—É—é –ĺ—á–Ķ—Ä–Ķ–ī—Ć, —ā–ĺ–≥–ī–į –ļ–į–ļ –ī—Ä—É–≥–ł–Ķ –Ī—É–ī—É—ā –Ķ—Č–Ķ –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä–ĺ–Ķ –≤—Ä–Ķ–ľ—Ź –ĺ—Ā—ā–į–≤–į—ā—Ć—Ā—Ź –≤¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź.
–Ę–Ķ–ľ –Ĺ–Ķ –ľ–Ķ–Ĺ–Ķ–Ķ —Ā–Ľ–Ķ–ī—É–Ķ—ā –Ĺ–į–Ņ–ĺ–ľ–Ĺ–ł—ā—Ć: –Ľ–ł—ą–Ķ–Ĺ–Ĺ—č–Ķ —Ā–Ĺ–į –∂–ł–≤–ĺ—ā–Ĺ—č–Ķ –Ņ–ĺ–≥–ł–Ī–į–Ľ–ł –Ĺ–Ķ –Ņ–ĺ—ā–ĺ–ľ—É, —á—ā–ĺ —Ā—ā–į–Ĺ–ĺ–≤–ł–Ľ–ł—Ā—Ć —Ā–Ľ–Ķ–Ņ—č–ľ–ł, –≥–Ľ—É—Ö–ł–ľ–ł, –∑–į–Ī—č–≤–į–Ľ–ł –Ņ—É—ā—Ć –ļ¬†–ļ–ĺ—Ä–ľ—É—ą–ļ–Ķ –ł–Ľ–ł –ł—Ā–Ņ—č—ā—č–≤–į–Ľ–ł —Ā–Ķ—Ä—Ć–Ķ–∑–Ĺ—č–Ķ —ā—Ä—É–ī–Ĺ–ĺ—Ā—ā–ł —Ā¬†–Ņ—Ä–ł–Ĺ—Ź—ā–ł–Ķ–ľ —Ä–Ķ—ą–Ķ–Ĺ–ł–Ļ, –į¬†–ł–∑-–∑–į –ľ–Ĺ–ĺ–∂–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ—č—Ö –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ä–į—Ā—Ā—ā—Ä–ĺ–Ļ—Ā—ā–≤ –≤–ĺ –≤—Ā–Ķ—Ö —Ā–ł—Ā—ā–Ķ–ľ–į—Ö –∂–ł–∑–Ĺ–Ķ–ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–Ķ–Ĺ–ł—Ź, –≤–ļ–Ľ—é—á–į—Ź –ł–ľ–ľ—É–Ĺ–Ĺ—É—é —Ā–ł—Ā—ā–Ķ–ľ—É [13]. –í¬†—ā–ĺ –∂–Ķ –≤—Ä–Ķ–ľ—Ź –ł–ľ–Ķ–Ĺ–Ĺ–ĺ –ľ–ĺ–∑–≥ –ĺ–ļ–į–∑—č–≤–į–Ľ—Ā—Ź –ĺ—Ä–≥–į–Ĺ–ĺ–ľ, –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ —É—Ā—ā–ĺ–Ļ—á–ł–≤—č–ľ –ļ¬†–ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł–ł —Ā–Ĺ–į.
–£—á–ł—ā—č–≤–į—Ź –≤—č—ą–Ķ—Ā–ļ–į–∑–į–Ĺ–Ĺ–ĺ–Ķ, –ľ–ĺ–∂–Ĺ–ĺ –Ņ—Ä–Ķ–ī–Ľ–ĺ–∂–ł—ā—Ć –ī—Ä—É–≥–ĺ–Ļ —Ą—É–Ĺ–ī–į–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ–ĺ –ĺ—ā–Ľ–ł—á–į—é—Č–ł–Ļ—Ā—Ź —Ā—Ü–Ķ–Ĺ–į—Ä–ł–Ļ. –ß—ā–ĺ –Ķ—Ā–Ľ–ł –ľ–ĺ–∑–≥, –ļ–į–ļ –ł¬†–ī—Ä—É–≥–ł–Ķ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–Ķ –ĺ—Ä–≥–į–Ĺ—č, –Ĺ–Ķ –Ĺ—É–∂–ī–į–Ķ—ā—Ā—Ź –≤¬†—Ā–Ņ–Ķ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–ľ –≤–ĺ—Ā—Ā—ā–į–Ĺ–ĺ–≤–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–ľ –ĺ—ā–ī—č—Ö–Ķ, —Ā–≤—Ź–∑–į–Ĺ–Ĺ–ĺ–ľ —Ā¬†–Ņ–ĺ–Ľ–Ĺ–ĺ–Ļ –Ņ–ĺ—ā–Ķ—Ä–Ķ–Ļ —Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–į–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł? –ß—ā–ĺ –Ķ—Ā–Ľ–ł –ľ–ĺ–∑–≥, –ļ–į–ļ –ł¬†–ļ–ĺ–ľ–Ņ—Ć—é—ā–Ķ—Ä—č, –ľ–ĺ–∂–Ķ—ā —Ä–į–Ī–ĺ—ā–į—ā—Ć —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ –Ĺ–Ķ–ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ–ĺ –ī–ĺ–Ľ–≥–ĺ, –į¬†–Ĺ–į–Ī–Ľ—é–ī–į–Ķ–ľ–į—Ź ¬ę—Ā–Ķ–Ĺ—Ā–ĺ—Ä–Ĺ–į—Ź –ł–∑–ĺ–Ľ—Ź—Ü–ł—Ź¬Ľ –ľ–ĺ–∑–≥–į –≤–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į –ĺ—ā—Ä–į–∂–į–Ķ—ā –Ĺ–Ķ –Ņ—Ä–Ķ–ļ—Ä–į—Č–Ķ–Ĺ–ł–Ķ —Ä–į–Ī–ĺ—ā—č, –į¬†–Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ –ī–Ľ—Ź –ĺ–Ī—Ä–į–Ī–ĺ—ā–ļ–ł –ī—Ä—É–≥–ĺ–≥–ĺ –Ņ–ĺ—ā–ĺ–ļ–į –≤—Ö–ĺ–ī–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł?
–≠–Ľ–Ķ–ļ—ā—Ä–ĺ—ć–Ĺ—Ü–Ķ—Ą–į–Ľ–ĺ–≥—Ä–į–ľ–ľ–į —Ā–Ĺ–į: –ĺ—ā—Ä–į–∂–Ķ–Ĺ–ł–Ķ –ĺ—Ā–ĺ–Ī–ĺ–≥–ĺ —Ä–Ķ–∂–ł–ľ–į —Ä–į–Ī–ĺ—ā—č –ľ–ĺ–∑–≥–į –≤–ĺ —Ā–Ĺ–Ķ –ł–Ľ–ł —Ö–į—Ä–į–ļ—ā–Ķ—Ä–į –≤—Ö–ĺ–ī–Ĺ–ĺ–Ļ –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł–ł?
–ě–Ī—Č–Ķ–Ņ—Ä–ł–∑–Ĺ–į–Ĺ–ĺ, —á—ā–ĺ –ļ–į—Ä—ā–ł–Ĺ—č —ć–Ľ–Ķ–ļ—ā—Ä–ĺ—ć–Ĺ—Ü–Ķ—Ą–į–Ľ–ĺ–≥—Ä–į–ľ–ľ—č (–≠–≠–ď) –ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–≤–ĺ–Ľ–Ĺ–ĺ–≤–ĺ–≥–ĺ —Ā–Ĺ–į –ł¬†–Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź —Ä–Ķ–∑–ļ–ĺ –ĺ—ā–Ľ–ł—á–į—é—ā—Ā—Ź. –≠—ā–ĺ –ľ–ĺ–∂–Ķ—ā —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤–ĺ–≤–į—ā—Ć –嬆–Ņ—Ä–ł–Ĺ—Ü–ł–Ņ–ł–į–Ľ—Ć–Ĺ–ĺ —Ä–į–∑–Ĺ–ĺ–ľ —Ö–į—Ä–į–ļ—ā–Ķ—Ä–Ķ —Ä–į–Ī–ĺ—ā—č –ľ–ĺ–∑–≥–į –≤¬†—ć—ā–ł—Ö —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł—Ź—Ö. –ě–ī–Ĺ–į–ļ–ĺ –ł–Ĺ—ā–Ķ—Ä–Ņ—Ä–Ķ—ā–į—Ü–ł—Ź —ć—ā–ĺ–≥–ĺ –Ĺ–į–Ī–Ľ—é–ī–Ķ–Ĺ–ł—Ź —ā–į–ļ–∂–Ķ –Ĺ–Ķ–ĺ–ī–Ĺ–ĺ–∑–Ĺ–į—á–Ĺ–į. –ö–į–ļ –Ņ—Ä–į–≤–ł–Ľ–ĺ, –≠–≠–ď –ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į —Ā—Ä–į–≤–Ĺ–ł–≤–į—é—ā —Ā¬†–≠–≠–ď –Ņ–į—Ā—Ā–ł–≤–Ĺ–ĺ–≥–ĺ –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź, –ļ–ĺ–≥–ī–į –Ľ—é–ī–ł –ł–Ľ–ł –∂–ł–≤–ĺ—ā–Ĺ—č–Ķ –Ĺ–į—Ö–ĺ–ī—Ź—ā—Ā—Ź –Ī–Ķ–∑ –ī–≤–ł–∂–Ķ–Ĺ–ł–Ļ –ł¬†—Ā–Ķ–Ĺ—Ā–ĺ—Ä–Ĺ—č–Ķ —Ā—ā–ł–ľ—É–Ľ—č –Ņ—Ä–į–ļ—ā–ł—á–Ķ—Ā–ļ–ł –ĺ—ā—Ā—É—ā—Ā—ā–≤—É—é—ā. –ė–ľ–Ķ–Ĺ–Ĺ–ĺ –Ņ–ĺ—ć—ā–ĺ–ľ—É –Ĺ–į–Ī–Ľ—é–ī–į–Ķ–ľ—č–Ķ —Ä–į–∑–Ľ–ł—á–ł—Ź –≤¬†–ļ–į—Ä—ā–ł–Ĺ–į—Ö –≠–≠–ď –ľ–ĺ–≥—É—ā –Ī—č—ā—Ć —Ā–≤—Ź–∑–į–Ĺ—č —Ā¬†—Ä–į–∑–Ľ–ł—á–Ĺ–ĺ–Ļ –ļ–ĺ—Ä–ļ–ĺ–≤–ĺ–Ļ –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł–Ķ–Ļ.
–Ē–Ľ—Ź —ā–ĺ–≥–ĺ —á—ā–ĺ–Ī—č –Ņ—Ä–ĺ–≤–Ķ—Ä–ł—ā—Ć —ć—ā—É –≥–ł–Ņ–ĺ—ā–Ķ–∑—É, –ľ—č —Ä–Ķ–≥–ł—Ā—ā—Ä–ł—Ä–ĺ–≤–į–Ľ–ł –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–Ĺ—É—é –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć –ł¬†–Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ—É—é —ć–Ľ–Ķ–ļ—ā—Ä–ĺ–ļ–ĺ—Ä—ā–ł–ļ–ĺ–≥—Ä–į–ľ–ľ—É —ɬ†–ļ–ĺ—ą–Ķ–ļ –ł¬†–ļ—Ä–ĺ–Ľ–ł–ļ–ĺ–≤. –Ē–Ľ—Ź –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –Ī—č–Ľ–ł –Ņ–ĺ–ī–ĺ–Ī—Ä–į–Ĺ—č —Ā—ā–ł–ľ—É–Ľ—č (–∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ—č–Ļ –ł–Ľ–ł —Ā–ĺ–ľ–į—ā–ł—á–Ķ—Ā–ļ–ł–Ļ), –≤—č–∑—č–≤–į—é—Č–ł–Ķ –į–ļ—ā–ł–≤–Ĺ—É—é –ł–ľ–Ņ—É–Ľ—Ć—Ā–į—Ü–ł—é –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤, –ł¬†—ć—ā–ł —Ā—ā–ł–ľ—É–Ľ—č –Ņ—Ä–Ķ–ī—ä—Ź–≤–Ľ—Ź–Ľ–ł —Ā¬†—á–į—Ā—ā–ĺ—ā–ĺ–Ļ, —Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤—É—é—Č–Ķ–Ļ —á–į—Ā—ā–ĺ—ā–Ķ –ī–Ķ–Ľ—Ć—ā–į-–≤–ĺ–Ľ–Ĺ. –í¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –į–ļ—ā–ł–≤–Ĺ–ĺ–≥–ĺ –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –∂–ł–≤–ĺ—ā–Ĺ–ĺ–≥–ĺ –ľ—č –ī–Ķ–Ļ—Ā—ā–≤–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –Ĺ–į–Ī–Ľ—é–ī–į–Ľ–ł –Ĺ–į –≠–≠–ď –Ņ–į—á–Ķ—á–Ĺ—É—é –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤ –ł¬†–ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ—č–Ķ –≤–ĺ–Ľ–Ĺ—č, –Ĺ–Ķ –ĺ—ā–Ľ–ł—á–ł–ľ—č–Ķ –ĺ—ā –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł –≤¬†—Ā—ā–į–ī–ł–ł –ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į (—Ä–ł—Ā.¬†1). –Ě–į —Ä–ł—Ā—É–Ĺ–ļ–Ķ¬†1–į –Ņ–ĺ–ļ–į–∑–į–Ĺ—č —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł—Ź –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ł¬†–ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į, –ĺ—ā–Ľ–ł—á–į—é—Č–Ķ–≥–ĺ—Ā—Ź –Ņ–ĺ—Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ–ľ –≤—č—Ā–ĺ–ļ–ĺ–į–ľ–Ņ–Ľ–ł—ā—É–ī–Ĺ—č—Ö –ī–Ķ–Ľ—Ć—ā–į-–≤–ĺ–Ľ–Ĺ. –†–ł—Ā—É–Ĺ–ĺ–ļ¬†1–Ī –ĺ—ā—Ä–į–∂–į–Ķ—ā —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ķ –į–ļ—ā–ł–≤–Ĺ–ĺ–≥–ĺ –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź¬†‚Äď –ļ—Ä–ĺ–Ľ–ł–ļ–į —Ā—ā–ł–ľ—É–Ľ–ł—Ä–ĺ–≤–į–Ľ–ł –Ĺ–Ķ—Ź—Ä–ļ–ł–ľ–ł –≤—Ā–Ņ—č—ą–ļ–į–ľ–ł —Ā–≤–Ķ—ā–į. –≠—ā–ł –≤—Ā–Ņ—č—ą–ļ–ł –≤—č–∑—č–≤–į–Ľ–ł –Ņ–ĺ—Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ –≤—č–∑–≤–į–Ĺ–Ĺ—č—Ö –ĺ—ā–≤–Ķ—ā–ĺ–≤, –ļ–ĺ—ā–ĺ—Ä—č–Ķ, —Ā–Ľ–ł–≤–į—Ź—Ā—Ć, –Ņ—Ä–ł–≤–ĺ–ī–ł–Ľ–ł –ļ¬†–ļ–į—Ä—ā–ł–Ĺ–Ķ, –Ĺ–Ķ–ĺ—ā–Ľ–ł—á–ł–ľ–ĺ–Ļ –ĺ—ā —ā–į–ļ–ĺ–≤–ĺ–Ļ –≤¬†–ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–ľ —Ā–Ĺ–Ķ. –ě—ā–≤–Ķ—ā—č –Ĺ–į –≤—Ā–Ņ—č—ą–ļ–ł –Ĺ–į–Ņ–ĺ–ľ–ł–Ĺ–į–Ľ–ł –ļ–į—Ä—ā–ł–Ĺ—É –≠–≠–ď –Ņ—Ä–ł –ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–ľ —Ā–Ĺ–Ķ. –ö–į–ļ –ī–Ľ—Ź –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ, —ā–į–ļ –ł¬†–ī–Ľ—Ź —Ā–ĺ–ľ–į—ā–ĺ—Ā–Ķ–Ĺ—Ā–ĺ—Ä–Ĺ–ĺ–Ļ –ļ–ĺ—Ä—č –ľ–ĺ–∂–Ĺ–ĺ –Ī—č–Ľ–ĺ –Ņ–ĺ–ī–ĺ–Ī—Ä–į—ā—Ć —ā–į–ļ–ł–Ķ —É—Ā–Ľ–ĺ–≤–ł—Ź —Ā–Ķ–Ĺ—Ā–ĺ—Ä–Ĺ–ĺ–Ļ —Ā—ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł, –Ņ—Ä–ł –ļ–ĺ—ā–ĺ—Ä—č—Ö —Ā–Ņ–Ķ–ļ—ā—Ä—č –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–Ļ —ć–Ľ–Ķ–ļ—ā—Ä–ĺ–ļ–ĺ—Ä—ā–ł–ļ–ĺ–≥—Ä–į–ľ–ľ—č –≤¬†–Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–ł –ł¬†–≤–ĺ —Ā–Ĺ–Ķ –Ĺ–Ķ –ĺ—ā–Ľ–ł—á–į–Ľ–ł—Ā—Ć. –ó–Ĺ–į—á–ł—ā, —Ö–į—Ä–į–ļ—ā–Ķ—Ä –≠–≠–ď –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ—Ź–Ķ—ā –ł–ľ–Ķ–Ĺ–Ĺ–ĺ –≤—Ö–ĺ–ī–Ĺ–į—Ź –ļ–ĺ—Ä–ļ–ĺ–≤–į—Ź –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł—Ź, –į¬†–Ĺ–Ķ —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ķ —Ā–Ĺ–į –ł–Ľ–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź.
