–Ď–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č–Ļ –į–Ĺ–į–Ľ–ł–∑ –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ—Ź—Ä–Ĺ–ĺ-–≥–Ķ–Ĺ–Ķ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤ –Ņ—Ä–ł –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł–ł –í1- –ł –≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤
- –ź–Ĺ–Ĺ–ĺ—ā–į—Ü–ł—Ź
- –°—ā–į—ā—Ć—Ź
- –°—Ā—č–Ľ–ļ–ł
- English
–í1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č –Ľ–ĺ–ļ–į–Ľ–ł–∑—É—é—ā—Ā—Ź –Ņ—Ä–Ķ–ł–ľ—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –≤ —Ā–Ķ—Ä–ĺ–∑–Ĺ—č—Ö –Ņ–ĺ–Ľ–ĺ—Ā—ā—Ź—Ö (–Ī—Ä—é—ą–Ĺ–ĺ–Ļ –ł –Ņ–Ľ–Ķ–≤—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ) –ł –Ľ–ł–ľ—Ą–ĺ–ł–ī–Ĺ–ĺ–Ļ —ā–ļ–į–Ĺ–ł —Ā–Ľ–ł–∑–ł—Ā—ā—č—Ö –ĺ–Ī–ĺ–Ľ–ĺ—á–Ķ–ļ. –í —Ā–Ľ–ł–∑–ł—Ā—ā–ĺ–Ļ –ĺ–Ī–ĺ–Ľ–ĺ—á–ļ–Ķ –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į –í1-–ļ–Ľ–Ķ—ā–ļ–ł –≤—č—Ä–į–Ī–į—ā—č–≤–į—é—ā –∑–Ĺ–į—á–ł—ā–Ķ–Ľ—Ć–Ĺ—É—é —á–į—Ā—ā—Ć –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–į (Ig) –ź.
–Ē–Ľ—Ź –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł—Ź –ł–∑–ĺ—ā–ł–Ņ–ĺ–≤ –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤ —ā—Ä–Ķ–Ī—É—é—ā—Ā—Ź —Ā–ł–≥–Ĺ–į–Ľ—č –ĺ—ā –Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤, –ĺ–ī–Ĺ–į–ļ–ĺ –≤ —ā–ĺ–Ĺ–ļ–ĺ–Ļ –ļ–ł—ą–ļ–Ķ –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ –ł–∑–ĺ—ā–ł–Ņ–į —Ā IgM –Ĺ–į IgA –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā –≤ T–Ě-–í1-–ļ–Ľ–Ķ—ā–ļ–į—Ö, –Ĺ–Ķ –Ņ–ĺ–Ľ—É—á–į—é—Č–ł—Ö –Ņ–ĺ–ľ–ĺ—Č–ł –Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ. –ü—Ä–Ķ–ī–Ņ–ĺ–Ľ–į–≥–į–Ķ—ā—Ā—Ź, —á—ā–ĺ –≤ —ć—ā–ĺ–ľ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–Ķ –ľ–ĺ–≥—É—ā —É—á–į—Ā—ā–≤–ĺ–≤–į—ā—Ć –≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č, –ļ–ĺ—ā–ĺ—Ä—č–Ķ, —ā–į–ļ –∂–Ķ –ļ–į–ļ B1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č, –≤ –ĺ–Ĺ—ā–ĺ–≥–Ķ–Ĺ–Ķ–∑–Ķ –Ņ–ĺ—Ź–≤–Ľ—Ź—é—ā—Ā—Ź —Ä–į–Ĺ—Ć—ą–Ķ –ī—Ä—É–≥–ł—Ö –Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ –ł –ł–ľ–Ķ—é—ā —Ā—Ö–ĺ–ī–Ĺ—É—é —Ā –í1-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł –Ľ–ĺ–ļ–į–Ľ–ł–∑–į—Ü–ł—é –≤ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–Ķ. –£—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ł–Ķ –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–ĺ–≤ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź –í1- –ł –≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤ –Ņ—Ä–ł –ł–ľ–ľ—É–Ĺ–Ĺ–ĺ–ľ –ĺ—ā–≤–Ķ—ā–Ķ –Ĺ–į –Ę–Ě-2-–į–Ĺ—ā–ł–≥–Ķ–Ĺ—č —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –į–ļ—ā—É–į–Ľ—Ć–Ĺ–ĺ–Ļ –∑–į–ī–į—á–Ķ–Ļ. –Ē–Ľ—Ź –Ņ—Ä–ĺ–≥–Ĺ–ĺ–∑–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č—Ö –Ņ—É—ā–Ķ–Ļ —ā–į–ļ–ĺ–≥–ĺ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź –Ņ—Ä–ł–ľ–Ķ–Ĺ—Ź–Ľ–ł –Ī–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č–Ļ –į–Ĺ–į–Ľ–ł–∑.
