–Т–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ –і–ї—П —Г–Ї—А–µ–њ–ї–µ–љ–Є—П –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ–Њ–є –Ј–∞—Й–Є—В—Л –≤ —Б–≤–µ—В–µ –Є–Љ–Љ—Г–љ–Њ–≥–Њ–Љ–µ–Њ—Б—В–∞—В–Є—З–µ—Б–Ї–Њ–є —А–Њ–ї–Є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л
- –Р–љ–љ–Њ—В–∞—Ж–Є—П
- –°—В–∞—В—М—П
- –°—Б—Л–ї–Ї–Є
- English
–Т–≤–µ–і–µ–љ–Є–µ
–Т –њ–Њ—Б–ї–µ–і–љ–Є–µ 2вАУ3¬†–і–µ—Б—П—В–Є–ї–µ—В–Є—П –њ—А–Њ–Є—Б—Е–Њ–і–Є—В —Б—В—А–µ–Љ–Є—В–µ–ї—М–љ–∞—П —Б–Љ–µ–љ–∞ –Є–Љ–Љ—Г–љ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –њ–∞—А–∞–і–Є–≥–Љ –Є¬†–њ–µ—А–µ–Њ—Б–Љ—Л—Б–ї–µ–љ–Є–µ —А–Њ–ї–Є —Б–Є—Б—В–µ–Љ—Л –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞ –Ї–∞–Ї –Њ–і–љ–Њ–є –Є–Ј –љ–∞–Є–±–Њ–ї–µ–µ –≤–∞–ґ–љ—Л—Е –≥–Њ–Љ–µ–Њ—Б—В–∞—В–Є—З–µ—Б–Ї–Є—Е —Б–Є—Б—В–µ–Љ –Њ—А–≥–∞–љ–Є–Ј–Љ–∞. –°—А–µ–і–Є –љ–∞–Є–±–Њ–ї–µ–µ –Ј–љ–∞—З–Є–Љ—Л—Е —Б–Њ–±—Л—В–Є–є –Ї–Њ–љ—Ж–∞ XX –≤–µ–Ї–∞¬†вАУ –љ–∞—З–∞–ї–∞ –љ—Л–љ–µ—И–љ–µ–≥–Њ —Б—В–Њ–ї–µ—В–Є—П –≤¬†–Њ–±–ї–∞—Б—В–Є –Є–Љ–Љ—Г–љ–Њ–ї–Њ–≥–Є–Є, –Њ–њ—А–µ–і–µ–ї—П—О—Й–Є—Е –≤–µ–Ї—В–Њ—А –µ–µ —А–∞–Ј–≤–Є—В–Є—П, —Б–ї–µ–і—Г–µ—В –≤—Л–і–µ–ї–Є—В—М –Њ—В–Ї—А—Л—В–Є–µ –њ–∞—В—В–µ—А–љ-—А–∞—Б–њ–Њ–Ј–љ–∞—О—Й–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –≤—А–Њ–ґ–і–µ–љ–љ–Њ–≥–Њ –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞ –Є¬†—Б—В—А—Г–Ї—В—Г—А–љ–Њ-—Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–≥–Њ –њ–Њ–ї–Є–Љ–Њ—А—Д–Є–Ј–Љ–∞ CD4+-–Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤. –Т¬†–Ї–Њ–љ—В–µ–Ї—Б—В–µ —Н—В–Є—Е –Њ—В–Ї—А—Л—В–Є–є –Љ–Њ–і–µ—А–љ–Є–Ј–Є—А—Г—О—В—Б—П –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є—П –Њ¬†—А–Њ–ї–Є –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –≤¬†–њ–Њ–і–і–µ—А–ґ–∞–љ–Є–Є –Є–Љ–Љ—Г–љ–љ–Њ–≥–Њ –≥–Њ–Љ–µ–Њ—Б—В–∞–Ј–∞. –С–∞–Ї—В–µ—А–Є–Є-–Ї–Њ–Љ–Љ–µ–љ—Б–∞–ї—Л, –≤–Њ –Љ–љ–Њ–≥–Њ–Љ –±–ї–∞–≥–Њ–і–∞—А—П –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—О –љ–∞ —А–µ—Ж–µ–њ—В–Њ—А—Л –≤—А–Њ–ґ–і–µ–љ–љ–Њ–≥–Њ –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞, –≤¬†–Ї–Њ–љ–µ—З–љ–Њ–Љ –Є—В–Њ–≥–µ –Љ–Њ–і—Г–ї–Є—А—Г—О—В –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–Ї—Г T-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ –≤¬†–љ–∞–њ—А–∞–≤–ї–µ–љ–Є–Є –Ї–∞–Ї —А–µ–≥—Г–ї—П—В–Њ—А–љ—Л—Е, —В–∞–Ї –Є¬†—Н—Д—Д–µ–Ї—В–Њ—А–љ—Л—Е –Ї–ї–µ—В–Њ–Ї, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—П –љ–∞—А—П–і—Г —Б¬†—В–Њ–ї–µ—А–Њ–≥–µ–љ–љ—Л–Љ–Є —Б–Є–≥–љ–∞–ї–∞–Љ–Є —Г–Ї—А–µ–њ–ї–µ–љ–Є–µ –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ–Њ–є –Ј–∞—Й–Є—В—Л –≤¬†–ґ–µ–ї—Г–і–Њ—З–љ–Њ-–Ї–Є—И–µ—З–љ–Њ–Љ —В—А–∞–Ї—В–µ (–Ц–Ъ–Ґ) –Є¬†–Ј–∞ –µ–≥–Њ –њ—А–µ–і–µ–ї–∞–Љ–Є.
–°–≤—П–Ј—М –Є–Љ–Љ—Г–љ–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –Є¬†–Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л —П–≤–ї—П–µ—В—Б—П –Њ–±—К–µ–Ї—В–Њ–Љ –њ—А–Є—Б—В–∞–ї—М–љ–Њ–≥–Њ –≤–љ–Є–Љ–∞–љ–Є—П –љ–µ —В–Њ–ї—М–Ї–Њ –Є–Љ–Љ—Г–љ–Њ–ї–Њ–≥–Њ–≤, –≥–∞—Б—В—А–Њ—Н–љ—В–µ—А–Њ–ї–Њ–≥–Њ–≤ –Є¬†–Љ–Є–Ї—А–Њ–±–Є–Њ–ї–Њ–≥–Њ–≤, –љ–Њ –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–µ–є –Љ–љ–Њ–≥–Є—Е –і—А—Г–≥–Є—Е –Љ–µ–і–Є–Ї–Њ-–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Б–њ–µ—Ж–Є–∞–ї—М–љ–Њ—Б—В–µ–є. –Т¬†–љ–∞—З–∞–ї–µ –Љ–∞—П 2013¬†–≥. –Ї—А—Г–њ–љ–µ–є—И–∞—П –Љ–µ–ґ–і—Г–љ–∞—А–Њ–і–љ–∞—П —Б–µ—В—М –њ–Њ–Є—Б–Ї–Њ–≤—Л—Е —Б–Є—Б—В–µ–Љ Google –љ–∞ –Ј–∞–њ—А–Њ—Б –Є–Ј –Ї–Њ–Љ–±–Є–љ–∞—Ж–Є–Є —Б–ї–Њ–≤ microbiota + immunity –≤—Л–і–∞–≤–∞–ї–∞ –Њ–Ї–Њ–ї–Њ 1,26¬†–Љ–ї–љ —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤, microflora + immunity¬†вАУ 2,63¬†–Љ–ї–љ. –Ф–Њ–њ–Њ–ї–љ–µ–љ–Є–µ –≤—Л—И–µ—Г–Ї–∞–Ј–∞–љ–љ—Л—Е –Ї–Њ–Љ–±–Є–љ–∞—Ж–Є–є –≤¬†–Ј–∞–њ—А–Њ—Б–µ —Б–ї–Њ–≤–Њ–Љ gut (–Ї–Є—И–µ—З–љ—Л–є) —Г–Љ–µ–љ—М—И–∞–ї–Њ —З–Є—Б–ї–Њ –Њ–±–љ–∞—А—Г–ґ–µ–љ–љ—Л—Е web-—Б—В—А–∞–љ–Є—Ж –њ—А–Є–Љ–µ—А–љ–Њ –≤–і–≤–Њ–µ, –Њ–і–љ–∞–Ї–Њ, –љ–µ—Б–Љ–Њ—В—А—П –љ–∞ —Н—В–Њ, –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ —Б—Б—Л–ї–Њ–Ї –≤–њ–µ—З–∞—В–ї—П–µ—В. –Я–Њ–Є—Б–Ї –≤¬†–∞–≤—В–Њ—А–Є—В–µ—В–љ–Њ–є –±–∞–Ј–µ –Љ–µ–і–Є–Ї–Њ-–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є –ї–Є—В–µ—А–∞—В—Г—А—Л PubMed –њ–Њ –Ї–ї—О—З–µ–≤—Л–Љ —Б–ї–Њ–≤–∞–Љ microbiota + immunity –≤—Л—П–≤–ї—П–ї –њ–Њ—З—В–Є 4¬†—В—Л—Б. –њ—Г–±–ї–Є–Ї–∞—Ж–Є–є, –∞¬†–њ–Њ –Ї–Њ–Љ–±–Є–љ–∞—Ж–Є–Є microbiota + immune¬†вАУ –±–Њ–ї–µ–µ 5¬†—В—Л—Б. –љ–∞—Г—З–љ—Л—Е —А–∞–±–Њ—В. –£—В–Њ—З–љ–µ–љ–Є–µ —Н—В–Є—Е –њ–Њ–Є—Б–Ї–Њ–≤—Л—Е –Ј–∞–њ—А–Њ—Б–Њ–≤ —Б–ї–Њ–≤–Њ–Љ gut –ї–Є—И—М –љ–µ–Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ —Б–љ–Є–ґ–∞–ї–Њ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –Њ–±–љ–∞—А—Г–ґ–µ–љ–љ—Л—Е –ї–Є—В–µ—А–∞—В—Г—А–љ—Л—Е –Є—Б—В–Њ—З–љ–Є–Ї–Њ–≤.
–†–∞—Б—В–µ—В —З–Є—Б–ї–Њ –њ—Г–±–ї–Є–Ї–∞—Ж–Є–є, –њ–Њ—Б–≤—П—Й–µ–љ–љ—Л—Е —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–Є–Љ, —Н—Д—Д–µ–Ї—В–∞–Љ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤. –Я—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ–∞—П –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –Ї–Њ—А—А–µ–Ї—Ж–Є–Є –Є¬†–њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є –Є–Љ–Љ—Г–љ–љ—Л—Е —А–∞—Б—Б—В—А–Њ–є—Б—В–≤ –Ј–∞ —Б—З–µ—В –њ–Њ—В—А–µ–±–ї–µ–љ–Є—П –ґ–Є–≤—Л—Е —Б–Є–Љ–±–Є–Њ–љ—В–љ—Л—Е –Љ–Є–Ї—А–Њ–±–Њ–≤ –≤¬†—Ж–µ–ї–Њ–Љ —Г–ґ–µ –љ–µ –≤—Л–Ј—Л–≤–∞–µ—В —Б–Њ–Љ–љ–µ–љ–Є—П [1]. –Ю–і–љ–∞–Ї–Њ —Б–Њ–Њ–±—Й–µ–љ–Є—П –Њ¬†—А–µ–Ј—Г–ї—М—В–∞—В–∞—Е –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ –њ—А–Є —В–µ—Е –Є–ї–Є –Є–љ—Л—Е –Є–Љ–Љ—Г–љ–Њ–Ј–∞–≤–Є—Б–Є–Љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П—Е –Ј–∞—З–∞—Б—В—Г—О –њ—А–Њ—В–Є–≤–Њ—А–µ—З–Є–≤—Л.
–Т—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –±–∞–Ї—В–µ—А–Є–є
–°—А–µ–і–Є –њ—А–∞–Ї—В–Є–Ї—Г—О—Й–Є—Е –≤—А–∞—З–µ–є –і–Є—Б–Ї—Г—В–Є—А—Г—О—В—Б—П –≤–Њ–њ—А–Њ—Б—Л —Б–Њ—Е—А–∞–љ–µ–љ–Є—П –ґ–Є–Ј–љ–µ—Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В–Є —Б–Є–Љ–±–Є–Њ–љ—В–љ—Л—Е –Љ–Є–Ї—А–Њ–±–Њ–≤ –љ–∞ —Н—В–∞–њ–∞—Е –њ—А–Њ–Є–Ј–≤–Њ–і—Б—В–≤–∞/—Е—А–∞–љ–µ–љ–Є—П –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ –Є¬†–њ—А–Є –њ—А–Њ—Е–Њ–ґ–і–µ–љ–Є–Є –∞–≥—А–µ—Б—Б–Є–≤–љ–Њ–є —Б—А–µ–і—Л –ґ–µ–ї—Г–і–Ї–∞ –Є¬†–≤–µ—А—Е–љ–Є—Е –Њ—В–і–µ–ї–Њ–≤ —В–Њ–љ–Ї–Њ–є –Ї–Є—И–Ї–Є. –Ю–±—Б—Г–ґ–і–∞–µ—В—Б—П —В–∞–Ї–ґ–µ —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –њ–µ—А–Њ—А–∞–ї—М–љ–Њ –≤–≤–µ–і–µ–љ–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є –Ї–Њ–ї–Њ–љ–Є–Ј–Є—А–Њ–≤–∞—В—М —Б–ї–Є–Ј–Є—Б—В—Л–µ –і–Є—Б—В–∞–ї—М–љ—Л—Е –Њ—В–і–µ–ї–Њ–≤ –њ–Є—Й–µ–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л. –Я–Њ –љ–∞—И–µ–Љ—Г –Љ–љ–µ–љ–Є—О, –і–∞–љ–љ—Л–µ –њ–∞—А–∞–Љ–µ—В—А—Л –Є–љ–і–Є–≤–Є–і—Г–∞–ї—М–љ—Л, –Ї–∞–ґ–і—Л–є –Ї–Њ–љ–Ї—А–µ—В–љ—Л–є –њ—А–Њ–і—Г–Ї—В –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞—В—М –Њ—В–і–µ–ї—М–љ–Њ. –Ь–љ–Њ–≥–Є–µ —А–∞–Ј—А–∞–±–Њ—В—З–Є–Ї–Є –Є¬†–њ—А–Њ–Є–Ј–≤–Њ–і–Є—В–µ–ї–Є —А–µ—И–Є–ї–Є —В–µ—Е–љ–Є—З–µ—Б–Ї–Є–µ –њ—А–Њ–±–ї–µ–Љ—Л, —Б–≤—П–Ј–∞–љ–љ—Л–µ —Б¬†–≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М—О –±–∞–Ї—В–µ—А–Є–є –Ї–∞–Ї –њ—А–Є –Є–Ј–≥–Њ—В–Њ–≤–ї–µ–љ–Є–Є/—Е—А–∞–љ–µ–љ–Є–Є –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, —В–∞–Ї –Є¬†–њ—А–Є –њ—А–Њ—Е–Њ–ґ–і–µ–љ–Є–Є –≤–µ—А—Е–љ–Є—Е –Њ—В–і–µ–ї–Њ–≤ –Ц–Ъ–Ґ, –Є¬†–і–Њ–Ї–∞–Ј–∞–ї–Є –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –±–Њ–ї–µ–µ –Є–ї–Є –Љ–µ–љ–µ–µ –і–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ –≤—Л—П–≤–ї–µ–љ–Є—П –≤–≤–µ–і–µ–љ–љ—Л—Е —И—В–∞–Љ–Љ–Њ–≤ –≤¬†–Њ–±—А–∞–Ј—Ж–∞—Е —Д–µ–Ї–∞–ї–Є–є [2]. –Я–Њ–і–Њ–±—А–∞–љ—Л –Њ–њ—В–Є–Љ–∞–ї—М–љ—Л–µ —Г—Б–ї–Њ–≤–Є—П –і–ї—П –ї–Є–Њ—Д–Є–ї—М–љ–Њ–є —Б—Г—И–Ї–Є –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—Й–Є–µ –Є—Е –ґ–Є–Ј–љ–µ—Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –Є¬†–і–ї–Є—В–µ–ї—М–љ—Г—О —Б—В–∞–±–Є–ї—М–љ–Њ—Б—В—М –њ—А–Є —Е—А–∞–љ–µ–љ–Є–Є [3]. –Ґ–µ—Е–љ–Њ–ї–Њ–≥–Є—П delayed release (–Ј–∞–Љ–µ–і–ї–µ–љ–љ–Њ–µ –≤—Л—Б–≤–Њ–±–Њ–ґ–і–µ–љ–Є–µ), –Є—Б–њ–Њ–ї—М–Ј—Г–µ–Љ–∞—П –њ—А–Є —Б–Њ–Ј–і–∞–љ–Є–Є —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л—Е –Ј–∞—Й–Є—Й–µ–љ–љ—Л—Е –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –≥–∞—А–∞–љ—В–Є—А—Г–µ—В –±–µ–Ј–Њ–њ–∞—Б–љ—Л–є –ґ–µ–ї—Г–і–Њ—З–љ—Л–є –Є¬†–і—Г–Њ–і–µ–љ–∞–ї—М–љ—Л–є —В—А–∞–љ–Ј–Є—В –ґ–Є–≤—Л—Е –±–∞–Ї—В–µ—А–Є–є –Є¬†–≤—Л—Б–≤–Њ–±–Њ–ґ–і–µ–љ–Є–µ –∞–Ї—В–Є–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–Њ–≤ –≤¬†–і–Є—Б—В–∞–ї—М–љ—Л—Е –Њ—В–і–µ–ї–∞—Е –Ц–Ъ–Ґ. –Я—А–Є —Н—В–Њ–Љ –≤¬†–Ї–∞—З–µ—Б—В–≤–µ —Б–Є—Б—В–µ–Љ –і–Њ—Б—В–∞–≤–Ї–Є —Б–Є–Љ–±–Є–Њ–љ—В–љ—Л—Е –Љ–Є–Ї—А–Њ–±–Њ–≤ –њ—А–µ–і–ї–Њ–ґ–µ–љ—Л –Ї–Є—И–µ—З–љ–Њ—А–∞—Б—В–≤–Њ—А–Є–Љ—Л–µ –Ї–∞–њ—Б—Г–ї—Л, –Ї–Є—Б–ї–Њ—В–Њ—Г—Б—В–Њ–є—З–Є–≤—Л–µ —В–∞–±–ї–µ—В–Ї–Є [4, 5] –Є¬†–Љ–Є–Ї—А–Њ–Ї–∞–њ—Б—Г–ї—Л [6], –Ї–Њ—В–Њ—А—Л–µ, –Ї—А–Њ–Љ–µ —В–Њ–≥–Њ, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—В –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –±–Њ–ї–µ–µ –і–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ —Е—А–∞–љ–µ–љ–Є—П –≥–Њ—В–Њ–≤–Њ–≥–Њ –њ—А–Њ–і—Г–Ї—В–∞ [7].
–Т–Љ–µ—Б—В–µ —Б¬†—В–µ–Љ –і–Њ–Ї–∞–Ј–∞–љ–Њ —Б–Њ—Е—А–∞–љ–µ–љ–Є–µ –ґ–Є–Ј–љ–µ—Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В–Є –љ–µ–Ї–Њ—В–Њ—А—Л—Е –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤ –њ—А–Є –њ—А–Њ—Е–Њ–ґ–і–µ–љ–Є–Є —З–µ—А–µ–Ј –Ц–Ъ–Ґ –Є¬†–≤ –љ–µ–Ј–∞—Й–Є—Й–µ–љ–љ–Њ–є —Д–Њ—А–Љ–µ, —З—В–Њ —Б–≤—П–Ј–∞–љ–Њ —Б¬†—Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В—М—О —Б–∞–Љ–Є—Е –±–∞–Ї—В–µ—А–Є–є –Ї¬†—Б–Њ–ї—П–љ–Њ–є –Ї–Є—Б–ї–Њ—В–µ, —Д–µ—А–Љ–µ–љ—В–∞–Љ –Є¬†–ґ–µ–ї—З–љ—Л–Љ –Ї–Є—Б–ї–Њ—В–∞–Љ. –≠—В–∞ —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В—М —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –≤–∞—А—М–Є—А—Г–µ—В –Љ–µ–ґ–і—Г –≤–Є–і–∞–Љ–Є –Є¬†—И—В–∞–Љ–Љ–∞–Љ–Є –±–∞–Ї—В–µ—А–Є–є, –љ–Њ –≤¬†—Б—А–µ–і–љ–µ–Љ 10вАУ25% –њ–Њ–≥–ї–Њ—Й–µ–љ–љ—Л—Е –±–∞–Ї—В–µ—А–Є–∞–ї—М–љ—Л—Е –Ї–ї–µ—В–Њ–Ї –≤—Л–ґ–Є–≤–∞—О—В –Є¬†–і–Њ—Б—В–Є–≥–∞—О—В –і–Є—Б—В–∞–ї—М–љ—Л—Е –Њ—В–і–µ–ї–Њ–≤ –Ї–Є—И–µ—З–љ–Є–Ї–∞ [8]. –Я—А–Њ–і–µ–Љ–Њ–љ—Б—В—А–Є—А–Њ–≤–∞–љ —Г—Б–њ–µ—И–љ—Л–є —В—А–∞–љ–Ј–Є—В —З–µ—А–µ–Ј –≤–µ—А—Е–љ–Є–µ –Њ—В–і–µ–ї—Л –Ц–Ъ–Ґ —А—П–і–∞ –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –Љ–Є–Ї—А–Њ–±–Њ–≤, —Б–Њ–і–µ—А–ґ–∞—Й–Є—Е—Б—П –≤¬†–є–Њ–≥—Г—А—В–µ [9, 10]. –Ю–і–љ–∞–Ї–Њ –≥–Њ—А–∞–Ј–і–Њ –±–Њ–ї—М—И–∞—П —Б—В–Њ–є–Ї–Њ—Б—В—М –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤ –≤¬†–є–Њ–≥—Г—А—В–∞—Е –Ї¬†–і–µ–є—Б—В–≤–Є—О —Б–Њ–ї—П–љ–Њ–є –Ї–Є—Б–ї–Њ—В—Л, —Д–µ—А–Љ–µ–љ—В–Њ–≤ –Є¬†–ґ–µ–ї—З–љ—Л—Е –Ї–Є—Б–ї–Њ—В –і–Њ—Б—В–Є–≥–∞–µ—В—Б—П –њ—Г—В–µ–Љ –Љ–Є–Ї—А–Њ–Ї–∞–њ—Б—Г–ї—П—Ж–Є–Є [11], —З—В–Њ –њ–Њ–і–і–µ—А–ґ–Є–≤–∞–µ—В —В–µ–Ј–Є—Б –Њ¬†–њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–∞—Е –Ј–∞—Й–Є—Й–µ–љ–љ—Л—Е –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤.
–Т–≤–µ–і–µ–љ–Є–µ –ґ–Є–≤—Л—Е –±–∞–Ї—В–µ—А–Є–є –≤¬†—Б–Њ—З–µ—В–∞–љ–Є–Є —Б¬†–њ—А–µ–±–Є–Њ—В–Є–Ї–∞–Љ–Є —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В –≤–µ—А–Њ—П—В–љ–Њ—Б—В—М –Ї–Њ–ї–Њ–љ–Є–Ј–∞—Ж–Є–Є —Б–ї–Є–Ј–Є—Б—В—Л—Е –Њ–±–Њ–ї–Њ—З–µ–Ї –Ї–Є—И–µ—З–љ–Є–Ї–∞ [12]. –≠—В–Њ —Г—З—В–µ–љ–Њ –љ–µ–Ї–Њ—В–Њ—А—Л–Љ–Є –њ—А–Њ–Є–Ј–≤–Њ–і–Є—В–µ–ї—П–Љ–Є –њ—А–Є —Б–Њ–Ј–і–∞–љ–Є–Є –Ї–Њ–Љ–±–Є–љ–Є—А–Њ–≤–∞–љ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ (—Б–Є–љ–±–Є–Њ—В–Є–Ї–Њ–≤), –≤–Ї–ї—О—З–∞—О—Й–Є—Е –љ–µ —В–Њ–ї—М–Ї–Њ –њ–Њ–ї–µ–Ј–љ—Л–µ –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ—Л, –љ–Њ –Є¬†–Є–љ–≥—А–µ–і–Є–µ–љ—В—Л, –Є–Ј–±–Є—А–∞—В–µ–ї—М–љ–Њ —Б—В–Є–Љ—Г–ї–Є—А—Г—О—Й–Є–µ —А–Њ—Б—В –Є/–Є–ї–Є –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–µ–є –љ–Њ—А–Љ–∞–ї—М–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л.
–Т –Ї–Њ–љ—В–µ–Ї—Б—В–µ –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В–Є –њ–µ—А–Њ—А–∞–ї—М–љ–Њ –≤–≤–µ–і–µ–љ–љ—Л—Е —Б–Є–Љ–±–Є–Њ–љ—В–љ—Л—Е –Љ–Є–Ї—А–Њ–±–Њ–≤ –Њ—В–Љ–µ—В–Є–Љ, —З—В–Њ –і–∞–ґ–µ –Љ–µ—А—В–≤—Л–µ –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–Є —А—П–і–∞ –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤ –Њ–Ї–∞–Ј—Л–≤–∞—О—В –Ј–љ–∞—З–Є–Љ—Л–µ –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л –љ–∞ –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ. –Ш–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–Є–µ —Б–≤–Њ–є—Б—В–≤–∞ –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤ –≤—Л—Е–Њ–і—П—В –Ј–∞ —А–∞–Љ–Ї–Є –Є—Е –ґ–Є–Ј–љ–µ—Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В–Є, –≤¬†—Б–≤—П–Ј–Є —Б¬†—З–µ–Љ –±—Л–ї –њ—А–µ–і–ї–Њ–ґ–µ–љ —В–µ—А–Љ–Є–љ ¬Ђ–њ–∞—А–∞–њ—А–Њ–±–Є–Њ—В–Є–Ї–Є¬ї –і–ї—П –Њ–±–Њ–Ј–љ–∞—З–µ–љ–Є—П –њ—А–Њ–і—Г–Ї—В–Њ–≤ —Б¬†–Є–љ–∞–Ї—В–Є–≤–Є—А–Њ–≤–∞–љ–љ—Л–Љ–Є —Б–Є–Љ–±–Є–Њ–љ—В–∞–Љ–Є, –Њ–Ї–∞–Ј—Л–≤–∞—О—Й–Є–Љ–Є –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –љ–∞ –Ј–і–Њ—А–Њ–≤—М–µ –њ–Њ—В—А–µ–±–Є—В–µ–ї–µ–є [13].