–§–Ķ–Ĺ–ĺ–ľ–Ķ–Ĺ –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į –≤¬†–ļ–ĺ–Ĺ—ā–Ķ–ļ—Ā—ā–Ķ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ¬†—ā–Ķ–ĺ—Ä–ł–ł —Ā–Ĺ–į
–ü–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ –ļ–ĺ—Ä—č –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į –Ĺ–į –į–Ĺ–į–Ľ–ł–∑ –ī—Ä—É–≥–ĺ–≥–ĺ –Ņ–ĺ—ā–ĺ–ļ–į –≤—Ö–ĺ–ī—Ź—Č–Ķ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł –ľ–ĺ–∂–Ķ—ā –Ľ–Ķ–∂–į—ā—Ć –≤¬†–ĺ—Ā–Ĺ–ĺ–≤–Ķ –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–į –ĺ—ā —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł—Ź –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ļ–ĺ —Ā–Ĺ—É. –ö–į–ļ–ł–Ķ –∂–Ķ —Ā–ł–≥–Ĺ–į–Ľ—č –ľ–ĺ–≥–Ľ–ł –Ī—č –ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–ł—ā—Ć —ā–į–ļ—É—é –Ņ–Ķ—Ä–ł–ĺ–ī–ł—á–Ķ—Ā–ļ—É—é –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł—é –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī —Ā–Ĺ–į? –°¬†—ā–ĺ—á–ļ–ł –∑—Ä–Ķ–Ĺ–ł—Ź —Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł–ł –ĺ—ā–≤–Ķ—ā –Ņ—Ä–ĺ—Ā—ā. –≠—ā–ĺ –ľ–ĺ–≥–Ľ–į –Ī—č—ā—Ć –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć —Ä–į–∑–Ĺ—č—Ö –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ā–ł—Ā—ā–Ķ–ľ, –Ĺ–į–Ņ—Ä–ł–ľ–Ķ—Ä —Ā–Ķ—Ä–ī–Ķ—á–Ĺ–ĺ-—Ā–ĺ—Ā—É–ī–ł—Ā—ā–ĺ–Ļ –ł–Ľ–ł –ī—č—Ö–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ, –∂–Ķ–Ľ—É–ī–ĺ—á–Ĺ–ĺ-–ļ–ł—ą–Ķ—á–Ĺ–ĺ–≥–ĺ —ā—Ä–į–ļ—ā–į.
–ú—č –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–ł–Ľ–ł, —á—ā–ĺ –≤–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ—č –ľ–ĺ–∑–≥–į, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –≤¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ĺ–Ī—Ä–į–Ī–į—ā—č–≤–į—é—ā —ć–ļ—Ā—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ—É—é –ł¬†–Ņ—Ä–ĺ–Ņ—Ä–ł–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ—É—é –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł—é, –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–į—é—ā—Ā—Ź –Ĺ–į –į–Ĺ–į–Ľ–ł–∑ –ł–Ĺ—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł, –Ņ—Ä–ł—Ö–ĺ–ī—Ź—Č–Ķ–Ļ –ĺ—ā —Ä–į–∑–Ĺ—č—Ö –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ā–ł—Ā—ā–Ķ–ľ. –†–ł—ā–ľ–ł—á–Ķ—Ā–ļ–į—Ź –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć —ć—ā–ł—Ö –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ā–ł—Ā—ā–Ķ–ľ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ—Ź–Ķ—ā –Ņ–Ķ—Ä–ł–ĺ–ī–ł—á–Ķ—Ā–ļ–ł–Ļ –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–Ĺ—č–Ļ –Ņ–ĺ—ā–ĺ–ļ –ļ¬†–ļ–ĺ—Ä–ļ–ĺ–≤—č–ľ –∑–ĺ–Ĺ–į–ľ, —á—ā–ĺ –ł¬†–ĺ—ā—Ä–į–∂–į–Ķ—ā –ļ–ĺ—Ä–ļ–ĺ–≤—č–Ļ —Ä–ł—ā–ľ –≠–≠–ď –ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–≤–ĺ–Ľ–Ĺ–ĺ–≤–ĺ–≥–ĺ —Ā–Ĺ–į. –Ę–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī —Ā–Ĺ–į –≤—č—Ā—ą–ł–Ķ –ĺ—ā–ī–Ķ–Ľ—č —Ü–Ķ–Ĺ—ā—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č –≤–ļ–Ľ—é—á–į—é—ā—Ā—Ź –≤¬†–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł.
–°–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –Ņ–Ķ—Ä–ł–ĺ–ī—č –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į¬†‚Äď —ć—ā–ĺ –Ĺ–Ķ —ā–Ķ –Ņ–Ķ—Ä–ł–ĺ–ī—č, –ļ–ĺ–≥–ī–į —É—Ā—ā–į–Ľ—č–Ļ –ľ–ĺ–∑–≥ –Ņ—Ä–Ķ–ļ—Ä–į—Č–į–Ķ—ā –ĺ–Ī—Ä–į–Ī–ĺ—ā–ļ—É —ć–ļ—Ā—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł –ī–Ľ—Ź –≤–ĺ—Ā—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ł—Ź —Ā–≤–ĺ–Ķ–Ļ —Ä–į–Ī–ĺ—ā–ĺ—Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ–ĺ—Ā—ā–ł, –į¬†–Ņ–Ķ—Ä–ł–ĺ–ī—č, –ļ–ĺ–≥–ī–į –Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ —Ä–į–Ī–ĺ—ā–į—é—Č–ł–Ķ –∑–ĺ–Ĺ—č –ľ–ĺ–∑–≥–į, —Ä–Ķ–į–≥–ł—Ä—É—Ź –Ĺ–į —Ā–ł–≥–Ĺ–į–Ľ—č —ā—Ä–Ķ–≤–ĺ–≥–ł, –ł–ī—É—Č–ł–Ķ –ĺ—ā –≤–Ĺ—É—ā—Ä–Ķ–Ĺ–Ĺ–ł—Ö –ĺ—Ä–≥–į–Ĺ–ĺ–≤, –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–į—é—ā—Ā—Ź –Ĺ–į –į–Ĺ–į–Ľ–ł–∑ –Ņ–ĺ—Ā—ā—É–Ņ–į—é—Č–Ķ–Ļ –ĺ—ā –Ĺ–ł—Ö –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł–ł.
–≠—ā–ĺ –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–ł–Ķ, —Ā¬†–ĺ–ī–Ĺ–ĺ–Ļ —Ā—ā–ĺ—Ä–ĺ–Ĺ—č, –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ—Ź–Ľ–ĺ—Ā—Ć —Ą–į–Ĺ—ā–į—Ā—ā–ł—á–Ķ—Ā–ļ–ł–ľ –ī–Ľ—Ź –ĺ–Ī—Č–Ķ–Ņ—Ä–ł–Ĺ—Ź—ā–ĺ–Ļ –Ņ–į—Ä–į–ī–ł–≥–ľ—č –ľ–ĺ–∑–≥–į, —Ā—Ą–ĺ—Ä–ľ–ł—Ä–ĺ–≤–į–Ĺ–Ĺ–ĺ–Ļ –Ĺ–į –ĺ—Ā–Ĺ–ĺ–≤–Ķ —ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–ĺ–≤, –≤—č–Ņ–ĺ–Ľ–Ĺ–Ķ–Ĺ–Ĺ—č—Ö –≤¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź. –°¬†–ī—Ä—É–≥–ĺ–Ļ —Ā—ā–ĺ—Ä–ĺ–Ĺ—č, —ć—ā–ĺ –ĺ—ā–ļ—Ä—č–≤–į–Ľ–ĺ –≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ—Ā—ā–ł –ī–Ľ—Ź –Ņ—Ä–ĺ–≤–Ķ—Ä–ļ–ł –ī–į–Ĺ–Ĺ–ĺ–≥–ĺ –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–ł—Ź –≤¬†–ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ –Ņ—Ä–ĺ—Ā—ā—č—Ö —É—Ā–Ľ–ĺ–≤–ł—Ź—Ö.
–†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č —Ä–į–∑–Ĺ–ĺ–Ņ–Ľ–į–Ĺ–ĺ–≤—č—Ö —ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–ĺ–≤ –Ĺ–į –∂–ł–≤—č—Ö –ľ–ĺ–ī–Ķ–Ľ—Ź—Ö (–ļ–ĺ—ą–ļ–ł, –ĺ–Ī–Ķ–∑—Ć—Ź–Ĺ—č, –ļ—Ä–ĺ–Ľ–ł–ļ–ł), –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–Ĺ—č—Ö —Ā¬†—Ü–Ķ–Ľ—Ć—é –Ņ—Ä–ĺ–≤–Ķ—Ä–ļ–ł –≤—č—Ā–ļ–į–∑–į–Ĺ–Ĺ–ĺ–Ļ –≥–ł–Ņ–ĺ—ā–Ķ–∑—č, –ĺ–Ņ—É–Ī–Ľ–ł–ļ–ĺ–≤–į–Ĺ—č [19, 20]. –Ď—č–Ľ–ĺ –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–Ķ–Ĺ–ĺ, —á—ā–ĺ –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī—č —Ā–Ĺ–į –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ—č –≤—Ā–Ķ—Ö –ł—Ā—Ā–Ľ–Ķ–ī—É–Ķ–ľ—č—Ö –∑–ĺ–Ĺ –ļ–ĺ—Ä—č –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į (–∑–į—ā—č–Ľ–ĺ—á–Ĺ–ĺ–Ļ, —ā–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ–Ļ –ł¬†–Ľ–ĺ–Ī–Ĺ–ĺ–Ļ) –Ĺ–į—á–ł–Ĺ–į–Ľ–ł —Ä–Ķ–į–≥–ł—Ä–ĺ–≤–į—ā—Ć –Ĺ–į —Ā–ł–≥–Ĺ–į–Ľ—č, –Ņ—Ä–ł—Ö–ĺ–ī—Ź—Č–ł–Ķ –ł–∑ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ā–ł—Ā—ā–Ķ–ľ. –°—Ä–į–∑—É –Ņ–ĺ—Ā–Ľ–Ķ –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł—Ź –∂–ł–≤–ĺ—ā–Ĺ–ĺ–≥–ĺ —Ā–≤—Ź–∑—Ć –ľ–ĺ–∑–≥–į —Ā¬†–≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–ľ–ł —Ā–ł—Ā—ā–Ķ–ľ–į–ľ–ł –Ņ—Ä–Ķ—Ä—č–≤–į–Ľ–į—Ā—Ć.
–≠—ā–ĺ—ā —Ą–į–ļ—ā –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź–Ķ—ā –≥–ĺ–≤–ĺ—Ä–ł—ā—Ć —É–∂–Ķ –嬆–≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —ā–Ķ–ĺ—Ä–ł–ł —Ā–Ĺ–į. –°–ĺ–≥–Ľ–į—Ā–Ĺ–ĺ —ć—ā–ĺ–Ļ —ā–Ķ–ĺ—Ä–ł–ł –ł–ľ–Ķ–Ĺ–Ĺ–ĺ –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł—Ź –ĺ—ā –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö –ĺ—Ä–≥–į–Ĺ–ĺ–≤ —Ā¬†—Ź–≤–Ĺ–ĺ –≤—č—Ä–į–∂–Ķ–Ĺ–Ĺ–ĺ–Ļ —Ä–ł—ā–ľ–ł—á–Ĺ–ĺ—Ā—ā—Ć—é –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ—Ź–Ķ—ā –≤–ĺ–Ľ–Ĺ–ĺ–≤—É—é –ļ–į—Ä—ā–ł–Ĺ—É –ļ–ĺ—Ä–ļ–ĺ–≤–ĺ–Ļ –į–ļ—ā–ł–≤–į—Ü–ł–ł –≤¬†—Ā—ā–į–ī–ł–ł –ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į. –Ē–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ł–∑–į—Ü–ł—Ź —ć–Ľ–Ķ–ļ—ā—Ä–ĺ—ć–Ĺ—Ü–Ķ—Ą–į–Ľ–ĺ–≥—Ä–į–ľ–ľ—č –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī—č –Ī—č—Ā—ā—Ä–ĺ–≥–ĺ —Ā–Ĺ–į –ľ–ĺ–∂–Ķ—ā —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤–ĺ–≤–į—ā—Ć –嬆–Ņ–ĺ—Ā—ā—É–Ņ–Ľ–Ķ–Ĺ–ł–ł –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł–ł –ĺ—ā –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö –ĺ—Ä–≥–į–Ĺ–ĺ–≤, –Ĺ–Ķ –ł–ľ–Ķ—é—Č–ł—Ö —Ź–≤–Ĺ–ĺ–Ļ –Ņ–Ķ—Ä–ł–ĺ–ī–ł—á–Ķ—Ā–ļ–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł,¬†‚Äď –Ņ–Ķ—á–Ķ–Ĺ–ł, –Ņ–ĺ—á–Ķ–ļ, –ĺ—Ä–≥–į–Ĺ–ĺ–≤ —Ä–Ķ–Ņ—Ä–ĺ–ī—É–ļ—ā–ł–≤–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č –ł¬†–ľ–ĺ–∑–≥–į.
–í —Ä–į–ľ–ļ–į—Ö –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —ā–Ķ–ĺ—Ä–ł–ł —Ā–Ĺ–į –ľ–ĺ–∑–≥ –ł–ľ–Ķ–Ķ—ā –ī–≤–ĺ–Ļ—Ā—ā–≤–Ķ–Ĺ–Ĺ—č–Ļ —Ā—ā–į—ā—É—Ā: —Ā¬†–ĺ–ī–Ĺ–ĺ–Ļ —Ā—ā–ĺ—Ä–ĺ–Ĺ—č, —ć—ā–ĺ —Ü–Ķ–Ĺ—ā—Ä–į–Ľ—Ć–Ĺ—č–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č–Ļ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ—Ä, –ļ–ĺ–Ĺ—ā—Ä–ĺ–Ľ–ł—Ä—É—é—Č–ł–Ļ –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ –≤¬†–ĺ–ļ—Ä—É–∂–į—é—Č–Ķ–Ļ —Ā—Ä–Ķ–ī–Ķ –≤–ĺ –≤—Ä–Ķ–ľ—Ź –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ł¬†–ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–ł–≤–į—é—Č–ł–Ļ –≤–ĺ—Ā—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ł–Ķ —Ä–į–Ī–ĺ—ā–ĺ—Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ–ĺ—Ā—ā–ł –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ā–ł—Ā—ā–Ķ–ľ –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī —Ā–Ĺ–į, –į¬†—Ā¬†–ī—Ä—É–≥–ĺ–Ļ¬†‚Äď —Ā–Ľ–ĺ–∂–Ĺ–Ķ–Ļ—ą–ł–Ļ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–Ļ –ĺ—Ä–≥–į–Ĺ, –ļ–ĺ—ā–ĺ—Ä—č–Ļ —ā–į–ļ–∂–Ķ –Ĺ—É–∂–ī–į–Ķ—ā—Ā—Ź –≤¬†–Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł—á–Ķ—Ā–ļ–ĺ–ľ –ĺ–Ī—Ā–Ľ—É–∂–ł–≤–į–Ĺ–ł–ł. –ö–į–ļ –ł¬†–ļ–ĺ–≥–ī–į —ā–į–ļ–ĺ–Ķ —Ā–į–ľ–ĺ–ĺ–Ī—Ā–Ľ—É–∂–ł–≤–į–Ĺ–ł–Ķ –ľ–ĺ–∑–≥–į –ĺ—Ā—É—Č–Ķ—Ā—ā–≤–Ľ—Ź–Ķ—ā—Ā—Ź, –Ņ–ĺ–ļ–į –Ĺ–Ķ —Ź—Ā–Ĺ–ĺ. –ú–ĺ–∂–Ĺ–ĺ –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–ł—ā—Ć, —á—ā–ĺ —ć—ā–ĺ –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā –≤¬†–ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ—č–Ķ –Ņ–Ķ—Ä–ł–ĺ–ī—č –Ī—č—Ā—ā—Ä–ĺ–≥–ĺ –ł–Ľ–ł –ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į –ł–Ľ–ł –∂–Ķ –ĺ—Ā—É—Č–Ķ—Ā—ā–≤–Ľ—Ź–Ķ—ā—Ā—Ź –Ņ–ĺ—Ā—ā–ĺ—Ź–Ĺ–Ĺ–ĺ, –Ĺ–į–Ņ—Ä–ł–ľ–Ķ—Ä, —Ā¬†–Ņ–ĺ–ľ–ĺ—Č—Ć—é –≥–Ľ–ł–į–Ľ—Ć–Ĺ—č—Ö –ļ–Ľ–Ķ—ā–ĺ–ļ. –Ě–Ķ–ī–į–≤–Ĺ–ĺ –ĺ—ā–ļ—Ä—č—ā—č–Ļ –≥–Ľ–ł–ľ—Ą–į—ā–ł—á–Ķ—Ā–ļ–ł–Ļ (glymphatic) –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ –ľ–ĺ–∂–Ķ—ā –Ī—č—ā—Ć —á–į—Ā—ā—Ć—é —ć—ā–ĺ–≥–ĺ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–į [21, 22].
–ü—Ä–Ķ–ī–Ľ–ĺ–∂–Ķ–Ĺ–Ĺ—č–Ļ –Ņ–ĺ–ī—Ö–ĺ–ī –Ņ—Ä–ĺ—Ź—Ā–Ĺ—Ź–Ķ—ā –∑–į–Ņ—É—Ā–ļ —Ā–Ĺ–į. –Ē–Ķ–Ļ—Ā—ā–≤–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –Ĺ–Ķ—Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–ł–Ķ —ā–Ķ–ļ—É—Č–ł—Ö –Ņ–į—Ä–į–ľ–Ķ—ā—Ä–ĺ–≤, –ĺ—ā—Ä–į–∂–į—é—Č–ł—Ö —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ķ –Ľ—é–Ī–ĺ–Ļ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č, –ł¬†–≥–Ķ–Ĺ–Ķ—ā–ł—á–Ķ—Ā–ļ–ł –∑–į–ī–į–Ĺ–Ĺ—č—Ö –Ī—É–ī–Ķ—ā –≤—č–∑—č–≤–į—ā—Ć –ĺ—Č—É—Č–Ķ–Ĺ–ł–Ķ —É—Ā—ā–į–Ľ–ĺ—Ā—ā–ł –ł–Ľ–ł –ī–į–≤–Ľ–Ķ–Ĺ–ł—Ź —Ā–Ĺ–į. –ē—Ā–Ľ–ł —Ā–ł—ā—É–į—Ü–ł—Ź –ī–ĺ–Ņ—É—Ā–ļ–į–Ķ—ā –Ĺ–Ķ–ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ—č–Ļ –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī –ļ–ĺ —Ā–Ĺ—É, –Ņ–ĺ—Ā–Ľ–Ķ–ī–Ĺ–ł–Ļ –Ī—É–ī–Ķ—ā –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā—Ć –≤–ĺ –≤—Ā–Ķ—Ö –ļ–ĺ—Ä–ļ–ĺ–≤—č—Ö –∑–ĺ–Ĺ–į—Ö. –ö–ĺ–≥–ī–į –≤¬†—ā–ĺ–Ļ –ł–Ľ–ł –ł–Ĺ–ĺ–Ļ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ–Ķ –Ņ–ĺ—ā—Ä–Ķ–Ī–Ĺ–ĺ—Ā—ā—Ć —Ā–Ĺ–į —Ä–Ķ–∑–ļ–ĺ –≤–ĺ–∑—Ä–į—Ā—ā–į–Ķ—ā, –Ĺ–ĺ –ĺ–ļ—Ä—É–∂–į—é—Č–į—Ź –ĺ–Ī—Ā—ā–į–Ĺ–ĺ–≤–ļ–į –Ĺ–Ķ –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź–Ķ—ā –Ņ–Ķ—Ä–Ķ–Ļ—ā–ł –ļ–ĺ —Ā–Ĺ—É, —Ā–ĺ–Ĺ –ľ–ĺ–∂–Ķ—ā –Ĺ–į—Ā—ā—É–Ņ–ł—ā—Ć –Ľ–ł—ą—Ć –≤¬†–Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö –ļ–ĺ—Ä–ļ–ĺ–≤—č—Ö –∑–ĺ–Ĺ–į—Ö –Ņ—Ä–ł —Ā–ĺ—Ö—Ä–į–Ĺ–Ķ–Ĺ–ł–ł –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ—á–Ķ—Ā–ļ–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł. –†–į–∑–≤–ł—ā–ł–Ķ —Ā–Ĺ–į –Ĺ–į—á–ł–Ĺ–į–Ķ—ā—Ā—Ź —Ā¬†—Ą–ł–Ľ–ĺ–≥–Ķ–Ĺ–Ķ—ā–ł—á–Ķ—Ā–ļ–ł –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –ľ–ĺ–Ľ–ĺ–ī—č—Ö –ļ–ĺ—Ä–ļ–ĺ–≤—č—Ö –∑–ĺ–Ĺ ¬ę–≤—č—Ā–ĺ–ļ–ĺ–≥–ĺ —É—Ä–ĺ–≤–Ĺ—Ź¬Ľ (–ĺ–Ĺ–ł –Ņ—Ä–Ķ–ĺ–Ī–Ľ–į–ī–į—é—ā –≤¬†–Ľ–ĺ–Ī–Ĺ—č—Ö –ĺ—ā–ī–Ķ–Ľ–į—Ö –ľ–ĺ–∑–≥–į). –°¬†—ć—ā–ł–ľ, –≤–ł–ī–ł–ľ–ĺ, –ł¬†—Ā–≤—Ź–∑–į–Ĺ –≤—č—Ź–≤–Ľ–Ķ–Ĺ–Ĺ—č–Ļ –≤¬†–≠–≠–ď –Ľ–ĺ–Ī–Ĺ–ĺ-–∑–į—ā—č–Ľ–ĺ—á–Ĺ—č–Ļ —ā—Ä–Ķ–Ĺ–ī —Ä–į–∑–≤–ł—ā–ł—Ź —Ā–Ĺ–į [16].