–ü—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–Ĺ–ĺ–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ –Ņ–ĺ–∑–≤–ĺ–Ľ–ł–Ľ–ĺ –Ņ–ĺ–Ľ—É—á–ł—ā—Ć –ī–į–Ĺ–Ĺ—č–Ķ –ĺ –ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č—Ö –ľ–į—Ä–ļ–Ķ—Ä–į—Ö –ł –ļ–Ľ—é—á–Ķ–≤—č—Ö —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ—č—Ö –Ņ—É—ā—Ź—Ö, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ľ–ĺ–≥—É—ā –Ī—č—ā—Ć –≤–ĺ–≤–Ľ–Ķ—á–Ķ–Ĺ—č –≤ —ć—ā–ĺ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ, –į —ā–į–ļ–∂–Ķ –ĺ —Ä–Ķ–≥—É–Ľ—Ź—ā–ĺ—Ä–Ĺ—č—Ö –≤–Ĺ—É—ā—Ä–ł–ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č—Ö —Ā—ā—Ä—É–ļ—ā—É—Ä–į—Ö ‚Äď –ľ–į–Ľ—č—Ö –†–Ě–ö, –ļ–ĺ—ā–ĺ—Ä—č–Ķ —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ—č –≤–Ľ–ł—Ź—ā—Ć –Ĺ–į –ĺ—ā–ī–Ķ–Ľ—Ć–Ĺ—č–Ķ –ļ–ĺ–ľ–Ņ–ĺ–Ĺ–Ķ–Ĺ—ā—č —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ—č—Ö –Ņ—É—ā–Ķ–Ļ.
–í1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č –Ľ–ĺ–ļ–į–Ľ–ł–∑—É—é—ā—Ā—Ź –Ņ—Ä–Ķ–ł–ľ—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –≤ —Ā–Ķ—Ä–ĺ–∑–Ĺ—č—Ö –Ņ–ĺ–Ľ–ĺ—Ā—ā—Ź—Ö (–Ī—Ä—é—ą–Ĺ–ĺ–Ļ –ł –Ņ–Ľ–Ķ–≤—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ) –ł –Ľ–ł–ľ—Ą–ĺ–ł–ī–Ĺ–ĺ–Ļ —ā–ļ–į–Ĺ–ł —Ā–Ľ–ł–∑–ł—Ā—ā—č—Ö –ĺ–Ī–ĺ–Ľ–ĺ—á–Ķ–ļ. –í —Ā–Ľ–ł–∑–ł—Ā—ā–ĺ–Ļ –ĺ–Ī–ĺ–Ľ–ĺ—á–ļ–Ķ –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į –í1-–ļ–Ľ–Ķ—ā–ļ–ł –≤—č—Ä–į–Ī–į—ā—č–≤–į—é—ā –∑–Ĺ–į—á–ł—ā–Ķ–Ľ—Ć–Ĺ—É—é —á–į—Ā—ā—Ć –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–į (Ig) –ź.