–С–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В—М –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ –Є¬†—А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В—М –Ї¬†–∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л–Љ —Б—А–µ–і—Б—В–≤–∞–Љ
–Р–Ї—В—Г–∞–ї—М–љ—Л–Љ –Є¬†—И–Є—А–Њ–Ї–Њ –Њ–±—Б—Г–ґ–і–∞–µ–Љ—Л–Љ –≤–Њ–њ—А–Њ—Б–Њ–Љ —П–≤–ї—П–µ—В—Б—П –±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В—М –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –±–∞–Ї—В–µ—А–Є–є, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –њ–Њ—В–µ–љ—Ж–Є–∞–ї—М–љ–∞—П –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –њ–µ—А–µ–і–∞—З–Є —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є –Ї¬†–∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л–Љ —Б—А–µ–і—Б—В–≤–∞–Љ –Њ—В —Н—В–Є—Е –±–∞–Ї—В–µ—А–Є–є –њ–∞—В–Њ–≥–µ–љ–∞–Љ –Є–ї–Є —Г—Б–ї–Њ–≤–љ–Њ-–њ–∞—В–Њ–≥–µ–љ–љ—Л–Љ –Љ–Є–Ї—А–Њ–±–∞–Љ. –Ф–µ–є—Б—В–≤–Є—В–µ–ї—М–љ–Њ, –Ї–Є—И–µ—З–љ—Л–є –Љ–Є–Ї—А–Њ–±–Є–Њ–Љ —З–µ–ї–Њ–≤–µ–Ї–∞ –Љ–Њ–ґ–µ—В —Б–ї—Г–ґ–Є—В—М —А–µ–Ј–µ—А–≤—Г–∞—А–Њ–Љ –≥–µ–љ–Њ–≤ —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В–Є –Ї¬†–∞–љ—В–Є–±–Є–Њ—В–Є–Ї–∞–Љ, –Є¬†—Б¬†—Г—З–µ—В–Њ–Љ –≤—Л—Б–Њ–Ї–Њ–є –њ–ї–Њ—В–љ–Њ—Б—В–Є –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤ –≤¬†—Н—В–Њ–є —Н–Ї–Њ—Б–Є—Б—В–µ–Љ–µ —Б—Г—Й–µ—Б—В–≤—Г–µ—В —А–Є—Б–Ї –≥–Њ—А–Є–Ј–Њ–љ—В–∞–ї—М–љ–Њ–є –і–Є—Б—Б–µ–Љ–Є–љ–∞—Ж–Є–Є —Г–Ї–∞–Ј–∞–љ–љ—Л—Е –≥–µ–љ–Њ–≤ –≤–љ—Г—В—А–Є –Љ–Є–Ї—А–Њ–±–љ–Њ–≥–Њ —Б–Њ–Њ–±—Й–µ—Б—В–≤–∞ [14]. –Т¬†–њ–Њ—Б–ї–µ–і–љ–Є–µ –≥–Њ–і—Л, –±–ї–∞–≥–Њ–і–∞—А—П —Б–Њ–≤–µ—А—И–µ–љ—Б—В–≤–Њ–≤–∞–љ–Є—О –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л—Е –Љ–µ—В–Њ–і–Њ–≤ –≤–µ—А–Є—Д–Є–Ї–∞—Ж–Є–Є –≥–µ–љ–Њ–≤ —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є, —Г—В–Њ—З–љ–µ–љ–∞ –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П –Њ¬†—А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–љ–Њ—Б—В–Є —Н—В–Є—Е –≥–µ–љ–Њ–≤ —Б—А–µ–і–Є –Љ–Є–Ї—А–Њ–±–Њ–≤ —А–∞–Ј–ї–Є—З–љ—Л—Е —В–∞–Ї—Б–Њ–љ–Њ–Љ–Є—З–µ—Б–Ї–Є—Е –≥—А—Г–њ–њ, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ —Б—А–µ–і–Є –Є–љ–і–Є–≥–µ–љ–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є. –†–∞—Б—И–Є—Д—А–Њ–≤–∞–љ—Л –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –њ–Њ—П–≤–ї–µ–љ–Є—П, —Б–µ–ї–µ–Ї—Ж–Є–Є, —Е—А–∞–љ–µ–љ–Є—П, –≤–љ—Г—В—А–Є- –Є¬†–Љ–µ–ґ–≤–Є–і–Њ–≤–Њ–є –њ–µ—А–µ–і–∞—З–Є –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ–Њ–є —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В–Є, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–∞ —А–Њ–ї—М –њ–ї–∞–Ј–Љ–Є–і, —В—А–∞—Б–њ–Њ–Ј–Њ–љ–Њ–≤ –Є¬†–Є–љ—В–µ–≥—А–Њ–љ–Њ–≤ [15]. –Э–µ–Ї–Њ—В–Њ—А—Л–µ –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ—Л —Б–∞–Љ–Є —Б–њ–Њ—Б–Њ–±–љ—Л —Б–Є–љ—В–µ–Ј–Є—А–Њ–≤–∞—В—М –∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л–µ –Љ–Њ–ї–µ–Ї—Г–ї—Л, –Є¬†–Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –µ—Б—В–µ—Б—В–≤–µ–љ–љ–Њ–є —Б–µ–ї–µ–Ї—Ж–Є–Є —Г—Б—В–Њ–є—З–Є–≤—Л—Е –Ї¬†–љ–Є–Љ –±–∞–Ї—В–µ—А–Є–є —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–Є—Б—М –µ—Й–µ –і–Њ –њ–Њ—П–≤–ї–µ–љ–Є—П —З–µ–ї–Њ–≤–µ–Ї–∞ –≤¬†—А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–ї–Њ–ґ–љ—Л—Е —Н–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Є¬†—Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ—Л—Е –Њ—В–љ–Њ—И–µ–љ–Є–є –Љ–µ–ґ–і—Г —Б–∞–Љ–Є–Љ–Є –Љ–Є–Ї—А–Њ–±–∞–Љ–Є [16]. –®–Є—А–Њ–Ї–Њ–µ –Є¬†–њ—А–Є —Н—В–Њ–Љ –і–∞–ї–µ–Ї–Њ –љ–µ –≤—Б–µ–≥–і–∞ —А–∞—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–µ –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Њ–≤ –≤¬†–њ–Њ—Б–ї–µ–і–љ–Є–µ –і–µ—Б—П—В–Є–ї–µ—В–Є—П —Г—Б–Є–ї–Є–ї–Њ —Б–µ–ї–µ–Ї—Ж–Є—О –Є¬†—А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є–µ —А–µ–Ј–Є—Б—В–µ–љ—В–љ—Л—Е —И—В–∞–Љ–Љ–Њ–≤, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ —Б—А–µ–і–Є –Є–љ–і–Є–≥–µ–љ–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є, –Ї–Њ—В–Њ—А—Л–µ –±–ї–∞–≥–Њ–і–∞—А—П —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–Њ –Ј–∞–Ї—А–µ–њ–ї–µ–љ–љ—Л–Љ –Ї–Њ–Љ–Љ—Г–љ–Є–Ї–∞—В–Є–≤–љ—Л–Љ –Љ–µ—Е–∞–љ–Є–Ј–Љ–∞–Љ –Љ–Њ–≥—Г—В –њ–µ—А–µ–і–∞–≤–∞—В—М —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г—О—Й–Є–µ –≥–µ–љ—Л –љ–µ —В–Њ–ї—М–Ї–Њ –і—А—Г–≥–Є–Љ –Ї–Њ–Љ–Љ–µ–љ—Б–∞–ї–∞–Љ, –љ–Њ –Є¬†–њ–∞—В–Њ–≥–µ–љ–∞–Љ.
–Я—А–Є–љ–Є–Љ–∞—П –≤–Њ –≤–љ–Є–Љ–∞–љ–Є–µ —В–Њ, —З—В–Њ —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В—М –Ї¬†–∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л–Љ —Б—А–µ–і—Б—В–≤–∞–Љ –і–ї–Є—В–µ–ї—М–љ–Њ–µ –≤—А–µ–Љ—П —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–ї–∞—Б—М –Љ–љ–Њ–≥–Є–Љ–Є —А–∞–Ј—А–∞–±–Њ—В—З–Є–Ї–∞–Љ–Є –Ї–∞–Ї –ґ–µ–ї–∞–µ–Љ–Њ–µ –Ї–∞—З–µ—Б—В–≤–Њ –њ—А–Є –≤—Л–±–Њ—А–µ –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤, –њ–Њ—В–µ–љ—Ж–Є–∞–ї—М–љ–∞—П —Г–≥—А–Њ–Ј–∞ –і–Є—Б—Б–µ–Љ–Є–љ–∞—Ж–Є–Є —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є —Б–Њ —Б—В–Њ—А–Њ–љ—Л –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ —Б—Г—Й–µ—Б—В–≤—Г–µ—В. –Т¬†—Н—В–Њ–є —Б–≤—П–Ј–Є –Я—А–Њ–і–Њ–≤–Њ–ї—М—Б—В–≤–µ–љ–љ–∞—П –Є¬†—Б–µ–ї—М—Б–Ї–Њ—Е–Њ–Ј—П–є—Б—В–≤–µ–љ–љ–∞—П –Њ—А–≥–∞–љ–Є–Ј–∞—Ж–Є—П –Ю–Ю–Э (Food and Agriculture Organization, FAO) –Є¬†–Т—Б–µ–Љ–Є—А–љ–∞—П –Њ—А–≥–∞–љ–Є–Ј–∞—Ж–Є—П –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П (World Health Organization, WHO) —Б—Д–Њ—А–Љ—Г–ї–Є—А–Њ–≤–∞–ї–Є —В—А–µ–±–Њ–≤–∞–љ–Є—П, –Ї–Њ—В–Њ—А—Л–Љ –і–Њ–ї–ґ–љ—Л —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Њ–≤–∞—В—М —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л–µ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є –Є¬†–≤—Е–Њ–і—П—Й–Є–µ –≤¬†–Є—Е —Б–Њ—Б—В–∞–≤ –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ—Л [17]. –Т¬†—З–∞—Б—В–љ–Њ—Б—В–Є, –Є—Б–њ–Њ–ї—М–Ј—Г–µ–Љ—Л–µ –≤¬†–Ї–Њ–љ–µ—З–љ—Л—Е –њ—А–Њ–і—Г–Ї—В–∞—Е –Є¬†–њ—А–µ–њ–∞—А–∞—В–∞—Е –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є–µ —И—В–∞–Љ–Љ—Л –Њ–±—П–Ј–∞—В–µ–ї—М–љ–Њ –і–Њ–ї–ґ–љ—Л –њ—А–Њ—Е–Њ–і–Є—В—М –Ї–Њ–љ—В—А–Њ–ї—М –љ–∞ –њ—А–µ–і–Љ–µ—В –љ–Њ—Б–Є—В–µ–ї—М—Б—В–≤–∞ –≥–µ–љ–Њ–≤ –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Њ—А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є –Є¬†–≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–Є –Є—Е –≥–Њ—А–Є–Ј–Њ–љ—В–∞–ї—М–љ–Њ–є –њ–µ—А–µ–і–∞—З–Є. –Т¬†—Б–Њ—Б—В–∞–≤ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –≤—Л–њ—Г—Б–Ї–∞–µ–Љ—Л—Е –≤¬†—Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Є–Є —Б¬†—В—А–µ–±–Њ–≤–∞–љ–Є—П–Љ–Є FAO/WHO, –≤—Е–Њ–і—П—В —В–Њ–ї—М–Ї–Њ —Е–Њ—А–Њ—И–Њ –Є–Ј—Г—З–µ–љ–љ—Л–µ, —В–∞–Ї—Б–Њ–љ–Њ–Љ–Є—З–µ—Б–Ї–Є –Ї–ї–∞—Б—Б–Є—Д–Є—Ж–Є—А–Њ–≤–∞–љ–љ—Л–µ –Ї—Г–ї—М—В—Г—А—Л, –і–µ–њ–Њ–љ–Є—А–Њ–≤–∞–љ–љ—Л–µ –≤¬†–Љ–µ–ґ–і—Г–љ–∞—А–Њ–і–љ–Њ –њ—А–Є–Ј–љ–∞–љ–љ—Л—Е –Ї–Њ–ї–ї–µ–Ї—Ж–Є—П—Е.
–Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ—Б–ї–µ–і–љ–Є—Е –ї–µ—В –і–µ–Љ–Њ–љ—Б—В—А–Є—А—Г—О—В, —З—В–Њ –њ—А–Њ–±–ї–µ–Љ–∞ –њ—А–Є–Њ–±—А–µ—В–µ–љ–Є—П/—Б–µ–ї–µ–Ї—Ж–Є–Є/–њ–µ—А–µ–і–∞—З–Є –≥–µ–љ–Њ–≤ —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В–Є –Ї¬†–∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л–Љ —Б—А–µ–і—Б—В–≤–∞–Љ —Б–≤—П–Ј–∞–љ–∞ –љ–µ —В–Њ–ї—М–Ї–Њ —Б¬†—И–Є—А–Њ–Ї–Є–Љ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Њ–≤. –Т–Њ –Љ–љ–Њ–≥–Є—Е —Б–ї—Г—З–∞—П—Е —А–µ–Ј–Є—Б—В–µ–љ—В–љ—Л–µ —И—В–∞–Љ–Љ—Л —Б—В–∞–љ–Њ–≤—П—В—Б—П –і–Њ–Љ–Є–љ–∞–љ—В–љ—Л–Љ–Є –Љ–Є–Ї—А–Њ–±–љ—Л–Љ–Є –њ–Њ–њ—Г–ї—П—Ж–Є—П–Љ–Є –і–∞–ґ–µ –≤¬†–Њ—В—Б—Г—В—Б—В–≤–Є–µ —Б–µ–ї–µ–Ї—В–Є–≤–љ–Њ–≥–Њ –і–∞–≤–ї–µ–љ–Є—П —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г—О—Й–Є—Е –∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤, –∞¬†–≥–µ–љ—Л —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В–Є –Ї¬†–∞–љ—В–Є–±–Є–Њ—В–Є–Ї–∞–Љ –≤—Л—П–≤–ї—П—О—В—Б—П —Г¬†–ї–Є—Ж –±–µ–Ј –Є—Б—В–Њ—А–Є–Є –њ—А—П–Љ–Њ–≥–Њ –і–µ–є—Б—В–≤–Є—П —Н—В–Є—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ [18]. –І–∞—Б—В–Њ –Ї–Њ–ї–Њ–љ–Є–Ј–∞—Ж–Є—П —Б–ї–Є–Ј–Є—Б—В—Л—Е –Њ–±–Њ–ї–Њ—З–µ–Ї —А–∞–Ј–ї–Є—З–љ—Л—Е –±–Є–Њ—В–Њ–њ–Њ–≤ —А–µ–Ј–Є—Б—В–µ–љ—В–љ—Л–Љ–Є –Ї¬†—Е–Є–Љ–Є–Њ–њ—А–µ–њ–∞—А–∞—В–∞–Љ —И—В–∞–Љ–Љ–∞–Љ–Є –Ї–Њ–Љ–Љ–µ–љ—Б–∞–ї–Њ–≤ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В —Г–ґ–µ –≤–Њ –≤—А–µ–Љ—П —А–Њ–і–Њ–≤, –≤—Б–Ї–∞—А–Љ–ї–Є–≤–∞–љ–Є—П –Є¬†–Ї–Њ–љ—В–∞–Ї—В–Њ–≤ –≤¬†—А–∞–љ–љ–µ–Љ –њ–Њ—Б—В–љ–∞—В–∞–ї—М–љ–Њ–Љ –њ–µ—А–Є–Њ–і–µ [19]. –Ь–љ–Њ–≥–Є–µ —И–Є—А–Њ–Ї–Њ –њ–Њ—В—А–µ–±–ї—П–µ–Љ—Л–µ –њ–Є—Й–µ–≤—Л–µ –њ—А–Њ–і—Г–Ї—В—Л, –Њ—Б–Њ–±–µ–љ–љ–Њ —Д–µ—А–Љ–µ–љ—В–Є—А–Њ–≤–∞–љ–љ—Л–µ –њ—А–Њ–і—Г–Ї—В—Л –Љ–Њ–ї–Њ—З–љ–Њ–≥–Њ, –Љ—П—Б–љ–Њ–≥–Њ –Є¬†—А–∞—Б—В–Є—В–µ–ї—М–љ–Њ–≥–Њ –њ—А–Њ–Є—Б—Е–Њ–ґ–і–µ–љ–Є—П, —Б–Њ–і–µ—А–ґ–∞—В –ґ–Є–≤—Л–µ –±–∞–Ї—В–µ—А–Є–Є, –љ–µ—Б—Г—Й–Є–µ –њ–µ—А–µ–і–∞–≤–∞–µ–Љ—Л–µ –≥–µ–љ—Л —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В–Є –Ї¬†–∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л–Љ —Б—А–µ–і—Б—В–≤–∞–Љ [20]. –≠—В–Є –≥–µ–љ—Л –≤—Л—П–≤–ї—П—О—В—Б—П —В–∞–Ї–ґ–µ –≤¬†–њ–Њ—З–≤–µ, –≤–Њ–і–µ –Є¬†–і—А—Г–≥–Є—Е –Њ–±—А–∞–Ј—Ж–∞—Е –Њ–Ї—А—Г–ґ–∞—О—Й–µ–є —Б—А–µ–і—Л [21, 22]. –Я–Њ—Б—В–Њ—П–љ–љ–Њ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –Њ–±–Љ–µ–љ –Љ–Є–Ї—А–Њ–±–∞–Љ–Є –Љ–µ–ґ–і—Г –ї—О–і—М–Љ–Є, –∞¬†—В–∞–Ї–ґ–µ –Љ–µ–ґ–і—Г —З–µ–ї–Њ–≤–µ–Ї–Њ–Љ –Є¬†–ґ–Є–≤–Њ—В–љ—Л–Љ–Є, –≤¬†—А–µ–Ј—Г–ї—М—В–∞—В–µ —З–µ–≥–Њ —В–∞–Ї –љ–∞–Ј—Л–≤–∞–µ–Љ—Л–є resistome¬†вАУ —Б–Њ–≤–Њ–Ї—Г–њ–љ–Њ—Б—В—М –≥–µ–љ–Њ–≤ –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ–Њ–є —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В–Є –љ–∞—И–µ–≥–Њ –Љ–Є–Ї—А–Њ–±–Є–Њ–Љ–∞¬†вАУ –њ–Њ–њ–Њ–ї–љ—П–µ—В—Б—П [14].
–£—З–Є—В—Л–≤–∞—П –≤—Л—И–µ—Б–Ї–∞–Ј–∞–љ–љ–Њ–µ, —А–Є—Б–Ї–Є –њ—А–Є–≤–љ–µ—Б–µ–љ–Є—П –љ–Њ–≤—Л—Е –≥–µ–љ–Њ–≤ —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є –Ї¬†–∞–љ—В–Є–±–Є–Њ—В–Є–Ї–∞–Љ, —Б–≤—П–Ј–∞–љ–љ—Л–µ —Б¬†–Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ —Е–Њ—А–Њ—И–Њ –Є–Ј—Г—З–µ–љ–љ—Л—Е –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –Є¬†–њ–µ—А–µ–і–∞—З–Є —Н—В–Є—Е –≥–µ–љ–Њ–≤ –і—А—Г–≥–Є–Љ —А–µ–Ј–Є–і–µ–љ—В–љ—Л–Љ –Є¬†—В—А–∞–љ–Ј–Є—В–Њ—А–љ—Л–Љ –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї—П–Љ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –≤¬†–Њ—Ж–µ–љ–Ї–∞—Е –Љ–љ–Њ–≥–Є—Е —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤ –≤—Л–≥–ї—П–і—П—В –њ—А–µ—Г–≤–µ–ї–Є—З–µ–љ–љ—Л–Љ–Є, –Ј–∞ –љ–µ–Ї–Њ—В–Њ—А—Л–Љ–Є –Є—Б–Ї–ї—О—З–µ–љ–Є—П–Љ–Є. –Ю—Б–Њ–±–Њ–µ –≤–љ–Є–Љ–∞–љ–Є–µ –Є¬†—В—Й–∞—В–µ–ї—М–љ—Л–є –Ї–Њ–љ—В—А–Њ–ї—М –±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В–Є –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л –і–ї—П –≤–Є–і–Њ–≤ –Є¬†—И—В–∞–Љ–Љ–Њ–≤ –±–∞–Ї—В–µ—А–Є–є, –Ї–Њ—В–Њ—А—Л–µ –љ–µ —В–Њ–ї—М–Ї–Њ –Є–Ј–≤–µ—Б—В–љ—Л –Ї–∞–Ї —З–∞—Б—В—Л–µ –љ–Њ—Б–Є—В–µ–ї–Є –њ–µ—А–µ–і–∞–≤–∞–µ–Љ—Л—Е –≥–µ–љ–Њ–≤ —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є, –љ–Њ –Є¬†–Ї–∞–Ї –њ–Њ—В–µ–љ—Ж–Є–∞–ї—М–љ—Л–µ –њ–∞—В–Њ–≥–µ–љ—Л. –Ґ–∞–Ї–Є–Љ–Є —Б–≤–Њ–є—Б—В–≤–∞–Љ–Є –Њ–±–ї–∞–і–∞—О—В, –љ–∞–њ—А–Є–Љ–µ—А, –Љ–љ–Њ–≥–Є–µ –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–Є —А–Њ–і–∞ Enterococcus, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –њ—А–Є–Љ–µ–љ—П–µ–Љ—Л–µ –≤¬†–Ї–∞—З–µ—Б—В–≤–µ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤. –Я–Њ–Љ–Є–Љ–Њ –і–Њ–Ї–∞–Ј–∞–љ–љ–Њ–є —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В–Є –њ–µ—А–µ–і–∞–≤–∞—В—М —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В—М –Ї¬†–∞–љ—В–Є–±–Є–Њ—В–Є–Ї–∞–Љ –і—А—Г–≥–Є–Љ –Љ–Є–Ї—А–Њ–±–∞–Љ, —Н—В–Є –±–∞–Ї—В–µ—А–Є–Є –і–Њ—Б—В–∞—В–Њ—З–љ–Њ —З–∞—Б—В–Њ —П–≤–ї—П—О—В—Б—П –≤–Њ–Ј–±—Г–і–Є—В–µ–ї—П–Љ–Є –љ–Њ–Ј–Њ–Ї–Њ–Љ–Є–∞–ї—М–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–є [23].
–І–∞—Б—В–Њ –Є—Б–њ–Њ–ї—М–Ј—Г–µ–Љ—Л–µ –≤¬†—Б–Њ—Б—В–∞–≤–µ –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –њ—А–Њ–і—Г–Ї—В–Њ–≤ –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–Є —А–Њ–і–∞ Bifidobacterium –≤¬†—А–µ–Ј—Г–ї—М—В–∞—В–µ –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є, –њ—А–Њ–≤–µ–і–µ–љ–љ—Л—Е –≤¬†—А–∞–Ј–љ—Л—Е —Б—В—А–∞–љ–∞—Е, –њ—А–Є–Ј–љ–∞–љ—Л –±–µ–Ј–Њ–њ–∞—Б–љ—Л–Љ–Є. –У–∞—Б—В—А–Њ–Є–љ—В–µ—Б—В–Є–љ–∞–ї—М–љ—Л–µ –Є¬†–≤–љ–µ–Ї–Є—И–µ—З–љ—Л–µ –Є–љ—Д–µ–Ї—Ж–Є–Є, —Б–≤—П–Ј–∞–љ–љ—Л–µ —Б¬†–±–∞–Ї—В–µ—А–Є—П–Љ–Є —Н—В–Њ–≥–Њ —А–Њ–і–∞, –≤—Б—В—А–µ—З–∞—О—В—Б—П –Ї—А–∞–є–љ–µ —А–µ–і–Ї–Њ [24]. –Ґ–µ–Љ –љ–µ –Љ–µ–љ–µ–µ –Њ—В–і–µ–ї—М–љ—Л–µ —И—В–∞–Љ–Љ—Л –±–Є—Д–Є–і–Њ–±–∞–Ї—В–µ—А–Є–є –љ–µ—Б—Г—В –≤¬†—Б–µ–±–µ –≥–µ–љ —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В–Є –Ї¬†—В–µ—В—А–∞—Ж–Є–Ї–ї–Є–љ—Г¬†вАУ tet(W) [25]. L. Masco –Є¬†—Б–Њ–∞–≤—В. –≤—Л—П–≤–Є–ї–Є —Н—В–Њ—В –≥–µ–љ —Г¬†15 –Є–Ј 100¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ—Л—Е —И—В–∞–Љ–Љ–Њ–≤ (50¬†–Є–Ј–Њ–ї—П—В–Њ–≤ –Є–Ј –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –њ—А–Њ–і—Г–Ї—В–Њ–≤ –Є¬†50¬†–і–µ–њ–Њ–љ–Є—А–Њ–≤–∞–љ–љ—Л—Е —И—В–∞–Љ–Љ–Њ–≤) —А–Њ–і–∞ Bifidobacterium, –њ—А–Є —Н—В–Њ–Љ –≤–Њ –≤—Б–µ—Е —Б–ї—Г—З–∞—П—Е –±–∞–Ї—В–µ—А–Є–Є¬†вАУ –љ–Њ—Б–Є—В–µ–ї–Є tet(W) –љ–µ —Б–Њ–і–µ—А–ґ–∞–ї–Є –Ї–Њ–љ—К—О–≥–∞—В–Є–≤–љ—Л–є —В—А–∞—Б–њ–Њ–Ј–Њ–љ TnB1230, –Ї–Њ—В–Њ—А—Л–є —Б—З–Є—В–∞–µ—В—Б—П –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ—Л–Љ –Ј–∞ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є–µ —Н—В–Њ–≥–Њ –≥–µ–љ–∞ –≤¬†–Њ–Ї—А—Г–ґ–∞—О—Й–µ–є —Б—А–µ–і–µ [26]. –≠—В–Њ —Б–Њ–≥–ї–∞—Б—Г–µ—В—Б—П —Б¬†–і–∞–љ–љ—Л–Љ–Є K.P. Scott –Є¬†—Б–Њ–∞–≤—В., –Ї–Њ—В–Њ—А—Л–µ –љ–µ —Б–Љ–Њ–≥–ї–Є –≤—Л—П–≤–Є—В—М —Н—В–Њ—В –Љ–Њ–±–Є–ї—М–љ—Л–є –≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є–є —Н–ї–µ–Љ–µ–љ—В –≤¬†—З–µ–ї–Њ–≤–µ—З–µ—Б–Ї–Є—Е –Є–Ј–Њ–ї—П—В–∞—Е B.¬†longum [27].