–ü–ĺ–≤–Ķ–ī–Ķ–Ĺ—á–Ķ—Ā–ļ–ł–Ķ –∑–į–ī–į—á–ł, –Ĺ–Ķ —ā—Ä–Ķ–Ī—É—é—Č–ł–Ķ –≤–ĺ–≤–Ľ–Ķ—á–Ķ–Ĺ–ł—Ź –≤—č—Ā—ą–ł—Ö –ļ–ĺ—Ä–ļ–ĺ–≤—č—Ö —Ä–Ķ—Ā—É—Ä—Ā–ĺ–≤, –ľ–ĺ–≥—É—ā –Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ —Ä–Ķ–į–Ľ–ł–∑–ĺ–≤—č–≤–į—ā—Ć—Ā—Ź –ī–į–∂–Ķ –≤¬†—É—Ā–Ľ–ĺ–≤–ł—Ź—Ö, –ļ–ĺ–≥–ī–į —á–į—Ā—ā—Ć –ļ–ĺ—Ä–ļ–ĺ–≤—č—Ö –∑–ĺ–Ĺ —É–∂–Ķ —Ā–Ņ–ł—ā. –ě–ī–Ĺ–į–ļ–ĺ –≤¬†—Ā–ł—ā—É–į—Ü–ł—Ź—Ö, –ļ–ĺ–≥–ī–į –≤—Ā—é –ľ–ĺ—Č—Ć –ļ–ĺ—Ä—č –ľ–ĺ–∑–≥–į –Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ–ĺ –∑–į–ī–Ķ–Ļ—Ā—ā–≤–ĺ–≤–į—ā—Ć –ī–Ľ—Ź –Ņ—Ä–ł–Ĺ—Ź—ā–ł—Ź —Ā–Ľ–ĺ–∂–Ĺ—č—Ö —Ä–Ķ—ą–Ķ–Ĺ–ł–Ļ, –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ—č–Ļ —Ā–ĺ–Ĺ –ľ–ĺ–∂–Ķ—ā –Ņ—Ä–ł–≤–Ķ—Ā—ā–ł –ļ¬†—Ā–Ķ—Ä—Ć–Ķ–∑–Ĺ—č–ľ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź–ľ.
–£—Ā–Ľ–ĺ–≤–ł—Ź –≤–ĺ–∑–Ĺ–ł–ļ–Ĺ–ĺ–≤–Ķ–Ĺ–ł—Ź –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į —É–ļ–į–∑—č–≤–į—é—ā –Ĺ–į —ā–ĺ, —á—ā–ĺ –ĺ–Ĺ –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –≤–Ķ—Ä–ĺ—Ź—ā–Ķ–Ĺ –≤¬†—Ā–ł—ā—É–į—Ü–ł—Ź—Ö, –ļ–ĺ–≥–ī–į –Ņ—Ä–ł—Ö–ĺ–ī–ł—ā—Ā—Ź –Ņ–ĺ–ī–ī–Ķ—Ä–∂–ł–≤–į—ā—Ć –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ –Ĺ–į —Ą–ĺ–Ĺ–Ķ –≤—č—Ā–ĺ–ļ–ĺ–≥–ĺ –ī–į–≤–Ľ–Ķ–Ĺ–ł—Ź —Ā–Ĺ–į. –Ē–Ľ—Ź –Ľ—é–ī–Ķ–Ļ —ć—ā–ĺ –ľ–ĺ–∂–Ķ—ā –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā—Ć –Ņ—Ä–ł —Ä–į–Ī–ĺ—ā–Ķ –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī—č –ľ–į–ļ—Ā–ł–ľ–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ā–ĺ–Ĺ–Ľ–ł–≤–ĺ—Ā—ā–ł.
–£ –Ľ—é–ī–Ķ–Ļ —Ā¬†–Ņ—Ä–ł–≤—č—á–Ĺ—č–ľ –ł–Ľ–ł –≤—č–Ĺ—É–∂–ī–Ķ–Ĺ–Ĺ—č–ľ —Ā–ĺ–ļ—Ä–į—Č–Ķ–Ĺ–Ĺ—č–ľ –Ĺ–ĺ—á–Ĺ—č–ľ —Ā–Ĺ–ĺ–ľ –ĺ—á–Ķ–Ĺ—Ć –≤—č—Ā–ĺ–ļ —Ä–ł—Ā–ļ —Ä–į–∑–≤–ł—ā–ł—Ź –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į. –Ē–Ľ—Ź –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö –≤–ł–ī–ĺ–≤ –ī–Ķ—Ź—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ—č–Ļ —Ā–ĺ–Ĺ, –≤—č–∑–≤–į–Ĺ–Ĺ—č–Ļ —Ö—Ä–ĺ–Ĺ–ł—á–Ķ—Ā–ļ–ĺ–Ļ –ī–Ķ–Ņ—Ä–ł–≤–į—Ü–ł–Ķ–Ļ —Ā–Ĺ–į, –ľ–ĺ–∂–Ķ—ā –ł¬†–Ĺ–Ķ —Ā–ĺ–∑–ī–į–≤–į—ā—Ć –ĺ—Ā–ĺ–Ī—č—Ö —ā—Ä—É–ī–Ĺ–ĺ—Ā—ā–Ķ–Ļ. –ě–ī–Ĺ–į–ļ–ĺ –ī–Ľ—Ź –ī–Ķ—Ź—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł, —Ā–≤—Ź–∑–į–Ĺ–Ĺ–ĺ–Ļ —Ā¬†–Ņ—Ä–ł–Ĺ—Ź—ā–ł–Ķ–ľ —Ā–Ľ–ĺ–∂–Ĺ—č—Ö –ł¬†–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–Ķ–Ĺ–Ĺ—č—Ö —Ä–Ķ—ą–Ķ–Ĺ–ł–Ļ, –ĺ—Ā–ĺ–Ī–Ķ–Ĺ–Ĺ–ĺ –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī—č –Ĺ–ĺ—á–Ĺ—č—Ö —Ā–ľ–Ķ–Ĺ, –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź –≤–ĺ–∑–Ĺ–ł–ļ–Ĺ–ĺ–≤–Ķ–Ĺ–ł—Ź –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į –ľ–ĺ–≥—É—ā –Ī—č—ā—Ć –Ņ–Ķ—á–į–Ľ—Ć–Ĺ—č, –Ņ–ĺ—Ā–ļ–ĺ–Ľ—Ć–ļ—É —É–≤–Ķ–Ľ–ł—á–ł–≤–į–Ķ—ā—Ā—Ź —Ä–ł—Ā–ļ –Ņ—Ä–ł–Ĺ—Ź—ā–ł—Ź –ĺ—ą–ł–Ī–ĺ—á–Ĺ—č—Ö —Ä–Ķ—ą–Ķ–Ĺ–ł–Ļ.
–ö—Ä–ĺ–ľ–Ķ –ĺ—ā—Ä–ł—Ü–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ –≤–Ľ–ł—Ź–Ĺ–ł—Ź —á–į—Ā—ā–ł—á–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į –Ĺ–į –ļ–ĺ–≥–Ĺ–ł—ā–ł–≤–Ĺ—č–Ķ —Ą—É–Ĺ–ļ—Ü–ł–ł, –Ĺ—É–∂–Ĺ–ĺ —É—á–ł—ā—č–≤–į—ā—Ć –ł¬†–≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č–Ķ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź —ć—ā–ĺ–≥–ĺ —Ź–≤–Ľ–Ķ–Ĺ–ł—Ź –ī–Ľ—Ź –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –∑–ī–ĺ—Ä–ĺ–≤—Ć—Ź. –ö–į–ļ –ī–Ľ—Ź —É—Ā–Ņ–Ķ—ą–Ĺ–ĺ–≥–ĺ —Ä–Ķ—ą–Ķ–Ĺ–ł—Ź —Ā–Ľ–ĺ–∂–Ĺ—č—Ö –Ņ—Ä–ĺ–Ī–Ľ–Ķ–ľ –≤¬†–Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–ł –≤–į–∂–Ĺ–ĺ —É—á–į—Ā—ā–ł–Ķ –≤—Ā–Ķ—Ö –ļ–ĺ—Ä–ļ–ĺ–≤—č—Ö –∑–ĺ–Ĺ, —ā–į–ļ –≤–į–∂–Ĺ–ĺ –ł¬†–≤–ĺ–≤–Ľ–Ķ—á–Ķ–Ĺ–ł–Ķ –≤—Ā–Ķ—Ö –ļ–ĺ—Ä–ļ–ĺ–≤—č—Ö –∑–ĺ–Ĺ –≤¬†–į–Ĺ–į–Ľ–ł–∑ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł –ī–Ľ—Ź —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ–≥–ĺ —É–Ņ—Ä–į–≤–Ľ–Ķ–Ĺ–ł—Ź –≤—Ā–Ķ–ľ–ł –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–ľ–ł —Ā–ł—Ā—ā–Ķ–ľ–į–ľ–ł. –ü–ĺ—Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī—č –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ľ–ĺ–∂–Ķ—ā —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤–ĺ–≤–į—ā—Ć –嬆–Ĺ–Ķ–ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ–Ļ –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł –ĺ–Ī—Č–Ķ–≥–ĺ —Ā–Ĺ–į –≤¬†–Ĺ–ĺ—á–Ĺ–ĺ–Ķ –≤—Ä–Ķ–ľ—Ź –ł–Ľ–ł —É–ļ–į–∑—č–≤–į—ā—Ć –Ĺ–į –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ—č–Ķ —Ā–ļ—Ä—č—ā—č–Ķ –Ņ—Ä–ĺ–Ī–Ľ–Ķ–ľ—č –≤¬†–≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ā—Ą–Ķ—Ä–Ķ.
–†–į—Ā—Ā–ľ–ĺ—ā—Ä–Ķ–Ĺ–Ĺ—č–Ķ –≤—č—ą–Ķ –≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ—Ā—ā–ł –į—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–Ĺ–ĺ–≥–ĺ —Ä–į–∑–≤–ł—ā–ł—Ź —Ā–Ĺ–į –≤¬†—Ä–į–∑–Ĺ—č—Ö –ĺ—ā–ī–Ķ–Ľ–į—Ö –ļ–ĺ—Ä—č –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ—Ź–Ľ–ł –≤–į—Ä–ł–į–Ĺ—ā—č –Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –ľ–ĺ–∑–≥–į. –°–≤—Ź–∑–į–Ĺ–Ĺ—č–Ķ —Ā¬†—ć—ā–ł–ľ —Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ–ľ –Ĺ–Ķ–≥–į—ā–ł–≤–Ĺ—č–Ķ –ī–Ľ—Ź –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–ĺ–≤ –ł¬†–ĺ–ļ—Ä—É–∂–Ķ–Ĺ–ł—Ź –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź –ĺ—ā—Ä–į–∂–į–Ľ–ł, —Ā–ļ–ĺ—Ä–Ķ–Ķ, –Ĺ–į—Ä—É—ą–Ķ–Ĺ–ł–Ķ ¬ę–Ņ—Ä–į–≤–ł–Ľ —ć–ļ—Ā–Ņ–Ľ—É–į—ā–į—Ü–ł–ł¬Ľ, —ā–ĺ –Ķ—Ā—ā—Ć —ā–ĺ, —á—ā–ĺ –Ņ—Ä–ł–Ĺ—Ź—ā–ĺ –Ĺ–į–∑—č–≤–į—ā—Ć —á–Ķ–Ľ–ĺ–≤–Ķ—á–Ķ—Ā–ļ–ł–ľ —Ą–į–ļ—ā–ĺ—Ä–ĺ–ľ.
–ě—Ä–≥–į–Ĺ–ł–∑–į—Ü–ł—Ź –ĺ—Ā–Ĺ–ĺ–≤–Ĺ—č—Ö –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č—Ö –Ņ–ĺ—ā–ĺ–ļ–ĺ–≤ –≤¬†–ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–Ķ
–í–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–į—Ź —ā–Ķ–ĺ—Ä–ł—Ź —Ā–Ĺ–į –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–į–≥–į–Ķ—ā –ł¬†–Ĺ–ĺ–≤—č–Ķ –Ņ–ĺ–ī—Ö–ĺ–ī—č –ļ¬†–Ņ–ĺ–Ĺ–ł–ľ–į–Ĺ–ł—é –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–ĺ–≤ –≤–ĺ–∑–Ĺ–ł–ļ–Ĺ–ĺ–≤–Ķ–Ĺ–ł—Ź —Ä—Ź–ī–į –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ļ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į, —Ā–≤—Ź–∑–į–Ĺ–Ĺ—č—Ö —ā–į–ļ–∂–Ķ —Ā¬†–Ĺ–į—Ä—É—ą–Ķ–Ĺ–ł–Ķ–ľ —Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–Ĺ–ĺ—Ā—ā–ł —Ä–į–Ī–ĺ—ā—č —Ā—ā—Ä—É–ļ—ā—É—Ä, —É—á–į—Ā—ā–≤—É—é—Č–ł—Ö –≤¬†—É–Ņ—Ä–į–≤–Ľ–Ķ–Ĺ–ł–ł –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č–ľ–ł –Ņ–ĺ—ā–ĺ–ļ–į–ľ–ł –≤¬†—Ü–ł–ļ–Ľ–Ķ ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ.
–†–į—Ā—Ā–ľ–ĺ—ā—Ä–ł–ľ –≤¬†—Ä–į–ľ–ļ–į—Ö –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —ā–Ķ–ĺ—Ä–ł–ł —Ā–Ĺ–į –ĺ—Ā–Ĺ–ĺ–≤–Ĺ—č–Ķ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č–Ķ –Ņ–ĺ—ā–ĺ–ļ–ł –≤¬†–∑–ī–ĺ—Ä–ĺ–≤–ĺ–ľ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–Ķ –≤¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ł¬†—Ā–Ĺ–į, –ļ–ĺ—ā–ĺ—Ä—č–Ķ —Ā—Ö–Ķ–ľ–į—ā–ł—á–Ķ—Ā–ļ–ł –ĺ—ā—Ä–į–∂–Ķ–Ĺ—č –Ĺ–į —Ä–ł—Ā—É–Ĺ–ļ–į—Ö 2 –ł¬†3. –ě—ā–ļ—Ä—č—ā—č–Ķ –Ņ—É—ā–ł –ī–Ľ—Ź –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł—Ź —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤ –Ņ–ĺ–ļ–į–∑–į–Ĺ—č —á–Ķ—Ä–Ĺ—č–ľ —Ü–≤–Ķ—ā–ĺ–ľ, –Ņ—É—ā–ł, –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤ –Ņ–ĺ –ļ–ĺ—ā–ĺ—Ä—č–ľ –Ņ—Ä–Ķ–ļ—Ä–į—Č–Ķ–Ĺ–ĺ,¬†‚Äď –Ņ—É–Ĺ–ļ—ā–ł—Ä–ĺ–ľ. –°—ā—Ä—É–ļ—ā—É—Ä—č, –Ī–Ľ–ĺ–ļ–ł—Ä—É—é—Č–ł–Ķ –Ņ–Ķ—Ä–Ķ–ī–į—á—É —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤, –ł–∑–ĺ–Ī—Ä–į–∂–Ķ–Ĺ—č –≤¬†–≤–ł–ī–Ķ –ļ—Ä—É–∂–ļ–ĺ–≤. –ü—Ä—Ź–ľ–ĺ—É–≥–ĺ–Ľ—Ć–Ĺ–ł–ļ–ł –≤–Ĺ—É—ā—Ä–ł –ļ—Ä—É–∂–ļ–ĺ–≤ –ĺ—ā—Ä–į–∂–į—é—ā —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ķ —Ā—ā—Ä—É–ļ—ā—É—Ä: –ļ–ĺ–≥–ī–į –ĺ–Ĺ–ł —Ä–į—Ā–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ—č –≤–ī–ĺ–Ľ—Ć –Ņ—Ä–ĺ–≤–ĺ–ī—Ź—Č–ł—Ö –Ņ—É—ā–Ķ–Ļ, –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ –ĺ—ā–ļ—Ä—č—ā–ĺ, –į¬†–Ņ–ĺ–Ņ–Ķ—Ä–Ķ–ļ¬†‚Äď –∑–į–ļ—Ä—č—ā–ĺ. –ö–į–∂–ī—č–Ļ –Ī–Ľ–ĺ–ļ –ł–ľ–Ķ–Ķ—ā –≤—ā–ĺ—Ä–ĺ–Ļ –≤—Ö–ĺ–ī, –Ņ–ĺ–ļ–į–∑–į–Ĺ–Ĺ—č–Ļ —Ā—ā—Ä–Ķ–Ľ–ļ–ĺ–Ļ. –≠—ā–ĺ –≤—Ö–ĺ–ī —É–Ņ—Ä–į–≤–Ľ—Ź—é—Č–Ķ–≥–ĺ —Ā–ł–≥–Ĺ–į–Ľ–į, –Ņ–ĺ –ļ–ĺ–ľ–į–Ĺ–ī–Ķ –ļ–ĺ—ā–ĺ—Ä–ĺ–≥–ĺ –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā –ĺ—ā–ļ—Ä—č—ā–ł–Ķ –ł–Ľ–ł –∑–į–ļ—Ä—č—ā–ł–Ķ –Ņ—É—ā–Ķ–Ļ. –ü–ĺ–ī–ĺ–Ī–Ĺ—č–Ķ —Ā—ā—Ä—É–ļ—ā—É—Ä—č –ł–∑–≤–Ķ—Ā—ā–Ĺ—č –≤¬†–Ĺ–Ķ–Ļ—Ä–ĺ—Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł–ł, –Ĺ–į–Ņ—Ä–ł–ľ–Ķ—Ä —ā—Ä–ł–į–ī–Ĺ—č–Ķ —Ā–ł–Ĺ–į–Ņ—Ā—č –ł–Ľ–ł —Ā–ł—Ā—ā–Ķ–ľ—č –Ņ—Ä–Ķ—Ā–ł–Ĺ–į–Ņ—ā–ł—á–Ķ—Ā–ļ–ĺ–≥–ĺ —ā–ĺ—Ä–ľ–ĺ–∂–Ķ–Ĺ–ł—Ź. –ě–Ņ–ł—Ā–į–Ĺ–Ĺ—č–Ķ –ľ–ĺ–ī—É–Ľ—Ź—ā–ĺ—Ä—č —Ā–ł–Ĺ–į–Ņ—ā–ł—á–Ķ—Ā–ļ–ĺ–Ļ –Ņ–Ķ—Ä–Ķ–ī–į—á–ł –Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č —ā–ĺ–∂–Ķ, —Ā–ļ–ĺ—Ä–Ķ–Ķ –≤—Ā–Ķ–≥–ĺ, —Ź–≤–Ľ—Ź—é—ā—Ā—Ź —ć–Ľ–Ķ–ľ–Ķ–Ĺ—ā–į–ľ–ł —ā–į–ļ–ł—Ö —É–Ņ—Ä–į–≤–Ľ—Ź—é—Č–ł—Ö —Ā–ł—Ā—ā–Ķ–ľ.