–Ē–Ľ—Ź –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł—Ź –ł–∑–ĺ—ā–ł–Ņ–ĺ–≤ –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤ —ā—Ä–Ķ–Ī—É—é—ā—Ā—Ź —Ā–ł–≥–Ĺ–į–Ľ—č –ĺ—ā –Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤, –ĺ–ī–Ĺ–į–ļ–ĺ –≤ —ā–ĺ–Ĺ–ļ–ĺ–Ļ –ļ–ł—ą–ļ–Ķ –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ –ł–∑–ĺ—ā–ł–Ņ–į —Ā IgM –Ĺ–į IgA –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā –≤ T–Ě-–í1-–ļ–Ľ–Ķ—ā–ļ–į—Ö, –Ĺ–Ķ –Ņ–ĺ–Ľ—É—á–į—é—Č–ł—Ö –Ņ–ĺ–ľ–ĺ—Č–ł –Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ. –ü—Ä–Ķ–ī–Ņ–ĺ–Ľ–į–≥–į–Ķ—ā—Ā—Ź, —á—ā–ĺ –≤ —ć—ā–ĺ–ľ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–Ķ –ľ–ĺ–≥—É—ā —É—á–į—Ā—ā–≤–ĺ–≤–į—ā—Ć –≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č, –ļ–ĺ—ā–ĺ—Ä—č–Ķ, —ā–į–ļ –∂–Ķ –ļ–į–ļ B1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č, –≤ –ĺ–Ĺ—ā–ĺ–≥–Ķ–Ĺ–Ķ–∑–Ķ –Ņ–ĺ—Ź–≤–Ľ—Ź—é—ā—Ā—Ź —Ä–į–Ĺ—Ć—ą–Ķ –ī—Ä—É–≥–ł—Ö –Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ –ł –ł–ľ–Ķ—é—ā —Ā—Ö–ĺ–ī–Ĺ—É—é —Ā –í1-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł –Ľ–ĺ–ļ–į–Ľ–ł–∑–į—Ü–ł—é –≤ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–Ķ. –£—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ł–Ķ –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–ĺ–≤ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź –í1- –ł –≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤ –Ņ—Ä–ł –ł–ľ–ľ—É–Ĺ–Ĺ–ĺ–ľ –ĺ—ā–≤–Ķ—ā–Ķ –Ĺ–į –Ę–Ě-2-–į–Ĺ—ā–ł–≥–Ķ–Ĺ—č —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –į–ļ—ā—É–į–Ľ—Ć–Ĺ–ĺ–Ļ –∑–į–ī–į—á–Ķ–Ļ. –Ē–Ľ—Ź –Ņ—Ä–ĺ–≥–Ĺ–ĺ–∑–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č—Ö –Ņ—É—ā–Ķ–Ļ —ā–į–ļ–ĺ–≥–ĺ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź –Ņ—Ä–ł–ľ–Ķ–Ĺ—Ź–Ľ–ł –Ī–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č–Ļ –į–Ĺ–į–Ľ–ł–∑.
–ü—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–Ĺ–ĺ–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ –Ņ–ĺ–∑–≤–ĺ–Ľ–ł–Ľ–ĺ –Ņ–ĺ–Ľ—É—á–ł—ā—Ć –ī–į–Ĺ–Ĺ—č–Ķ –ĺ –ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č—Ö –ľ–į—Ä–ļ–Ķ—Ä–į—Ö –ł –ļ–Ľ—é—á–Ķ–≤—č—Ö —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ—č—Ö –Ņ—É—ā—Ź—Ö, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ľ–ĺ–≥—É—ā –Ī—č—ā—Ć –≤–ĺ–≤–Ľ–Ķ—á–Ķ–Ĺ—č –≤ —ć—ā–ĺ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ, –į —ā–į–ļ–∂–Ķ –ĺ —Ä–Ķ–≥—É–Ľ—Ź—ā–ĺ—Ä–Ĺ—č—Ö –≤–Ĺ—É—ā—Ä–ł–ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č—Ö —Ā—ā—Ä—É–ļ—ā—É—Ä–į—Ö ‚Äď –ľ–į–Ľ—č—Ö –†–Ě–ö, –ļ–ĺ—ā–ĺ—Ä—č–Ķ —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ—č –≤–Ľ–ł—Ź—ā—Ć –Ĺ–į –ĺ—ā–ī–Ķ–Ľ—Ć–Ĺ—č–Ķ –ļ–ĺ–ľ–Ņ–ĺ–Ĺ–Ķ–Ĺ—ā—č —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ—č—Ö –Ņ—É—ā–Ķ–Ļ.