–Т —Ж–µ–ї–Њ–Љ –±–µ–Ј–Њ–њ–∞—Б–љ—Л–Љ–Є —В–∞–Ї–ґ–µ –њ—А–Є–Ј–љ–∞—О—В—Б—П –±–Њ–ї—М—И–Є–љ—Б—В–≤–Њ –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤ —А–Њ–і–∞ Lactobacillus [28]. –Х–і–Є–љ–Є—З–љ—Л–µ —Б–ї—Г—З–∞–Є –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї—П—А–љ–Њ–є –Є–љ—Д–µ–Ї—Ж–Є–Є –љ–µ —Б–Њ–њ–Њ—Б—В–∞–≤–Є–Љ—Л —Б¬†–Њ–≥—А–Њ–Љ–љ—Л–Љ–Є –Љ–∞—Б—И—В–∞–±–∞–Љ–Є —Б–ї—Г—З–∞–є–љ–Њ–≥–Њ –Є¬†–љ–∞–Љ–µ—А–µ–љ–љ–Њ–≥–Њ –њ–Њ–≥–ї–Њ—Й–µ–љ–Є—П –ґ–Є–≤—Л—Е –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї. –†–µ—В—А–Њ—Б–њ–µ–Ї—В–Є–≤–љ—Л–є –∞–љ–∞–ї–Є–Ј –њ–Њ–Ї–∞–Ј–∞–ї, —З—В–Њ –≤¬†—А–µ–Ј—Г–ї—М—В–∞—В–µ —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Г–≤–µ–ї–Є—З–µ–љ–Є—П –њ–Њ—В—А–µ–±–ї–µ–љ–Є—П L.¬†rhamnosus GG (–Њ–і–љ–Њ–≥–Њ –Є–Ј –љ–∞–Є–±–Њ–ї–µ–µ –Є–Ј—Г—З–µ–љ–љ—Л—Е –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤) –≤¬†–§–Є–љ–ї—П–љ–і–Є–Є —З–∞—Б—В–Њ—В–∞ —Б–ї—Г—З–∞–µ–≤ –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї—П—А–љ–Њ–є –±–∞–Ї—В–µ—А–Є–µ–Љ–Є–Є –љ–µ —Г–≤–µ–ї–Є—З–Є–ї–∞—Б—М [29]. –†–Є—Б–Ї —А–∞–Ј–≤–Є—В–Є—П –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї–µ–Љ–Є–Є –љ–µ –њ—А–µ–≤—Л—И–∞–µ—В 1¬†—Б–ї—Г—З–∞–є –љ–∞ –Љ–Є–ї–ї–Є–Њ–љ –њ–Њ—В—А–µ–±–Є—В–µ–ї–µ–є [30]. –Ш–Ј–≤–µ—Б—В–љ–∞—П —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В—М –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї –Ї¬†–љ–µ–Ї–Њ—В–Њ—А—Л–Љ –∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л–Љ —Б—А–µ–і—Б—В–≤–∞–Љ, –љ–∞–њ—А–Є–Љ–µ—А –Ї¬†–≤–∞–љ–Ї–Њ–Љ–Є—Ж–Є–љ—Г –Є¬†—В—А–Є–Љ–µ—В–Њ–њ—А–Є–Љ—Г, —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–µ—В—Б—П –Ї–∞–Ї –њ—А–Њ—П–≤–ї–µ–љ–Є–µ –µ—Б—В–µ—Б—В–≤–µ–љ–љ–Њ–є —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є, –Ї–Њ—В–Њ—А–∞—П a priori –љ–µ –њ–µ—А–µ–і–∞–µ—В—Б—П –і—А—Г–≥–Є–Љ –Љ–Є–Ї—А–Њ–±–∞–Љ. –Т–Њ –Љ–љ–Њ–≥–Є—Е –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–∞—Е –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї –≤—Л—П–≤–ї—П–ї–Є—Б—М –Є¬†–њ—А–Є–Њ–±—А–µ—В–µ–љ–љ—Л–µ –≥–µ–љ—Л —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є, –Ї–Њ—В–Њ—А—Л–µ –њ–Њ—В–µ–љ—Ж–Є–∞–ї—М–љ–Њ –Љ–Њ–≥—Г—В –њ–µ—А–µ–і–∞–≤–∞—В—М—Б—П –і—А—Г–≥–Є–Љ –±–∞–Ї—В–µ—А–Є—П–Љ, —З–∞—Й–µ –і—А—Г–≥–Є—Е –≥–µ–љ—Л —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В–Є –Ї¬†—В–µ—В—А–∞—Ж–Є–Ї–ї–Є–љ—Г tet(M), —Н—А–Є—В—А–Њ–Љ–Є—Ж–Є–љ—Г erm(B) –Є¬†—Е–ї–Њ—А–∞–Љ—Д–µ–љ–Є–Ї–Њ–ї—Г cat. –Ю–і–љ–∞–Ї–Њ –і–ї—П –±–Њ–ї—М—И–Є–љ—Б—В–≤–∞ –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–µ–є —А–Њ–і–∞ Lactobacillus –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –≥–Њ—А–Є–Ј–Њ–љ—В–∞–ї—М–љ–Њ–є –њ–µ—А–µ–і–∞—З–Є —Н—В–Є—Е –≥–µ–љ–Њ–≤ –љ–µ –і–Њ–Ї–∞–Ј–∞–љ–∞. –Ы–Є—И—М —Г¬†–љ–µ—Б–Ї–Њ–ї—М–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤ –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї –Њ–±–љ–∞—А—Г–ґ–µ–љ–∞ —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –њ–µ—А–µ–і–∞–≤–∞—В—М –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ—Г—О —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В—М –і—А—Г–≥–Є–Љ –≤–Є–і–∞–Љ, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї—П–Љ —А–Њ–і–∞ Enterococcus [31]. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –і–ї—П –љ–µ–њ–∞—В–Њ–≥–µ–љ–љ—Л—Е –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤ –Њ–њ–∞—Б–љ–Њ—Б—В—М –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В –љ–µ —Б—В–Њ–ї—М–Ї–Њ —Б–∞–Љ–∞ —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В—М –Ї¬†–∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л–Љ —Б—А–µ–і—Б—В–≤–∞–Љ, —Б–Ї–Њ–ї—М–Ї–Њ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –µ–µ –њ–µ—А–µ–і–∞—З–Є –і—А—Г–≥–Є–Љ –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї—П–Љ –Љ–Є–Ї—А–Њ–±–љ–Њ–≥–Њ —Б–Њ–Њ–±—Й–µ—Б—В–≤–∞.
–Т –ї—О–±–Њ–Љ —Б–ї—Г—З–∞–µ –±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В—М –≤—Е–Њ–і—П—Й–Є—Е –≤¬†—Б–Њ—Б—В–∞–≤ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤ –і–Њ–ї–ґ–љ–∞ –±—Л—В—М –≤—Б–µ—Б—В–Њ—А–Њ–љ–љ–µ –Є–Ј—Г—З–µ–љ–∞ —Б–Њ–≥–ї–∞—Б–љ–Њ –њ—А–Є–љ—Ж–Є–њ–∞–Љ, –Ї–Њ—В–Њ—А—Л–µ —Б—Д–Њ—А–Љ—Г–ї–Є—А–Њ–≤–∞–љ—Л –Ї–∞–Ї –≤¬†–≤—Л—И–µ—Г–њ–Њ–Љ—П–љ—Г—В–Њ–Љ –і–Њ–Ї—Г–Љ–µ–љ—В–µ FAO/WHO, —В–∞–Ї –Є¬†–≤¬†–і—А—Г–≥–Є—Е –Љ–µ–ґ–і—Г–љ–∞—А–Њ–і–љ—Л—Е –Є–љ–Є—Ж–Є–∞—В–Є–≤–∞—Е: PROSAFE, QPS, ACE-ART [32]. –Я–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –Ї–Њ—В–Њ—А—Л–µ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г—О—В —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л–Љ –Љ–µ–ґ–і—Г–љ–∞—А–Њ–і–љ—Л–Љ —Б—В–∞–љ–і–∞—А—В–∞–Љ —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є –Є¬†–±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В–Є, –љ–∞ –Ј–і–Њ—А–Њ–≤—М–µ —З–µ–ї–Њ–≤–µ–Ї–∞, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –љ–∞ –Є–Љ–Љ—Г–љ–љ—Л–є –≥–Њ–Љ–µ–Њ—Б—В–∞–Ј –Њ—А–≥–∞–љ–Є–Ј–Љ–∞, —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –њ—А–µ–≤–Њ—Б—Е–Њ–і–Є—В –Ј–љ–∞—З–Є–Љ–Њ—Б—В—М –њ–Њ—В–µ–љ—Ж–Є–∞–ї—М–љ—Л—Е —А–Є—Б–Ї–Њ–≤, —Б–≤—П–Ј–∞–љ–љ—Л—Е —Б¬†–Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ —Г–Ї–∞–Ј–∞–љ–љ—Л—Е –њ—А–Њ–і—Г–Ї—В–Њ–≤.
–Т–љ–Є–Љ–∞–љ–Є—П –Ј–∞—Б–ї—Г–ґ–Є–≤–∞—О—В —Б–Њ–Њ–±—Й–µ–љ–Є—П –Њ¬†–њ–Њ–њ—Л—В–Ї–∞—Е –њ—А–µ–і–Њ—В–≤—А–∞—В–Є—В—М –Ї–Њ–ї–Њ–љ–Є–Ј–∞—Ж–Є—О –Њ—А–≥–∞–љ–Є–Ј–Љ–∞ —А–µ–Ј–Є—Б—В–µ–љ—В–љ—Л–Љ–Є —И—В–∞–Љ–Љ–∞–Љ–Є –Ј–∞ —Б—З–µ—В –њ–µ—А–Њ—А–∞–ї—М–љ–Њ–≥–Њ –≤–≤–µ–і–µ–љ–Є—П –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Њ—З—Г–≤—Б—В–≤–Є—В–µ–ї—М–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є [33], –љ–µ–Ї–Њ—В–Њ—А—Л–µ –Є–Ј –љ–Є—Е –±—Л–ї–Є –≤–µ—Б—М–Љ–∞ —Г—Б–њ–µ—И–љ—Л–Љ–Є [34]. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є –Љ–Њ–≥—Г—В –±—Л—В—М –љ–µ —В–Њ–ї—М–Ї–Њ —Д–∞–Ї—В–Њ—А–Њ–Љ —А–Є—Б–Ї–∞, –љ–Њ –Є¬†–Є–љ—Б—В—А—Г–Љ–µ–љ—В–Њ–Љ –±–Њ—А—М–±—Л —Б¬†—Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В—М—О –Ї¬†–∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л–Љ —Б—А–µ–і—Б—В–≤–∞–Љ.
–†–Њ–ї—М –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –≤¬†—Н–≤–Њ–ї—О—Ж–Є–Є –Є–Љ–Љ—Г–љ–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е
–Ф–Њ–ї–≥–Њ–µ –≤—А–µ–Љ—П —Н–≤–Њ–ї—О—Ж–Є—П –Є–Љ–Љ—Г–љ–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –ґ–Є–≤–Њ—В–љ—Л—Е —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–ї–∞—Б—М –≤¬†–Ї–Њ–љ—В–µ–Ї—Б—В–µ –њ–Њ—Б—В–Њ—П–љ–љ–Њ–≥–Њ –њ—А–µ—Б—Б–Є–љ–≥–∞ —Б–Њ —Б—В–Њ—А–Њ–љ—Л –њ–∞—В–Њ–≥–µ–љ–Њ–≤. –Я—А–µ–Њ–±–ї–∞–і–∞–ї–Њ –Љ–љ–µ–љ–Є–µ, —З—В–Њ –Є–Љ–µ–љ–љ–Њ –њ–Њ–і –≤–ї–Є—П–љ–Є–µ–Љ –њ–Њ—Б—В–Њ—П–љ–љ—Л—Е –≤—Л–Ј–Њ–≤–Њ–≤ –Њ—В –Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Л—Е –∞–≥–µ–љ—В–Њ–≤ —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–∞—Б—М —Б–ї–Њ–ґ–љ–∞—П —Б–Є—Б—В–µ–Љ–∞ –≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Њ–є –≤–∞—А–Є–∞–±–µ–ї—М–љ–Њ—Б—В–Є, –њ–Њ–Ј–≤–Њ–ї—П—О—Й–∞—П –≥–µ–љ–µ—А–Є—А–Њ–≤–∞—В—М –Њ–≥—А–Њ–Љ–љ–Њ–µ —З–Є—Б–ї–Њ –Љ–Њ–ї–µ–Ї—Г–ї, —В–∞–Ї–Є—Е –Ї–∞–Ї –∞–љ—В–Є—В–µ–ї–∞, —Б¬†–њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –љ–µ–Њ–≥—А–∞–љ–Є—З–µ–љ–љ—Л–Љ–Є —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—П–Љ–Є —А–∞—Б–њ–Њ–Ј–љ–∞–≤–∞—В—М –≤–љ–Њ–≤—М –њ–Њ—П–≤–ї—П—О—Й–Є–µ—Б—П –њ–∞—В–Њ–≥–µ–љ—Л. –Т¬†–њ–Њ–і–і–µ—А–ґ–Ї—Г —Н—В–Њ–≥–Њ –Љ–љ–µ–љ–Є—П –њ—А–Є–≤–Њ–і–Є–ї—Б—П –њ—А–Є–Љ–µ—А –≤–Ј–∞–Є–Љ–Њ—Б–≤—П–Ј–Є –њ–Њ—П–≤–ї–µ–љ–Є—П —З–µ–ї—О—Б—В–µ–є —Г¬†—А—Л–± –Є¬†—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—П –∞–і–∞–њ—В–Є–≤–љ–Њ–≥–Њ –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞ [35]. –§–Є–ї–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є –њ–Њ—П–≤–ї–µ–љ–Є–µ —З–µ–ї—О—Б—В–µ–є –і–∞–ї–Њ —А—Л–±–∞–Љ –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–Њ –≤¬†–њ–Є—В–∞–љ–Є–Є, –њ–Њ–Ј–≤–Њ–ї–Є–ї–Њ —Г–≤–µ–ї–Є—З–Є—В—М –Є–Љ —Б–≤–Њ–Є —А–∞–Ј–Љ–µ—А—Л –Є¬†–њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М –ґ–Є–Ј–љ–Є, –љ–Њ –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ –≤—Л—А–Њ—Б —А–Є—Б–Ї —В—А–∞–≤–Љ–∞—В–Є–Ј–∞—Ж–Є–Є –Є¬†–Є–љ—Д–Є—Ж–Є—А–Њ–≤–∞–љ–Є—П, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –њ–Њ–≤—В–Њ—А–љ–Њ–≥–Њ, —Б–ї–Є–Ј–Є—Б—В—Л—Е –Њ–±–Њ–ї–Њ—З–µ–Ї –Ї–Є—И–µ—З–љ–Є–Ї–∞. –°—З–Є—В–∞–ї–Њ—Б—М, —З—В–Њ —Н—В–Њ—В –њ—А–Њ—Ж–µ—Б—Б –њ–Њ—Б–ї—Г–ґ–Є–ї –Њ—В–њ—А–∞–≤–љ–Њ–є —В–Њ—З–Ї–Њ–є –і–ї—П —А–∞–Ј–≤–Є—В–Є—П –Є–Љ–Љ—Г–љ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є –њ–∞–Љ—П—В–Є [36].
–Ю–і–љ–∞–Ї–Њ —Б–µ–є—З–∞—Б –њ—А–Њ–Є—Б—Е–Њ–і–Є—В —В—А–∞–љ—Б—Д–Њ—А–Љ–∞—Ж–Є—П –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є–є. –Я–Њ–ґ–∞–ї—Г–є, –±–Њ–ї–µ–µ –ї–Њ–≥–Є—З–љ—Л–Љ–Є –Є¬†–Њ–±–Њ—Б–љ–Њ–≤–∞–љ–љ—Л–Љ–Є —П–≤–ї—П—О—В—Б—П –і–Њ–≤–Њ–і—Л –Њ¬†—В–Њ–Љ, —З—В–Њ –Ї–Њ–Љ–Љ–µ–љ—Б–∞–ї—Л –Њ–Ї–∞–Ј—Л–≤–∞—О—В –±–Њ–ї—М—И–µ–µ, —З–µ–Љ –њ–∞—В–Њ–≥–µ–љ—Л, —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–Њ–µ –і–∞–≤–ї–µ–љ–Є–µ –љ–∞ –Є–Љ–Љ—Г–љ–љ—Г—О —Б–Є—Б—В–µ–Љ—Г –Є¬†–њ–Њ—В–Њ–Љ—Г –Є–Љ –њ—А–Є–љ–∞–і–ї–µ–ґ–Є—В –±–Њ–ї—М—И–∞—П —А–Њ–ї—М –≤¬†—Д–Є–ї–Њ–≥–µ–љ–µ–Ј–µ —Н—В–Њ–є —Б–Є—Б—В–µ–Љ—Л [37, 38]. –Я–Њ —Б–∞–Љ—Л–Љ —Б–і–µ—А–ґ–∞–љ–љ—Л–Љ –Њ—Ж–µ–љ–Ї–∞–Љ, –Љ–љ–Њ–≥–Њ–Ї–ї–µ—В–Њ—З–љ—Л–µ –Њ—А–≥–∞–љ–Є–Ј–Љ—Л –Є–Љ–µ—О—В —А–µ–Ј–Є–і–µ–љ—В–љ—Г—О –Љ–Є–Ї—А–Њ—Д–ї–Њ—А—Г –±–Њ–ї–µ–µ –њ–Њ–ї—Г–Љ–Є–ї–ї–Є–∞—А–і–∞ –ї–µ—В [39, 40]. –Т—Б–µ —Н—В–Њ –≤—А–µ–Љ—П –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ—Л –Є¬†–Љ–Є–Ї—А–Њ–±—Л –Ї–Њ-—Н–≤–Њ–ї—О—Ж–Є–Њ–љ–Є—А—Г—О—В –Є¬†–∞–і–∞–њ—В–Є—А—Г—О—В—Б—П –і—А—Г–≥ –Ї¬†–і—А—Г–≥—Г. –Ц–Є–≤–Њ—В–љ—Л–µ —П–≤–Є–ї–Є—Б—М –і–ї—П –Ј–∞—Б–µ–ї–Є–≤—И–Є—Е –Є—Е –±–∞–Ї—В–µ—А–Є–є —Б—В–∞–±–Є–ї—М–љ—Л–Љ–Є —Н–Ї–Њ—Б–Є—Б—В–µ–Љ–∞–Љ–Є, –Є–Ј–Њ–±–Є–ї—Г—О—Й–Є–Љ–Є –љ—Г—В—А–Є–µ–љ—В–∞–Љ–Є. –Т¬†—Н—В–Њ–є —Б–≤—П–Ј–Є –Ј–і–Њ—А–Њ–≤—М–µ —Е–Њ–Ј—П–Є–љ–∞ –њ—А–Є–Њ–±—А–µ–ї–Њ –њ–µ—А–≤–Њ—Б—В–µ–њ–µ–љ–љ–Њ–µ –Ј–љ–∞—З–µ–љ–Є–µ –і–ї—П –µ–≥–Њ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л [38].
–Ь–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ—Л –Ї–Њ–ї–Њ–љ–Є–Ј–Є—А–Њ–≤–∞–ї–Є —А–∞–Ј–ї–Є—З–љ—Л–µ —Г—З–∞—Б—В–Ї–Є –љ–∞—И–µ–≥–Њ —В–µ–ї–∞, –≤–Ї–ї—О—З–∞—П –Ї–Њ–ґ—Г, –њ–Њ–ї–Њ—Б—В—М —А—В–∞, —Б–ї–Є–Ј–Є—Б—В—Л–µ –Љ–Њ—З–µ–њ–Њ–ї–Њ–≤–Њ–є –Є¬†–і—Л—Е–∞—В–µ–ї—М–љ–Њ–є —Б–Є—Б—В–µ–Љ, —Г—Е–Њ, –Њ–і–љ–∞–Ї–Њ –Є–Љ–µ–љ–љ–Њ –љ–Є–ґ–љ–Є–µ –Њ—В–і–µ–ї—Л –Ц–Ъ–Ґ —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—В—Б—П –љ–∞–Є–±–Њ–ї—М—И–µ–є –њ–ї–Њ—В–љ–Њ—Б—В—М—О –Є¬†—А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–Є–µ–Љ –Ї–Њ–Љ–Љ–µ–љ—Б–∞–ї–Њ–≤. –Т¬†–і–Є—Б—В–∞–ї—М–љ–Њ–є —З–∞—Б—В–Є —В–Њ–љ–Ї–Њ–є –Ї–Є—И–Ї–Є –Є¬†–Њ—Б–Њ–±–µ–љ–љ–Њ –≤¬†—В–Њ–ї—Б—В–Њ–є –Ї–Є—И–Ї–µ –Њ–±–Є—В–∞—О—В –±–∞–Ї—В–µ—А–Є–Є, –∞—А—Е–µ–Є, –≥—А–Є–±—Л, –њ—А–Њ—Б—В–µ–є—И–Є–µ –Є¬†–≤ –љ–µ–Ї–Њ—В–Њ—А—Л—Е —Б–ї—Г—З–∞—П—Е –Љ–љ–Њ–≥–Њ–Ї–ї–µ—В–Њ—З–љ—Л–µ –≥–µ–ї—М–Љ–Є–љ—В—Л. –Ю–і–љ–∞–Ї–Њ –±–∞–Ї—В–µ—А–Є–Є, –±–µ–Ј—Г—Б–ї–Њ–≤–љ–Њ, –і–Њ–Љ–Є–љ–Є—А—Г—О—В: –Є—Е –Њ–±—Й–µ–µ —З–Є—Б–ї–Њ –≤¬†—В–Њ–ї—Б—В–Њ–є –Ї–Є—И–Ї–µ –і–Њ—Б—В–Є–≥–∞–µ—В 100¬†—В—А–ї–љ. –Э–µ–і–∞–≤–љ–Є–µ –Љ–µ—В–∞–≥–µ–љ–Њ–Љ–љ—Л–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л —З–µ–ї–Њ–≤–µ–Ї–∞ –≤—Л—П–≤–Є–ї–Є –Њ–Ї–Њ–ї–Њ 3,3¬†–Љ–ї–љ –≥–µ–љ–Њ–≤, —З—В–Њ –њ—А–Є–Љ–µ—А–љ–Њ –≤¬†150¬†—А–∞–Ј –њ—А–µ–≤–Њ—Б—Е–Њ–і–Є—В —З–Є—Б–ї–Њ –≥–µ–љ–Њ–≤ —Б–∞–Љ–Њ–≥–Њ —З–µ–ї–Њ–≤–µ–Ї–∞ [41].
–Ь–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є–µ —Д–Є–ї–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є –љ–∞—Г—З–Є–ї–Є—Б—М –Є–Ј–≤–ї–µ–Ї–∞—В—М –њ–Њ–ї—М–Ј—Г –Њ—В —В–∞–Ї–Њ–є –Њ–≥—А–Њ–Љ–љ–Њ–є –Љ–∞—Б—Б—Л —А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–љ—Л—Е —А–µ–Ј–Є–і–µ–љ—В–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є, –Ї–Њ—В–Њ—А—Л–µ, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є, –њ–Њ–Љ–Њ–≥–∞—О—В –њ–µ—А–µ–≤–∞—А–Є–≤–∞—В—М –Ї–Њ–Љ–њ–ї–µ–Ї—Б–љ—Л–µ —Г–≥–ї–µ–≤–Њ–і—Л –Є¬†—Б–љ–∞–±–ґ–∞—О—В –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ –≤–∞–ґ–љ–µ–є—И–Є–Љ–Є –љ—Г—В—А–Є–µ–љ—В–∞–Љ–Є. –Ю–і–љ–∞–Ї–Њ —А–Њ–ї—М –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –і–∞–ї–µ–Ї–Њ –љ–µ –Њ–≥—А–∞–љ–Є—З–Є–≤–∞–µ—В—Б—П —Г—З–∞—Б—В–Є–µ–Љ –≤¬†–њ–Є—Й–µ–≤–∞—А–µ–љ–Є–Є.
–Т—Б–µ –±–Њ–ї—М—И–µ –љ–∞—Г—З–љ—Л—Е –њ–Њ–і—В–≤–µ—А–ґ–і–µ–љ–Є–є –љ–∞—Е–Њ–і–Є—В –њ—А–Є–≤–µ–і–µ–љ–љ–Њ–µ –љ–∞–Љ–Є —А–∞–љ–µ–µ –Њ–±—А–∞–Ј–љ–Њ–µ –≤—Л—Б–Ї–∞–Ј—Л–≤–∞–љ–Є–µ –Њ¬†—В–Њ–Љ, —З—В–Њ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є–µ, –≤–Ї–ї—О—З–∞—П —З–µ–ї–Њ–≤–µ–Ї–∞, –Ј–∞–Ї–ї—О—З–Є–ї–Є —Б–Њ –Љ–љ–Њ–≥–Є–Љ–Є –Є–Ј —А–µ–Ј–Є–і–µ–љ—В–љ—Л—Е –Љ–Є–Ї—А–Њ–±–Њ–≤ –љ–µ —В–Њ–ї—М–Ї–Њ ¬Ђ–њ–∞–Ї—В –Њ¬†–љ–µ–љ–∞–њ–∞–і–µ–љ–Є–Є¬ї, –љ–Њ –Є¬†¬Ђ–і–Њ–≥–Њ–≤–Њ—А –Њ¬†–≤–Ј–∞–Є–Љ–Њ–њ–Њ–Љ–Њ—Й–Є¬ї –љ–∞ —Б–ї—Г—З–∞–є –≤–љ–µ—И–љ–Є—Е —Г–≥—А–Њ–Ј, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –Є—Б—Е–Њ–і—П—Й–Є—Е –Њ—В –њ–∞—В–Њ–≥–µ–љ–љ—Л—Е –≤–Є—А—Г—Б–Њ–≤ –Є¬†–±–∞–Ї—В–µ—А–Є–є [1].