–Ě–į —Ä–ł—Ā—É–Ĺ–ļ–Ķ 2 –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ĺ —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ķ –Ņ—Ä–ĺ–≤–ĺ–ī—Ź—Č–ł—Ö –Ņ—É—ā–Ķ–Ļ –Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č –≤¬†–Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–ł. –°–ł–≥–Ĺ–į–Ľ—č –ł–∑ –ĺ–ļ—Ä—É–∂–į—é—Č–Ķ–Ļ —Ā—Ä–Ķ–ī—č –ł–Ľ–ł –嬆–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–ł–ł —ā–Ķ–Ľ–į –∂–ł–≤–ĺ—ā–Ĺ–ĺ–≥–ĺ, –Ņ—Ä–Ķ–ĺ–Ī—Ä–į–∑–ĺ–≤–į–Ĺ–Ĺ—č–Ķ —ć–ļ—Ā—ā–Ķ—Ä–ĺ- –ł¬†–Ņ—Ä–ĺ–Ņ—Ä–ł–ĺ—Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–į–ľ–ł –≤¬†–Ĺ–Ķ—Ä–≤–Ĺ—č–Ķ –ł–ľ–Ņ—É–Ľ—Ć—Ā—č, –Ņ—Ä–ĺ—Ö–ĺ–ī—Ź—ā —á–Ķ—Ä–Ķ–∑ –ĺ—ā–ļ—Ä—č—ā–ĺ–Ķ –Ī–Ľ–ĺ–ļ–ł—Ä—É—é—Č–Ķ–Ķ —É—Ā—ā—Ä–ĺ–Ļ—Ā—ā–≤–ĺ –ł¬†–Ĺ–į–Ņ—Ä–į–≤–Ľ—Ź—é—ā—Ā—Ź –ī–Ľ—Ź –į–Ĺ–į–Ľ–ł–∑–į –≤¬†—Ü–Ķ–Ĺ—ā—Ä–į–Ľ—Ć–Ĺ—č–Ļ –Ī–Ľ–ĺ–ļ (¬ę–ö–ĺ—Ä–į –ľ–ĺ–∑–≥–į¬Ľ). –†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –≤—č–Ņ–ĺ–Ľ–Ĺ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –∑–ī–Ķ—Ā—Ć –į–Ĺ–į–Ľ–ł–∑–į –Ņ–Ķ—Ä–Ķ–ī–į—é—ā—Ā—Ź –≤¬†–Ī–Ľ–ĺ–ļ, –ĺ—ā–≤–Ķ—á–į—é—Č–ł–Ļ –∑–į –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ –ł¬†–ī–≤–ł–≥–į—ā–Ķ–Ľ—Ć–Ĺ—É—é –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć. –ü–į—Ä–į–Ľ–Ľ–Ķ–Ľ—Ć–Ĺ–ĺ –≤—č—Ö–ĺ–ī—Ź—Č–ł–Ķ –ł–∑ –ļ–ĺ—Ä—č —Ā–ł–≥–Ĺ–į–Ľ—č –Ņ–Ķ—Ä–Ķ–ī–į—é—ā—Ā—Ź –≤¬†–Ī–Ľ–ĺ–ļ ¬ę–°–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ¬Ľ, –į–ļ—ā–ł–≤–į—Ü–ł—Ź –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤ –ļ–ĺ—ā–ĺ—Ä–ĺ–≥–ĺ –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†–Ņ–ĺ—Ź–≤–Ľ–Ķ–Ĺ–ł—é –ĺ—Č—É—Č–Ķ–Ĺ–ł–Ļ —Ā–Ķ–Ī—Ź –ł¬†–ĺ–ļ—Ä—É–∂–į—é—Č–Ķ–≥–ĺ –ľ–ł—Ä–į. –Ę—Ä–į–ī–ł—Ü–ł–ĺ–Ĺ–Ĺ–ĺ —Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ —Ā–≤—Ź–∑—č–≤–į—é—ā —Ā¬†–į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć—é –ļ–ĺ—Ä—č –ľ–ĺ–∑–≥–į. –ě–ī–Ĺ–į–ļ–ĺ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź —Ā–Ĺ–į –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ–ĺ —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤—É—é—ā, —á—ā–ĺ —ć—ā–ĺ –Ĺ–Ķ —ā–į–ļ. –í–ĺ–Ņ—Ä–ĺ—Ā –嬆–≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ–Ļ –Ľ–ĺ–ļ–į–Ľ–ł–∑–į—Ü–ł–ł ¬ę—Ā—ā—Ä—É–ļ—ā—É—Ä —Ā–ĺ–∑–Ĺ–į–Ĺ–ł—Ź¬Ľ –≤¬†–ļ–ĺ–Ĺ—ā–Ķ–ļ—Ā—ā–Ķ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–ĺ–≤ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź —Ā–Ĺ–į —É–∂–Ķ —Ä–į—Ā—Ā–ľ–ĺ—ā—Ä–Ķ–Ĺ [19], –ł¬†–ľ—č –Ĺ–Ķ –Ī—É–ī–Ķ–ľ –Ĺ–į –Ĺ–Ķ–ľ –ĺ—Ā—ā–į–Ĺ–į–≤–Ľ–ł–≤–į—ā—Ć—Ā—Ź.
–í–ĺ –≤—Ä–Ķ–ľ—Ź –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł—Ź –嬆—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –≤–Ĺ—É—ā—Ä–Ķ–Ĺ–Ĺ–ł—Ö –ĺ—Ä–≥–į–Ĺ–ĺ–≤, –Ņ—Ä–Ķ–ĺ–Ī—Ä–į–∑–ĺ–≤–į–Ĺ–Ĺ–į—Ź –ł–Ĺ—ā–Ķ—Ä–ĺ—Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–į–ľ–ł –≤¬†–Ĺ–Ķ—Ä–≤–Ĺ—č–Ķ –ł–ľ–Ņ—É–Ľ—Ć—Ā—č, –Ņ–ĺ—Ā—ā—É–Ņ–į–Ķ—ā –≤¬†—Ā—ā—Ä—É–ļ—ā—É—Ä—č –į–≤—ā–ĺ–Ĺ–ĺ–ľ–Ĺ–ĺ–Ļ –Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č. –ö–ĺ–Ĺ—ā—Ä–ĺ–Ľ—Ć –≤–Ĺ—É—ā—Ä–Ķ–Ĺ–Ĺ–ł—Ö –ĺ—Ä–≥–į–Ĺ–ĺ–≤ –≤¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ĺ—Ā—É—Č–Ķ—Ā—ā–≤–Ľ—Ź–Ķ—ā—Ā—Ź –Ņ–ĺ –Ľ–ł–Ĺ–ł—Ź–ľ –ĺ–Ī—Ä–į—ā–Ĺ—č—Ö –Ņ—Ä–ĺ–Ķ–ļ—Ü–ł–Ļ¬†‚Äď –ĺ—ā –į–≤—ā–ĺ–Ĺ–ĺ–ľ–Ĺ–ĺ–Ļ –Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č, –ļ–ĺ—ā–ĺ—Ä–į—Ź –ĺ—Ä–≥–į–Ĺ–ł–∑–ĺ–≤–į–Ĺ–į —Ā–Ķ–≥–ľ–Ķ–Ĺ—ā–į—Ä–Ĺ–ĺ, —á—ā–ĺ –ł¬†–Ņ—Ä–Ķ–Ņ—Ź—ā—Ā—ā–≤—É–Ķ—ā –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—é —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤ –ł–∑ —Ä–į–∑–Ĺ—č—Ö –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ā–ł—Ā—ā–Ķ–ľ [23]. –Ě–Ķ—Ā–ľ–ĺ—ā—Ä—Ź –Ĺ–į —ć—ā–ĺ, –į–≤—ā–ĺ–Ĺ–ĺ–ľ–Ĺ–į—Ź –Ĺ–Ķ—Ä–≤–Ĺ–į—Ź —Ā–ł—Ā—ā–Ķ–ľ–į –ľ–ĺ–∂–Ķ—ā –Ņ–ĺ–ī–ī–Ķ—Ä–∂–ł–≤–į—ā—Ć –≥–ĺ–ľ–Ķ–ĺ—Ā—ā–į–∑ –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –ĺ–≥—Ä–į–Ĺ–ł—á–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –Ņ–Ķ—Ä–ł–ĺ–ī–į.
–ě–ī–Ĺ–į–ļ–ĺ —Ā–ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ķ–ľ –ł–Ĺ—ā–Ķ—Ä–ĺ—Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä—č –Ĺ–į—á–ł–Ĺ–į—é—ā –Ņ–Ķ—Ä–Ķ–ī–į–≤–į—ā—Ć —Ā–ł–≥–Ĺ–į–Ľ—č –ĺ–Ī –ĺ—ā–ļ–Ľ–ĺ–Ĺ–Ķ–Ĺ–ł–ł —ā–Ķ–ļ—É—Č–ł—Ö –Ņ–į—Ä–į–ľ–Ķ—ā—Ä–ĺ–≤ –ĺ—ā –≥–Ķ–Ĺ–Ķ—ā–ł—á–Ķ—Ā–ļ–ł –∑–į–ī–į–Ĺ–Ĺ—č—Ö –Ĺ–ĺ—Ä–ľ. –í–ł–ī–ł–ľ–ĺ, –ł–ľ–Ķ–Ĺ–Ĺ–ĺ —ć—ā–ł —Ā–ł–≥–Ĺ–į–Ľ—č –≤–ĺ—Ā–Ņ—Ä–ł–Ĺ–ł–ľ–į—é—ā—Ā—Ź –∂–ł–≤–ĺ—ā–Ĺ—č–ľ–ł –ļ–į–ļ —á—É–≤—Ā—ā–≤–ĺ —É—Ā—ā–į–Ľ–ĺ—Ā—ā–ł –ł¬†–∑–į—Ā—ā–į–≤–Ľ—Ź—é—ā –ł—Ö –ł—Ā–ļ–į—ā—Ć –Ī–Ķ–∑–ĺ–Ņ–į—Ā–Ĺ–ĺ–Ķ –ł¬†—É–ī–ĺ–Ī–Ĺ–ĺ–Ķ –ľ–Ķ—Ā—ā–ĺ –ī–Ľ—Ź —Ā–Ĺ–į. –Ď–ĺ–Ľ–Ķ–Ķ –ī–Ķ—ā–į–Ľ—Ć–Ĺ–ĺ –ī–ł–Ĺ–į–ľ–ł–ļ–į –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–į –ļ–ĺ —Ā–Ĺ—É –≤¬†—Ā–≤–Ķ—ā–Ķ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —ā–Ķ–ĺ—Ä–ł–ł —Ä–į—Ā—Ā–ľ–ĺ—ā—Ä–Ķ–Ĺ–į —Ä–į–Ĺ–Ķ–Ķ [20].
–í–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā —Ä–į–ī–ł–ļ–į–Ľ—Ć–Ĺ–ĺ–Ķ –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł–Ķ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č—Ö –Ņ–ĺ—ā–ĺ–ļ–ĺ–≤ –≤¬†–Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ–Ķ (—Ä–ł—Ā. 2). –í¬†–ł–ī–Ķ–į–Ľ–Ķ –Ņ–ĺ—Ā–Ľ–Ķ –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ–ł—Ź —Ä–į–∑—Ä–Ķ—ą–Ķ–Ĺ–ł—Ź –Ĺ–į —Ā–ĺ–Ĺ –≤—Ā–Ķ –Ī–Ľ–ĺ–ļ–ł—Ä—É—é—Č–ł–Ķ —Ā–ł—Ā—ā–Ķ–ľ—č –ĺ–ī–Ĺ–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ –ľ–Ķ–Ĺ—Ź—é—ā –Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–ł–Ķ. –í–ĺ —Ā–Ĺ–Ķ –Ņ—Ä–Ķ–ļ—Ä–į—Č–į–Ķ—ā—Ā—Ź –Ņ–Ķ—Ä–Ķ–ī–į—á–į —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤ –ĺ—ā —ć–ļ—Ā—ā–Ķ—Ä–ĺ- –ł¬†–Ņ—Ä–ĺ–Ņ—Ä–ł–ĺ—Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–ĺ–≤ –≤¬†–ļ–ĺ—Ä—É –ľ–ĺ–∑–≥–į. –Ě–ĺ –Ņ–ĺ —ā–Ķ–ľ –∂–Ķ –≤—Ö–ĺ–ī–Ĺ—č–ľ –ļ–į–Ĺ–į–Ľ–į–ľ –≤¬†–ļ–ĺ—Ä—É –Ĺ–į—á–ł–Ĺ–į–Ķ—ā –Ņ–ĺ—Ā—ā—É–Ņ–į—ā—Ć –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł—Ź –ł–∑ –į–≤—ā–ĺ–Ĺ–ĺ–ľ–Ĺ–ĺ–Ļ –Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č –ĺ¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –≤—Ā–Ķ—Ö –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ā–ł—Ā—ā–Ķ–ľ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į.
–°–ļ–ĺ—Ä–Ķ–Ķ –≤—Ā–Ķ–≥–ĺ, –ĺ—Ā–Ĺ–ĺ–≤–Ĺ–ĺ–Ļ —Ā—ā—Ä—É–ļ—ā—É—Ä–ĺ–Ļ, –≤¬†–ļ–ĺ—ā–ĺ—Ä–ĺ–Ļ –ĺ—Ā—É—Č–Ķ—Ā—ā–≤–Ľ—Ź–Ķ—ā—Ā—Ź –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ –ļ–ĺ—Ä–ļ–ĺ–≤–ĺ–Ļ –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł–ł —Ā¬†—ć–ļ—Ā—ā–Ķ—Ä–ĺ- –ł¬†–Ņ—Ä–ĺ–Ņ—Ä–ł–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ—č—Ö –≤—Ö–ĺ–ī–ĺ–≤ –Ĺ–į –ł–Ĺ—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ—č–Ķ, —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź —ā–į–Ľ–į–ľ—É—Ā. –í–Ķ—Ā—Ć–ľ–į –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ, —á—ā–ĺ —ć—ā–ĺ –ł¬†–Ķ—Ā—ā—Ć –Ķ–≥–ĺ –ĺ—Ā–Ĺ–ĺ–≤–Ĺ–į—Ź —Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–į–Ľ—Ć–Ĺ–į—Ź –Ĺ–į–≥—Ä—É–∑–ļ–į. –ü—Ä–ł —Ā–ľ–Ķ–Ĺ–Ķ –ļ–ĺ—Ä–ļ–ĺ–≤–ĺ–Ļ –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł–ł —Ā¬†—ć–ļ—Ā—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ–ĺ–Ļ –Ĺ–į –ł–Ĺ—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ—É—é –Ķ—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –ĺ–∂–ł–ī–į—ā—Ć –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł–Ļ –ł¬†–≤¬†—ć—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–Ĺ—č—Ö –ļ–ĺ—Ä–ļ–ĺ–≤—č—Ö –Ņ—Ä–ĺ–Ķ–ļ—Ü–ł—Ź—Ö. –ü–ĺ—ā–ĺ–ļ–ł –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł, –≤—č—Ö–ĺ–ī—Ź—Č–ł–Ķ –ł–∑ –ļ–ĺ—Ä—č –≤–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į, –ĺ—ā—Ä–į–∂–į—é—ā —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –ĺ–Ī—Ä–į–Ī–ĺ—ā–ļ–ł –ļ–ĺ—Ä–ĺ–Ļ –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł, –ł¬†–ĺ–Ĺ–ł –Ĺ–Ķ –ī–ĺ–Ľ–∂–Ĺ—č –Ĺ–į–Ņ—Ä–į–≤–Ľ—Ź—ā—Ć—Ā—Ź –≤¬†—Ā—ā—Ä—É–ļ—ā—É—Ä—č, —Ā–≤—Ź–∑–į–Ĺ–Ĺ—č–Ķ —Ā¬†–ī–≤–ł–≥–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć—é, –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ–ľ –ł¬†—Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ–ľ. –Ě–į —Ā—Ö–Ķ–ľ–Ķ –Ī–Ľ–ĺ–ļ–ł—Ä—É—é—Č–ł–Ķ —Ā–ł—Ā—ā–Ķ–ľ—č –Ĺ–į —ć—ā–ł—Ö –Ņ—É—ā—Ź—Ö –≤–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į –∑–į–ļ—Ä—č—ā—č.