–í–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ
–í1-–ļ–Ľ–Ķ—ā–ļ–ł¬†‚Äď –ĺ—ā–ī–Ķ–Ľ—Ć–Ĺ–į—Ź —Ā—É–Ī–Ņ–ĺ–Ņ—É–Ľ—Ź—Ü–ł—Ź B-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤ –≤¬†–ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–Ķ —á–Ķ–Ľ–ĺ–≤–Ķ–ļ–į –ł¬†–ľ—č—ą–Ķ–Ļ.
–í¬†–ĺ—ā–Ľ–ł—á–ł–Ķ –ĺ—ā¬†B2-–ļ–Ľ–Ķ—ā–ĺ–ļ, –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–Ĺ—č—Ö –Ņ—Ä–Ķ–ł–ľ—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –≤¬†—Ā–Ķ–Ľ–Ķ–∑–Ķ–Ĺ–ļ–Ķ, –Ľ–ł–ľ—Ą–ĺ—É–∑–Ľ–į—Ö –ł¬†–ļ–ĺ—Ā—ā–Ĺ–ĺ–ľ –ľ–ĺ–∑–≥–Ķ, –í1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č –Ľ–ĺ–ļ–į–Ľ–ł–∑—É—é—ā—Ā—Ź –≤¬†—Ā–Ķ—Ä–ĺ–∑–Ĺ—č—Ö –Ņ–ĺ–Ľ–ĺ—Ā—ā—Ź—Ö (–Ņ–Ķ—Ä–ł—ā–ĺ–Ĺ–Ķ–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł¬†–Ī—Ä—é—ą–Ĺ–ĺ–Ļ), –į¬†—ā–į–ļ–∂–Ķ –≤¬†–Ľ–ł–ľ—Ą–ĺ–ł–ī–Ĺ–ĺ–Ļ —ā–ļ–į–Ĺ–ł —Ā–Ľ–ł–∑–ł—Ā—ā—č—Ö –ĺ–Ī–ĺ–Ľ–ĺ—á–Ķ–ļ, –≤¬†—á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł –≤¬†—Ā–Ľ–ł–∑–ł—Ā—ā–ĺ–Ļ –ĺ–Ī–ĺ–Ľ–ĺ—á–ļ–Ķ –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į.