–Ш–љ—В–µ—А–µ—Б–љ—Л–є –∞–ї—М—В–µ—А–љ–∞—В–Є–≤–љ—Л–є –≤–Ј–≥–ї—П–і –љ–∞ —Д–Є–ї–Њ–≥–µ–љ–µ–Ј –Є¬†—Д—Г–љ–Ї—Ж–Є—О –Є–Љ–Љ—Г–љ–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –њ—А–Є–≤–Њ–і–Є—В E.T. Harvill [42]. –Э–µ—Б–Љ–Њ—В—А—П –љ–∞ ¬Ђ—Б–Њ–≤–µ—А—И–µ–љ–љ—Г—О¬ї –Ј–∞—Й–Є—В—Г, —Б–≤—П–Ј–∞–љ–љ—Г—О —Б¬†–≤—Л—А–∞–±–Њ—В–Ї–Њ–є –Њ–≥—А–Њ–Љ–љ–Њ–≥–Њ —А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–Є—П –њ–∞—В–Њ–≥–µ–љ-—Б–њ–µ—Ж–Є—Д–Є—З–µ—Б–Ї–Є—Е –Љ–Њ–ї–µ–Ї—Г–ї, –љ–∞—И –Њ—А–≥–∞–љ–Є–Ј–Љ –љ–µ —Б–≤–Њ–±–Њ–і–µ–љ –Њ—В –±–Њ–ї–µ–Ј–љ–µ—В–≤–Њ—А–љ—Л—Е –Љ–Є–Ї—А–Њ–±–Њ–≤ –Є¬†–≤—А—П–і –ї–Є –±–Њ–ї–µ–µ —Г—Б–њ–µ—И–µ–љ –≤¬†–±–Њ—А—М–±–µ —Б¬†–њ–Њ—Б–ї–µ–і–љ–Є–Љ–Є, —З–µ–Љ —А–∞—Б—В–µ–љ–Є—П –Є¬†–љ–∞—Б–µ–Ї–Њ–Љ—Л–µ, –Ї–Њ—В–Њ—А—Л–µ –љ–µ –Є–Љ–µ—О—В –∞–і–∞–њ—В–Є–≤–љ–Њ–≥–Њ –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞. –Т¬†–љ–∞—З–∞–ї–µ 1970-—Е –≥–≥. –±—Л–ї–∞ –≤—Л–і–≤–Є–љ—Г—В–∞ –≥–Є–њ–Њ—В–µ–Ј–∞ вАЬRed QueenвАЭ (–≤ —А—Г—Б—Б–Ї–Њ–Љ –њ–µ—А–µ–≤–Њ–і–µ¬†вАУ ¬Ђ–І–µ—А–љ–Њ–є –Ъ–Њ—А–Њ–ї–µ–≤—Л¬ї), –≤¬†—Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Є–Є —Б¬†–Ї–Њ—В–Њ—А–Њ–є –Љ–µ–ґ–і—Г –њ–∞—В–Њ–≥–µ–љ–∞–Љ–Є –Є¬†–Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–Љ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В ¬Ђ—Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–∞—П –≥–Њ–љ–Ї–∞ –≤–Њ–Њ—А—Г–ґ–µ–љ–Є–є¬ї: —З–µ–Љ –±–Њ–ї–µ–µ —Б–ї–Њ–ґ–љ–Њ–є –Є¬†—Б–Њ–≤–µ—А—И–µ–љ–љ–Њ–є —Б—В–∞–љ–Њ–≤–Є—В—Б—П –Є–Љ–Љ—Г–љ–љ–∞—П –Ј–∞—Й–Є—В–∞ —Е–Њ–Ј—П–Є–љ–∞, —В–µ–Љ –ґ–µ—Б—В—З–µ —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ—Л–є –Њ—В–±–Њ—А –њ–∞—А–∞–Ј–Є—В–Њ–≤, –Є¬†–љ–∞–Њ–±–Њ—А–Њ—В [43]. –≠—В–Њ –і–∞–µ—В —Б–µ–ї–µ–Ї—В–Є–≤–љ—Л–µ –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–∞ —Е–Њ–Ј—П–Є–љ—Г –њ–µ—А–µ–і –і—А—Г–≥–Є–Љ–Є –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–∞–Љ–Є, –Њ–і–љ–∞–Ї–Њ —Б–Њ—А–µ–≤–љ–Њ–≤–∞–љ–Є–µ —Б¬†–±–Њ–ї–µ–Ј–љ–µ—В–≤–Њ—А–љ—Л–Љ–Є –Љ–Є–Ї—А–Њ–±–∞–Љ–Є a¬†priori –Њ–±—А–µ—З–µ–љ–Њ –љ–∞ –љ–µ—Г–і–∞—З—Г –≤¬†—Б–≤—П–Ј–Є —Б¬†–±–Њ–ї—М—И–µ–є —Б–Ї–Њ—А–Њ—Б—В—М—О —Н–≤–Њ–ї—О—Ж–Є–Є –њ–∞—В–Њ–≥–µ–љ–Њ–≤. –Ъ–∞–Ї —Б–Ї–∞–Ј–∞–ї–∞ –І–µ—А–љ–∞—П –Ъ–Њ—А–Њ–ї–µ–≤–∞ –Є–Ј –њ—А–Њ–Є–Ј–≤–µ–і–µ–љ–Є—П –Ы—М—О–Є—Б–∞ –Ъ—Н—А—А–Њ–ї–ї–∞ ¬Ђ–Р–ї–Є—Б–∞ –≤¬†–Ч–∞–Ј–µ—А–Ї–∞–ї—М–µ¬ї, ¬Ђ–њ—А–Є—Е–Њ–і–Є—В—Б—П –±–µ–ґ–∞—В—М —Б–Њ –≤—Б–µ—Е –љ–Њ–≥, —З—В–Њ–±—Л —В–Њ–ї—М–Ї–Њ –Њ—Б—В–∞—В—М—Б—П –љ–∞ —В–Њ–Љ –ґ–µ –Љ–µ—Б—В–µ¬ї.
E.T. Harvill —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–µ—В –Є–Љ–Љ—Г–љ–Є—В–µ—В (–≤–Ї–ї—О—З–∞—П –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Г—О –Ј–∞—Й–Є—В—Г –Є¬†—В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В—М) –Ї–∞–Ї –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –Ї–Њ–Љ–њ–ї–µ–Ї—Б–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л, –≤–∞–ґ–љ–µ–є—И–µ–є –Ј–∞–і–∞—З–µ–є –Ї–Њ—В–Њ—А–Њ–є —П–≤–ї—П–µ—В—Б—П –њ–Њ–і–і–µ—А–ґ–∞–љ–Є–µ –Љ–Є–Ї—А–Њ–±–Є–Њ–Љ–∞ (–Ї–Њ–Љ–Љ–µ–љ—Б–∞–ї–Њ–≤ –Є¬†—Б–Є–Љ–±–Є–Њ–љ—В–Њ–≤), –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—Й–µ–≥–Њ –Є–ї–Є —Г–ї—Г—З—И–∞—О—Й–µ–≥–Њ –Љ–љ–Њ–≥–Є–µ –ґ–Є–Ј–љ–µ–љ–љ–Њ –≤–∞–ґ–љ—Л–µ —Д—Г–љ–Ї—Ж–Є–Є –љ–∞—И–µ–≥–Њ –Њ—А–≥–∞–љ–Є–Ј–Љ–∞ [42]. –Я–Њ –µ–≥–Њ –Љ–љ–µ–љ–Є—О, –Љ–љ–Њ–≥–Є–µ –Є–Ј–≤–µ—Б—В–љ—Л–µ –Є¬†–љ–µ–Є–Ј–≤–µ—Б—В–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –Ј–∞—Й–Є—В—Л –Њ—В –њ–∞—В–Њ–≥–µ–љ–Њ–≤¬†вАУ —Н—В–Њ —З–∞—Б—В—М —Б–Є—Б—В–µ–Љ—Л –њ–Њ–і–і–µ—А–ґ–∞–љ–Є—П –Ј–і–Њ—А–Њ–≤–Њ–≥–Њ –Љ–Є–Ї—А–Њ–±–Є–Њ–Љ–∞. –Т¬†—Н—В–Њ–є —Б–≤—П–Ј–Є –љ–µ —Г–і–Є–≤–Є—В–µ–ї—М–љ–Њ, —З—В–Њ –њ—А–µ–Њ–±–ї–∞–і–∞—О—Й–Є–є –Є–Ј–Њ—В–Є–њ –∞–љ—В–Є—В–µ–ї —Б–ї–Є–Ј–Є—Б—В—Л—Е –Њ–±–Њ–ї–Њ—З–µ–Ї IgA –љ–∞—Ж–µ–ї–µ–љ –љ–∞ —А–µ–Ј–Є–і–µ–љ—В–љ—Г—О –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Г, –љ–Њ –њ—А–Є —Н—В–Њ–Љ –Њ–±–µ—Б–њ–µ—З–Є–≤–∞–µ—В –љ–µ –Ј–∞—Й–Є—В—Г –Њ—В —Б–Є–Љ–±–Є–Њ–љ—В–Њ–≤, –∞¬†–Є–Љ–Љ—Г–љ–љ—Г—О —В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В—М –Ї¬†–љ–Є–Љ. –Т–њ–Њ–ї–љ–µ –≤–µ—А–Њ—П—В–љ–Њ, —З—В–Њ —Д–Є–ї–Њ–≥–µ–љ–µ–Ј —Б–Є—Б—В–µ–Љ—Л –∞–і–∞–њ—В–Є–≤–љ–Њ–≥–Њ –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞ —Б–≤—П–Ј–∞–љ —Б¬†–љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ—Б—В—М—О –±—Л—Б—В—А–Њ –њ—А–Є—Б–њ–Њ—Б–∞–±–ї–Є–≤–∞—В—М—Б—П –Ї¬†–љ–Њ–≤—Л–Љ –Ї–Њ–Љ–Љ–µ–љ—Б–∞–ї–∞–Љ –Є¬†—Б–Є–Љ–±–Є–Њ–љ—В–∞–Љ, –Ї–Њ—В–Њ—А—Л–µ –њ–Њ–њ–∞–і–∞—О—В –≤¬†–љ–∞—И –Њ—А–≥–∞–љ–Є–Ј–Љ —Б¬†–љ–Њ–≤–Њ–є –њ–Є—Й–µ–є –Є¬†–њ–Њ–Љ–Њ–≥–∞—О—В —Е–Њ–Ј—П–Є–љ—Г –і–µ—В–Њ–Ї—Б–Є—Ж–Є—А–Њ–≤–∞—В—М –Є¬†–њ–µ—А–µ–≤–∞—А–Є–≤–∞—В—М —Н—В—Г –њ–Є—Й—Г –Є, –Ї—А–Њ–Љ–µ —В–Њ–≥–Њ, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—В –Ї–Њ–ї–Њ–љ–Є–Ј–∞—Ж–Є–Њ–љ–љ—Г—О —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В—М –Є¬†–і—А—Г–≥–Є–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –Ј–∞—Й–Є—В—Л –Њ—В –њ–∞—В–Њ–≥–µ–љ–Њ–≤.
–Ь–Є–Ї—А–Њ–±–Є–Њ—В–∞ –љ–µ —В–Њ–ї—М–Ї–Њ —Г–њ—А–∞–≤–ї—П–µ—В —Д–Є–ї–Њ–≥–µ–љ–µ–Ј–Њ–Љ –Є–Љ–Љ—Г–љ–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л, –љ–Њ –Є¬†–Њ–њ—А–µ–і–µ–ї—П–µ—В —Е–∞—А–∞–Ї—В–µ—А –Њ–љ—В–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Њ–≥–Њ —А–∞–Ј–≤–Є—В–Є—П –Љ—Г–Ї–Њ–Ј–∞–ї—М–љ–Њ–≥–Њ –Є¬†–Њ–±—Й–µ–≥–Њ –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞, –∞¬†—В–∞–Ї–ґ–µ —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М –Є¬†–∞–і–µ–Ї–≤–∞—В–љ–Њ—Б—В—М –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ–Њ–≥–Њ –Њ—В–≤–µ—В–∞. –Э–∞—А—Г—И–µ–љ–Є–µ —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–Њ —Б–ї–Њ–ґ–Є–≤—И–Є—Е—Б—П —Н—В–∞–њ–Њ–≤ –љ–µ–Њ–љ–∞—В–∞–ї—М–љ–Њ–≥–Њ –Є¬†–њ–Њ—Б—В–љ–∞—В–∞–ї—М–љ–Њ–≥–Њ –Ј–∞—Б–µ–ї–µ–љ–Є—П –Љ–Є–Ї—А–Њ–±–∞–Љ–Є –љ–∞—И–µ–≥–Њ –Њ—А–≥–∞–љ–Є–Ј–Љ–∞, –Њ—Б–Њ–±–µ–љ–љ–Њ –Ц–Ъ–Ґ, –≤–µ–і–µ—В –Ї¬†–Є–Љ–Љ—Г–љ–Њ–Ј–∞–≤–Є—Б–Є–Љ—Л–Љ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П–Љ [44]. –≠—В–Њ –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В—Б—П –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П–Љ–Є –Є–Љ–Љ—Г–љ–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л —Г¬†–≥–љ–Њ—В–Њ–±–Є–Њ–љ—В–Њ–≤ (–±–µ–Ј–Љ–Є–Ї—А–Њ–±–љ—Л—Е –ґ–Є–≤–Њ—В–љ—Л—Е), —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—Й–Є—Е—Б—П —Б–ї–∞–±–Њ—А–∞–Ј–≤–Є—В–Њ–є –Љ—Г–Ї–Њ–Ј–∞-–∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ–љ–Њ–є –ї–Є–Љ—Д–Њ–Є–і–љ–Њ–є —В–Ї–∞–љ—М—О, –≥–Є–њ–Њ–њ–ї–∞—Б—В–Є—З–µ—Б–Ї–Є–Љ–Є –њ–µ–є–µ—А–Њ–≤—Л–Љ–Є –±–ї—П—И–Ї–∞–Љ–Є, —Г–Љ–µ–љ—М—И–µ–љ–Є–µ–Љ —З–Є—Б–ї–∞ CD4+-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ –Є¬†IgA-–њ—А–Њ–і—Г—Ж–Є—А—Г—О—Й–Є—Е –њ–ї–∞–Ј–Љ–∞—В–Є—З–µ—Б–Ї–Є—Е –Ї–ї–µ—В–Њ–Ї –≤¬†—Б–Њ–±—Б—В–≤–µ–љ–љ–Њ–є –њ–ї–∞—Б—В–Є–љ–Ї–µ —Б–ї–Є–Ј–Є—Б—В–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є, –њ—А–Є —Н—В–Њ–Љ –≤¬†–ї–Є–Љ—Д–Њ–Є–і–љ—Л—Е –Њ—А–≥–∞–љ–∞—Е –Њ—В–Љ–µ—З–∞–µ—В—Б—П —Б–і–≤–Є–≥ –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–Ї–Є –Ґ-–Ї–ї–µ—В–Њ–Ї –≤¬†—Б—В–Њ—А–Њ–љ—Г T-—Е–µ–ї–њ–µ—А–Њ–≤ 2¬†—В–Є–њ–∞ (ThвВВ) –Є¬†—Б–љ–Є–ґ–µ–љ–љ–∞—П –њ—А–Њ–і—Г–Ї—Ж–Є—П –Є–љ—В–µ—А—Д–µ—А–Њ–љ–∞ (–Ш–§–Э) –≥–∞–Љ–Љ–∞ [45, 46].
–Ь–Є–Ї—А–Њ–±–Є–Њ—В–∞ –Є¬†–Ј–і–Њ—А–Њ–≤—М–µ —Е–Њ–Ј—П–Є–љ–∞
–Т –њ–Њ—Б–ї–µ–і–љ–Є–µ –≥–Њ–і—Л —Г–≤–µ–ї–Є—З–Є–ї–Њ—Б—М —З–Є—Б–ї–Њ —А–∞–±–Њ—В –њ–Њ –Є–Ј—Г—З–µ–љ–Є—О –Љ–Є–Ї—А–Њ–±–Є–Њ–Љ–∞ –≤¬†–Ї–Њ–љ—В–µ–Ї—Б—В–µ –Ј–і–Њ—А–Њ–≤—М—П –Є¬†–Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є —З–µ–ї–Њ–≤–µ–Ї–∞ [47]. –≠—В–Є–Њ–ї–Њ–≥–Є—П –Љ–љ–Њ–≥–Є—Е –±–Њ–ї–µ–Ј–љ–µ–є –≤–Ї–ї—О—З–∞–µ—В –љ–µ —В–Њ–ї—М–Ї–Њ –≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є–µ, –љ–Њ –Є¬†—Н–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ —Д–∞–Ї—В–Њ—А—Л. –І–∞—Б—В–Њ—В–∞ —В–∞–Ї–Є—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є, –Ї–∞–Ї –Њ–ґ–Є—А–µ–љ–Є–µ, –і–Є–∞–±–µ—В, –∞—В–µ—А–Њ—Б–Ї–ї–µ—А–Њ–Ј, –љ–µ—Г–Ї–ї–Њ–љ–љ–Њ —А–∞—Б—В–µ—В –≤¬†–њ–Њ—Б–ї–µ–і–љ–Є–µ –і–µ—Б—П—В–Є–ї–µ—В–Є—П, —З—В–Њ –≤¬†–Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–є —Б—В–µ–њ–µ–љ–Є —Б–≤—П–Ј—Л–≤–∞—О—В —Б¬†—Д–∞–Ї—В–Њ—А–∞–Љ–Є –Њ–Ї—А—Г–ґ–∞—О—Й–µ–є —Б—А–µ–і—Л [48]. –Ш–Ј–Љ–µ–љ–µ–љ–Є–µ –Њ–±—А–∞–Ј–∞ –Є¬†—Е–∞—А–∞–Ї—В–µ—А–∞ –њ–Є—В–∞–љ–Є—П, –∞¬†—В–∞–Ї–ґ–µ –њ—А—П–Љ–Њ–µ –Є¬†–Њ–њ–Њ—Б—А–µ–і–Њ–≤–∞–љ–љ–Њ–µ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ –∞–љ—В–Є–Љ–Є–Ї—А–Њ–±–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Њ–Ї–∞–Ј—Л–≤–∞—О—В —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –љ–∞ —Б–Њ—Б—В–∞–≤ –Є¬†—Д—Г–љ–Ї—Ж–Є—О –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л —З–µ–ї–Њ–≤–µ–Ї–∞, —З—В–Њ —П–≤–ї—П–µ—В—Б—П –≤–∞–ґ–љ–µ–є—И–Є–Љ —Н–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ —Д–∞–Ї—В–Њ—А–Њ–Љ —А–∞–Ј–≤–Є—В–Є—П —Г–Ї–∞–Ј–∞–љ–љ—Л—Е –±–Њ–ї–µ–Ј–љ–µ–є [49]. –Ь–Є–Ї—А–Њ–±–Є–Њ–Љ –Є–≥—А–∞–µ—В —Б—Г—Й–µ—Б—В–≤–µ–љ–љ—Г—О —А–Њ–ї—М –≤¬†—Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–Њ–Љ –±–∞–ї–∞–љ—Б–µ, —Н–љ–і–Њ–Ї—А–Є–љ–љ—Л—Е —Б–Є–≥–љ–∞–ї–∞—Е –Є¬†–Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е —А–∞—Б—Б—В—А–Њ–є—Б—В–≤–∞—Е, –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ–љ—Л—Е —Б¬†–Њ–ґ–Є—А–µ–љ–Є–µ–Љ, —В–∞–Ї–Є—Е –Ї–∞–Ї –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–∞—П —Н–љ–і–Њ—В–Њ–Ї—Б–µ–Љ–Є—П [50].
–Я–Њ—Б—В–µ–њ–µ–љ–љ–Њ –њ—А–Њ—П—Б–љ—П–µ—В—Б—П —Д—Г–љ–Ї—Ж–Є—П –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –≤¬†–Ї–Њ–љ—В–µ–Ї—Б—В–µ –≤–Ј–∞–Є–Љ–Њ—Б–≤—П–Ј–Є –Ц–Ъ–Ґ –Є¬†—Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л (–¶–Э–°). C –Њ—Б—М—О ¬Ђ–Љ–Є–Ї—А–Њ–±–Є–Њ—В–∞¬†вАУ –Ї–Є—И–µ—З–љ–Є–Ї¬†вАУ –Љ–Њ–Ј–≥¬ї –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ—Л –≤–Њ–Ј—А–∞—Б—В–љ–∞—П —Н–≤–Њ–ї—О—Ж–Є—П –Є¬†—А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–љ—Л–µ —Д—Г–љ–Ї—Ж–Є–Є –¶–Э–°, –≤–Ї–ї—О—З–∞—П –њ–Њ–≤–µ–і–µ–љ–Є–µ –Є¬†–љ–∞—Б—В—А–Њ–µ–љ–Є–µ. –≠—В–∞ –Њ—Б—М, –≤–µ—А–Њ—П—В–љ–Њ, –Є–Љ–µ–µ—В –±–Њ–ї—М—И–Њ–µ –Ј–љ–∞—З–µ–љ–Є–µ –≤¬†—А–∞–Ј–≤–Є—В–Є–Є —В–∞–Ї–Є—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є —З–µ–ї–Њ–≤–µ–Ї–∞, –Ї–∞–Ї –∞—Г—В–Є–Ј–Љ. –Ф–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є, –≤–Ї–ї—О—З–∞—О—Й–Є–µ –±–Є—Д–Є–і–Њ–±–∞–Ї—В–µ—А–Є–Є –Є¬†–ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї—Л, —Б–љ–Є–ґ–∞—О—В —В—А–µ–≤–Њ–≥—Г —Г¬†–ґ–Є–≤–Њ—В–љ—Л—Е, –∞¬†—Г¬†–±–Њ–ї—М–љ—Л—Е –ї—О–і–µ–є –Њ–±—Г—Б–ї–Њ–≤–ї–Є–≤–∞—О—В –Љ–љ–Њ–≥–Є–µ –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ—Л–µ —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ —Н—Д—Д–µ–Ї—В—Л, –≤–Ї–ї—О—З–∞—П —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П –Ї–Њ—А—В–Є–Ј–Њ–ї–∞ –≤¬†—Б—Л–≤–Њ—А–Њ—В–Ї–µ –Ї—А–Њ–≤–Є [51].
–Т—Л—П–≤–ї–µ–љ—Л –≤—Л—А–∞–ґ–µ–љ–љ—Л–µ –Є–Ј–Љ–µ–љ–µ–љ–Є—П –Љ–Є–Ї—А–Њ–±–Є–Њ—Ж–µ–љ–Њ–Ј–∞ –Ц–Ъ–Ґ —Г¬†–±–Њ–ї—М–љ—Л—Е —А–µ–≤–Љ–∞—В–Њ–Є–і–љ—Л–Љ –∞—А—В—А–Є—В–Њ–Љ, —Б–Є—Б—В–µ–Љ–љ–Њ–є –Ї—А–∞—Б–љ–Њ–є –≤–Њ–ї—З–∞–љ–Ї–Њ–є –Є¬†–љ–µ–Ї–Њ—В–Њ—А—Л–Љ–Є –і—А—Г–≥–Є–Љ–Є –∞—Г—В–Њ–Є–Љ–Љ—Г–љ–љ—Л–Љ–Є –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П–Љ–Є [52]. –Э–µ –≤—Л–Ј—Л–≤–∞–µ—В —Б–Њ–Љ–љ–µ–љ–Є–є —Б–≤—П–Ј—М —А–∞—Б—Б—В—А–Њ–є—Б—В–≤ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л —Б¬†—А–∞–Ј–≤–Є—В–Є–µ–Љ –±–Њ–ї–µ–Ј–љ–Є –Ъ—А–Њ–љ–∞ –Є¬†—П–Ј–≤–µ–љ–љ–Њ–≥–Њ –Ї–Њ–ї–Є—В–∞ [53]. –Р—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ–љ—Л–µ —Б¬†–і–Є—Б–±–Є–Њ–Ј–Њ–Љ –љ–∞—А—Г—И–µ–љ–Є—П –Є–Љ–Љ—Г–љ–љ–Њ–≥–Њ –≥–Њ–Љ–µ–Њ—Б—В–∞–Ј–∞, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤ —В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В–Є, —Г–≤–µ–ї–Є—З–Є–≤–∞—О—В —А–Є—Б–Ї –∞–ї–ї–µ—А–≥–Є—З–µ—Б–Ї–Є—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є [54].
–Т –љ–µ–і–∞–≤–љ–µ–є —А–∞–±–Њ—В–µ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ–Њ –њ–Њ–і—В–≤–µ—А–ґ–і–µ–љ–∞ –≥–Є–њ–Њ—В–µ–Ј–∞ –Њ¬†—В–Њ–Љ, —З—В–Њ –Љ–Є–Ї—А–Њ–±–Є–Њ—В–∞ –Є–≥—А–∞–µ—В —А–Њ–ї—М –≤¬†–њ–Њ–і–і–µ—А–ґ–∞–љ–Є–Є ¬Ђ—Б–Є—П–љ–Є—П –Ј–і–Њ—А–Њ–≤—М—П¬ї, —В–Њ –µ—Б—В—М —В–∞–Ї–Є—Е —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї, –Ї–∞–Ї –±–ї–µ—Б—В—П—Й–Є–µ –≤–Њ–ї–Њ—Б—Л –Є¬†–Ј–і–Њ—А–Њ–≤–∞—П –Ї–Њ–ґ–∞, –Ї–Њ—В–Њ—А—Л–µ —Б–≤—П–Ј—Л–≤–∞—О—В —Б¬†—Д–µ—А—В–Є–ї—М–љ–Њ—Б—В—М—О –Є¬†–Љ–Њ–ї–Њ–і–Њ—Б—В—М—О. –Я—А–Є —Н—В–Њ–Љ –і–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є –Љ–Њ–≥—Г—В –≤–Њ—Б—Б—В–∞–љ–∞–≤–ї–Є–≤–∞—В—М —Н—В–Є –≤–љ–µ—И–љ–Є–µ –њ—А–Њ—П–≤–ї–µ–љ–Є—П —А–µ–њ—А–Њ–і—Г–Ї—В–Є–≤–љ—Л—Е —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В–µ–є –њ—А–Є —Б—В–∞—А–µ–љ–Є–Є –Њ—А–≥–∞–љ–Є–Ј–Љ–∞ [55].