–§–į–ļ—ā –ĺ—ā–ļ–Ľ—é—á–Ķ–Ĺ–ł—Ź —Ā–ĺ–∑–Ĺ–į–Ĺ–ł—Ź –ĺ—ā –≤–Ĺ–Ķ—ą–Ĺ–Ķ–≥–ĺ –ľ–ł—Ä–į –≤–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į —Ö–ĺ—Ä–ĺ—ą–ĺ –ł–∑–≤–Ķ—Ā—ā–Ķ–Ĺ –ļ–į–∂–ī–ĺ–ľ—É –ł–∑ —Ā–ĺ–Ī—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –ĺ–Ņ—č—ā–į. –ö—Ä–ĺ–ľ–Ķ —ā–ĺ–≥–ĺ, –ł–∑ –į–Ĺ–į—ā–ĺ–ľ–ł–ł –ł¬†—Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł–ł –ł–∑–≤–Ķ—Ā—ā–Ĺ–ĺ, —á—ā–ĺ –≤–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į –į–ļ—ā–ł–≤–Ĺ–ĺ –Ī–Ľ–ĺ–ļ–ł—Ä—É–Ķ—ā—Ā—Ź –Ņ–Ķ—Ä–Ķ–ī–į—á–į —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤ –ł–∑ –ļ–ĺ—Ä—č –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į –Ĺ–į –ľ–ĺ—ā–ĺ–Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ—č —Ā–Ņ–ł–Ĺ–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į. –≠—ā–ĺ –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†—Ä–į—Ā—Ā–Ľ–į–Ī–Ľ–Ķ–Ĺ–ł—é –ľ—č—ą—Ü —ā–Ķ–Ľ–į, –ļ–ĺ—ā–ĺ—Ä–ĺ–Ķ –ī–ĺ—Ā—ā–ł–≥–į–Ķ—ā –ľ–į–ļ—Ā–ł–ľ—É–ľ–į –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī—č –Ī—č—Ā—ā—Ä–ĺ–≥–ĺ —Ā–Ĺ–į¬†‚Äď —ā–į–ļ –Ĺ–į–∑—č–≤–į–Ķ–ľ–į—Ź –į—ā–ĺ–Ĺ–ł—Ź —Ā–Ĺ–į [24].
–ě–ī–Ĺ–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ –≤–ĺ —Ā–Ĺ–Ķ –ī–ĺ–Ľ–∂–Ĺ–ĺ –ĺ—ā–ļ—Ä—č–≤–į—ā—Ć—Ā—Ź –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ –Ņ–ĺ –Ĺ–ĺ–≤–ĺ–ľ—É –Ņ—É—ā–ł –ł–∑ –ļ–ĺ—Ä—č –ľ–ĺ–∑–≥–į –Ĺ–į —Ā—ā—Ä—É–ļ—ā—É—Ä—č, —Ā–≤—Ź–∑–į–Ĺ–Ĺ—č–Ķ —Ā¬†–į—Ā—Ā–ĺ—Ü–ł–į—ā–ł–≤–Ĺ–ĺ–Ļ –ĺ–Ī—Ä–į–Ī–ĺ—ā–ļ–ĺ–Ļ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł, –≥–ī–Ķ –∑–į–ľ—č–ļ–į–Ķ—ā—Ā—Ź –Ņ–Ķ—ā–Ľ—Ź —É–Ņ—Ä–į–≤–Ľ–Ķ–Ĺ–ł—Ź –≤—Ā–Ķ–ľ–ł –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–ľ–ł —Ā–ł—Ā—ā–Ķ–ľ–į–ľ–ł –≤–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į. –ú–ĺ–∂–Ĺ–ĺ –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–ł—ā—Ć, —á—ā–ĺ —Ü–Ķ–Ĺ—ā—Ä–ĺ–ľ –į—Ā—Ā–ĺ—Ü–ł–į—ā–ł–≤–Ĺ–ĺ–Ļ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –≥–ł–Ņ–ĺ—ā–į–Ľ–į–ľ—É—Ā.
–ö–ĺ–≥–ī–į –≤—Ā–Ķ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–Ķ –Ņ–į—Ä–į–ľ–Ķ—ā—Ä—č –≤¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –≤–ļ–Ľ—é—á–Ķ–Ĺ–ł—Ź –ļ–ĺ—Ä—č –≤¬†–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł–Ĺ—ā–Ķ–≥—Ä–į—Ü–ł–ł –Ī—É–ī—É—ā –Ņ—Ä–ł–≤–Ķ–ī–Ķ–Ĺ—č –≤¬†–Ĺ–ĺ—Ä–ľ—É, –ļ–ĺ–ľ–į–Ĺ–ī–į –Ĺ–į —Ā–ĺ–Ĺ —Ā–Ĺ–ł–ľ–į–Ķ—ā—Ā—Ź, –≤—Ā–Ķ –Ī–Ľ–ĺ–ļ–ł—Ä—É—é—Č–ł–Ķ —Ā–ł—Ā—ā–Ķ–ľ—č –ľ–Ķ–Ĺ—Ź—é—ā –Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–ł–Ķ –ł¬†–∂–ł–≤–ĺ—ā–Ĺ–ĺ–Ķ –Ņ—Ä–ĺ–Ī—É–∂–ī–į–Ķ—ā—Ā—Ź.
–Ę–į–ļ –≤—č–≥–Ľ—Ź–ī–ł—ā –ł–ī–Ķ–į–Ľ—Ć–Ĺ–į—Ź –ļ–į—Ä—ā–ł–Ĺ–į –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł—Ź –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č—Ö –Ņ–ĺ—ā–ĺ–ļ–ĺ–≤ –≤¬†—Ü–ł–ļ–Ľ–Ķ ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ. –ě–ī–Ĺ–į–ļ–ĺ –ĺ—Ā–Ĺ–ĺ–≤–Ĺ—č–Ķ —ć–Ľ–Ķ–ľ–Ķ–Ĺ—ā—č —ć—ā–ĺ–Ļ —Ā—Ö–Ķ–ľ—謆‚Äď —Ā–ł—Ā—ā–Ķ–ľ—č, –Ī–Ľ–ĺ–ļ–ł—Ä—É—é—Č–ł–Ķ –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ –Ņ–ĺ —ā–ĺ–ľ—É –ł–Ľ–ł –ł–Ĺ–ĺ–ľ—É –Ņ—É—ā–ł,¬†‚Äď —ć—ā–ĺ –Ĺ–Ķ —ć–Ľ–Ķ–ļ—ā—Ä–ĺ–Ĺ–Ĺ—č–Ķ —Ä–Ķ–Ľ–Ķ, –į¬†—Ö–ł–ľ–ł—á–Ķ—Ā–ļ–ł–Ķ —Ā–ł–Ĺ–į–Ņ—Ā—č. –≠—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć –ł—Ö —Ä–į–Ī–ĺ—ā—č –∑–į–≤–ł—Ā–ł—ā –ĺ—ā –ľ–Ĺ–ĺ–≥–ł—Ö –≤–Ĺ–Ķ—ą–Ĺ–ł—Ö –Ņ–ĺ –ĺ—ā–Ĺ–ĺ—ą–Ķ–Ĺ–ł—é –ļ¬†—ć—ā–ĺ–ľ—É —Ā–ł–Ĺ–į–Ņ—Ā—É —É—Ā–Ľ–ĺ–≤–ł–Ļ.
–Ę–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, –į–Ĺ–į–Ľ–ł–∑–ł—Ä—É—Ź —Ä–į–Ī–ĺ—ā—É –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–Ĺ–ĺ–Ļ —Ā—Ö–Ķ–ľ—č, –Ĺ—É–∂–Ĺ–ĺ –Ņ–ĺ–ľ–Ĺ–ł—ā—Ć, —á—ā–ĺ –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ –Ī–Ľ–ĺ–ļ–ł—Ä—É—é—Č–ł—Ö —Ā–ł—Ā—ā–Ķ–ľ –Ņ—Ä–ł –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö —Ą–į–ļ—ā–ĺ—Ä–į—Ö –ľ–ĺ–∂–Ķ—ā —Ä–į–∑–≤–ł–≤–į—ā—Ć—Ā—Ź –į—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–Ĺ–ĺ –ł¬†—Ä–į—Ā—ā—Ź–Ĺ—É—ā—Ć—Ā—Ź –Ĺ–į –∑–Ĺ–į—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ķ –≤—Ä–Ķ–ľ—Ź. –í¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –Ĺ–į —Ą–ĺ–Ĺ–Ķ –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź —á–į—Ā—ā—Ć —Ā—ā—Ä—É–ļ—ā—É—Ä –ľ–ĺ–∑–≥–į –ľ–ĺ–∂–Ķ—ā –Ī—č—ā—Ć –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–į –≤¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ķ —Ā–Ĺ–į. –ė¬†–Ĺ–į–ĺ–Ī–ĺ—Ä–ĺ—ā, –≤¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł –ĺ–Ī—Č–Ķ–≥–ĺ —Ā–Ĺ–į –ĺ—ā–ī–Ķ–Ľ—Ć–Ĺ—č–Ķ –Ņ—Ä–ĺ–≤–ĺ–ī—Ź—Č–ł–Ķ –Ņ—É—ā–ł –ľ–ĺ–≥—É—ā –ĺ–ļ–į–∑–į—ā—Ć—Ā—Ź –ĺ—ā–ļ—Ä—č—ā—č–ľ–ł, –ļ–į–ļ –≤¬†–Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–ł. –í–ĺ–∑–Ĺ–ł–ļ–į—é—Č–į—Ź –ł–∑-–∑–į —ć—ā–ĺ–≥–ĺ –į—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–Ĺ–ĺ—Ā—ā—Ć –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł—Ź –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č—Ö –Ņ–ĺ—ā–ĺ–ļ–ĺ–≤, –Ī–Ķ–∑—É—Ā–Ľ–ĺ–≤–Ĺ–ĺ, –Ņ—Ä–ł–≤–Ķ–ī–Ķ—ā –ļ¬†—Ā–ľ–Ķ—ą–Ķ–Ĺ–ł—é —ć–ļ—Ā—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ–ĺ–Ļ –ł¬†–ł–Ĺ—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł –Ĺ–į –≤—Ö–ĺ–ī–Ķ –≤¬†–ļ–ĺ—Ä—É –ľ–ĺ–∑–≥–į. –í—č—Ö–ĺ–ī—Ź—Č–ł–Ķ —Ā–ł–≥–Ĺ–į–Ľ—č –ł–∑ –ļ–ĺ—Ä—č –ľ–ĺ–∑–≥–į –ľ–ĺ–≥—É—ā –Ī—č—ā—Ć –Ĺ–į–Ņ—Ä–į–≤–Ľ–Ķ–Ĺ—č –Ĺ–Ķ —ā–Ķ–ľ –į–ī—Ä–Ķ—Ā–į—ā–į–ľ. –Ē–į–Ľ–Ķ–Ķ –ľ—č —Ä–į—Ā—Ā–ľ–ĺ—ā—Ä–ł–ľ –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č–Ķ –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ—č–Ķ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź —ā–į–ļ–ł—Ö –Ľ–ĺ–∂–Ĺ—č—Ö –Ņ–Ķ—Ä–Ķ–į–ī—Ä–Ķ—Ā–į—Ü–ł–Ļ.
–ü—Ä–ĺ—Ā–ĺ–Ĺ–ĺ—á–Ĺ—č–Ķ –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—Ü–ł–ł
–ß–į—Ā—ā–ĺ –Ņ—Ä–ł –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–Ķ –ł–∑ —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł—Ź –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ļ–ĺ —Ā–Ĺ—É –Ĺ–į–Ī–Ľ—é–ī–į—é—ā—Ā—Ź –Ņ—Ä–ĺ—Ā–ĺ–Ĺ–ĺ—á–Ĺ—č–Ķ (–≥–ł–Ņ–Ĺ–į–≥–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ) –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—Ü–ł–ł –≤¬†–≤–ł–ī–Ķ —Ā—É—Č–Ķ—Ā—ā–≤, –Ņ–ĺ–Ľ–∑—É—Č–ł—Ö –Ņ–ĺ —Ā—ā–Ķ–Ĺ–Ķ, –ł–Ľ–ł –ī–≤–ł–∂—É—Č–ł—Ö—Ā—Ź –ł–Ľ–ł –ī–Ķ—Ą–ĺ—Ä–ľ–ł—Ä—É—é—Č–ł—Ö—Ā—Ź –Ņ—Ä–Ķ–ī–ľ–Ķ—ā–ĺ–≤ –ł–∑ –ĺ–ļ—Ä—É–∂–į—é—Č–Ķ–Ļ –ĺ–Ī—Ā—ā–į–Ĺ–ĺ–≤–ļ–ł [25]. –ü—Ä–ĺ—Ā–ĺ–Ĺ–ĺ—á–Ĺ—č–Ķ –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—Ü–ł–ł –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č, –ļ–ĺ–≥–ī–į –Ņ–ĺ–ī –ī–į–≤–Ľ–Ķ–Ĺ–ł–Ķ–ľ —Ā–Ĺ–į —É–∂–Ķ –ĺ—ā–ļ—Ä—č–≤–į–Ķ—ā—Ā—Ź –Ņ—É—ā—Ć –ī–Ľ—Ź –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł–ł –≤¬†–ļ–ĺ—Ä—É, –Ĺ–ĺ –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł –Ķ—Č–Ķ –Ĺ–Ķ –Ī–Ľ–ĺ–ļ–ł—Ä–ĺ–≤–į–Ĺ–ĺ –ł¬†—Ā–ĺ—Ö—Ä–į–Ĺ–Ķ–Ĺ–į —Ā–≤—Ź–∑—Ć –ļ–ĺ—Ä—č —Ā¬†–Ī–Ľ–ĺ–ļ–į–ľ–ł –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł—Ź –ł¬†—Ā–ĺ–∑–Ĺ–į–Ĺ–ł—Ź. –ě—ā–ļ—Ä—č–≤—ą–ł–Ļ—Ā—Ź –Ņ–ĺ—ā–ĺ–ļ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤ –Ņ–ĺ –ł–Ĺ—ā–Ķ–Ĺ—Ā–ł–≤–Ĺ–ĺ—Ā—ā–ł –Ī—É–ī–Ķ—ā —ā–į–ļ–ł–ľ –∂–Ķ, –ļ–į–ļ –ł¬†–Ņ–ĺ—ā–ĺ–ļ –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤. –í—Ā–Ņ–Ľ–Ķ—Ā–ļ–ł –ł–ľ–Ņ—É–Ľ—Ć—Ā–Ĺ–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł –ĺ—ā –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö –≤—Ö–ĺ–ī–ĺ–≤, –Ņ–ĺ–Ņ–į–≤ –≤¬†—Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ, –≤—č–∑–ĺ–≤—É—ā –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ—č–Ķ –ĺ–Ī—Ä–į–∑—č, –Ĺ–į–Ľ–ĺ–∂–Ķ–Ĺ–Ĺ—č–Ķ –Ĺ–į –≤—Ā–Ķ –Ķ—Č–Ķ –≤–ĺ—Ā–Ņ—Ä–ł–Ĺ–ł–ľ–į–Ķ–ľ—É—é —Ä–Ķ–į–Ľ—Ć–Ĺ—É—é –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ—É—é —Ā—Ü–Ķ–Ĺ—É.
–í–∑–ī—Ä–į–≥–ł–≤–į–Ĺ–ł–Ķ –≤–ĺ —Ā–Ĺ–Ķ (—Ā—ā–į—Ä—ā—č —Ā–Ĺ–į)
–ė–Ĺ–ĺ–≥–ī–į –Ņ—Ä–ł –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–Ķ –ĺ—ā –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ļ–ĺ —Ā–Ĺ—É –≤—č—Ö–ĺ–ī –ł–∑ –ļ–ĺ—Ä—č –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į –≤¬†—Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ –Ņ–Ķ—Ä–Ķ–ļ—Ä—č–≤–į–Ķ—ā—Ā—Ź –Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ, –į¬†–≤—č—Ö–ĺ–ī –≤¬†—Ā—ā—Ä—É–ļ—ā—É—Ä—č, —Ā–≤—Ź–∑–į–Ĺ–Ĺ—č–Ķ —Ā¬†–ī–≤–ł–≥–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć—é, –ĺ—Ā—ā–į–Ķ—ā—Ā—Ź –ĺ—ā–ļ—Ä—č—ā—č–ľ. –ü—Ä–ł —Ä–į–∑–≤–ł—ā–ł–ł —Ā–Ĺ–į –į–ļ—ā–ł–≤–į—Ü–ł—Ź –≤¬†–ľ–ĺ—ā–ĺ—Ä–Ĺ—č—Ö –ĺ—ā–ī–Ķ–Ľ–į—Ö –ļ–ĺ—Ä—č, –≤—č–∑–≤–į–Ĺ–Ĺ–į—Ź —É–∂–Ķ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–ľ–ł –≤—Ö–ĺ–ī–į–ľ–ł, –Ĺ–Ķ –Ī–Ľ–ĺ–ļ–ł—Ä—É–Ķ—ā—Ā—Ź, –į¬†–Ņ–Ķ—Ä–Ķ–ī–į–Ķ—ā—Ā—Ź –Ĺ–į –ľ–ĺ—ā–ĺ–Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ—č —Ā–Ņ–ł–Ĺ–Ĺ–ĺ–≥–ĺ –ľ–ĺ–∑–≥–į. –í–ĺ–∑–Ī—É–∂–ī–Ķ–Ĺ–ł–Ķ –ī–≤–ł–≥–į—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ĺ–≤ –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†—Ä–Ķ–∑–ļ–ł–ľ –ī–≤–ł–∂–Ķ–Ĺ–ł—Ź–ľ —ā—É–Ľ–ĺ–≤–ł—Č–į –ł¬†–ļ–ĺ–Ĺ–Ķ—á–Ĺ–ĺ—Ā—ā–Ķ–Ļ, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –Ņ—Ä–Ķ—Ä—č–≤–į—é—ā —Ä–į–∑–≤–ł–≤–į—é—Č–ł–Ļ—Ā—Ź —Ā–ĺ–Ĺ. –≠—ā–ĺ –ľ–ĺ–ī–Ķ–Ľ—Ć —Ä–į—Ā–Ņ—Ä–ĺ—Ā—ā—Ä–į–Ĺ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –ł¬†–ł–∑–≤–Ķ—Ā—ā–Ĺ–ĺ–≥–ĺ –ľ–Ĺ–ĺ–≥–ł–ľ —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł—Ź –≤–∑–ī—Ä–į–≥–ł–≤–į–Ĺ–ł–Ļ –≤–ĺ —Ā–Ĺ–Ķ, —á–į—Ā—ā–ĺ —Ā–ĺ–Ņ—Ä–ĺ–≤–ĺ–∂–ī–į—é—Č–Ķ–≥–ĺ—Ā—Ź –ĺ—Č—É—Č–Ķ–Ĺ–ł–Ķ–ľ –Ņ–į–ī–Ķ–Ĺ–ł—Ź [26].