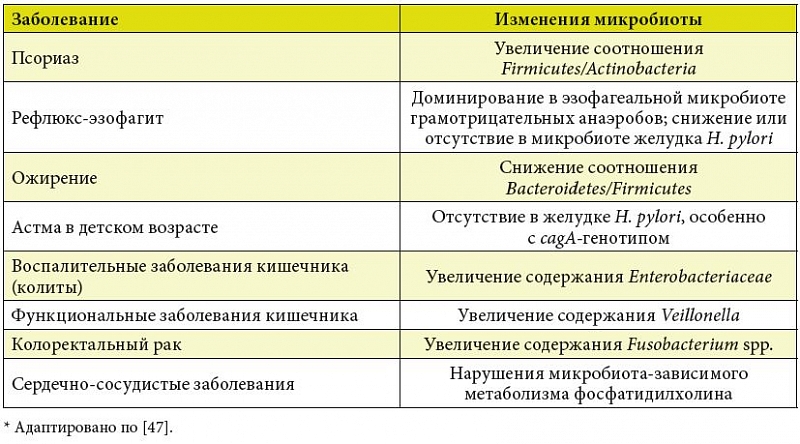
I. Cho –Є¬†M.J. Blaser –Њ–±–Њ–±—Й–Є–ї–Є –і–∞–љ–љ—Л–µ –њ–Њ—Б–ї–µ–і–љ–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –≤–Ј–∞–Є–Љ–Њ—Б–≤—П–Ј–Є –њ–∞—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Б–Њ—Б—В–Њ—П–љ–Є–є —З–µ–ї–Њ–≤–µ–Ї–∞ –Є¬†—А–∞—Б—Б—В—А–Њ–є—Б—В–≤ –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л (—В–∞–±–ї.¬†1)¬†[47]. –Я–ї–µ–є–Њ—В—А–Њ–њ–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –Љ–Є–Ї—А–Њ–±–Є–Њ–Љ–∞ –љ–∞ —Б–Њ—Б—В–Њ—П–љ–Є–µ –Ј–і–Њ—А–Њ–≤—М—П –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –≤–Ї–ї—О—З–∞—П —З–µ–ї–Њ–≤–µ–Ї–∞, –њ–Њ–Ј–≤–Њ–ї—П–µ—В —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞—В—М –Є—Е –љ–µ –њ—А–Њ—Б—В–Њ –Ї–∞–Ї –Є–Ј–Њ–ї–Є—А–Њ–≤–∞–љ–љ—Л–µ –≤–Є–і—Л, –∞¬†–Ї–∞–Ї —Б–ї–Њ–ґ–љ—Л–µ —Н–Ї–Њ—Б–Є—Б—В–µ–Љ—Л, –≤¬†–Ї–Њ—В–Њ—А—Л—Е –Њ–≥—А–Њ–Љ–љ–Њ–µ —З–Є—Б–ї–Њ –Љ–Є–Ї—А–Њ–±–Њ–≤ –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤—Г–µ—В —Б¬†–Ї–ї–µ—В–Ї–∞–Љ–Є, –Њ—А–≥–∞–љ–∞–Љ–Є –Є¬†—Б–Є—Б—В–µ–Љ–∞–Љ–Є –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–∞, –∞¬†—В–∞–Ї–ґ–µ –і—А—Г–≥ —Б¬†–і—А—Г–≥–Њ–Љ, –Є—Б–њ–Њ–ї—М–Ј—Г—П —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–Њ –Ј–∞–Ї—А–µ–њ–ї–µ–љ–љ—Л–µ —Б–Є–≥–љ–∞–ї—М–љ—Л–µ –Є¬†–і—А—Г–≥–Є–µ –Ї–Њ–Љ–Љ—Г–љ–Є–Ї–∞—В–Є–≤–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л.
–†–Њ–ї—М –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –≤¬†–њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ–Њ–є –Ј–∞—Й–Є—В–µ
–Т–∞–ґ–љ–∞—П —А–Њ–ї—М –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –≤¬†–Ј–∞—Й–Є—В–µ –Њ—В –њ–∞—В–Њ–≥–µ–љ–Њ–≤ –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –≤¬†–њ–Є—Й–µ–≤–∞—А–Є—В–µ–ї—М–љ–Њ–Љ —В—А–∞–Ї—В–µ, –Њ–±—Г—Б–ї–Њ–≤–ї–µ–љ–љ–∞—П —А–∞–Ј–ї–Є—З–љ—Л–Љ–Є –Љ–µ—Е–∞–љ–Є–Ј–Љ–∞–Љ–Є –Ї–Њ–ї–Њ–љ–Є–Ј–∞—Ж–Є–Њ–љ–љ–Њ–є —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є [56вАУ58], –і–∞–≤–љ–Њ —Г–ґ–µ –љ–µ –≤—Л–Ј—Л–≤–∞–µ—В —Б–Њ–Љ–љ–µ–љ–Є–є. –Ю—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –љ–µ–і–∞–≤–љ–Њ –њ–Њ–ї—Г—З–µ–љ—Л –і–Њ–Ї–∞–Ј–∞—В–µ–ї—М—Б—В–≤–∞ —В–Њ–≥–Њ, —З—В–Њ –Є–љ–і–Є–≥–µ–љ–љ—Л–µ –±–∞–Ї—В–µ—А–Є–Є —А–µ–≥—Г–ї–Є—А—Г—О—В –Ј–∞—Й–Є—В–љ—Л–µ –Є–Љ–Љ—Г–љ–љ—Л–µ —А–µ–∞–Ї—Ж–Є–Є –Є¬†–Ј–∞ –њ—А–µ–і–µ–ї–∞–Љ–Є –њ–Є—Й–µ–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –≤¬†—А–µ—Б–њ–Є—А–∞—В–Њ—А–љ–Њ–Љ —В—А–∞–Ї—В–µ. –£—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ –≤–ї–Є—П–љ–Є–µ –Ї–Є—И–µ—З–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є-–Ї–Њ–Љ–Љ–µ–љ—Б–∞–ї–Њ–≤ –љ–∞ —А–µ–∞–ї–Є–Ј–∞—Ж–Є—О –Є–Љ–Љ—Г–љ–љ–Њ–≥–Њ –Њ—В–≤–µ—В–∞ –њ—А–Њ—В–Є–≤ –≤–Є—А—Г—Б–∞ –≥—А–Є–њ–њ–∞, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –љ–∞ —Б–Њ–Ј—А–µ–≤–∞–љ–Є–µ –≤–Є—А—Г—Б-—Б–њ–µ—Ж–Є—Д–Є—З–µ—Б–Ї–Є—Е CD4+- –Є¬†–°D8+-–Ґ-–Ї–ї–µ—В–Њ–Ї. –Т¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞—Е in vivo –Є—Б—В–Њ—Й–µ–љ–Є–µ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –њ–µ—А–Њ—А–∞–ї—М–љ—Л–Љ –њ—А–Є–µ–Љ–Њ–Љ –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Њ–≤ —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ —Б–љ–Є–ґ–∞–ї–Њ —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В—М –Ї¬†–Є–љ—В—А–∞–љ–∞–Ј–∞–ї—М–љ–Њ–Љ—Г –Ј–∞—А–∞–ґ–µ–љ–Є—О –≤–Є—А—Г—Б–Њ–Љ –≥—А–Є–њ–њ–∞ –Р¬†–Є¬†—Г–≤–µ–ї–Є—З–Є–≤–∞–ї–Њ –ї–µ—В–∞–ї—М–љ–Њ—Б—В—М –Є–љ—Д–Є—Ж–Є—А–Њ–≤–∞–љ–љ—Л—Е –ґ–Є–≤–Њ—В–љ—Л—Е. –£–Ї–∞–Ј–∞–љ–љ—Л–µ –і–µ—Д–µ–Ї—В—Л –њ—А–Њ—В–Є–≤–Њ–≤–Є—А—Г—Б–љ–Њ–є –Ј–∞—Й–Є—В—Л –±—Л–ї–Є —Б–≤—П–Ј–∞–љ—Л —Б–Њ —Б–љ–Є–ґ–µ–љ–Є–µ–Љ –Ґ- –Є¬†–Т-–Ї–ї–µ—В–Њ—З–љ—Л—Е –Є–Љ–Љ—Г–љ–љ—Л—Е —А–µ–∞–Ї—Ж–Є–є, –∞¬†—В–∞–Ї–ґ–µ –њ–Њ–і–∞–≤–ї–µ–љ–Є–µ–Љ —Д—Г–љ–Ї—Ж–Є–Є –Є¬†–Љ–Є–≥—А–∞—Ж–Є–Є –і–µ–љ–і—А–Є—В–љ—Л—Е –Ї–ї–µ—В–Њ–Ї –≤¬†—А–µ—Б–њ–Є—А–∞—В–Њ—А–љ–Њ–Љ —В—А–∞–Ї—В–µ [59]. –≠—В–Є –і–∞–љ–љ—Л–µ —Б–Њ–≥–ї–∞—Б—Г—О—В—Б—П —Б¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–Љ–Є –љ–∞–±–ї—О–і–µ–љ–Є—П–Љ–Є, –≥–Њ–≤–Њ—А—П—Й–Є–Љ–Є –Њ–± —Г—В—П–ґ–µ–ї–µ–љ–Є–Є —В–µ—З–µ–љ–Є—П –≥—А–Є–њ–њ–∞ –Є¬†–і—А—Г–≥–Є—Е —А–µ—Б–њ–Є—А–∞—В–Њ—А–љ—Л—Е –≤–Є—А—Г—Б–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–є –њ—А–Є –њ—А–Є–µ–Љ–µ –∞–љ—В–Є–±–∞–Ї—В–µ—А–Є–∞–ї—М–љ—Л—Е —Б—А–µ–і—Б—В–≤, –≤–µ–і—Г—Й–µ–Љ –Ї¬†–њ–Њ–і–∞–≤–ї–µ–љ–Є—О –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л.
–Т –љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П —А–∞—Б–Ї—А—Л—В—Л –љ–µ–Ї–Њ—В–Њ—А—Л–µ –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л, –Ј–∞ —Б—З–µ—В –Ї–Њ—В–Њ—А—Л—Е –Ї–Є—И–µ—З–љ—Л–µ –±–∞–Ї—В–µ—А–Є–Є —Г—Б–Є–ї–Є–≤–∞—О—В –њ—А–Њ—В–Є–≤–Њ–≤–Є—А—Г—Б–љ—Л–є –Њ—В–≤–µ—В.
–Т–Њ-–њ–µ—А–≤—Л—Е, —Н—В–Њ —Б—В–Є–Љ—Г–ї—П—Ж–Є—П –Њ–±—А–∞–Ј-—А–∞—Б–њ–Њ–Ј–љ–∞—О—Й–Є—Е Toll-–њ–Њ–і–Њ–±–љ—Л—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ (TLR). TLR-2 —А–∞—Б–њ–Њ–Ј–љ–∞—О—В –ї–Є–њ–Њ—В–µ–є—Е–Њ–µ–≤—Л–µ –Ї–Є—Б–ї–Њ—В—Л –Є¬†–ї–Є–њ–Њ–њ—А–Њ—В–µ–Є–љ—Л –±–∞–Ї—В–µ—А–Є–∞–ї—М–љ—Л—Е –Ї–ї–µ—В–Њ—З–љ—Л—Е —Б—В–µ–љ–Њ–Ї. TLR-4/MD-2 —П–≤–ї—П—О—В—Б—П —Б–µ–љ—Б–Њ—А–∞–Љ–Є –ї–Є–њ–Њ–њ–Њ–ї–Є—Б–∞—Е–∞—А–Є–і–Њ–≤ –≥—А–∞–Љ–Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є. TLR-9 —А–∞—Б–њ–Њ–Ј–љ–∞—О—В –љ–µ–Љ–µ—В–Є–ї–Є—А–Њ–≤–∞–љ–љ—Л–µ –°pG-–њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ—Б—В–Є –±–∞–Ї—В–µ—А–Є–∞–ї—М–љ–Њ–є –Ф–Э–Ъ. –†–µ–Ї—В–∞–ї—М–љ–Њ–µ –≤–≤–µ–і–µ–љ–Є–µ –ї–Є–≥–∞–љ–і–Њ–≤ TLR –Љ—Л—И–∞–Љ –≤–Њ—Б—Б—В–∞–љ–∞–≤–ї–Є–≤–∞–ї–Њ –њ–Њ–і–∞–≤–ї–µ–љ–љ—Л–є –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–∞–Љ–Є –њ—А–Њ—В–Є–≤–Њ–≤–Є—А—Г—Б–љ—Л–є –Є–Љ–Љ—Г–љ–љ—Л–є –Њ—В–≤–µ—В –Є¬†—Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В—М –Ї¬†–Є–љ—В—А–∞–љ–∞–Ј–∞–ї—М–љ–Њ–Љ—Г –Ј–∞—А–∞–ґ–µ–љ–Є—О –≤–Є—А—Г—Б–Њ–Љ –≥—А–Є–њ–њ–∞ –Р¬†[59]. –≠—В–Њ –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В —В–µ–Ј–Є—Б –Њ¬†—В–Њ–Љ, —З—В–Њ —Б—В–Є–Љ—Г–ї–Є—А—Г—О—Й–Є–µ –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Г—О –Ј–∞—Й–Є—В—Г —Б–Є–≥–љ–∞–ї—Л, –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ –≤¬†–љ–Є–ґ–љ–Є—Е –Њ—В–і–µ–ї–∞—Е –Ц–Ъ–Ґ, –њ–µ—А–µ–і–∞—О—В—Б—П –≤¬†—Б–ї–Є–Ј–Є—Б—В—Л–µ –і—А—Г–≥–Є—Е –±–Є–Њ—В–Њ–њ–Њ–≤, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –≤¬†—А–µ—Б–њ–Є—А–∞—В–Њ—А–љ—Л–є —В—А–∞–Ї—В.
–Т–Њ-–≤—В–Њ—А—Л—Е, —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–∞ —А–Њ–ї—М NOD-–њ–Њ–і–Њ–±–љ—Л—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ (NLR) –Є¬†–≤–Ї–ї—О—З–∞—О—Й–Є—Е –Є—Е –Є–љ—Д–ї–∞–Љ–Љ–∞—Б–Њ–Љ –Ї–∞–Ї —Б–µ–љ—Б–Њ—А–Њ–≤ –±–∞–Ї—В–µ—А–Є–∞–ї—М–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–Њ–≤, –∞–Ї—В–Є–≤–Є—А—Г—О—Й–Є—Е –њ—А–Њ—В–Є–≤–Њ–≤–Є—А—Г—Б–љ—Л–є –Њ—В–≤–µ—В [60]. –Я—А–Њ–і—Г–Ї—В—Л –і–µ–≥—А–∞–і–∞—Ж–Є–Є –Ї–ї–µ—В–Њ—З–љ—Л—Е —Б—В–µ–љ–Њ–Ї –±–∞–Ї—В–µ—А–Є–є-–Ї–Њ–Љ–Љ–µ–љ—Б–∞–ї–Њ–≤, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –Љ—Г—А–∞–Љ–Є–ї–њ–µ–њ—В–Є–і—Л, —П–≤–ї—П—П—Б—М –ї–Є–≥–∞–љ–і–∞–Љ–Є —Н—В–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ [61], —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ —Г—Б–Є–ї–Є–≤–∞—О—В –њ—А–Њ—В–Є–≤–Њ–≤–Є—А—Г—Б–љ—Г—О –Ј–∞—Й–Є—В—Г –Њ—А–≥–∞–љ–Є–Ј–Љ–∞, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –Њ—В –≤–Њ–Ј–±—Г–і–Є—В–µ–ї–µ–є –≥—А–Є–њ–њ–∞ [62].
–Я—А–Њ–±–Є–Њ—В–Є–Ї–Є –Ї–∞–Ї –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї—П—В–Њ—А—Л
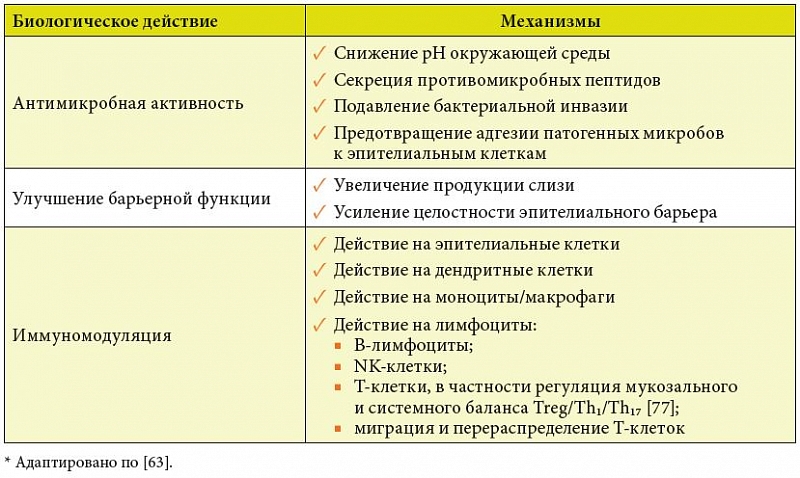
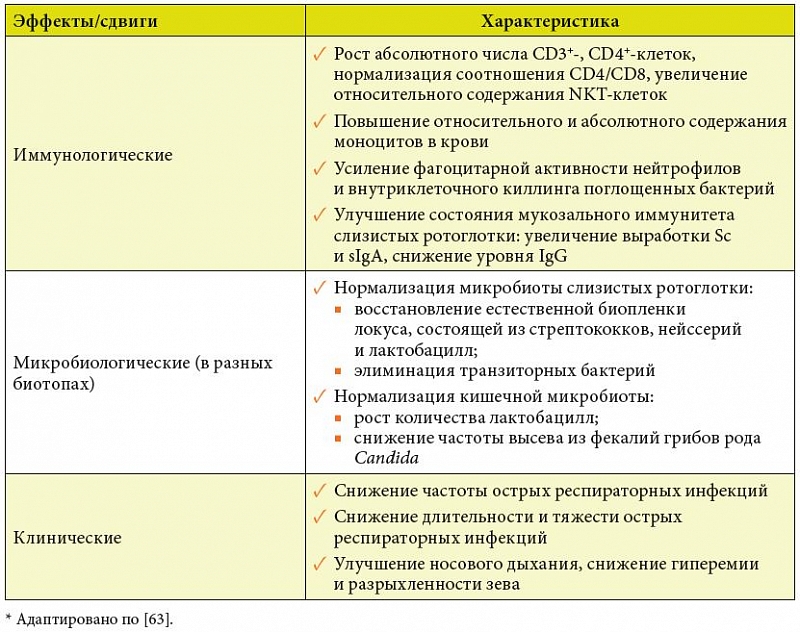
–£–±–µ–і–Є—В–µ–ї—М–љ—Л–µ –і–Њ–Ї–∞–Ј–∞—В–µ–ї—М—Б—В–≤–∞, –≤–Њ-–њ–µ—А–≤—Л—Е, —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–Њ –Ј–∞–Ї—А–µ–њ–ї–µ–љ–љ–Њ–є —А–Њ–ї–Є –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –≤¬†—А–µ–≥—Г–ї—П—Ж–Є–Є –Є–Љ–Љ—Г–љ–љ–Њ–≥–Њ –≥–Њ–Љ–µ–Њ—Б—В–∞–Ј–∞, –≤–Њ-–≤—В–Њ—А—Л—Е, –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–Є –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ–Њ –Є¬†–Ї–∞—З–µ—Б—В–≤–µ–љ–љ–Њ –Ї–Њ—А—А–Є–≥–Є—А–Њ–≤–∞—В—М –Љ–Є–Ї—А–Њ–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Є¬†–Є–Љ–Љ—Г–љ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є –њ—А–Є –њ–µ—А–Њ—А–∞–ї—М–љ–Њ–Љ –≤–≤–µ–і–µ–љ–Є–Є —Б–Є–Љ–±–Є–Њ–љ—В–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є –Њ–Ї–Њ–љ—З–∞—В–µ–ї—М–љ–Њ —В—А–∞–љ—Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–Є –≤–Њ–њ—А–Њ—Б –Њ¬†—Ж–µ–ї–µ—Б–Њ–Њ–±—А–∞–Ј–љ–Њ—Б—В–Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ –≤¬†–Ї–∞—З–µ—Б—В–≤–µ –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї—П—В–Њ—А–Њ–≤ –Є–Ј –і–Є—Б–Ї—Г—Б—Б–Є–Њ–љ–љ–Њ–≥–Њ –≤¬†—А–Є—В–Њ—А–Є—З–µ—Б–Ї–Є–є [1]. –Ф–∞–љ–љ—Л–µ –њ–Њ –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є –Є¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ–Њ –њ–Њ–і—В–≤–µ—А–ґ–і–µ–љ–љ—Л–Љ –Љ–µ—Е–∞–љ–Є–Ј–Љ–∞–Љ –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ–Њ–≥–Њ –і–µ–є—Б—В–≤–Є—П –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ —Б—Г–Љ–Љ–Є—А–Њ–≤–∞–љ—Л –≤¬†—В–∞–±–ї–Є—Ж–µ¬†2 [63].
–Т –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–є —Б—В–µ–њ–µ–љ–Є —Б¬†–Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–µ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В—М—О —Б–≤—П–Ј–∞–љ—Л —Б–ї–µ–і—Г—О—Й–Є–µ –і–Њ–Ї–∞–Ј–∞–љ–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –±–∞–Ї—В–µ—А–Є–є –љ–∞ –Њ—А–≥–∞–љ–Є–Ј–Љ —З–µ–ї–Њ–≤–µ–Ї–∞ [64]:
1) –њ—А–µ–і–Њ—В–≤—А–∞—Й–µ–љ–Є–µ –Є/–Є–ї–Є —Б–љ–Є–ґ–µ–љ–Є–µ –≤—Л—А–∞–ґ–µ–љ–љ–Њ—Б—В–Є –Є–љ—Д–µ–Ї—Ж–Є–є —А–µ—Б–њ–Є—А–∞—В–Њ—А–љ–Њ–≥–Њ —В—А–∞–Ї—В–∞ –Є¬†–Љ–Њ—З–µ–њ–Њ–ї–Њ–≤–Њ–є —Б–Є—Б—В–µ–Љ—Л;
2) –њ—А–µ–і–Њ—В–≤—А–∞—Й–µ–љ–Є–µ –Є/–Є–ї–Є —Б–љ–Є–ґ–µ–љ–Є–µ –≤—Л—А–∞–ґ–µ–љ–љ–Њ—Б—В–Є –∞–ї–ї–µ—А–≥–Є—З–µ—Б–Ї–Є—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є —Г¬†–і–µ—В–µ–є;
3) —Б–љ–Є–ґ–µ–љ–Є–µ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –Ї–∞–љ—Ж–µ—А–Њ–≥–µ–љ–љ—Л—Е —Н–љ–Ј–Є–Љ–Њ–≤ –Є/–Є–ї–Є –≥–љ–Є–ї–Њ—Б—В–љ—Л—Е (–±–∞–Ї—В–µ—А–Є–∞–ї—М–љ—Л—Е) –Љ–µ—В–∞–±–Њ–ї–Є—В–Њ–≤ –≤¬†–Ї–Є—И–µ—З–љ–Є–Ї–µ;
4) –њ—А–µ–і–Њ—В–≤—А–∞—Й–µ–љ–Є–µ –Є–ї–Є —Б–љ–Є–ґ–µ–љ–Є–µ —В—П–ґ–µ—Б—В–Є –Є¬†–њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В–Є —А–Њ—В–∞–≤–Є—А—Г—Б–љ–Њ–є –Є–љ—Д–µ–Ї—Ж–Є–Є –Є¬†–∞–љ—В–Є–±–Є–Њ—В–Є–Ї-–∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ–љ–Њ–є –і–Є–∞—А–µ–Є;
5) –Ї–Њ—А—А–µ–Ї—Ж–Є—П —А–∞—Б—Б—В—А–Њ–є—Б—В–≤ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Є¬†—Б–љ–Є–ґ–µ–љ–Є–µ –≤—Л—А–∞–ґ–µ–љ–љ–Њ—Б—В–Є –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є—Е –њ—А–Њ—П–≤–ї–µ–љ–Є–є –њ—А–Є –≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л—Е –ґ–µ–ї—Г–і–Њ—З–љ–Њ-–Ї–Є—И–µ—З–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П—Е, –Є–Ј–±—Л—В–Њ—З–љ–Њ–Љ –±–∞–Ї—В–µ—А–Є–∞–ї—М–љ–Њ–Љ —А–Њ—Б—В–µ, –∞¬†—В–∞–Ї–ґ–µ –њ—А–Є –Є–љ—Д–µ–Ї—Ж–Є–Є, –≤—Л–Ј–≤–∞–љ–љ–Њ–є Helicobacter pylori;
6) –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–∞ –њ–Њ—Б–ї–µ–Њ–њ–µ—А–∞—Ж–Є–Њ–љ–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Л—Е –Њ—Б–ї–Њ–ґ–љ–µ–љ–Є–є —Г¬†–±–Њ–ї—М–љ—Л—Е –Ї–Њ–ї–Њ—А–µ–Ї—В–∞–ї—М–љ—Л–Љ [65вАУ67] –Є¬†–±–Є–ї–Є–∞—А–љ—Л–Љ —А–∞–Ї–Њ–Љ [68вАУ71].
–Я–Њ–ї—Г—З–µ–љ—Л —В–∞–Ї–ґ–µ –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ—Л–µ –і–∞–љ–љ—Л–µ –њ–Њ –њ—А–Є–Љ–µ–љ–µ–љ–Є—О –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ –њ—А–Є –љ–µ–Ї–Њ—В–Њ—А—Л—Е –∞—Г—В–Њ–Є–Љ–Љ—Г–љ–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П—Е [72].
–°–ї–µ–і—Г–µ—В –≤—Л–і–µ–ї–Є—В—М —А—П–і —Д–µ–љ–Њ–Љ–µ–љ–Њ–≤ –Є¬†—Б–≤–Њ–є—Б—В–≤ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –Ї–Њ—В–Њ—А—Л–µ –Њ—В–ї–Є—З–∞—О—В –Є—Е –Њ—В –і—А—Г–≥–Є—Е –Є–Љ–Љ—Г–љ–Њ—Б—В–Є–Љ—Г–ї–Є—А—Г—О—Й–Є—Е –Є¬†–Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–Є—Е —Б—А–µ–і—Б—В–≤.
–Т–Њ-–њ–µ—А–≤—Л—Е, —Н—В–Њ —Б—В–Њ–є–Ї–Њ—Б—В—М –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ–Њ–≥–Њ –і–µ–є—Б—В–≤–Є—П. –Ъ–Њ—А—А–µ–Ї—Ж–Є—П –Є¬†—Б—В–∞–±–Є–ї–Є–Ј–∞—Ж–Є—П –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ—Л—Е –Є¬†–Ї–∞—З–µ—Б—В–≤–µ–љ–љ—Л—Е –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–µ–є –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л, –і–Њ—Б—В–Є–≥–љ—Г—В—Л–µ –≤¬†—А–µ–Ј—Г–ї—М—В–∞—В–µ –Ї—Г—А—Б–Њ–≤–Њ–≥–Њ –њ—А–Є–Љ–µ–љ–µ–љ–Є—П –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, —П–≤–ї—П—О—В—Б—П —Д—Г–љ–і–∞–Љ–µ–љ—В–Њ–Љ –Є—Е –і–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ –≤–ї–Є—П–љ–Є—П –љ–∞ –Є–Љ–Љ—Г–љ–љ—Л–є –≥–Њ–Љ–µ–Њ—Б—В–∞–Ј. –С–Њ–ї—М—И–Є–љ—Б—В–≤–Њ –і—А—Г–≥–Є—Е –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Њ—Б—В–∞–≤–ї—П—О—В –ї–Є—И—М –Ї—А–∞—В–Ї–Њ–≤—А–µ–Љ–µ–љ–љ—Л–є —Б–ї–µ–і –њ–Њ—Б–ї–µ –Ј–∞–≤–µ—А—И–µ–љ–Є—П –Є—Е –њ—А–Є–Љ–µ–љ–µ–љ–Є—П.
–Т–Њ-–≤—В–Њ—А—Л—Е, –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є–µ –±–∞–Ї—В–µ—А–Є–Є –њ–µ—А–µ–і–∞—О—В –љ–µ —В–Њ–ї—М–Ї–Њ —Б–Є–≥–љ–∞–ї—Л, –∞–Ї—В–Є–≤–Є—А—Г—О—Й–Є–µ –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Г—О –Ј–∞—Й–Є—В—Г, –љ–Њ –Є¬†—Б–Є–≥–љ–∞–ї—Л —В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В–Є –Ї¬†—Б–Њ–±—Б—В–≤–µ–љ–љ—Л–Љ –Є¬†—З—Г–ґ–µ—А–Њ–і–љ—Л–Љ –∞–љ—В–Є–≥–µ–љ–∞–Љ, —В–µ–Љ —Б–∞–Љ—Л–Љ —Б–љ–Є–ґ–∞—П –≤–µ—А–Њ—П—В–љ–Њ—Б—В—М —А–∞–Ј–≤–Є—В–Є—П –∞–ї–ї–µ—А–≥–Є—З–µ—Б–Ї–Є—Е –Є¬†–∞—Г—В–Њ–Є–Љ–Љ—Г–љ–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є. –С–ї–∞–≥–Њ–і–∞—А—П —Н—В–Њ–Љ—Г —Б–≤–Њ–є—Б—В–≤—Г –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ –њ—А–Є –Є—Е –њ—А–Є–Љ–µ–љ–µ–љ–Є–Є –Љ–Є–љ–Є–Љ–Є–Ј–Є—А–Њ–≤–∞–љ —А–Є—Б–Ї –Є–Ј–±—Л—В–Њ—З–љ–Њ–є —Б—В–Є–Љ—Г–ї—П—Ж–Є–Є –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞, –Ї–Њ—В–Њ—А—Л–є —Б—Г—Й–µ—Б—В–≤—Г–µ—В –њ—А–Є –њ—А–Є–Љ–µ–љ–µ–љ–Є–Є –Љ–љ–Њ–≥–Є—Е –Є–љ—Л—Е –Є–Љ–Љ—Г–љ–Њ—Б—В–Є–Љ—Г–ї–Є—А—Г—О—Й–Є—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤. –Т¬†–њ–Њ–і—В–≤–µ—А–ґ–і–µ–љ–Є–µ —В–Њ–ї–µ—А–Њ–≥–µ–љ–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л, –∞¬†—В–∞–Ї–ґ–µ –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤ –Љ–Њ–ґ–љ–Њ –њ—А–Є–≤–µ—Б—В–Є –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —Д–∞–Ї—В–Њ–≤. –Я—А–Є–Љ–µ–љ–µ–љ–Є–µ –∞–љ—В–Є–±–∞–Ї—В–µ—А–Є–∞–ї—М–љ—Л—Е —Б—А–µ–і—Б—В–≤ –Є¬†–Њ—В—Б—Г—В—Б—В–≤–Є–µ/–љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ—Б—В—М –≥—А—Г–і–љ–Њ–≥–Њ –≤—Б–Ї–∞—А–Љ–ї–Є–≤–∞–љ–Є—П –≤¬†–Љ–ї–∞–і–µ–љ—З–µ—Б—В–≤–µ, –≤—Л–Ј—Л–≤–∞—П –і–Є—Б–±–Є–Њ—В–Є—З–µ—Б–Ї–Є–µ —А–∞—Б—Б—В—А–Њ–є—Б—В–≤–∞, —Г–≤–µ–ї–Є—З–Є–≤–∞—О—В –≤–µ—А–Њ—П—В–љ–Њ—Б—В—М —А–∞–Ј–≤–Є—В–Є—П –њ–Є—Й–µ–≤–Њ–є –∞–ї–ї–µ—А–≥–Є–Є –Є¬†–∞—В–Њ–њ–Є—З–µ—Б–Ї–Њ–≥–Њ –і–µ—А–Љ–∞—В–Є—В–∞ [73]. –Э–∞–Њ–±–Њ—А–Њ—В, –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ –≤¬†—А–∞–љ–љ–µ–Љ –≤–Њ–Ј—А–∞—Б—В–µ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є —Б–Њ–і–µ—А–ґ–∞—Й–Є—Е –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї—Л –Є¬†–±–Є—Д–Є–і–Њ–±–∞–Ї—В–µ—А–Є–Є, —Б–љ–Є–ґ–∞–µ—В –≤–µ—А–Њ—П—В–љ–Њ—Б—В—М —А–∞–Ј–≤–Є—В–Є—П –∞–ї–ї–µ—А–≥–Є—З–µ—Б–Ї–Є—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є [74]. –¶–µ–ї—Л–є —А—П–і –∞—Г—В–Њ–Є–Љ–Љ—Г–љ–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є —Б–≤—П–Ј–∞–љ —Б¬†–Ї–Є—И–µ—З–љ—Л–Љ –і–Є—Б–±–Є–Њ–Ј–Њ–Љ [75], —В–Њ–≥–і–∞ –Ї–∞–Ї –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є —Г–Љ–µ–љ—М—И–∞—О—В –≤—Л—А–∞–ґ–µ–љ–љ–Њ—Б—В—М –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є—Е –њ—А–Њ—П–≤–ї–µ–љ–Є–є —А–µ–≤–Љ–∞—В–Њ–Є–і–љ–Њ–≥–Њ –∞—А—В—А–Є—В–∞ [72]. –†–∞—Б–Ї—А—Л—В–Є–µ –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л—Е –Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤ –±–Њ–ї–µ–Ј–љ–Є –Ъ—А–Њ–љ–∞, –њ—А–Є –Ї–Њ—В–Њ—А–Њ–є –≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є–є –і–µ—Д–µ–Ї—В –њ–µ—А–µ–і–∞—З–Є –њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л—Е —Б–Є–≥–љ–∞–ї–Њ–≤ –Њ—В –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–Њ–≤ –±–∞–Ї—В–µ—А–Є–∞–ї—М–љ—Л—Е –Ї–ї–µ—В–Њ—З–љ—Л—Е —Б—В–µ–љ–Њ–Ї¬†вАУ –Љ—Г—А–∞–Љ–Є–ї–њ–µ–њ—В–Є–і–Њ–≤¬†вАУ —З–µ—А–µ–Ј NOD2-—А–µ—Ж–µ–њ—В–Њ—А—Л –њ—А–Є–≤–Њ–і–Є—В –Ї¬†—А–∞–Ј–≤–Є—В–Є—О —В—П–ґ–µ–ї–Њ–≥–Њ –≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П –Ї–Є—И–µ—З–љ–Є–Ї–∞, —В–∞–Ї–ґ–µ –≤¬†–Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–є —Б—В–µ–њ–µ–љ–Є –њ–Њ–і–і–µ—А–ґ–Є–≤–∞–µ—В —В–µ–Ј–Є—Б –Њ¬†—В–Њ–ї–µ—А–Њ–≥–µ–љ–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є —Б–Є–Љ–±–Є–Њ–љ—В–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є [61].
–Т-—В—А–µ—В—М–Є—Е, —Б—Г—Й–µ—Б—В–≤—Г—О—В —Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–Њ –Ј–∞–Ї—А–µ–њ–ї–µ–љ–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –њ–µ—А–µ–і–∞—З–Є —Б–Є–≥–љ–∞–ї–Њ–≤, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—Й–Є—Е –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Г—О –Ј–∞—Й–Є—В—Г –Є¬†–Є–Љ–Љ—Г–љ–љ—Г—О —В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В—М, –Є–Ј –Ц–Ъ–Ґ –≤¬†–і—А—Г–≥–Є–µ –±–Є–Њ—В–Њ–њ—Л –Њ—А–≥–∞–љ–Є–Ј–Љ–∞, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –≤¬†—Б–ї–Є–Ј–Є—Б—В—Л–µ —А–µ—Б–њ–Є—А–∞—В–Њ—А–љ–Њ–≥–Њ —В—А–∞–Ї—В–∞ –Є¬†–Љ–Њ—З–µ–њ–Њ–ї–Њ–≤–Њ–є —Б–Є—Б—В–µ–Љ—Л. –Ю—Б–љ–Њ–≤–љ—Л–µ –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л —Н—В–Њ–≥–Њ —Д–µ–љ–Њ–Љ–µ–љ–∞, –њ–Њ–ї—Г—З–Є–≤—И–µ–≥–Њ –љ–∞–Ј–≤–∞–љ–Є–µ ¬Ђ–Є–Љ–Љ—Г–љ–љ–∞—П —Б–Њ–ї–Є–і–∞—А–љ–Њ—Б—В—М —Б–ї–Є–Ј–Є—Б—В—Л—Е¬ї [76], —А–∞—Б–Ї—А—Л—В—Л. –Я—А–Є–Љ–µ–љ—П—П –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є, –Љ—Л –Є–Љ–Є—В–Є—А—Г–µ–Љ –µ—Б—В–µ—Б—В–≤–µ–љ–љ—Л–є –њ—Г—В—М —А–µ–≥—Г–ї—П—Ж–Є–Є –Є–Љ–Љ—Г–љ–љ—Л—Е —А–µ–∞–Ї—Ж–Є–є –≤–љ–µ –њ–Є—Й–µ–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л, –Ї–Њ—В–Њ—А—Л–є –Њ–±—Г—Б–ї–Њ–≤–ї–µ–љ —Ж–Є—А–Ї—Г–ї—П—Ж–Є–µ–є –±–∞–Ї—В–µ—А–Є–∞–ї—М–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–Њ–≤ –Є¬†–њ—А–Њ–і—Г–Ї—В–Њ–≤, –∞¬†—В–∞–Ї–ґ–µ –Є–Љ–Љ—Г–љ–љ—Л—Е –Љ–µ–і–Є–∞—В–Њ—А–Њ–≤ –Є¬†–Ї–ї–µ—В–Њ–Ї –Є–Ј –Ц–Ъ–Ґ –≤¬†–і—А—Г–≥–Є–µ –Њ—А–≥–∞–љ—Л –Є¬†—В–Ї–∞–љ–Є –љ–∞—И–µ–≥–Њ –Њ—А–≥–∞–љ–Є–Ј–Љ–∞.
–†–∞–Ј–ї–Є—З–Є—П –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –±–∞–Ї—В–µ—А–Є–є –≤¬†–Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є
–Т—Л—П–≤–ї–µ–љ—Л —Б—Г—Й–µ—Б—В–≤–µ–љ–љ—Л–µ –Љ–µ–ґ—А–Њ–і–Њ–≤—Л–µ, –Љ–µ–ґ–≤–Є–і–Њ–≤—Л–µ –Є¬†–Љ–µ–ґ—И—В–∞–Љ–Љ–Њ–≤—Л–µ —А–∞–Ј–ї–Є—З–Є—П –≤¬†–Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–µ–Љ –і–µ–є—Б—В–≤–Є–Є —Б–Є–Љ–±–Є–Њ–љ—В–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є. –Я—А–Є—З–µ–Љ —Н—В–Є —А–∞–Ј–ї–Є—З–Є—П –Ї–∞—Б–∞—О—В—Б—П –љ–µ —В–Њ–ї—М–Ї–Њ –≤—Л—А–∞–ґ–µ–љ–љ–Њ—Б—В–Є, –љ–Њ –Є¬†—Е–∞—А–∞–Ї—В–µ—А–∞ –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤.
–Т –Ї–∞—З–µ—Б—В–≤–µ –њ—А–Є–Љ–µ—А–∞ –њ—А–Є–≤–µ–і–µ–Љ –і–∞–љ–љ—Л–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–µ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є 21¬†—И—В–∞–Љ–Љ–∞ —З–µ—В—Л—А–µ—Е –≤–Є–і–Њ–≤ –±–Є—Д–Є–і–Њ–±–∞–Ї—В–µ—А–Є–є [77]. –Я–µ—А–≤–∞—П –≥—А—Г–њ–њ–∞ —Н—В–Є—Е —И—В–∞–Љ–Љ–Њ–≤ –≤¬†–Ї—Г–ї—М—В—Г—А–µ –Љ–Њ–љ–Њ–љ—Г–Ї–ї–µ–∞—А–љ—Л—Е –Ї–ї–µ—В–Њ–Ї —З–µ–ї–Њ–≤–µ–Ї–∞ in vitro –Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–ї–∞ –њ—А–Њ–і—Г–Ї—Ж–Є—О —Ж–Є—В–Њ–Ї–Є–љ–Њ–≤ ThвВБ-–њ—А–Њ—Д–Є–ї—П: –Ш–§–Э-–≥–∞–Љ–Љ–∞ –Є¬†—Д–∞–Ї—В–Њ—А–∞ –љ–µ–Ї—А–Њ–Ј–∞ –Њ–њ—Г—Е–Њ–ї–Є (–§–Э–Ю). –Т—В–Њ—А–∞—П –≥—А—Г–њ–њ–∞ –≥–ї–∞–≤–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ –≤—Л–Ј—Л–≤–∞–ї–∞ –≤—Л—А–∞–±–Њ—В–Ї—Г –Є–љ—В–µ—А–ї–µ–є–Ї–Є–љ–∞-17 –њ—А–Є –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –љ–Є–Ј–Ї–Њ–є –њ—А–Њ–і—Г–Ї—Ж–Є–Є –Ш–§–Э-–≥–∞–Љ–Љ–∞ –Є¬†–§–Э–Ю (ThвВБвВЗ-–њ—А–Њ—Д–Є–ї—М). –Ґ—А–µ—В—М—П –≥—А—Г–њ–њ–∞ –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ —Б—В–Є–Љ—Г–ї–Є—А–Њ–≤–∞–ї–∞ –≤—Л—А–∞–±–Њ—В–Ї—Г –Є–љ—В–µ—А–ї–µ–є–Ї–Є–љ–∞-10 –њ—А–Є –љ–Є–Ј–Ї–Њ–Љ —Г—А–Њ–≤–љ–µ –њ—А–Њ–і—Г–Ї—Ж–Є–Є ThвВБ- –Є¬†ThвВБвВЗ-—Ж–Є—В–Њ–Ї–Є–љ–Њ–≤ (Treg-–њ—А–Њ—Д–Є–ї—М). –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, —А–∞–Ј–љ—Л–µ —И—В–∞–Љ–Љ—Л –±–Є—Д–Є–і–Њ–±–∞–Ї—В–µ—А–Є–є –Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–ї–Є –ї–Є–±–Њ –њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л–µ —Ж–Є—В–Њ–Ї–Є–љ—Л, –ї–Є–±–Њ –њ—А–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л–µ –Љ–µ–і–Є–∞—В–Њ—А—Л, –њ—А–Є —Н—В–Њ–Љ —Б–њ–µ–Ї—В—А –њ–Њ—Б–ї–µ–і–љ–Є—Е —В–Њ–ґ–µ –≤–∞—А—М–Є—А–Њ–≤–∞–ї. –С–Њ–ї–µ–µ —В–Њ–≥–Њ, –Є–љ–Ї—Г–±–∞—Ж–Є—П —Б¬†—А–∞–Ј–ї–Є—З–љ—Л–Љ–Є –±–∞–Ї—В–µ—А–Є–∞–ї—М–љ—Л–Љ–Є —И—В–∞–Љ–Љ–∞–Љ–Є –њ–Њ-—А–∞–Ј–љ–Њ–Љ—Г –Є–Ј–Љ–µ–љ—П–ї–∞ —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –і–µ–љ–і—А–Є—В–љ—Л—Е –Ї–ї–µ—В–Њ–Ї –њ–Њ–ї—П—А–Є–Ј–Њ–≤–∞—В—М –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–Ї—Г –љ–µ–Ј—А–µ–ї—Л—Е T-–Ї–ї–µ—В–Њ–Ї, –Њ–њ—А–µ–і–µ–ї—П—П –±–∞–ї–∞–љ—Б —Б—Г–±–њ–Њ–њ—Г–ї—П—Ж–Є–є CD4+-–Ґ-–Ї–ї–µ—В–Њ–Ї: ThвВБ/ThвВБвВЗ/Treg.
–Э–∞–њ–Њ–Љ–љ–Є–Љ, —З—В–Њ ThвВБ-–Ї–ї–µ—В–Ї–Є –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л –і–ї—П –Є–Љ–Љ—Г–љ–љ–Њ–≥–Њ –Њ—В–≤–µ—В–∞ –њ—А–Њ—В–Є–≤ –≤–Є—А—Г—Б–Њ–≤ –Є¬†–±–∞–Ї—В–µ—А–Є–є —Б¬†–≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ—Л–Љ –њ–∞—А–∞–Ј–Є—В–Є—А–Њ–≤–∞–љ–Є–µ–Љ (–Љ–Є–Ї–Њ–њ–ї–∞–Ј–Љ—Л, —Е–ї–∞–Љ–Є–і–Є–Є, —Г—А–µ–∞–њ–ї–∞–Ј–Љ—Л –Є¬†–і—А.), ThвВБвВЗ¬†вАУ –њ—А–Њ—В–Є–≤ –≤–љ–µ–Ї–ї–µ—В–Њ—З–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є –Є¬†–≥—А–Є–±–Њ–≤, —В–Њ–≥–і–∞ –Ї–∞–Ї Treg –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—В –Є–Љ–Љ—Г–љ–љ—Г—О —В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В—М, —В–Њ –µ—Б—В—М –њ—А–µ–і–Њ—В–≤—А–∞—Й–∞—О—В –Є–Ј–±—Л—В–Њ—З–љ—Л–µ –Є–Љ–Љ—Г–љ–љ—Л–µ —А–µ–∞–Ї—Ж–Є–Є –≤¬†–Њ—В–љ–Њ—И–µ–љ–Є–Є —Б–Њ–±—Б—В–≤–µ–љ–љ—Л—Е –Є¬†—З—Г–ґ–µ—А–Њ–і–љ—Л—Е —Б—Г–±—Б—В–∞–љ—Ж–Є–є.
–Т –і—А—Г–≥–Њ–є —А–∞–±–Њ—В–µ –њ—А–Њ–і–µ–Љ–Њ–љ—Б—В—А–Є—А–Њ–≤–∞–љ—Л —А–∞–Ј–ї–Є—З–Є—П –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–Є—Е —Н—Д—Д–µ–Ї—В–Њ–≤ 4¬†—И—В–∞–Љ–Љ–Њ–≤ –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї –Є¬†2¬†—И—В–∞–Љ–Љ–Њ–≤ –±–Є—Д–Є–і–Њ–±–∞–Ї—В–µ—А–Є–є (Bifidobacterium longum SP 07/3 –Є¬†Bifidobacterium bifidum MF 20/5) [78]. –Т—Б–µ –±–∞–Ї—В–µ—А–Є–Є —Г–≤–µ–ї–Є—З–Є–≤–∞–ї–Є —Н–Ї—Б–њ—А–µ—Б—Б–Є—О –∞–Ї—В–Є–≤–∞—Ж–Є–Њ–љ–љ—Л—Е –Љ–∞—А–Ї–µ—А–Њ–≤ –љ–∞ –Ґ-–Ї–ї–µ—В–Ї–∞—Е –Є¬†–µ—Б—В–µ—Б—В–≤–µ–љ–љ—Л—Е –Ї–Є–ї–ї–µ—А–∞—Е (NK), –∞¬†—В–∞–Ї–ґ–µ —Г—Б–Є–ї–Є–≤–∞–ї–Є —Ж–Є—В–Њ—В–Њ–Ї—Б–Є—З–µ—Б–Ї—Г—О –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –њ–Њ—Б–ї–µ–і–љ–Є—Е. –Я—А–Є —Н—В–Њ–Љ –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї—Л —Б—В–Є–Љ—Г–ї–Є—А–Њ–≤–∞–ї–Є –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –њ—А–Њ–і—Г–Ї—Ж–Є—О —Ж–Є—В–Њ–Ї–Є–љ–Њ–≤ ThвВБ-–њ—А–Њ—Д–Є–ї—П, –∞¬†—Г–Ї–∞–Ј–∞–љ–љ—Л–µ —И—В–∞–Љ–Љ—Л –±–Є—Д–Є–і–Њ–±–∞–Ї—В–µ—А–Є–є¬†вАУ –њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л—Е –Љ–µ–і–Є–∞—В–Њ—А–Њ–≤. –Я—А–Є–Љ–µ—З–∞—В–µ–ї—М–љ–∞ —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М Bifidobacterium longum SP 07/3 –Є¬†Bifidobacterium bifidum MF 20/5 –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ —Г—Б–Є–ї–Є–≤–∞—В—М –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –Њ—Б–љ–Њ–≤–љ—Л—Е –Ї–ї–µ—В–Њ—З–љ—Л—Е –Ј–≤–µ–љ—М–µ–≤ –Є–Љ–Љ—Г–љ–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л, —Г—З–∞—Б—В–≤—Г—О—Й–Є—Е –≤¬†–њ—А–Њ—В–Є–≤–Њ–≤–Є—А—Г—Б–љ–Њ–Љ –Њ—В–≤–µ—В–µ, –Є¬†–Є–љ–і—Г—Ж–Є—А–Њ–≤–∞—В—М —В–Њ–ї–µ—А–Њ–≥–µ–љ–љ—Л–µ —Б–Є–≥–љ–∞–ї—Л.
–Т—Л—И–µ—Б–Ї–∞–Ј–∞–љ–љ–Њ–µ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г–µ—В –Њ¬†—Ж–µ–ї–µ—Б–Њ–Њ–±—А–∞–Ј–љ–Њ—Б—В–Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –≤¬†–Ї–∞—З–µ—Б—В–≤–µ –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї—П—В–Њ—А–Њ–≤ –ї–Є—И—М —В–µ—Е –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –Ї–Њ—В–Њ—А—Л–µ –≤–Ї–ї—О—З–∞—О—В —И—В–∞–Љ–Љ—Л —Б¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ–Њ –Є–Ј—Г—З–µ–љ–љ—Л–Љ –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є –і–Њ–Ї–∞–Ј–∞–љ–љ—Л–Љ –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ—Л–Љ –і–µ–є—Б—В–≤–Є–µ–Љ. –Я–Њ–і—З–µ—А–Ї–љ–µ–Љ, —З—В–Њ –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –Љ–љ–Њ–≥–Є—Е –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є –Є—Б–њ–Њ–ї—М–Ј—Г–µ–Љ—Л—Е –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–∞ –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ –ї–Є–±–Њ –љ–µ –Є–Ј—Г—З–µ–љ–∞ –≤–Њ–Њ–±—Й–µ.
–Т —Б–ї—Г—З–∞–µ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П —Б–Є–Љ–±–Є–Њ—В–Є–Ї–Њ–≤, —В–Њ –µ—Б—В—М –Ї–Њ–Љ–њ–ї–µ–Ї—Б–Њ–≤ –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤, –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ —Г—З–Є—В—Л–≤–∞—В—М –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М —Б–Є–љ–µ—А–≥–Є–Ј–Љ–∞ –≤¬†—А–µ–∞–ї–Є–Ј–∞—Ж–Є–Є –Њ–і–љ–Є—Е –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –Є¬†–∞–љ—В–∞–≥–Њ–љ–Є–Ј–Љ–∞¬†вАУ –і—А—Г–≥–Є—Е, —З—В–Њ —В—А–µ–±—Г–µ—В –і–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ—Л—Е —Б–њ–µ—Ж–Є–∞–ї—М–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є.
–≠—Д—Д–µ–Ї—В–Є–≤–љ—Л–Љ–Є –њ—А–Є–Ј–љ–∞–љ—Л –Ї–Њ–Љ–±–Є–љ–∞—Ж–Є–Є –љ–µ–Ї–Њ—В–Њ—А—Л—Е –ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї –Є¬†–±–Є—Д–Є–і–Њ–±–∞–Ї—В–µ—А–Є–є. –ѓ—А–Ї–Є–Љ –њ—А–Є–Љ–µ—А–Њ–Љ —П–≤–ї—П–µ—В—Б—П —Б–Њ—З–µ—В–∞–љ–Є–µ –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤ Lactobacillus gasseri PA 16/8, Bifidobacterium bifidum MF 20/5 –Є¬†Bifidobacterium longum SP 07/3. –Ґ–∞–Ї–∞—П –Ї–Њ–Љ–±–Є–љ–∞—Ж–Є—П –њ—А–Є –Ї—Г—А—Б–Њ–≤–Њ–Љ –њ—А–Є–Љ–µ–љ–µ–љ–Є–Є –љ–µ —В–Њ–ї—М–Ї–Њ –Ї–Њ—А—А–Є–≥–Є—А–Њ–≤–∞–ї–∞ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є –Ї–ї–µ—В–Њ—З–љ–Њ–≥–Њ –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞ –Є¬†—Г–≤–µ–ї–Є—З–Є–≤–∞–ї–∞ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ –±–Є—Д–Є–і–Њ–±–∞–Ї—В–µ—А–Є–є –Є¬†–ї–∞–Ї—В–Њ–±–∞—Ж–Є–ї–ї –≤¬†—Д–µ–Ї–∞–ї–Є—П—Е –Є—Б–њ—Л—В—Г–µ–Љ—Л—Е, –љ–Њ –Є¬†–њ—А–µ–і–Њ—В–≤—А–∞—Й–∞–ї–∞ —А–∞–Ј–≤–Є—В–Є–µ –Є¬†—Б–љ–Є–ґ–∞–ї–∞ –≤—Л—А–∞–ґ–µ–љ–љ–Њ—Б—В—М —А–µ—Б–њ–Є—А–∞—В–Њ—А–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–є –≤¬†–Ј–Є–Љ–љ–µ-–≤–µ—Б–µ–љ–љ–Є–є –њ–µ—А–Є–Њ–і [79, 80]. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, —И—В–∞–Љ–Љ—Л Lactobacillus gasseri PA 16/8 –Є¬†Bifidobacterium bifidum MF 20/5 –њ—А–Њ—П–≤–ї—П–ї–Є —Б—Г—Й–µ—Б—В–≤–µ–љ–љ—Л–є —Б–Є–љ–µ—А–≥–Є–Ј–Љ –≤¬†–њ–Њ–і–∞–≤–ї–µ–љ–Є–Є 6¬†—З–∞—Б—В–Њ –≤—Б—В—А–µ—З–∞—О—Й–Є—Е—Б—П –≤¬†–Ц–Ъ–Ґ –њ–∞—В–Њ–≥–µ–љ–Њ–≤ [81].