–°–Ĺ–ĺ–≤–ł–ī–Ķ–Ĺ–ł—Ź
–°–į–ľ–ĺ–Ļ —á–į—Ā—ā–ĺ –≤—Ā—ā—Ä–Ķ—á–į—é—Č–Ķ–Ļ—Ā—Ź –ł¬†–Ī–Ķ–∑–ĺ–Ī–ł–ī–Ĺ–ĺ–Ļ –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł–Ķ–Ļ —Ā–Ĺ–į —Ź–≤–Ľ—Ź—é—ā—Ā—Ź —Ā–Ĺ–ĺ–≤–ł–ī–Ķ–Ĺ–ł—Ź. –ü—Ä–ł—á–ł–Ĺ–į –ł—Ö –≤–ĺ–∑–Ĺ–ł–ļ–Ĺ–ĺ–≤–Ķ–Ĺ–ł—Ź, —Ā¬†–Ĺ–į—ą–Ķ–Ļ —ā–ĺ—á–ļ–ł –∑—Ä–Ķ–Ĺ–ł—Ź, –ĺ—á–Ķ–Ĺ—Ć –Ņ—Ä–ĺ—Ā—ā–į. –Ď–Ľ–ĺ–ļ–ł—Ä—É—é—Č–į—Ź —Ā–ł—Ā—ā–Ķ–ľ–į –Ĺ–į –Ņ—É—ā–ł –ł–∑ –ļ–ĺ—Ä—č –≤¬†—Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ –≤¬†—Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–ł —Ā–Ĺ–į –ł–Ľ–ł –≤¬†–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–Ķ –Ķ–≥–ĺ —Ä–į–∑–≤–ł—ā–ł—Ź –ĺ–ļ–į–∑—č–≤–į–Ķ—ā—Ā—Ź –Ĺ–Ķ –Ņ–ĺ–Ľ–Ĺ–ĺ—Ā—ā—Ć—é –Ņ–Ķ—Ä–Ķ–ļ—Ä—č—ā–ĺ–Ļ. –Ě–Ķ–Ľ—Ć–∑—Ź –ł—Ā–ļ–Ľ—é—á–ł—ā—Ć –ł¬†—ā–ĺ, —á—ā–ĺ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–Ķ —Ā–ł–≥–Ĺ–į–Ľ—č –≤¬†–ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ—č–Ķ –ľ–ĺ–ľ–Ķ–Ĺ—ā—č –ī–ĺ—Ā—ā–ł–≥–į—é—ā –ł–Ĺ—ā–Ķ–Ĺ—Ā–ł–≤–Ĺ–ĺ—Ā—ā–ł, –ļ–ĺ—ā–ĺ—Ä–į—Ź –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź–Ķ—ā –ł–ľ –Ņ—Ä–Ķ–ĺ–ī–ĺ–Ľ–Ķ—ā—Ć –≤–ĺ–∑—Ä–ĺ—Ā—ą–ł–Ķ –Ņ–ĺ—Ä–ĺ–≥–ł –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł—Ź –Ņ–ĺ –ļ–į–Ĺ–į–Ľ–į–ľ –≤¬†—Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ. –í–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–Ķ —Ā–ł—Ā—ā–Ķ–ľ—č –Ĺ–Ķ –ł–ľ–Ķ—é—ā –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł—Ź –≤¬†–Ĺ–į—ą–Ķ–ľ —Ā–ĺ–∑–Ĺ–į–Ĺ–ł–ł, –Ņ–ĺ—ć—ā–ĺ–ľ—É –ī–Ľ—Ź –Ĺ–Ķ–≥–ĺ –ł—Ö —Ā–ł–≥–Ĺ–į–Ľ—č —á–į—Č–Ķ –≤—Ā–Ķ–≥–ĺ —Ź–≤–Ľ—Ź—é—ā—Ā—Ź –Ņ—Ä–ĺ—Ā—ā–ĺ —ą—É–ľ–ĺ–ľ. –ź¬†—ą—É–ľ –ľ–ĺ–∂–Ķ—ā –≤–ĺ–∑–Ī—É–ī–ł—ā—Ć –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ—č —Ā¬†—Ā–į–ľ—č–ľ–ł –Ĺ–ł–∑–ļ–ł–ľ–ł –Ņ–ĺ—Ä–ĺ–≥–į–ľ–ł –ĺ—ā–≤–Ķ—ā–ĺ–≤, —ā–ĺ –Ķ—Ā—ā—Ć –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ—č, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –į–ļ—ā–ł–≤–Ĺ–ĺ —Ä–į–Ī–ĺ—ā–į–Ľ–ł –≤¬†–Ņ—Ä–Ķ–ī—č–ī—É—Č–ł–Ļ –Ņ–Ķ—Ä–ł–ĺ–ī –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź.
–ú–ĺ–∂–Ĺ–ĺ —ā–į–ļ–∂–Ķ –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–ł—ā—Ć, —á—ā–ĺ –≤¬†–ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ—č–Ķ –ľ–ĺ–ľ–Ķ–Ĺ—ā—č –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł –Ņ—Ä–ĺ—Ā—ā—Ä–į–Ĺ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–Ķ —Ä–į—Ā–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–ł–Ķ –≤–ĺ–∑–Ī—É–∂–ī–Ķ–Ĺ–ł—Ź, –ł–ī—É—Č–Ķ–≥–ĺ –ł–∑ –ļ–ĺ—Ä—č –ł¬†–ĺ—ā—Ä–į–∂–į—é—Č–Ķ–≥–ĺ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā –į–Ĺ–į–Ľ–ł–∑–į –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł, –ĺ–ļ–į–∂–Ķ—ā—Ā—Ź –Ī–Ľ–ł–∑–ļ–ł–ľ –ļ¬†–Ņ—Ä–ĺ—Ā—ā—Ä–į–Ĺ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–ľ—É –Ņ—Ä–ĺ—Ą–ł–Ľ—é —Ā—Ą–ĺ—Ä–ľ–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č—Ö –≤¬†–Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–ł –ĺ–Ī—Ä–į–∑–ĺ–≤ –ļ–į–ļ–ł—Ö-–Ľ–ł–Ī–ĺ —Ä–Ķ–į–Ľ—Ć–Ĺ—č—Ö –ĺ–Ī—ä–Ķ–ļ—ā–ĺ–≤. –í¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –≤—Ā–Ņ—č—Ö–Ĺ—É–≤—ą–ł–Ļ –ĺ–Ī—Ä–į–∑ –Ņ–ĺ –į—Ā—Ā–ĺ—Ü–ł–į—ā–ł–≤–Ĺ—č–ľ —Ā–≤—Ź–∑—Ź–ľ –≤¬†—Ā–ł—Ā—ā–Ķ–ľ–Ķ —Ā–ĺ–∑–Ĺ–į–Ĺ–ł—Ź –∑–į–Ņ—É—Ā—ā–ł—ā —Ä–į–∑–≤–ł—ā–ł–Ķ —Ą–į–Ĺ—ā–į—Ā—ā–ł—á–Ķ—Ā–ļ–ł—Ö —Ā—é–∂–Ķ—ā–ĺ–≤.
–°–ļ–ĺ—Ä–Ķ–Ķ –≤—Ā–Ķ–≥–ĺ, —Ā–Ĺ–ĺ–≤–ł–ī–Ķ–Ĺ–ł—Ź –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ—Ź—é—ā —Ā–ĺ–Ī–ĺ–Ļ —Ź–≤–Ľ–Ķ–Ĺ–ł—Ź –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–Ĺ–ĺ–≥–ĺ –Ņ–Ķ—Ä–ł–ĺ–ī–į –ĺ—ā –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ļ–ĺ —Ā–Ĺ—É –ł–Ľ–ł —á–į—Č–Ķ –ĺ—ā —Ā–Ĺ–į –ļ¬†–Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—é, –ļ–ĺ–≥–ī–į –Ī–Ľ–ĺ–ļ–ł—Ä—É—é—Č–į—Ź —Ā–ł—Ā—ā–Ķ–ľ–į –Ĺ–į –≤—Ö–ĺ–ī–Ķ –≤¬†—Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ –Ķ—Č–Ķ –Ĺ–Ķ –Ņ–ĺ–Ľ–Ĺ–ĺ—Ā—ā—Ć—é –∑–į–ļ—Ä—č—ā–į –ł–Ľ–ł —É–∂–Ķ –Ĺ–Ķ–ľ–Ĺ–ĺ–≥–ĺ –Ņ—Ä–ł–ĺ—ā–ļ—Ä—č—ā–į. –≠—ā–į –Ī–Ķ–∑–≤—Ä–Ķ–ī–Ĺ–į—Ź –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł—Ź –ł–ľ–Ķ–Ķ—ā —Ā–≤–ĺ–ł –Ĺ–Ķ–Ņ—Ä–ł—Ź—ā–Ĺ—č–Ķ —Ā—ā–ĺ—Ä–ĺ–Ĺ—č. –ö–ĺ–≥–ī–į –Ī–Ľ–ĺ–ļ –Ĺ–į –≤—Ö–ĺ–ī–Ķ –≤¬†–ĺ—ā–ī–Ķ–Ľ —Ā–ĺ–∑–Ĺ–į–Ĺ–ł—Ź –Ĺ–Ķ —É–ī–į–Ķ—ā—Ā—Ź –Ņ–Ķ—Ä–Ķ–ļ—Ä—č—ā—Ć –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –ī–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł, –≤–ĺ–∑–Ĺ–ł–ļ–į—é—ā –Ĺ–ĺ—á–Ĺ—č–Ķ –ļ–ĺ—ą–ľ–į—Ä—č, –Ņ—Ä–Ķ—Ä—č–≤–į—é—Č–ł–Ķ —ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į. –≠—ā–ĺ –≤—Ā—ā—Ä–Ķ—á–į–Ķ—ā—Ā—Ź –Ņ—Ä–ł –Ĺ–į–Ľ–ł—á–ł–ł –≥–Ľ—É–Ī–ĺ–ļ–ĺ–Ļ –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł–ł.
–ü–į—Ä–į–Ľ–ł—á —Ā–Ĺ–į
–Ě–Ķ—Ä–Ķ–ī–ļ–ĺ –Ņ—Ä–ł –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–Ķ –ĺ—ā —Ā–Ĺ–į –ļ¬†–Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—é –Ņ—Ä–ĺ—Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź —ā–į–ļ –Ĺ–į–∑—č–≤–į–Ķ–ľ—č–Ļ –Ņ–į—Ä–į–Ľ–ł—á —Ā–Ĺ–į. –ß–Ķ–Ľ–ĺ–≤–Ķ–ļ –Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ –Ņ—Ä–ĺ—Ā—č–Ņ–į–Ķ—ā—Ā—Ź, –į–ī–Ķ–ļ–≤–į—ā–Ĺ–ĺ –≤–ĺ—Ā–Ņ—Ä–ł–Ĺ–ł–ľ–į–Ķ—ā –ĺ–ļ—Ä—É–∂–į—é—Č—É—é –ī–Ķ–Ļ—Ā—ā–≤–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā—Ć –ł¬†—Ā–ĺ–Ī—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–Ķ —ā–Ķ–Ľ–ĺ, –Ĺ–ĺ –Ĺ–Ķ —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ķ–Ĺ –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä–ĺ–≥–ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł (–ĺ—ā –Ĺ–Ķ—Ā–ļ–ĺ–Ľ—Ć–ļ–ł—Ö –ī–Ķ—Ā—Ź—ā–ļ–ĺ–≤ —Ā–Ķ–ļ—É–Ĺ–ī –ī–ĺ –ľ–ł–Ĺ—É—ā—č) —Ā–ĺ–≤–Ķ—Ä—ą–ł—ā—Ć –ļ–į–ļ–ĺ–Ķ-–Ľ–ł–Ī–ĺ –Ņ—Ä–ĺ–ł–∑–≤–ĺ–Ľ—Ć–Ĺ–ĺ–Ķ –ī–≤–ł–∂–Ķ–Ĺ–ł–Ķ. –°–ĺ–≥–Ľ–į—Ā–Ĺ–ĺ –Ĺ–į—ą–Ķ–Ļ —Ā—Ö–Ķ–ľ–Ķ –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ–Ļ –Ņ—Ä–ł—á–ł–Ĺ–ĺ–Ļ —ć—ā–ĺ–≥–ĺ —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –∑–į–Ņ–į–∑–ī—č–≤–į–Ĺ–ł–Ķ —Ā–Ĺ—Ź—ā–ł—Ź –Ī–Ľ–ĺ–ļ–į –Ņ–Ķ—Ä–Ķ–ī–į—á–ł –ľ–ĺ—ā–ĺ—Ä–Ĺ—č—Ö –ļ–ĺ–ľ–į–Ĺ–ī –ł–∑ –ļ–ĺ—Ä—č –≤¬†–ĺ—ā–ī–Ķ–Ľ –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł—Ź –ł¬†–ī–≤–ł–≥–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł, –ļ–ĺ–≥–ī–į —Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ —É–∂–Ķ –Ņ—Ä–ĺ—Ā–Ĺ—É–Ľ–ĺ—Ā—Ć, –į¬†—Ā–ł—Ā—ā–Ķ–ľ–į —É–Ņ—Ä–į–≤–Ľ–Ķ–Ĺ–ł—Ź –ī–≤–ł–∂–Ķ–Ĺ–ł—Ź–ľ–ł –Ķ—Č–Ķ –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–į–Ķ—ā —Ā–Ņ–į—ā—Ć.
–°–ĺ–ľ–Ĺ–į–ľ–Ī—É–Ľ–ł–∑–ľ (—Ā–Ĺ–ĺ—Ö–ĺ–∂–ī–Ķ–Ĺ–ł–Ķ)
–í—Ā—ā—Ä–Ķ—á–į—é—ā—Ā—Ź –ł¬†–Ņ—Ä—Ź–ľ–ĺ –Ņ—Ä–ĺ—ā–ł–≤–ĺ–Ņ–ĺ–Ľ–ĺ–∂–Ĺ—č–Ķ —Ā–Ľ—É—á–į–ł. –ě—ā–ļ—Ä—č–≤–į–Ķ—ā—Ā—Ź –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤ –ł–∑ –ļ–ĺ—Ä—č –≤¬†–ĺ—ā–ī–Ķ–Ľ –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł—Ź –ł¬†–ī–≤–ł–≥–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł, –į¬†–Ņ—É—ā—Ć –≤¬†–ĺ—ā–ī–Ķ–Ľ —Ā–ĺ–∑–Ĺ–į–Ĺ–ł—Ź –≤—Ā–Ķ –Ķ—Č–Ķ –∑–į–ļ—Ä—č—ā (—Ä–ł—Ā. 2 –ł¬†3). –≠—ā–ĺ –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†—Ā–Ĺ–ĺ—Ö–ĺ–∂–ī–Ķ–Ĺ–ł—Ź–ľ¬†‚Äď –ī–ĺ–≤–ĺ–Ľ—Ć–Ĺ–ĺ —á–į—Ā—ā–ĺ –≤—Ā—ā—Ä–Ķ—á–į—é—Č–Ķ–ľ—É—Ā—Ź —Ź–≤–Ľ–Ķ–Ĺ–ł—é, –ĺ—Ā–ĺ–Ī–Ķ–Ĺ–Ĺ–ĺ –≤¬†–ī–Ķ—ā—Ā–ļ–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ. –õ—é–ī–ł –≤—Ā—ā–į—é—ā –Ĺ–ĺ—á—Ć—é —Ā¬†–Ņ–ĺ—Ā—ā–Ķ–Ľ–ł –ł¬†—Ā–ĺ–≤–Ķ—Ä—ą–į—é—ā —Ä–į–∑–Ĺ–ĺ–ĺ–Ī—Ä–į–∑–Ĺ—č–Ķ –ī–Ķ–Ļ—Ā—ā–≤–ł—Ź –ł–Ľ–ł –Ņ—Ä–ĺ–≥—É–Ľ–ļ–ł, –ī–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā—Ć –ļ–ĺ—ā–ĺ—Ä—č—Ö –ľ–ĺ–∂–Ķ—ā –Ī—č—ā—Ć —Ā–į–ľ–ĺ–Ļ —Ä–į–∑–Ĺ–ĺ–Ļ. –ď–Ľ–į–∑–į –Ņ—Ä–ł —Ö–ĺ–∂–ī–Ķ–Ĺ–ł–ł –≤–ĺ —Ā–Ĺ–Ķ –ĺ–Ī—č—á–Ĺ–ĺ –ĺ—ā–ļ—Ä—č—ā—č. –õ—é–ī–ł –Ĺ–Ķ –Ĺ–į—ā—č–ļ–į—é—ā—Ā—Ź –Ĺ–į –Ņ—Ä–Ķ–Ņ—Ź—ā—Ā—ā–≤–ł—Ź, –ł—Ö –ī–≤–ł–∂–Ķ–Ĺ–ł—Ź —Ö–ĺ—Ä–ĺ—ą–ĺ –ļ–ĺ–ĺ—Ä–ī–ł–Ĺ–ł—Ä–ĺ–≤–į–Ĺ—č. –ß–į—Ā—ā–ĺ –≤¬†–ļ–ĺ–Ĺ—Ü–Ķ —ā–į–ļ–ł—Ö –Ņ—Ä–ĺ–≥—É–Ľ–ĺ–ļ –ĺ–Ĺ–ł –ľ–ĺ–≥—É—ā –Ĺ–į–Ļ—ā–ł –ī—Ä—É–≥–ĺ–Ķ –ľ–Ķ—Ā—ā–ĺ –ī–Ľ—Ź —Ā–Ĺ–į, –į¬†–Ņ–ĺ—Ā–Ľ–Ķ –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł—Ź –Ĺ–Ķ –Ņ–ĺ–Ĺ–ł–ľ–į—é—ā, –ļ–į–ļ —Ā—é–ī–į –Ņ–ĺ–Ņ–į–Ľ–ł [27]. –Į–≤–Ľ–Ķ–Ĺ–ł–Ķ —Ā–ĺ–ľ–Ĺ–į–ľ–Ī—É–Ľ–ł–∑–ľ–į —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –Ķ—Č–Ķ –ĺ–ī–Ĺ–ł–ľ –į—Ä–≥—É–ľ–Ķ–Ĺ—ā–ĺ–ľ –≤¬†–Ņ–ĺ–Ľ—Ć–∑—É —ā–ĺ–≥–ĺ, —á—ā–ĺ —Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ –ĺ—ā–ī–Ķ–Ľ–Ķ–Ĺ–ĺ –ĺ—ā –ļ–ĺ—Ä—č –ľ–ĺ–∑–≥–į –ł¬†—Ā—ā—Ä—É–ļ—ā—É—Ä, –Ņ—Ä–ĺ–≥—Ä–į–ľ–ľ–ł—Ä—É—é—Č–ł—Ö –ī–≤–ł–∂–Ķ–Ĺ–ł—Ź —ā–Ķ–Ľ–į.