–Т—Л–±–Њ—А –њ—А–Њ–±–Є–Њ—В–Є–Ї–∞ –і–ї—П —Г–Ї—А–µ–њ–ї–µ–љ–Є—П –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ–Њ–є –Ј–∞—Й–Є—В—Л
–Я—А–Њ–±–Є–Њ—В–Є–Ї–Є —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ —А–∞–Ј–ї–Є—З–∞—О—В—Б—П –љ–µ —В–Њ–ї—М–Ї–Њ –њ–Њ —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В–Є –≤–ї–Є—П—В—М –љ–∞ –Ї–Є—И–µ—З–љ—Г—О –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Г, –љ–Њ –Є¬†–њ–Њ –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є. –Э–∞ —Н—В–Є —А–∞–Ј–ї–Є—З–Є—П –≤–ї–Є—П—О—В –њ—А–µ–ґ–і–µ –≤—Б–µ–≥–Њ —Б–≤–Њ–є—Б—В–≤–∞ –Є—Б–њ–Њ–ї—М–Ј—Г–µ–Љ—Л—Е –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –Љ–Є–Ї—А–Њ–±–Њ–≤, —Б—В–µ–њ–µ–љ—М –Є—Е –Ј–∞—Й–Є—В—Л –Њ—В –∞–≥—А–µ—Б—Б–Є–≤–љ—Л—Е –ґ–µ–ї—Г–і–Њ—З–љ—Л—Е –Є¬†–і—Г–Њ–і–µ–љ–∞–ї—М–љ—Л—Е —Д–∞–Ї—В–Њ—А–Њ–≤, –∞¬†—В–∞–Ї–ґ–µ –љ–∞–ї–Є—З–Є–µ –і–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ—Л—Е –Є–љ–≥—А–µ–і–Є–µ–љ—В–Њ–≤, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—Й–Є—Е —Н—Д—Д–µ–Ї—В–Є–≤–љ—Г—О —А–µ–∞–ї–Є–Ј–∞—Ж–Є—О –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –њ–Њ—В—А–µ–±–ї—П–µ–Љ—Л–Љ–Є —Б–Є–Љ–±–Є–Њ–љ—В–∞–Љ–Є.
–†–∞—Ж–Є–Њ–љ–∞–ї—М–љ—Л–µ –њ—А–Є–љ—Ж–Є–њ—Л –≤—Л–±–Њ—А–∞ –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ–Њ–≥–Њ –њ—А–Њ–±–Є–Њ—В–Є–Ї–∞ –Њ—Б–≤–µ—Й–µ–љ—Л —А–∞–љ–µ–µ [1]. –Ъ¬†—Н—В–Є–Љ –њ—А–Є–љ—Ж–Є–њ–∞–Љ –Љ–Њ–ґ–љ–Њ –і–Њ–±–∞–≤–Є—В—М —Ж–µ–ї–µ—Б–Њ–Њ–±—А–∞–Ј–љ–Њ—Б—В—М –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П —В–Њ–ї—М–Ї–Њ —В–µ—Е –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –Ї–Њ—В–Њ—А—Л–µ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г—О—В —В—А–µ–±–Њ–≤–∞–љ–Є—П–Љ FAO/WHO [18], –Њ—Б–Њ–±–µ–љ–љ–Њ –њ–Њ –Ї—А–Є—В–µ—А–Є—П–Љ –±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В–Є.
–Ъ–Њ–Љ–њ–∞–љ–Є—П Merck KGaA (–У–µ—А–Љ–∞–љ–Є—П) —А–∞–Ј—А–∞–±–Њ—В–∞–ї–∞ –Є¬†–≤—Л–≤–µ–ї–∞ –љ–∞ —А—Л–љ–Њ–Ї –±–µ–Ј–Њ–њ–∞—Б–љ—Л–µ –Є¬†–≤—Л—Б–Њ–Ї–Њ—Н—Д—Д–µ–Ї—В–Є–≤–љ—Л–µ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є –С–Є–Њ–љ¬Ѓ3 (–і–ї—П –≤–Ј—А–Њ—Б–ї—Л—Е –Є¬†–њ–Њ–і—А–Њ—Б—В–Ї–Њ–≤ –Њ—В 14¬†–ї–µ—В) –Є¬†–С–Є–Њ–љ¬Ѓ3¬†–Ъ–Є–і (–і–ї—П –і–µ—В–µ–є –Њ—В¬†4 –і–Њ 14¬†–ї–µ—В), –Њ–±–ї–∞–і–∞—О—Й–Є–µ –і–Њ–Ї–∞–Ј–∞–љ–љ–Њ–є –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В—М—О –Є¬†–њ–Њ–ї–љ–Њ—Б—В—М—О –Њ—В–≤–µ—З–∞—О—Й–Є–µ –Ї—А–Є—В–µ—А–Є—П–Љ FAO/WHO. –≠—В–Є –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є –≤–Ї–ї—О—З–∞—О—В –≤¬†—Б–µ–±—П –Ї–Њ–Љ–±–Є–љ–∞—Ж–Є—О Tribion HarmonisвДҐ –Є–Ј —В—А–µ—Е —В—Й–∞—В–µ–ї—М–љ–Њ –њ–Њ–і–Њ–±—А–∞–љ–љ—Л—Е —И—В–∞–Љ–Љ–Њ–≤ —Б–Є–Љ–±–Є–Њ–љ—В–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є: Lactobacillus gasseri PA 16/8, Bifidobacterium bifidum MF 20/5 –Є¬†Bifidobacterium longum SP 07/3. –Ю–± –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–Є—Е —Б–≤–Њ–є—Б—В–≤–∞—Е —Н—В–Є—Е —И—В–∞–Љ–Љ–Њ–≤ –Є¬†—Б–Є–љ–µ—А–≥–Є–Ј–Љ–µ –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –њ—А–Є –Є—Е —Б–Њ—З–µ—В–∞–љ–љ–Њ–Љ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–Є —Г–ґ–µ –≥–Њ–≤–Њ—А–Є–ї–Њ—Б—М –≤—Л—И–µ.
–Т —Б–Њ—Б—В–∞–≤ –С–Є–Њ–љ¬Ѓ3, –Ї—А–Њ–Љ–µ —В–Њ–≥–Њ, –≤—Е–Њ–і—П—В –њ—А–µ–±–Є–Њ—В–Є–Ї –Є–љ—Г–ї–Є–љ, 12¬†–≤–Є—В–∞–Љ–Є–љ–Њ–≤ –Є¬†12¬†–Љ–∞–Ї—А–Њ- –Є¬†–Љ–Є–Ї—А–Њ—Н–ї–µ–Љ–µ–љ—В–Њ–≤, –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л—Е –і–ї—П –њ–Њ–ї–љ–Њ—Ж–µ–љ–љ–Њ–≥–Њ —Д—Г–љ–Ї—Ж–Є–Њ–љ–Є—А–Њ–≤–∞–љ–Є—П –Є–Љ–Љ—Г–љ–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л. –С–Є–Њ–љ¬Ѓ3¬†–Ъ–Є–і –њ–Њ —Б–Њ—Б—В–∞–≤—Г –Њ—В–ї–Є—З–∞–µ—В—Б—П —В–µ–Љ, —З—В–Њ –Є–Ј –Љ–Є–љ–µ—А–∞–ї—М–љ—Л—Е –≤–µ—Й–µ—Б—В–≤ –≤–Ї–ї—О—З–∞–µ—В —В–Њ–ї—М–Ї–Њ –Ї–∞–ї—М—Ж–Є–є, —Ж–Є–љ–Ї –Є¬†–ґ–µ–ї–µ–Ј–Њ.
–Я–Њ–Љ–Є–Љ–Њ —А–∞—Ж–Є–Њ–љ–∞–ї—М–љ–Њ –њ–Њ–і–Њ–±—А–∞–љ–љ–Њ–є –Ї–Њ–Љ–±–Є–љ–∞—Ж–Є–Є 3¬†—Б–Є–Љ–±–Є–Њ–љ—В–љ—Л—Е —И—В–∞–Љ–Љ–Њ–≤ –Є¬†–Ї–Њ–Љ–њ–ї–µ–Ї—Б–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞, –≤–∞–ґ–љ—Л–Љ —Б–ї–∞–≥–∞–µ–Љ—Л–Љ —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є –С–Є–Њ–љ¬Ѓ3 —П–≤–ї—П–µ—В—Б—П –Ј–∞–њ–∞—В–µ–љ—В–Њ–≤–∞–љ–љ–∞—П —В–µ—Е–љ–Њ–ї–Њ–≥–Є—П —В–∞–±–ї–µ—В–Є—А–Њ–≤–∞–љ–Є—П: –Њ—А–Є–≥–Є–љ–∞–ї—М–љ–∞—П —В—А–µ—Е—Б–ї–Њ–є–љ–∞—П —В–∞–±–ї–µ—В–Ї–∞, –њ–Њ–Ї—А—Л—В–∞—П –Ј–∞—Й–Є—В–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Њ–є, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞–µ—В delayed release (–Ј–∞–Љ–µ–і–ї–µ–љ–љ–Њ–µ –≤—Л—Б–≤–Њ–±–Њ–ґ–і–µ–љ–Є–µ), –∞¬†—В–∞–Ї–ґ–µ –≥–∞—А–∞–љ—В–Є—А—Г–µ—В –і–Њ—Б—В–∞–≤–Ї—Г –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –Ї—Г–ї—М—В—Г—А –≤¬†–љ–Є–ґ–љ–Є–µ –Њ—В–і–µ–ї—Л –Ц–Ъ–Ґ –ґ–Є–≤—Л–Љ–Є –Є¬†–≤—Л—Б–Њ–Ї—Г—О –±–Є–Њ–і–Њ—Б—В—Г–њ–љ–Њ—Б—В—М –і—А—Г–≥–Є—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–Њ–≤ –њ—А–µ–њ–∞—А–∞—В–∞.
–С–ї–∞–≥–Њ–і–∞—А—П —Г–Ї–∞–Ј–∞–љ–љ—Л–Љ —Б–≤–Њ–є—Б—В–≤–∞–Љ –С–Є–Њ–љ¬Ѓ3 —Е–Њ—А–Њ—И–Њ –Ј–∞—А–µ–Ї–Њ–Љ–µ–љ–і–Њ–≤–∞–ї —Б–µ–±—П –љ–µ —В–Њ–ї—М–Ї–Њ –Ї–∞–Ї —Б—А–µ–і—Б—В–≤–Њ –і–ї—П –њ–Њ–і–і–µ—А–ґ–∞–љ–Є—П –Ј–і–Њ—А–Њ–≤–Њ–≥–Њ –±–∞–ї–∞–љ—Б–∞ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Ц–Ъ–Ґ, –љ–Њ –Є¬†–Ї–∞–Ї –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї—П—В–Њ—А —Б¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є –Є¬†–ї–∞–±–Њ—А–∞—В–Њ—А–љ–Њ –і–Њ–Ї–∞–Ј–∞–љ–љ–Њ–є —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М—О. –Я–Њ—Б—В–µ–њ–µ–љ–љ–Њ –љ–∞–Ї–∞–њ–ї–Є–≤–∞–µ—В—Б—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П –Њ–± –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–µ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є—Е —Н—Д—Д–µ–Ї—В–∞—Е –С–Є–Њ–љ¬Ѓ3¬†–Ъ–Є–і.
–І–∞—Б—В—Л–µ —Н–њ–Є–Ј–Њ–і—Л –Њ—Б—В—А—Л—Е —А–µ—Б–њ–Є—А–∞—В–Њ—А–љ—Л—Е –≤–Є—А—Г—Б–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–є (–Ю–†–Т–Ш) —П–≤–ї—П—О—В—Б—П –≤–∞–ґ–љ–µ–є—И–Є–Љ –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–Љ –Љ–∞—А–Ї–µ—А–Њ–Љ –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ—Б—В–Є –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ–Њ–є –Ј–∞—Й–Є—В—Л. –С–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В—М –Є¬†—Б—В–Њ–є–Ї–Њ—Б—В—М –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤, –Ї–Њ—В–Њ—А—Л–Љ–Є —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—В—Б—П –љ–µ–Ї–Њ—В–Њ—А—Л–µ —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л–µ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є, —П–≤–ї—П—О—В—Б—П –≤–∞–ґ–љ–µ–є—И–Є–Љ–Є –Ї–∞—З–µ—Б—В–≤–∞–Љ–Є –њ—А–Є –≤—Л–±–Њ—А–µ —Б—А–µ–і—Б—В–≤–∞ –љ–µ—Б–њ–µ—Ж–Є—Д–Є—З–µ—Б–Ї–Њ–є —Б–µ–Ј–Њ–љ–љ–Њ–є –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є –Ю–†–Т–Ш.
–Т —Н—В–Њ–є —Б–≤—П–Ј–Є –Є–љ—В–µ—А–µ—Б –њ—А–µ–і—Б—В–∞–≤–ї—П—О—В —А–µ–Ј—Г–ї—М—В–∞—В—Л –і–≤—Г—Е –њ—А–Њ–≤–µ–і–µ–љ–љ—Л—Е –≤¬†–У–µ—А–Љ–∞–љ–Є–Є –Ї—А—Г–њ–љ—Л—Е –і–≤–Њ–є–љ—Л—Е —Б–ї–µ–њ—Л—Е —А–∞–љ–і–Њ–Љ–Є–Ј–Є—А–Њ–≤–∞–љ–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є –С–Є–Њ–љ¬Ѓ3 –Ї–∞–Ї —Б—А–µ–і—Б—В–≤–∞ –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є –Ю–†–Т–Ш.
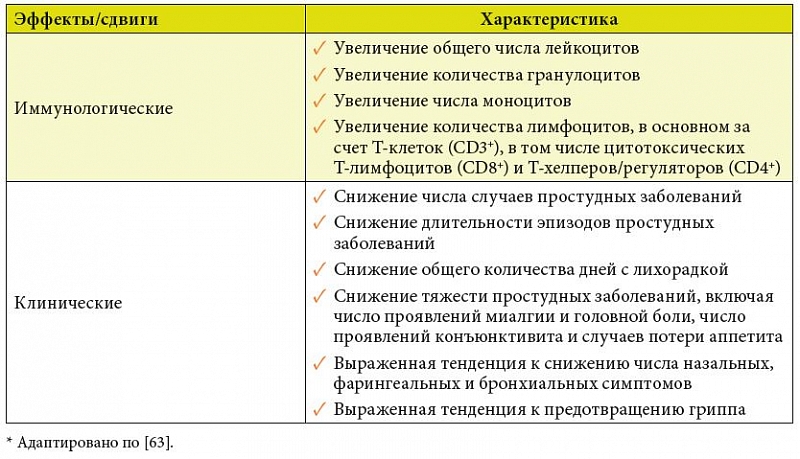
–Т –Њ–і–љ–Њ–Љ –Є–Ј –љ–Є—Е (n = 477) —Г¬†–і–Њ–±—А–Њ–≤–Њ–ї—М—Ж–µ–≤, –њ—А–Є–љ–Є–Љ–∞–≤—И–Є—Е –µ–ґ–µ–і–љ–µ–≤–љ–Њ –њ–Њ 1¬†—В–∞–±–ї–µ—В–Ї–µ –С–Є–Њ–љ¬Ѓ3 –≤¬†–Ј–Є–Љ–љ–Є–є –Є–ї–Є –Ј–Є–Љ–љ–µ-–≤–µ—Б–µ–љ–љ–Є–є –њ–µ—А–Є–Њ–і, –≤¬†—Б—А–∞–≤–љ–µ–љ–Є–Є —Б¬†—В–∞–Ї–Њ–≤—Л–Љ–Є, –њ–Њ–ї—Г—З–∞–≤—И–Є–Љ–Є –њ–ї–∞—Ж–µ–±–Њ, —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ —Б–љ–Є–ґ–∞–ї–Њ—Б—М —З–Є—Б–ї–Њ —Б–ї—Г—З–∞–µ–≤ –њ—А–Њ—Б—В—Г–і–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є, –Њ–±—Й–µ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –і–љ–µ–є —Б¬†–ї–Є—Е–Њ—А–∞–і–Ї–Њ–є, —З–Є—Б–ї–Њ –њ—А–Њ—П–≤–ї–µ–љ–Є–є –Љ–Є–∞–ї–≥–Є–Є –Є¬†–≥–Њ–ї–Њ–≤–љ–Њ–є –±–Њ–ї–Є –њ—А–Є –њ—А–Њ—Б—В—Г–і–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П—Е, —З–Є—Б–ї–Њ –њ—А–Њ—П–≤–ї–µ–љ–Є–є –Ї–Њ–љ—К—О–љ–Ї—В–Є–≤–Є—В–∞ –Є¬†—Б–ї—Г—З–∞–µ–≤ –њ–Њ—В–µ—А–Є –∞–њ–њ–µ—В–Є—В–∞. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –Њ–±–љ–∞—А—Г–ґ–µ–љ–∞ –≤—Л—А–∞–ґ–µ–љ–љ–∞—П —В–µ–љ–і–µ–љ—Ж–Є—П –Ї¬†—Б–љ–Є–ґ–µ–љ–Є—О –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В–Є –њ—А–Њ—Б—В—Г–і–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є, —З–Є—Б–ї–∞ —Б–ї—Г—З–∞–µ–≤ –≥—А–Є–њ–њ–∞, —З–Є—Б–ї–∞ –љ–∞–Ј–∞–ї—М–љ—Л—Е, —Д–∞—А–Є–љ–≥–µ–∞–ї—М–љ—Л—Е –Є¬†–±—А–Њ–љ—Е–Є–∞–ї—М–љ—Л—Е —Б–Є–Љ–њ—В–Њ–Љ–Њ–≤ [82].
–Т–Њ –≤—В–Њ—А–Њ–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є, –≤¬†–Ї–Њ—В–Њ—А–Њ–Љ —Г—З–∞—Б—В–≤–Њ–≤–∞–ї–Њ 479¬†–і–Њ–±—А–Њ–≤–Њ–ї—М—Ж–µ–≤, –Ї—Г—А—Б–Њ–≤–Њ–є –њ—А–Є–µ–Љ –С–Є–Њ–љ¬Ѓ3 –≤¬†–Ј–Є–Љ–љ–µ-–≤–µ—Б–µ–љ–љ–Є–є –њ–µ—А–Є–Њ–і —Б–Њ–Ї—А–∞—Й–∞–ї –љ–∞ 2¬†–і–љ—П —Б—А–µ–і–љ—О—О –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М —Н–њ–Є–Ј–Њ–і–Њ–≤ –њ—А–Њ—Б—В—Г–і–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є –Є¬†—Б–љ–Є–ґ–∞–ї –≤—Л—А–∞–ґ–µ–љ–љ–Њ—Б—В—М –Є—Е —Б–Є–Љ–њ—В–Њ–Љ–Њ–≤ –≤¬†—Б—А–∞–≤–љ–µ–љ–Є–Є —Б¬†–Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є –≥—А—Г–њ–њ–Њ–є, –≤¬†–Ї–Њ—В–Њ—А–Њ–є –Є—Б–њ—Л—В—Г–µ–Љ—Л–µ –њ—А–Є–љ–Є–Љ–∞–ї–Є –∞–љ–∞–ї–Њ–≥–Є—З–љ—Л–є –Љ—Г–ї—М—В–Є–≤–Є—В–∞–Љ–Є–љ–љ–Њ-–Љ—Г–ї—М—В–Є–Љ–Є–љ–µ—А–∞–ї—М–љ—Л–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б, –љ–Њ –±–µ–Ј –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –±–∞–Ї—В–µ—А–Є–є [79]. –≠—В–Њ –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В –≤–µ–і—Г—Й—Г—О —А–Њ–ї—М –≤¬†—Г–Ї—А–µ–њ–ї–µ–љ–Є–Є –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ–Њ–є –Ј–∞—Й–Є—В—Л –њ—А–Њ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е —И—В–∞–Љ–Љ–Њ–≤ –≤¬†—Б–Њ—Б—В–∞–≤–µ –С–Є–Њ–љ¬Ѓ3, –∞¬†–Ј–љ–∞—З–µ–љ–Є–µ –≤–Є—В–∞–Љ–Є–љ–Њ–≤ –Є¬†–Љ–Є–љ–µ—А–∞–ї–Њ–≤ –≤¬†—Н—В–Њ–Љ –Њ—В–љ–Њ—И–µ–љ–Є–Є –Ј–∞–Ї–ї—О—З–∞–µ—В—Б—П –≥–ї–∞–≤–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ –≤¬†—Б–Њ–Ј–і–∞–љ–Є–Є –±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л—Е —Г—Б–ї–Њ–≤–Є–є –і–ї—П —А–µ–∞–ї–Є–Ј–∞—Ж–Є–Є –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤ —Б–Є–Љ–±–Є–Њ–љ—В–љ—Л—Е –±–∞–Ї—В–µ—А–Є–є.
–Т –Њ–±–µ–Є—Е —А–∞–±–Њ—В–∞—Е –љ–∞ —Д–Њ–љ–µ –њ—А–Є–µ–Љ–∞ –С–Є–Њ–љ¬Ѓ3 –≤—Л—П–≤–ї–µ–љ—Л —Б—Г—Й–µ—Б—В–≤–µ–љ–љ—Л–µ –Є–Љ–Љ—Г–љ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Є–Ј–Љ–µ–љ–µ–љ–Є—П, —Б–Њ–≤–Њ–Ї—Г–њ–љ–Њ—Б—В—М –Ї–Њ—В–Њ—А—Л—Е –Љ–Њ–ґ–љ–Њ —А–∞—Б—Ж–µ–љ–Є—В—М –Ї–∞–Ї —Б–Є—Б—В–µ–Љ–љ—Г—О –∞–Ї—В–Є–≤–∞—Ж–Є—О –≤—А–Њ–ґ–і–µ–љ–љ—Л—Е –Є¬†–∞–і–∞–њ—В–Є–≤–љ—Л—Е, –≤¬†–њ–µ—А–≤—Г—О –Њ—З–µ—А–µ–і—М –Ї–ї–µ—В–Њ—З–љ—Л—Е, –Ј–≤–µ–љ—М–µ–≤ –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞.
–Ю—Б–љ–Њ–≤–љ—Л–µ –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–Є–µ –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–µ —Н—Д—Д–µ–Ї—В—Л –С–Є–Њ–љ¬Ѓ3, –і–Њ–Ї–∞–Ј—Л–≤–∞—О—Й–Є–µ –µ–≥–Њ —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М –Ї–∞–Ї –љ–∞–і–µ–ґ–љ–Њ–≥–Њ —Б—А–µ–і—Б—В–≤–∞ –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є –Ю–†–Т–Ш, –њ–Њ —А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ –і–≤—Г—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Б—Г–Љ–Љ–Є—А–Њ–≤–∞–љ—Л –≤¬†—В–∞–±–ї–Є—Ж–µ¬†3 [80, 81].
–Ш–Ј–≤–µ—Б—В–µ–љ —Д–∞–Ї—В —А–∞–Ј–≤–Є—В–Є—П —Г¬†–Њ–љ–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –±–Њ–ї—М–љ—Л—Е –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ—Б—В–Є –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ–Њ–є –Ј–∞—Й–Є—В—Л –≤¬†–њ–Њ—Б–ї–µ–Њ–њ–µ—А–∞—Ж–Є–Њ–љ–љ–Њ–Љ –њ–µ—А–Є–Њ–і–µ, —З—В–Њ —Б–≤—П–Ј–∞–љ–Њ –Ї–∞–Ї —Б¬†–Њ—Б–љ–Њ–≤–љ—Л–Љ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–µ–Љ, —В–∞–Ї –Є¬†—Б –њ—А–Њ–≤–Њ–і–Є–Љ—Л–Љ –ї–µ—З–µ–љ–Є–µ–Љ. –Я–µ—А–Є–Њ–њ–µ—А–∞—Ж–Є–Њ–љ–љ–∞—П –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Њ–њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–∞ –Є¬†–њ—А–Њ—В–Є–≤–Њ–Љ–Є–Ї—А–Њ–±–љ–∞—П —В–µ—А–∞–њ–Є—П —Г–ґ–µ –≤–Њ–Ј–љ–Є–Ї—И–Є—Е –Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Л—Е –Њ—Б–ї–Њ–ґ–љ–µ–љ–Є–є —Г¬†—Н—В–Є—Е –±–Њ–ї—М–љ—Л—Е –њ–Њ–і–∞–≤–ї—П—О—В —А–µ–Ј–Є–і–µ–љ—В–љ—Л–µ –Ї–Є—И–µ—З–љ—Л–µ –±–∞–Ї—В–µ—А–Є–Є, —Г—Б—Г–≥—Г–±–ї—П—П –Є–Љ–Љ—Г–љ–љ—Л–µ –љ–∞—А—Г—И–µ–љ–Є—П.
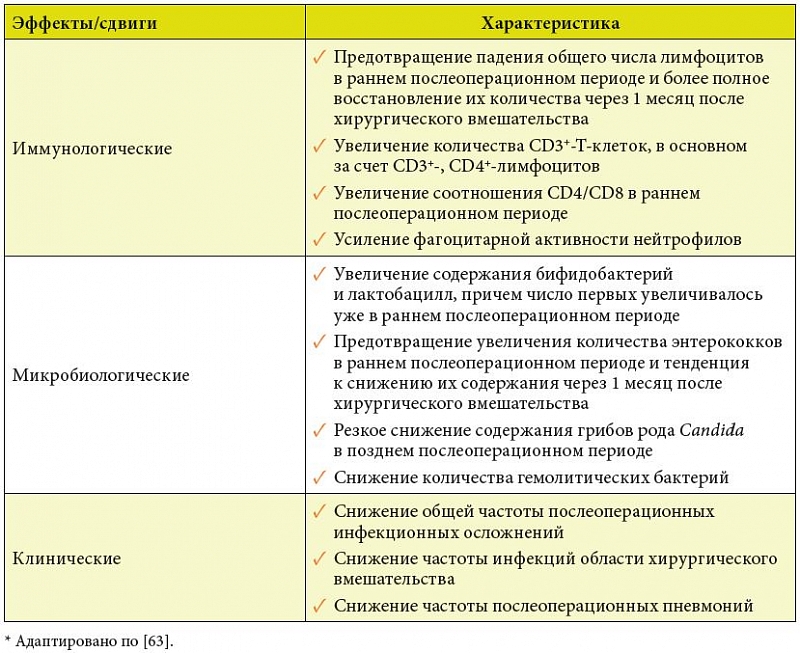
–£—З–Є—В—Л–≤–∞—П –≤—Л—И–µ—Б–Ї–∞–Ј–∞–љ–љ–Њ–µ, –∞¬†—В–∞–Ї–ґ–µ —А–Њ–ї—М —А–∞—Б—Б—В—А–Њ–є—Б—В–≤ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –≤¬†—А–∞–Ј–≤–Є—В–Є–Є –Є¬†–њ—А–Њ–≥—А–µ—Б—Б–Є—А–Њ–≤–∞–љ–Є–Є –Ї–Њ–ї–Њ—А–µ–Ї—В–∞–ї—М–љ–Њ–≥–Њ —А–∞–Ї–∞ [82], –Є–Ј—Г—З–µ–љ–Њ –≤–ї–Є—П–љ–Є–µ –С–Є–Њ–љ¬Ѓ3 –љ–∞ —А–∞–Ј–≤–Є—В–Є–µ –њ–Њ—Б–ї–µ–Њ–њ–µ—А–∞—Ж–Є–Њ–љ–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Л—Е –Њ—Б–ї–Њ–ґ–љ–µ–љ–Є–є, –Є–Љ–Љ—Г–љ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є –Ї—А–Њ–≤–Є –Є¬†–Љ–Є–Ї—А–Њ–±–Є–Њ—Ж–µ–љ–Њ–Ј –Ї–Є—И–µ—З–љ–Є–Ї–∞ —Г¬†–±–Њ–ї—М–љ—Л—Е, –Њ–њ–µ—А–Є—А–Њ–≤–∞–љ–љ—Л—Е –њ–Њ –њ–Њ–≤–Њ–і—Г —А–∞–Ї–∞ —В–Њ–ї—Б—В–Њ–є –Є¬†–њ—А—П–Љ–Њ–є –Ї–Є—И–Ї–Є [65]. –Т¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є —Г—З–∞—Б—В–≤–Њ–≤–∞–ї–Њ 50¬†–±–Њ–ї—М–љ—Л—Е, –Ї–Њ—В–Њ—А—Л–µ –±—Л–ї–Є —А–∞–љ–і–Њ–Љ–Є–Ј–Є—А–Њ–≤–∞–љ—Л –љ–∞ 2¬†–≥—А—Г–њ–њ—Л –њ–Њ 25¬†—З–µ–ї–Њ–≤–µ–Ї. –Т¬†–Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є –≥—А—Г–њ–њ–µ –њ–∞—Ж–Є–µ–љ—В—Л –њ–Њ–ї—Г—З–∞–ї–Є —Б—В–∞–љ–і–∞—А—В–љ–Њ–µ –ї–µ—З–µ–љ–Є–µ, –≤–Ї–ї—О—З–∞—О—Й–µ–µ –њ–µ—А–Є–Њ–њ–µ—А–∞—Ж–Є–Њ–љ–љ—Г—О –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Њ–њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї—Г. –Т¬†—В–µ—Б—В-–≥—А—Г–њ–њ–µ –±–Њ–ї—М–љ—Л–µ, –Ї—А–Њ–Љ–µ —В–Њ–≥–Њ, –њ–Њ–ї—Г—З–∞–ї–Є –С–Є–Њ–љ¬Ѓ3 –њ–Њ 1¬†—В–∞–±–ї–µ—В–Ї–µ –≤¬†—Б—Г—В–Ї–Є –≤¬†—В–µ—З–µ–љ–Є–µ 5¬†–і–љ–µ–є –і–Њ –Њ–њ–µ—А–∞—Ж–Є–Є –Є¬†30¬†—Б—Г—В–Њ–Ї –њ–Њ—Б–ї–µ –љ–µ–µ, –љ–∞—З–Є–љ–∞—П —Б¬†5-–≥–Њ –і–љ—П –њ–Њ—Б–ї–µ –Њ–њ–µ—А–∞—Ж–Є–Є. –£¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤, –њ–Њ–ї—Г—З–∞–≤—И–Є—Е –С–Є–Њ–љ¬Ѓ3, —Б–љ–Є–ґ–∞–ї–∞—Б—М —З–∞—Б—В–Њ—В–∞ —А–∞–Ј–≤–Є—В–Є—П –Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Л—Е –Њ—Б–ї–Њ–ґ–љ–µ–љ–Є–є, —З—В–Њ –Ї–Њ—А—А–µ–ї–Є—А–Њ–≤–∞–ї–Њ —Б¬†—Ж–µ–ї—Л–Љ —А—П–і–Њ–Љ –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ—Л—Е –Є–Љ–Љ—Г–љ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Є¬†–Љ–Є–Ї—А–Њ–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Б–і–≤–Є–≥–Њ–≤ (—В–∞–±–ї.¬†4) [65].
–†–µ–Ј—Г–ї—М—В–∞—В—Л —Н—В–Њ–є —А–∞–±–Њ—В—Л –і–Њ–Ї–∞–Ј—Л–≤–∞—О—В —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –С–Є–Њ–љ¬Ѓ3 —Г—Б–Є–ї–Є–≤–∞—В—М –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Г—О –Ј–∞—Й–Є—В—Г —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–Є—Б—Е–Њ–і–љ—Л–Љ –Є–Љ–Љ—Г–љ–Њ–і–µ—Д–Є—Ж–Є—В–љ—Л–Љ —Б–Њ—Б—В–Њ—П–љ–Є–µ–Љ, –∞¬†—В–∞–Ї–ґ–µ –љ–Њ—А–Љ–∞–ї–Є–Ј–Њ–≤–∞—В—М —Б–Њ—Б—В–Њ—П–љ–Є–µ –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Є¬†—Г–≤–µ–ї–Є—З–Є–≤–∞—В—М —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М –Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤ –Ї–Њ–ї–Њ–љ–Є–Ј–∞—Ж–Є–Њ–љ–љ–Њ–є —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤, –Њ–њ–µ—А–Є—А–Њ–≤–∞–љ–љ—Л—Е –њ–Њ –њ–Њ–≤–Њ–і—Г –Ї–Њ–ї–Њ—А–µ–Ї—В–∞–ї—М–љ–Њ–≥–Њ —А–∞–Ї–∞. –Я—А–Є–љ–Є–Љ–∞—П –≤–Њ –≤–љ–Є–Љ–∞–љ–Є–µ, —З—В–Њ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є –Ї–ї–µ—В–Њ—З–љ–Њ–≥–Њ –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞ —Г¬†–±–Њ–ї—М–љ—Л—Е —Б–Њ –Љ–љ–Њ–≥–Є–Љ–Є –Ј–ї–Њ–Ї–∞—З–µ—Б—В–≤–µ–љ–љ—Л–Љ–Є –Њ–њ—Г—Е–Њ–ї—П–Љ–Є —П–≤–ї—П—О—В—Б—П –Ї—А–Є—В–µ—А–Є—П–Љ–Є –њ—А–Њ–≥–љ–Њ–Ј–∞ –љ–µ —В–Њ–ї—М–Ї–Њ —А–∞–Ј–≤–Є—В–Є—П –њ–Њ—Б–ї–µ–Њ–њ–µ—А–∞—Ж–Є–Њ–љ–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Л—Е –Њ—Б–ї–Њ–ґ–љ–µ–љ–Є–є, –љ–Њ –Є¬†—В–µ—З–µ–љ–Є—П —Б–∞–Љ–Њ–≥–Њ –љ–µ–Њ–њ–ї–∞—Б—В–Є—З–µ—Б–Ї–Њ–≥–Њ –њ—А–Њ—Ж–µ—Б—Б–∞ [83вАУ85], –≤—Л—П–≤–ї–µ–љ–љ—Л–µ –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л –С–Є–Њ–љ¬Ѓ3 –њ—А–µ–і—Б—В–∞–≤–ї—П—О—В—Б—П –Љ–љ–Њ–≥–Њ–Њ–±–µ—Й–∞—О—Й–Є–Љ–Є –≤¬†–њ–ї–∞–љ–µ –њ—А–Њ–≥–љ–Њ–Ј–∞ –Њ—Б–љ–Њ–≤–љ–Њ–≥–Њ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П.
–С—Л–ї–∞ –њ—А–Њ–≤–µ–і–µ–љ–∞ –Њ—Ж–µ–љ–Ї–∞ —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є –Є¬†–±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В–Є –С–Є–Њ–љ¬Ѓ3¬†–Ъ–Є–і –≤¬†–Ї–∞—З–µ—Б—В–≤–µ —Б—А–µ–і—Б—В–≤–∞ –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є —А–µ—Б–њ–Є—А–∞—В–Њ—А–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–є —Г¬†—З–∞—Б—В–Њ –±–Њ–ї–µ—О—Й–Є—Е –і–µ—В–µ–є –≤¬†–≤–Њ–Ј—А–∞—Б—В–µ 7вАУ8¬†–ї–µ—В, –Ї–Њ—В–Њ—А—Л–µ –љ–∞—Е–Њ–і–Є–ї–Є—Б—М –љ–∞ –і–Є—Б–њ–∞–љ—Б–µ—А–љ–Њ–Љ –љ–∞–±–ї—О–і–µ–љ–Є–Є –њ–Њ –Љ–µ—Б—В—Г –ґ–Є—В–µ–ї—М—Б—В–≤–∞. –†–∞–±–Њ—В–∞ –≤—Л–њ–Њ–ї–љ–µ–љ–∞ –њ–Њ–і —А—Г–Ї–Њ–≤–Њ–і—Б—В–≤–Њ–Љ –њ—А–Њ—Д–µ—Б—Б–Њ—А–∞ –Ы.–Т.¬†–§–µ–Ї–ї–Є—Б–Њ–≤–Њ–є¬†вАУ —А—Г–Ї–Њ–≤–Њ–і–Є—В–µ–ї—П –Њ—В–і–µ–ї–µ–љ–Є—П –і–µ—В—Б–Ї–Є—Е –Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Л—Е –±–Њ–ї–µ–Ј–љ–µ–є –Ь–Њ—Б–Ї–Њ–≤—Б–Ї–Њ–≥–Њ –Њ–±–ї–∞—Б—В–љ–Њ–≥–Њ –љ–∞—Г—З–љ–Њ-–Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М—Б–Ї–Њ–≥–Њ –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Њ–≥–Њ –Є–љ—Б—В–Є—В—Г—В–∞ –Є–Љ.¬†–Ь.–§.¬†–Т–ї–∞–і–Є–Љ–Є—А—Б–Ї–Њ–≥–Њ. –Т¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –≤–Ї–ї—О—З–µ–љ 61¬†—А–µ–±–µ–љ–Њ–Ї, –њ—А–Є–±—Л–≤—И–Є–є –љ–∞ –Њ–Ј–і–Њ—А–Њ–≤–Є—В–µ–ї—М–љ–Њ–µ –ї–µ—З–µ–љ–Є–µ –≤¬†–¶–µ–љ—В—А–∞–ї—М–љ—Л–є –і–µ—В—Б–Ї–Є–є –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–є —Б–∞–љ–∞—В–Њ—А–Є–є ¬Ђ–Ь–∞–ї–∞—Е–Њ–≤–Ї–∞¬ї –≤¬†–њ–µ—А–Є–Њ–і –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ–≥–Њ —Б–µ–Ј–Њ–љ–љ–Њ–≥–Њ –њ–Њ–і—К–µ–Љ–∞ –Ј–∞–±–Њ–ї–µ–≤–∞–µ–Љ–Њ—Б—В–Є –Њ—Б—В—А—Л–Љ–Є —А–µ—Б–њ–Є—А–∞—В–Њ—А–љ—Л–Љ–Є –Є–љ—Д–µ–Ї—Ж–Є—П–Љ–Є. –Т–Ї–ї—О—З–µ–љ–љ—Л–µ –≤¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –і–µ—В–Є –±—Л–ї–Є —А–∞–љ–і–Њ–Љ–Є–Ј–Є—А–Њ–≤–∞–љ—Л –љ–∞ –≥—А—Г–њ–њ—Г (n = 28), –≤¬†–Ї–Њ—В–Њ—А–Њ–є –њ—А–Њ–≤–Њ–і–Є–ї—Б—П –Ї–Њ–Љ–њ–ї–µ–Ї—Б –Њ–±—Й–µ–њ—А–Є–љ—П—В—Л—Е –Њ–Ј–і–Њ—А–Њ–≤–Є—В–µ–ї—М–љ—Л—Е –Љ–µ—А–Њ–њ—А–Є—П—В–Є–є, –Є¬†–≥—А—Г–њ–њ—Г (n = 33), –≤¬†–Ї–Њ—В–Њ—А–Њ–є —Н—В–Њ—В –Ї–Њ–Љ–њ–ї–µ–Ї—Б –і–Њ–њ–Њ–ї–љ—П–ї–Є –њ—А–Є–µ–Љ–Њ–Љ –С–Є–Њ–љ¬Ѓ3¬†–Ъ–Є–і –њ–Њ 1¬†–ґ–µ–≤–∞—В–µ–ї—М–љ–Њ–є —В–∞–±–ї–µ—В–Ї–µ –µ–ґ–µ–і–љ–µ–≤–љ–Њ –≤¬†—В–µ—З–µ–љ–Є–µ 30¬†–і–љ–µ–є. –С–∞–Ј–Є—Б–љ—Л–µ –Њ–Ј–і–Њ—А–Њ–≤–Є—В–µ–ї—М–љ—Л–µ –Љ–µ—А–Њ–њ—А–Є—П—В–Є—П –≤¬†—Б–∞–љ–∞—В–Њ—А–Є–Є –≤–Ї–ї—О—З–∞–ї–Є –Љ–∞—Б—Б–∞–ґ, —Д–Є–Ј–Є–Њ–њ—А–Њ—Ж–µ–і—Г—А—Л, –ї–µ—З–µ–±–љ—Г—О —Д–Є–Ј–Ї—Г–ї—М—В—Г—А—Г, –њ–Њ—Б–µ—Й–µ–љ–Є–µ –≥–∞–ї–Њ–Ї–∞–Љ–µ—А—Л, –±–∞—Б—Б–µ–є–љ–∞, –њ–Њ—В—А–µ–±–ї–µ–љ–Є–µ –Љ–Є–љ–µ—А–∞–ї—М–љ–Њ–є –≤–Њ–і—Л –Є¬†–Ї–Є—Б–ї–Њ—А–Њ–і–љ—Л—Е –Ї–Њ–Ї—В–µ–є–ї–µ–є. –Ф—А—Г–≥–Є–µ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є –Є¬†–Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї—П—В–Њ—А—Л –≤¬†–Њ–±–µ–Є—Е –≥—А—Г–њ–њ–∞—Е –љ–µ –њ—А–Є–Љ–µ–љ—П–ї–Є—Б—М. –Я–Њ–і—А–Њ–±–љ–Њ–µ –Њ–њ–Є—Б–∞–љ–Є–µ —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤ —Н—В–Њ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П —Б—В–∞–љ–µ—В –њ—А–µ–і–Љ–µ—В–Њ–Љ –Њ—В–і–µ–ї—М–љ–Њ–є –љ–∞—Г—З–љ–Њ–є –њ—Г–±–ї–Є–Ї–∞—Ж–Є–Є. –Я—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ—Л–µ –і–∞–љ–љ—Л–µ –њ–Њ –Ї–ї–Є–љ–Є–Ї–Њ-–ї–∞–±–Њ—А–∞—В–Њ—А–љ–Њ–є —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є –С–Є–Њ–љ¬Ѓ3¬†–Ъ–Є–і —Г¬†—З–∞—Б—В–Њ –±–Њ–ї–µ—О—Й–Є—Е –і–µ—В–µ–є —Б—Г–Љ–Љ–Є—А–Њ–≤–∞–љ—Л –≤¬†—В–∞–±–ї–Є—Ж–µ¬†5. –Т—Л—П–≤–ї–µ–љ–љ—Л–є –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є—З–µ—Б–Ї–Є–є —Н—Д—Д–µ–Ї—В –С–Є–Њ–љ¬Ѓ3¬†–Ъ–Є–і –≤¬†–Њ—В–љ–Њ—И–µ–љ–Є–Є –Њ—Б—В—А—Л—Е —А–µ—Б–њ–Є—А–∞—В–Њ—А–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–є, —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞—О—Й–Є–є—Б—П –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ—Л–Љ–Є –Є–Љ–Љ—Г–љ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ–Є –Є¬†–Љ–Є–Ї—А–Њ–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ–Є –Є–Ј–Љ–µ–љ–µ–љ–Є—П–Љ–Є, –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В —Ж–µ–ї–µ—Б–Њ–Њ–±—А–∞–Ј–љ–Њ—Б—В—М –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П —Н—В–Њ–≥–Њ –њ—А–µ–њ–∞—А–∞—В–∞ —Г¬†—З–∞—Б—В–Њ –±–Њ–ї–µ—О—Й–Є—Е –і–µ—В–µ–є —Б¬†—Ж–µ–ї—М—О —Г–Ї—А–µ–њ–ї–µ–љ–Є—П –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ–Њ–є –Ј–∞—Й–Є—В—Л.
–Ч–∞–Ї–ї—О—З–µ–љ–Є–µ
–Э–∞–±–ї—О–і–∞–µ–Љ–∞—П –≤¬†–њ–Њ—Б–ї–µ–і–љ–Є–µ –≥–Њ–і—Л –≤–Њ–ї–љ–∞ —А–∞–±–Њ—В –њ–Њ –Є–Ј—Г—З–µ–љ–Є—О –Љ–љ–Њ–≥–Њ–≥—А–∞–љ–љ—Л—Е —Б–Є–Љ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –≤–Ј–∞–Є–Љ–Њ–Њ—В–љ–Њ—И–µ–љ–Є–є –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–∞ –Є¬†–Ї–Њ–ї–Њ–љ–Є–Ј–Є—А—Г—О—Й–Є—Е –µ–≥–Њ –±–∞–Ї—В–µ—А–Є–є –љ–µ –Њ—Б—В–∞–≤–ї—П–µ—В –њ–Њ–≤–Њ–і–∞ –і–ї—П —Б–Њ–Љ–љ–µ–љ–Є–є –≤¬†–≤–∞–ґ–љ–µ–є—И–µ–є —А–Њ–ї–Є –Ї–Њ–Љ–Љ–µ–љ—Б–∞–ї–Њ–≤ –≤¬†–њ–Њ–і–і–µ—А–ґ–∞–љ–Є–Є –Є–Љ–Љ—Г–љ–љ–Њ–≥–Њ –≥–Њ–Љ–µ–Њ—Б—В–∞–Ј–∞, –∞¬†—В–∞–Ї–ґ–µ –≤¬†—Д–Є–ї–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Њ–є –Є¬†–Њ–љ—В–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Њ–є —Н–≤–Њ–ї—О—Ж–Є–Є –Є–Љ–Љ—Г–љ–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –Є¬†–њ–Њ–і–і–µ—А–ґ–∞–љ–Є–Є –Є–Љ–Љ—Г–љ–љ–Њ–≥–Њ –≥–Њ–Љ–µ–Њ—Б—В–∞–Ј–∞ —Е–Њ–Ј—П–Є–љ–∞. –≠—В–Њ –Њ–±—Г—Б–ї–Њ–≤–Є–ї–Њ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –њ–Њ–Є—Б–Ї–∞ —Б—А–µ–і—Б—В–≤ –Ї–Њ—А—А–µ–Ї—Ж–Є–Є –Є–Љ–Љ—Г–љ–љ—Л—Е —А–∞—Б—Б—В—А–Њ–є—Б—В–≤ —Б—А–µ–і–Є –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤. –†–Њ—Б—В –Њ–±—К–µ–Љ–Њ–≤ –њ–Њ—В—А–µ–±–ї–µ–љ–Є—П –њ–Њ—Б–ї–µ–і–љ–Є—Е –≤—Л–і–≤–Є–љ—Г–ї –љ–∞ –њ–µ—А–≤—Л–є –њ–ї–∞–љ –≤–Њ–њ—А–Њ—Б –Њ–± –Є—Е –±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В–Є. –°–Њ–±–ї—О–і–µ–љ–Є–µ —А–∞–Ј—А–∞–±–Њ—В—З–Є–Ї–∞–Љ–Є –Є¬†–њ—А–Њ–Є–Ј–≤–Њ–і–Є—В–µ–ї—П–Љ–Є —В—А–µ–±–Њ–≤–∞–љ–Є–є –Ї–Њ–љ—В—А–Њ–ї—П —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є –Є¬†–±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В–Є, —Б—Д–Њ—А–Љ—Г–ї–Є—А–Њ–≤–∞–љ–љ—Л—Е –∞–≤—В–Њ—А–Є—В–µ—В–љ—Л–Љ–Є –Љ–µ–ґ–і—Г–љ–∞—А–Њ–і–љ—Л–Љ–Є –Њ—А–≥–∞–љ–Є–Ј–∞—Ж–Є—П–Љ–Є, —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В –њ–Њ—П–≤–ї–µ–љ–Є—О –љ–∞ —А—Л–љ–Ї–µ –±–µ–Ј–Њ–њ–∞—Б–љ—Л—Е –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –Ї–Њ—В–Њ—А—Л–µ –Љ–Њ–≥—Г—В —Б–ї—Г–ґ–Є—В—М –і–µ–є—Б—В–≤–µ–љ–љ—Л–Љ –Є–љ—Б—В—А—Г–Љ–µ–љ—В–Њ–Љ –і–ї—П —Г–Ї—А–µ–њ–ї–µ–љ–Є—П –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ–Њ–є –Ј–∞—Й–Є—В—Л. –Ю–і–љ–∞–Ї–Њ –њ—А–Њ–±–Є–Њ—В–Є–Ї–Є —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ —А–∞–Ј–ї–Є—З–∞—О—В—Б—П –њ–Њ –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є, –±–Њ–ї–µ–µ —В–Њ–≥–Њ, –≤–ї–Є—П–љ–Є–µ –Љ–љ–Њ–≥–Є—Е –Є–Ј –љ–Є—Е –љ–∞ –њ—А–Њ—В–Є–≤–Њ–Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Г—О –Ј–∞—Й–Є—В—Г –љ–µ –Є–Ј—Г—З–µ–љ–Њ. –С–Є–Њ–љ¬Ѓ3 –Є¬†–С–Є–Њ–љ¬Ѓ3¬†–Ъ–Є–і, —П–≤–ї—П—П—Б—М –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї—П–Љ–Є –њ–Њ—Б–ї–µ–і–љ–µ–≥–Њ –њ–Њ–Ї–Њ–ї–µ–љ–Є—П –Ї–Њ–Љ–њ–ї–µ–Ї—Б–љ—Л—Е –њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –Њ—В–≤–µ—З–∞—О—Й–Є—Е —В—А–µ–±–Њ–≤–∞–љ–Є—П–Љ FAO/WHO –Є¬†–Њ–±–ї–∞–і–∞—О—Й–Є—Е –і–Њ–Ї–∞–Ј–∞–љ–љ–Њ–є –Є–Љ–Љ—Г–љ–Њ—В—А–Њ–њ–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В—М—О –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Њ–є —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М—О, –њ—А–µ–і—Б—В–∞–≤–ї—П—О—В —Б–Њ–±–Њ–є —Б—А–µ–і—Б—В–≤–∞ –≤—Л–±–Њ—А–∞ –і–ї—П –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є —А–µ—Б–њ–Є—А–∞—В–Њ—А–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–є –Є¬†–њ–Њ—Б–ї–µ–Њ–њ–µ—А–∞—Ж–Є–Њ–љ–љ—Л—Е –Є–љ—Д–µ–Ї—Ж–Є–Њ–љ–љ—Л—Е –Њ—Б–ї–Њ–ґ–љ–µ–љ–Є–є —Г¬†–≤–Ј—А–Њ—Б–ї—Л—Е –Є¬†–і–µ—В–µ–є.
Probiotics for anti-infectious protection in context of immunogomeostatic role of microbiota
O.V. Kalyuzhin
I.M. Sechenov First Moscow State Medical University of the Ministry of Health of the Russian Federation, Chair of Clinical Immunology and Allergy
Contact person: Oleg Vitalyevich Kalyuzhin, kalyuzhin@list.ru
Several debating points concerning efficacy and safety of probiotics are discussed. Up-to-date information on the immunogomeostatic role of intestinal microbiota and use of viable bacterial cells for the restoration/correction of immune gomeostasis has been presented. Probiotics exhibiting properties of synbiotics and symbiotics and containing well-balanced vitamin and mineral complex have been described. Probiotics that meet international safety requirements and possess proven immunomodulatory activities and clinical efficacy are recommended for the correction of immune disorders in adult and infants and for the prevention of acute respiratory diseases.
–£–≤–∞–ґ–∞–µ–Љ—Л–є –њ–Њ—Б–µ—В–Є—В–µ–ї—М uMEDp!
–£–≤–µ–і–Њ–Љ–ї—П–µ–Љ –Т–∞—Б –Њ —В–Њ–Љ, —З—В–Њ –Ј–і–µ—Б—М —Б–Њ–і–µ—А–ґ–Є—В—Б—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П, –њ—А–µ–і–љ–∞–Ј–љ–∞—З–µ–љ–љ–∞—П –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –і–ї—П —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П.
–Х—Б–ї–Є –Т—Л –љ–µ —П–≤–ї—П–µ—В–µ—Б—М —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–Љ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П, –∞–і–Љ–Є–љ–Є—Б—В—А–∞—Ж–Є—П –љ–µ –љ–µ—Б–µ—В –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ—Б—В–Є –Ј–∞ –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ –њ–Њ—Б–ї–µ–і—Б—В–≤–Є—П, –≤–Њ–Ј–љ–Є–Ї—И–Є–µ –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–∞–Љ–Њ—Б—В–Њ—П—В–µ–ї—М–љ–Њ–≥–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Т–∞–Љ–Є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є —Б –њ–Њ—А—В–∞–ї–∞ –±–µ–Ј –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є –Ї–Њ–љ—Б—Г–ї—М—В–∞—Ж–Є–Є —Б –≤—А–∞—З–Њ–Љ.
–Э–∞–ґ–Є–Љ–∞—П –љ–∞ –Ї–љ–Њ–њ–Ї—Г ¬Ђ–Т–Њ–є—В–Є¬ї, –Т—Л –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В–µ, —З—В–Њ —П–≤–ї—П–µ—В–µ—Б—М –≤—А–∞—З–Њ–Љ –Є–ї–Є —Б—В—Г–і–µ–љ—В–Њ–Љ –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–≥–Њ –≤—Г–Ј–∞.