–í–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–Ķ –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—Ü–ł–ł
–í —Ä–į–∑–ī–Ķ–Ľ–Ķ –嬆–Ņ—Ä–ĺ—Ā–ĺ–Ĺ–ĺ—á–Ĺ—č—Ö –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—Ü–ł—Ź—Ö –ľ—č –≥–ĺ–≤–ĺ—Ä–ł–Ľ–ł –ĺ–Ī –ĺ—Ā–ĺ–Ī–Ķ–Ĺ–Ĺ–ĺ—Ā—ā—Ź—Ö –≤–ĺ—Ā–Ņ—Ä–ł—Ź—ā–ł—Ź, –≤—č–∑–≤–į–Ĺ–Ĺ—č—Ö —Ā–ľ–Ķ—ą–Ķ–Ĺ–ł–Ķ–ľ —ć–ļ—Ā—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ–ĺ–Ļ –ł¬†–ł–Ĺ—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł –Ĺ–į –≤—Ö–ĺ–ī–Ķ –≤¬†–ļ–ĺ—Ä—É, –ļ–ĺ—ā–ĺ—Ä–ĺ–Ķ –ĺ–ļ–į–∑—č–≤–į–Ķ—ā—Ā—Ź –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č–ľ –≤¬†–Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–Ĺ—č–Ķ –Ņ–Ķ—Ä–ł–ĺ–ī—č –ĺ—ā –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ļ–ĺ —Ā–Ĺ—É, –ļ–ĺ–≥–ī–į –ĺ–ī–Ĺ–ł –Ņ—Ä–ĺ–≤–ĺ–ī—Ź—Č–ł–Ķ –Ņ—É—ā–ł –Ķ—Č–Ķ –Ĺ–Ķ –Ņ–ĺ–Ľ–Ĺ–ĺ—Ā—ā—Ć—é –∑–į–ļ—Ä—č–Ľ–ł—Ā—Ć, –į¬†–ī—Ä—É–≥–ł–Ķ –Ķ—Č–Ķ –Ĺ–Ķ –Ņ–ĺ–Ľ–Ĺ–ĺ—Ā—ā—Ć—é –ĺ—ā–ļ—Ä—č–Ľ–ł—Ā—Ć. –í¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –ł–Ĺ—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ—č–Ķ —Ā–ł–≥–Ĺ–į–Ľ—č –Ņ—Ä–ĺ–Ķ—Ü–ł—Ä—É—é—ā—Ā—Ź –Ĺ–į —Ä–Ķ–į–Ľ—Ć–Ĺ—É—é –∂–ł–∑–Ĺ–Ķ–Ĺ–Ĺ—É—é —Ā–ł—ā—É–į—Ü–ł—é –ł¬†–≤–ĺ—Ā–Ņ—Ä–ł–Ĺ–ł–ľ–į—é—ā—Ā—Ź –Ľ—é–ī—Ć–ľ–ł –ļ–į–ļ –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—Ü–ł–ł.
–ü–ĺ—Ö–ĺ–∂–Ķ–Ķ —Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ —ā–Ķ–ĺ—Ä–Ķ—ā–ł—á–Ķ—Ā–ļ–ł –ľ–ĺ–∂–Ķ—ā —Ä–į–∑–≤–ł–≤–į—ā—Ć—Ā—Ź –ł¬†–≤¬†–Ņ—Ä–ĺ—ā–ł–≤–ĺ–Ņ–ĺ–Ľ–ĺ–∂–Ĺ–ĺ–ľ –Ĺ–į–Ņ—Ä–į–≤–Ľ–Ķ–Ĺ–ł–ł. –ü–ĺ –į–Ĺ–į–Ľ–ĺ–≥–ł–ł –ľ—č –Ĺ–į–∑–≤–į–Ľ–ł –Ķ–≥–ĺ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–ľ–ł –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—Ü–ł—Ź–ľ–ł. –ú–ĺ–∂–Ĺ–ĺ –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–ł—ā—Ć, —á—ā–ĺ –Ņ–ĺ–ī –ī–į–≤–Ľ–Ķ–Ĺ–ł–Ķ–ľ —Ā–Ĺ–į –Ĺ–į—á–Ĺ—É—ā –ĺ—ā–ļ—Ä—č–≤–į—ā—Ć—Ā—Ź –Ņ—É—ā–ł, —Ā–≤—Ź–∑—č–≤–į—é—Č–ł–Ķ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–Ķ —Ā–ł—Ā—ā–Ķ–ľ—č —Ā¬†–ļ–ĺ—Ä–ĺ–Ļ –ľ–ĺ–∑–≥–į (—Ä–ł—Ā. 2 –ł¬†3), –Ĺ–ĺ –∂–ł–∑–Ĺ–Ķ–Ĺ–Ĺ—č–Ķ –ĺ–Ī—Ā—ā–ĺ—Ź—ā–Ķ–Ľ—Ć—Ā—ā–≤–į –Ĺ–Ķ –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź—é—ā –Ĺ–Ķ–ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ –Ņ–Ķ—Ä–Ķ–Ļ—ā–ł –ļ–ĺ —Ā–Ĺ—É. –°–≤—Ź–∑—Ć –ļ–ĺ—Ä—č —Ā¬†—ć–ļ—Ā—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ—č–ľ–ł –≤—Ö–ĺ–ī–į–ľ–ł –ł¬†–≤—č—Ö–ĺ–ī—č –Ĺ–į —Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ –ł¬†–ī–≤–ł–≥–į—ā–Ķ–Ľ—Ć–Ĺ—É—é –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć –ĺ—Ā—ā–į—é—ā—Ā—Ź –ĺ—ā–ļ—Ä—č—ā—č–ľ–ł. –ü—Ä–ł —ć—ā–ĺ–ľ –ľ–ĺ–∂–Ķ—ā –ĺ–ļ–į–∑–į—ā—Ć—Ā—Ź, —á—ā–ĺ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –į–Ĺ–į–Ľ–ł–∑–į —ć–ļ—Ā—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ—č—Ö —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤ –Ī—É–ī—É—ā –Ņ–ĺ–Ņ–į–ī–į—ā—Ć –≤¬†–Ī–Ľ–ĺ–ļ –į—Ā—Ā–ĺ—Ü–ł–į—ā–ł–≤–Ĺ–ĺ–Ļ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł–Ĺ—ā–Ķ–≥—Ä–į—Ü–ł–ł –ł¬†—Ä–į—Ā—Ü–Ķ–Ĺ–ł–≤–į—ā—Ć—Ā—Ź —ā–į–ľ –ļ–į–ļ —Ā–ł–≥–Ĺ–į–Ľ—č –ļ–ĺ—Ä—č –į–Ĺ–į–Ľ–ł–∑–į –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö –≤—Ö–ĺ–ī–ĺ–≤. –Ę–Ķ–Ņ–Ķ—Ä—Ć —É–Ņ—Ä–į–≤–Ľ—Ź—ā—Ć –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–ľ–ł —Ā–ł—Ā—ā–Ķ–ľ–į–ľ–ł –Ī—É–ī—É—ā –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—ā–ĺ—Ä–Ĺ—č–Ķ —Ā–ł–≥–Ĺ–į–Ľ—č. –ü–ĺ–Ĺ—Ź—ā–Ĺ–ĺ, —á—ā–ĺ –Ĺ–ł –ļ¬†—á–Ķ–ľ—É —Ö–ĺ—Ä–ĺ—ą–Ķ–ľ—É –≤¬†–ĺ—ā–Ĺ–ĺ—ą–Ķ–Ĺ–ł–ł –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č —ć—ā–ĺ –Ĺ–Ķ –Ņ—Ä–ł–≤–Ķ–ī–Ķ—ā. –ė–∑–≤–Ķ—Ā—ā–Ĺ—č–Ķ –Ĺ–Ķ–≥–į—ā–ł–≤–Ĺ—č–Ķ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–Ķ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź –ĺ—Ā—ā—Ä–ĺ–≥–ĺ —Ā—ā—Ä–Ķ—Ā—Ā–į, –≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ, –ł–ľ–Ķ—é—ā —ā–į–ļ–ĺ–Ļ –∂–Ķ –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ —Ä–į–∑–≤–ł—ā–ł—Ź.
–Ē–į–Ĺ–Ĺ—č–ľ –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–ĺ–ľ –ľ–ĺ–∂–Ĺ–ĺ –ĺ–Ī—ä—Ź—Ā–Ĺ–ł—ā—Ć –ł¬†–Ņ–ĺ—Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö –ļ–ĺ–ľ–Ņ–ĺ–Ĺ–Ķ–Ĺ—ā–ĺ–≤ (—ā–ĺ—ą–Ĺ–ĺ—ā—č, —Ä–≤–ĺ—ā—č), –Ī–ĺ–Ľ–Ķ–∑–Ĺ–ł –ī–≤–ł–∂–Ķ–Ĺ–ł—Ź –ł–Ľ–ł —É–ļ–į—á–ł–≤–į–Ĺ–ł—Ź. –†–ł—ā–ľ–ł—á–Ĺ–ĺ–Ķ –Ņ–ĺ–ļ–į—á–ł–≤–į–Ĺ–ł–Ķ, –ł–Ľ–ł –Ņ–Ķ—Ä–ł–ĺ–ī–ł—á–Ķ—Ā–ļ–ĺ–Ķ, –ļ–į–ļ –Ĺ–į –ļ–į—á–Ķ–Ľ—Ź—Ö, –ł–Ľ–ł –Ņ–ĺ—Ā—ā–ĺ—Ź–Ĺ–Ĺ–ĺ–Ķ, –ļ–į–ļ –≤¬†–ļ–ĺ—Ā–ľ–ł—á–Ķ—Ā–ļ–ĺ–ľ –Ņ–ĺ–Ľ–Ķ—ā–Ķ, –į¬†—ā–į–ļ–∂–Ķ —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł–Ķ –≥—Ä–į–≤–ł—ā–į—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–Ļ –Ĺ–į–≥—Ä—É–∑–ļ–ł —Ä–Ķ–∑–ļ–ĺ –ľ–Ķ–Ĺ—Ź—é—ā –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–Ĺ—É—é –ł–ľ–Ņ—É–Ľ—Ć—Ā–į—Ü–ł—é –ĺ—ā –ľ–Ķ—Ö–į–Ĺ–ĺ—Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–ĺ–≤ –≤¬†—Ā—ā–Ķ–Ĺ–ļ–į—Ö –ĺ—Ä–≥–į–Ĺ–ĺ–≤ –∂–Ķ–Ľ—É–ī–ĺ—á–Ĺ–ĺ-–ļ–ł—ą–Ķ—á–Ĺ–ĺ–≥–ĺ —ā—Ä–į–ļ—ā–į. –Ě–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ–ĺ—Ā—ā—Ć —ć–ļ—Ā—ā—Ä–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –į–Ĺ–į–Ľ–ł–∑–į –Ņ—Ä–ł—á–ł–Ĺ —ā–į–ļ–ĺ–≥–ĺ –Ĺ–Ķ–ĺ–Ī—č—á–Ĺ–ĺ–≥–ĺ –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł—Ź –į—Ą—Ą–Ķ—Ä–Ķ–Ĺ—ā–į—Ü–ł–ł –Ņ—Ä–ł–≤–Ķ–ī–Ķ—ā –ļ¬†—É—Ā–ł–Ľ–Ķ–Ĺ–ł—é –ī–į–≤–Ľ–Ķ–Ĺ–ł—Ź —Ā–Ĺ–į. –ü–ĺ–ī —ć—ā–ł–ľ –ī–į–≤–Ľ–Ķ–Ĺ–ł–Ķ–ľ –Ī—É–ī–Ķ—ā –Ņ—Ä–ł–ĺ—ā–ļ—Ä—č—ā –ļ–ĺ—Ä–ļ–ĺ–≤—č–Ļ –≤—č—Ö–ĺ–ī –≤¬†—Ā—ā—Ä—É–ļ—ā—É—Ä—č –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –į–Ĺ–į–Ľ–ł–∑–į –ł,¬†–≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ, –≤—Ö–ĺ–ī –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł –≤¬†–ļ–ĺ—Ä—É. –Ě–ĺ –≤—č—Ā–ĺ–ļ–į—Ź –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ—á–Ķ—Ā–ļ–į—Ź –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć –≤¬†—ć—ā–ĺ –≤—Ä–Ķ–ľ—Ź –ł—Ā–ļ–Ľ—é—á–į–Ķ—ā –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī –ļ–ĺ —Ā–Ĺ—É, –ł¬†–ļ–ĺ—Ä–į –ľ–ĺ–∑–≥–į –ĺ—Ā—ā–į–Ķ—ā—Ā—Ź —Ā–≤—Ź–∑–į–Ĺ–Ĺ–ĺ–Ļ —Ā¬†–ľ–ĺ—Č–Ĺ—č–ľ –Ņ–ĺ—ā–ĺ–ļ–ĺ–ľ —ć–ļ—Ā—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ—č—Ö –≤—Ö–ĺ–ī–ĺ–≤. –≠—ā–ĺ—ā –Ņ–ĺ—ā–ĺ–ļ —ć–ļ—Ā—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ—č—Ö —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤ –Ĺ–į—á–ł–Ĺ–į–Ķ—ā –∑–į–Ī—Ä–į—Ā—č–≤–į—ā—Ć—Ā—Ź –≤¬†–ĺ—ā–ī–Ķ–Ľ—č –į–Ĺ–į–Ľ–ł–∑–į –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł, —á—ā–ĺ –ī–Ķ–Ľ–į–Ķ—ā –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č–Ļ –Ņ–ĺ—ā–ĺ–ļ –Ĺ–Ķ–Ņ—Ä–ł–≤—č—á–Ĺ—č–ľ. –°–ł—Ā—ā–Ķ–ľ–į —Ä–į—Ā—Ü–Ķ–Ĺ–ł–≤–į–Ķ—ā –Ķ–≥–ĺ –ļ–į–ļ –Ņ–ĺ—Ā—ā—É–Ņ–į—é—Č–ł–Ļ –ł–∑ –ĺ—Ä–≥–į–Ĺ–ĺ–≤ –∂–Ķ–Ľ—É–ī–ĺ—á–Ĺ–ĺ-–ļ–ł—ą–Ķ—á–Ĺ–ĺ–≥–ĺ —ā—Ä–į–ļ—ā–į. –ē—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–Ļ —Ä–Ķ–į–ļ—Ü–ł–Ķ–Ļ –Ĺ–į —ć—ā–ł —Ā–ł–≥–Ĺ–į–Ľ—č –Ī—É–ī–Ķ—ā –ļ–ĺ–ľ–į–Ĺ–ī–į —ć–ļ—Ā—ā—Ä–Ķ–Ĺ–Ĺ–ĺ–Ļ –ĺ—á–ł—Ā—ā–ļ–ł –∂–Ķ–Ľ—É–ī–ļ–į –ĺ—ā –ł—Ā—ā–ĺ—á–Ĺ–ł–ļ–ĺ–≤ –Ņ—É–≥–į—é—Č–ł—Ö —Ā–ł–≥–Ĺ–į–Ľ–ĺ–≤, —ā–ĺ –Ķ—Ā—ā—Ć —Ä–≤–ĺ—ā–Ĺ—č–Ļ —Ä–Ķ—Ą–Ľ–Ķ–ļ—Ā.
–Ě–į—Ā–ļ–ĺ–Ľ—Ć–ļ–ĺ –Ĺ–į–ľ –ł–∑–≤–Ķ—Ā—ā–Ĺ–ĺ, –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ, –≤–Ķ–ī—É—Č–ł–Ļ –ļ¬†–≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č–ľ –≥–į–Ľ–Ľ—é—Ü–ł–Ĺ–į—Ü–ł—Ź–ľ, —Ä–į–Ĺ–Ķ–Ķ –Ĺ–ł–ļ–Ķ–ľ –Ĺ–Ķ —Ä–į—Ā—Ā–ľ–į—ā—Ä–ł–≤–į–Ľ—Ā—Ź. –í–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ, —á—ā–ĺ –≤¬†—Ā–ĺ–Ĺ–Ĺ–ĺ-–ī–Ķ–Ņ—Ä–ł–≤–ł—Ä–ĺ–≤–į–Ĺ–Ĺ–ĺ–ľ –ĺ–Ī—Č–Ķ—Ā—ā–≤–Ķ –Ĺ–Ķ–≥–į—ā–ł–≤–Ĺ—č–Ķ –ī–Ľ—Ź –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ā–ł—Ā—ā–Ķ–ľ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź —Ä–į–Ī–ĺ—ā—č –Ņ–ĺ–ī–ĺ–Ī–Ĺ–ĺ–≥–ĺ –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–į –ľ–ĺ–≥—É—ā –Ī—č—ā—Ć —Ā—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ—č–ľ–ł. –°–ļ–ĺ—Ä–Ķ–Ķ –≤—Ā–Ķ–≥–ĺ, –ł–ľ–Ķ–Ĺ–Ĺ–ĺ –ł–Ĺ—ā–Ķ—Ä—Ą–Ķ—Ä–Ķ–Ĺ—Ü–ł—Ź —ć–ļ—Ā—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ–ĺ–Ļ –ł¬†–ł–Ĺ—ā–Ķ—Ä–ĺ—Ü–Ķ–Ņ—ā–ł–≤–Ĺ–ĺ–Ļ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł –Ĺ–į –Ņ—É—ā–ł –≤¬†–ļ–ĺ—Ä—É –ł¬†–Ĺ–į –≤—č—Ö–ĺ–ī–į—Ö –ł–∑ –ļ–ĺ—Ä—č –ľ–ĺ–∂–Ķ—ā –Ī—č—ā—Ć –Ņ—Ä–ł—á–ł–Ĺ–ĺ–Ļ –ľ–Ĺ–ĺ–≥–ł—Ö –Ņ—Ā–ł—Ö–ĺ—Ā–ĺ–ľ–į—ā–ł—á–Ķ—Ā–ļ–ł—Ö –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ.
–Ē–Ķ—ā–į–Ľ—Ć–Ĺ–ĺ–Ķ –≤—č—Ź—Ā–Ĺ–Ķ–Ĺ–ł–Ķ –ī–ł–Ĺ–į–ľ–ł–ļ–ł —É—á–į—Ā—ā–ł—Ź –≤—č—Ā—ą–ł—Ö –ĺ—ā–ī–Ķ–Ľ–ĺ–≤ –ľ–ĺ–∑–≥–į –≤¬†—Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö —Ą—É–Ĺ–ļ—Ü–ł–Ļ –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī —Ā–Ĺ–į¬†‚Äď –∑–į–ī–į—á–į –Ĺ–į –Ī—É–ī—É—Č–Ķ–Ķ. –ü–ĺ–ļ–į –∂–Ķ —Ā–ľ–Ķ–Ľ–ĺ –ľ–ĺ–∂–Ĺ–ĺ —É—ā–≤–Ķ—Ä–∂–ī–į—ā—Ć, —á—ā–ĺ —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ—č–Ļ –ł¬†–ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ—č–Ļ —Ā–ĺ–Ŭ†‚Äď —ć—ā–ĺ —Ā–į–ľ—č–Ļ –ī–Ķ—ą–Ķ–≤—č–Ļ, –Ī–Ķ–∑–ĺ–Ņ–į—Ā–Ĺ—č–Ļ –ł¬†–Ņ—Ä–ł—Ź—ā–Ĺ—č–Ļ —Ā–Ņ–ĺ—Ā–ĺ–Ī —É–Ľ—É—á—ą–Ķ–Ĺ–ł—Ź –ļ–į–ļ –Ņ—Ā–ł—Ö–ł—á–Ķ—Ā–ļ–ĺ–≥–ĺ, —ā–į–ļ –ł¬†—Ą–ł–∑–ł—á–Ķ—Ā–ļ–ĺ–≥–ĺ –∑–ī–ĺ—Ä–ĺ–≤—Ć—Ź.
1. –Ē–ĺ—Ä–ĺ—Ö–ĺ–≤ –í.–Ď. –°–ĺ–ľ–Ĺ–ĺ–Ľ–ĺ–≥–ł—Ź –ł –Ī–Ķ–∑–ĺ–Ņ–į—Ā–Ĺ–ĺ—Ā—ā—Ć –Ņ—Ä–ĺ—Ą–Ķ—Ā—Ā–ł–ĺ- –Ĺ–į–Ľ—Ć–Ĺ–ĺ–Ļ –ī–Ķ—Ź—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł // –Ė—É—Ä–Ĺ–į–Ľ –≤—č—Ā—ą–Ķ–Ļ –Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ –ī–Ķ- —Ź—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł –ł–ľ. –ė.–ü. –ü–į–≤–Ľ–ĺ–≤–į. 2013. –Ę. 63. ‚ĄĖ 1. –°. 33‚Äď47.
2. –ö–ĺ–≤–į–Ľ—Ć–∑–ĺ–Ĺ –í.–ú. –ě—Ā–Ĺ–ĺ–≤—č —Ā–ĺ–ľ–Ĺ–ĺ–Ľ–ĺ–≥–ł–ł: —Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—Ź –ł –Ĺ–Ķ–Ļ—Ä–ĺ—Ö–ł–ľ–ł—Ź —Ü–ł–ļ–Ľ–į ¬ę–Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ ‚Äď —Ā–ĺ–ŬĽ. –ú.: –Ď–ł–Ĺ–ĺ–ľ. –õ–į–Ī–ĺ—Ä–į—ā–ĺ—Ä–ł—Ź –∑–Ĺ–į–Ĺ–ł–Ļ, 2012.
3. Akerstedt T., Hallvig D., Anund A. et al. Having to stop driving at night because of dangerous sleepiness ‚Äď awareness, physiology and behaviour // J. Sleep Res. 2013. Vol. 22. ‚ĄĖ 4. P. 380‚Äď388.
4. Maquet P. The role of sleep in learning and memory // Science. 2001. Vol. 294. ‚ĄĖ 5544. P. 1048‚Äď1052.
5. Diekelmann S., Born J. The memory function of sleep // Nat. Rev. Neurosci. 2010. Vol. 11. ‚ĄĖ 2. P. 114‚Äď126.
6. Harrison Y., Horne J.A. The impact of sleep deprivation on decision making: a review // J. Exp. Psychol. Appl. 2000. Vol. 6. ‚ĄĖ 3. P. 236‚Äď249.
7. Chuah L.Y., Chee M.W. Cholinergic augmentation modulates visual task performance in sleep-deprived young adults // J. Neurosci. 2008. Vol. 28. ‚ĄĖ 44. P. 11369‚Äď11377.
8. Genzel L., Dresler M., Wehrle R. et al. Slow wave sleep and REM sleep awakenings do not affect sleep dependent memory consolidation // Sleep. 2009. Vol. 32. ‚ĄĖ 3. P. 302‚Äď310.
9. Krueger J.M., Ob√°l F. A neuronal group theory of sleep function // J. Sleep Res. 1993. Vol. 2. ‚ĄĖ 2. P. 63‚Äď69.
10. Tononi G., Cirelli C. Sleep function and synaptic homeostasis // Sleep Med. Rev. 2006. Vol. 10. ‚ĄĖ 1. P. 49‚Äď62.
11. Everson C.A., Bergmann B.M., Rechtschaffen A. Sleep deprivation in the rat: III. Total sleep deprivation // Sleep. 1989. Vol. 12. ‚ĄĖ 1. P. 13‚Äď21.
12. Cirelli C., Shaw P.J., Rechtschaffen A. et al. No evidence of brain cell degeneration after long-term sleep deprivation in rats // Brain Res. 1999. Vol. 840. ‚ĄĖ 1‚Äď2. P. 184‚Äď193.
13. Rechtshaffen A., Bergmann B.M. Sleep deprivation in the rat: an update of the 1989 paper // Sleep. 2002. Vol. 25. ‚ĄĖ 1. P. 18‚Äď24.
14. –ü–ł–≥–į—Ä–Ķ–≤ –ė.–Ě. –Ě–ĺ–≤–į—Ź –∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–į—Ź –∑–ĺ–Ĺ–į –Ĺ–į –Ĺ–ł–∂- –Ĺ–Ķ–Ļ —Ā—ā–Ķ–Ĺ–ļ–Ķ –ļ—Ä–Ķ—Ā—ā–ĺ–≤–ł–ī–Ĺ–ĺ–Ļ –Ī–ĺ—Ä–ĺ–∑–ī—č –ľ–ĺ–∑–≥–į –ļ–ĺ—ą–Ķ–ļ // –Ě–Ķ–Ļ—Ä–ĺ—Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—Ź. 1984. –Ę. 16. ‚ĄĖ 6. –°. 766‚Äď773.
15. Pigarev I.N., Nothdurft H.C., Kastner S. Evidence for asynchronous development of sleep in cortical areas // Neuroreport. 1997. Vol. 8. ‚ĄĖ 11. P. 2557‚Äď2560.
16. Werth E., Achermann P., Borbely A.A. Brain topography of the human sleep EEG: antero-posterior shifts of spectral power // Neuroreport. 1996. Vol. 8. ‚ĄĖ 1. P. 123‚Äď127.
17. Ferrara M., De Gennaro L. Going local: insights from EEG and stereo-EEG studies of the human sleep-wake cycle // Curr. Top. Med. Chem. 2011. Vol. 11. ‚ĄĖ 19. P. 2423‚Äď2437.
18. Vyazovskiy V.V., Olcese U., Hanlon E.C. et al. Local sleep in awake rats // Nature. 2011. Vol. 472. ‚ĄĖ 6. P. 443‚Äď447.
19. –ü–ł–≥–į—Ä–Ķ–≤ –ė.–Ě. –í–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć–Ĺ–į—Ź —ā–Ķ–ĺ—Ä–ł—Ź —Ā–Ĺ–į // –Ė—É—Ä–Ĺ–į–Ľ –≤—č- —Ā—ą–Ķ–Ļ –Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ –ī–Ķ—Ź—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł –ł–ľ. –ė.–ü. –ü–į–≤–Ľ–ĺ–≤–į. 2013. –Ę. 63. ‚ĄĖ 1. –°. 86‚Äď104.
20. –ü–ł–≥–į—Ä–Ķ–≤ –ė.–Ě., –ü–ł–≥–į—Ä–Ķ–≤–į –ú.–õ. –°–ĺ–Ĺ, —ć–ľ–ĺ—Ü–ł–ł –ł –≤–ł—Ā—Ü–Ķ—Ä–į–Ľ—Ć- –Ĺ—č–Ļ –ļ–ĺ–Ĺ—ā—Ä–ĺ–Ľ—Ć // –§–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—Ź —á–Ķ–Ľ–ĺ–≤–Ķ–ļ–į. 2013. –Ę. 39. ‚ĄĖ 6. –°. 31‚Äď44.
21. Nedergaard M. Neuroscience. Garbage truck of the brain // Science. 2013. Vol. 340. ‚ĄĖ 6140. P. 1529‚Äď1530.
22. Xie L., Kang H., Xu Q. et al. Sleep drives metabolite clearance from the adult brain // Science. 2013. Vol. 342. ‚ĄĖ 6156. P. 373‚Äď377.
23. –Ě–ĺ–∑–ī—Ä–į—á–Ķ–≤ –ź.–Ē. –§–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—Ź –≤–Ķ–≥–Ķ—ā–į—ā–ł–≤–Ĺ–ĺ–Ļ –Ĺ–Ķ—Ä–≤–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č. –õ.: –ú–Ķ–ī–ł—Ü–ł–Ĺ–į, 1983.
24. Chase M.H. Confirmation of the consensus that glycinergic postsynaptic inhibition is responsible for the atonia of REM sleep // Sleep. 2008. Vol. 31. ‚ĄĖ 11. P. 1487‚Äď1491.
25. Sturzenegger C., Bassetti C.L. The clinical spectrum of narcolepsy with cataplexy: a reappraisal // J. Sleep Res. 2004. Vol. 13. ‚ĄĖ 4. P. 395‚Äď406.
26. Walters A.S. Clinical identification of the simple sleep-related movement disorders // Chest. 2007. Vol. 131. ‚ĄĖ 4. P. 1260‚Äď1266.
27. Kavey N.B., Whyte J., Resor S.R. Jr. et al. Somnambulism in adults // Neurology. 1990. Vol. 40. ‚ĄĖ 5. P. 749‚Äď752.
Asynchronous development of sleep – a plausible cause of cognitive impairment and a number of pathological conditions related to sleep-wake cycle
I.N. Pigarev1, M.L. Pigareva2
1 A.A. Kharkevich Institute of Information Transfer Issues of the Russian Academy of Sciences
2 Institute of Higher Nervous Function and Neurophysiology of the Russian Academy of Sciences
Contact person: Ivan Nikolayevich Pigarev, pigarev@iitp.ru
The article gives a review to several theories of sleep. According to them, sleep is regarded as a period of recovery of effective brain functioning after active wakefulness. Several studies have demonstrated the association between decreased cognition and increased sleepiness. According to the alternative hypothesis, cognitive impairment is explained by shutdown of certain cortical zones due to their local sleep rather than by impaired brain performance. The studies of asynchronous development of sleep in the cortical zones are described. Causes and conditions for the development of local sleep are discussed in the framework of authirs’ visceral theory of sleep. The¬†theory is based upon an assumption that, during sleep, central parts of the nervous system shift from the analysis of external information to the analysis of internal (visceral) signals. Hypothetical switching network diagram of the main information pathways in the sleep-wake cycle is presented. The authors emphasize that asynchronous switching may explain the development of the number of the sleep-wake cycle-associated pathological conditions (i.e. waking hallucinations, sleep starts, dreams/anxiety dreams, sleepwaking and sleep paralysis). This theory may predict emergence of ‘visceral hallucinations‚Äô which are regarded as a potential cause of seasickness, motion-sickness and some other psychosomatic disorders.
–Ě–ĺ–≤–ĺ—Ā—ā–ł –Ĺ–į —ā–Ķ–ľ—É
10.08.2022 01:00:00
07.07.2022 10:00:00
–ė–Ě–°–Ę–†–£–ú–ē–Ě–Ę–ę
–ě—ā–Ņ—Ä–į–≤–ł—ā—Ć —Ā—ā–į—ā—Ć—é –Ņ–ĺ —ć–Ľ–Ķ–ļ—ā—Ä–ĺ–Ĺ–Ĺ–ĺ–Ļ –Ņ–ĺ—á—ā–Ķ
–í–į—ą –į–ī—Ä–Ķ—Ā —ć–Ľ–Ķ–ļ—ā—Ä–ĺ–Ĺ–Ĺ–ĺ–Ļ –Ņ–ĺ—á—ā—č:
–ź–ī—Ä–Ķ—Ā —ć–Ľ–Ķ–ļ—ā—Ä–ĺ–Ĺ–Ĺ–ĺ–Ļ –Ņ–ĺ—á—ā—č –Ņ–ĺ–Ľ—É—á–į—ā–Ķ–Ľ—Ź:
–†–į–∑–ī–Ķ–Ľ–ł—ā–Ķ –Ĺ–Ķ—Ā–ļ–ĺ–Ľ—Ć–ļ–ĺ –į–ī—Ä–Ķ—Ā–ĺ–≤ —ć–Ľ–Ķ–ļ—ā—Ä–ĺ–Ĺ–Ĺ–ĺ–Ļ –Ņ–ĺ—á—ā—č –∑–į–Ņ—Ź—ā–ĺ–Ļ
–°–ĺ–ĺ–Ī—Č–Ķ–Ĺ–ł–Ķ(–Ĺ–Ķ –ĺ–Ī—Ź–∑–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ)
–Ě–Ķ –Ī–ĺ–Ľ–Ķ–Ķ 1500 —Ā–ł–ľ–≤–ĺ–Ľ–ĺ–≤
–ź–Ĺ—ā–ł —Ā–Ņ–į–ľ:
–Ē–Ľ—Ź –Ņ—Ä–Ķ–ī–ĺ—ā–≤—Ä–į—Č–Ķ–Ĺ–ł—Ź —Ā–Ņ–į–ľ–į, –Ņ–ĺ–∂–į–Ľ—É–Ļ—Ā—ā–į, –≤–≤–Ķ–ī–ł—ā–Ķ –≤ –Ņ–ĺ–Ľ–Ķ —Ā–Ľ–ĺ–≤–ĺ, –ļ–ĺ—ā–ĺ—Ä–ĺ–Ķ –≤–ł–ī–ł—ā–Ķ –Ĺ–ł–∂–Ķ.
–ě–Ī–Ĺ–ĺ–≤–ł—ā—Ć –ļ–ĺ–ī
* –į–ī—Ä–Ķ—Ā–į –Ņ—Ä–Ķ–ī–ĺ—Ā—ā–į–≤–Ľ–Ķ–Ĺ–Ĺ—č–Ķ –í–į–ľ–ł –Ī—É–ī—É—ā –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į—ā—Ć—Ā—Ź —ā–ĺ–Ľ—Ć–ļ–ĺ –ī–Ľ—Ź –ĺ—ā–Ņ—Ä–į–≤–ļ–ł —ć–Ľ–Ķ–ļ—ā—Ä–ĺ–Ĺ–Ĺ–ĺ–Ļ –Ņ–ĺ—á—ā—č.
–£–≤–į–∂–į–Ķ–ľ—č–Ļ –Ņ–ĺ—Ā–Ķ—ā–ł—ā–Ķ–Ľ—Ć uMEDp!
–£–≤–Ķ–ī–ĺ–ľ–Ľ—Ź–Ķ–ľ –í–į—Ā –ĺ —ā–ĺ–ľ, —á—ā–ĺ –∑–ī–Ķ—Ā—Ć —Ā–ĺ–ī–Ķ—Ä–∂–ł—ā—Ā—Ź –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł—Ź, –Ņ—Ä–Ķ–ī–Ĺ–į–∑–Ĺ–į—á–Ķ–Ĺ–Ĺ–į—Ź –ł—Ā–ļ–Ľ—é—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –ī–Ľ—Ź —Ā–Ņ–Ķ—Ü–ł–į–Ľ–ł—Ā—ā–ĺ–≤ –∑–ī—Ä–į–≤–ĺ–ĺ—Ö—Ä–į–Ĺ–Ķ–Ĺ–ł—Ź.
–ē—Ā–Ľ–ł –í—č –Ĺ–Ķ —Ź–≤–Ľ—Ź–Ķ—ā–Ķ—Ā—Ć —Ā–Ņ–Ķ—Ü–ł–į–Ľ–ł—Ā—ā–ĺ–ľ –∑–ī—Ä–į–≤–ĺ–ĺ—Ö—Ä–į–Ĺ–Ķ–Ĺ–ł—Ź, –į–ī–ľ–ł–Ĺ–ł—Ā—ā—Ä–į—Ü–ł—Ź –Ĺ–Ķ –Ĺ–Ķ—Ā–Ķ—ā –ĺ—ā–≤–Ķ—ā—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ—Ā—ā–ł –∑–į –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č–Ķ –ĺ—ā—Ä–ł—Ü–į—ā–Ķ–Ľ—Ć–Ĺ—č–Ķ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź, –≤–ĺ–∑–Ĺ–ł–ļ—ą–ł–Ķ –≤ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ —Ā–į–ľ–ĺ—Ā—ā–ĺ—Ź—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–ł—Ź –í–į–ľ–ł –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł —Ā –Ņ–ĺ—Ä—ā–į–Ľ–į –Ī–Ķ–∑ –Ņ—Ä–Ķ–ī–≤–į—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –ļ–ĺ–Ĺ—Ā—É–Ľ—Ć—ā–į—Ü–ł–ł —Ā –≤—Ä–į—á–ĺ–ľ.
–Ě–į–∂–ł–ľ–į—Ź –Ĺ–į –ļ–Ĺ–ĺ–Ņ–ļ—É ¬ę–í–ĺ–Ļ—ā–ł¬Ľ, –í—č –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–į–Ķ—ā–Ķ, —á—ā–ĺ —Ź–≤–Ľ—Ź–Ķ—ā–Ķ—Ā—Ć –≤—Ä–į—á–ĺ–ľ –ł–Ľ–ł —Ā—ā—É–ī–Ķ–Ĺ—ā–ĺ–ľ –ľ–Ķ–ī–ł—Ü–ł–Ĺ—Ā–ļ–ĺ–≥–ĺ –≤—É–∑–į.