–í¬†–ĺ–Ĺ—ā–ĺ–≥–Ķ–Ĺ–Ķ–∑–Ķ –í1-–ļ–Ľ–Ķ—ā–ļ–ł –Ņ–ĺ—Ź–≤–Ľ—Ź—é—ā—Ā—Ź —Ä–į–Ĺ—Ć—ą–Ķ –í2-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤. –ü—Ä–Ķ–ī—ą–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ł–ļ–ł –í1-–ļ–Ľ–Ķ—ā–ĺ–ļ –ľ–ł–≥—Ä–ł—Ä—É—é—ā –ł–∑¬†—Ą–Ķ—ā–į–Ľ—Ć–Ĺ–ĺ–Ļ –Ņ–Ķ—á–Ķ–Ĺ–ł –≤¬†—Ā–Ķ—Ä–ĺ–∑–Ĺ—č–Ķ –Ņ–ĺ–Ľ–ĺ—Ā—ā–ł. –°—á–ł—ā–į–Ķ—ā—Ā—Ź, —á—ā–ĺ –ĺ–Ĺ–ł –ľ–ĺ–≥—É—ā —Ā–ĺ—Ö—Ä–į–Ĺ—Ź—ā—Ć—Ā—Ź –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –≤—Ā–Ķ–Ļ –∂–ł–∑–Ĺ–ł. –≠—ā–ĺ –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–į–Ķ—ā—Ā—Ź –∑–į—Ä–ĺ–ī—č—ą–Ķ–≤–ĺ–Ļ –ĺ—Ä–≥–į–Ĺ–ł–∑–į—Ü–ł–Ķ–Ļ¬†–≥–Ķ–Ĺ–ĺ–≤ –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤ (Ig) –í1-–ļ–Ľ–Ķ—ā–ĺ–ļ. –ě–ī–Ĺ–į–ļ–ĺ –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ—č –ī–į–Ĺ–Ĺ—č–Ķ –嬆—ā–ĺ–ľ, —á—ā–ĺ –Ņ–嬆–ļ—Ä–į–Ļ–Ĺ–Ķ–Ļ –ľ–Ķ—Ä–Ķ —á–į—Ā—ā—Ć –í1-–ļ–Ľ–Ķ—ā–ĺ–ļ –≤–∑—Ä–ĺ—Ā–Ľ–ĺ–≥–ĺ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į –ľ–ĺ–∂–Ķ—ā –ĺ–Ī—Ä–į–∑–ĺ–≤—č–≤–į—ā—Ć—Ā—Ź –≤¬†–ļ–ĺ—Ā—ā–Ĺ–ĺ–ľ –ľ–ĺ–∑–≥–Ķ.
–í1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č –ĺ—ā–≤–Ķ—á–į—é—ā –Ņ—Ä–Ķ–ł–ľ—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –Ĺ–į¬†–Ę-–Ĺ–Ķ–∑–į–≤–ł—Ā–ł–ľ—č–Ķ (–Ę–Ě) –į–Ĺ—ā–ł–≥–Ķ–Ĺ—č 1-–≥–嬆–ł 2-–≥–嬆—ā–ł–Ņ–ĺ–≤.
–Ę–Ě-1-–į–≥–Ķ–Ĺ—ā—č –≤¬†–ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ –≤—č—Ā–ĺ–ļ–ł—Ö –ļ–ĺ–Ĺ—Ü–Ķ–Ĺ—ā—Ä–į—Ü–ł—Ź—Ö —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ—č –ł–Ĺ–ī—É—Ü–ł—Ä–ĺ–≤–į—ā—Ć –Ņ–ĺ–Ľ–ł–ļ–Ľ–ĺ–Ĺ–į–Ľ—Ć–Ĺ—É—é –į–ļ—ā–ł–≤–į—Ü–ł—é B-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤ (–∑—Ä–Ķ–Ľ—č—Ö –ł¬†–Ĺ–Ķ–∑—Ä–Ķ–Ľ—č—Ö) –ł¬†–Ņ—Ä–ĺ–ī—É–ļ—Ü–ł—é Ig. –ö–Ľ–į—Ā—Ā–ł—á–Ķ—Ā–ļ–ł–ľ –Ņ—Ä–ł–ľ–Ķ—Ä–ĺ–ľ –Ę–Ě-1-–į–Ĺ—ā–ł–≥–Ķ–Ĺ–į —Ā–Ľ—É–∂–ł—ā –Ľ–ł–Ņ–ĺ–Ņ–ĺ–Ľ–ł—Ā–į—Ö–į—Ä–ł–ī.
–Ę–Ě-2-–į–≥–Ķ–Ĺ—ā—č –ł–ľ–Ķ—é—ā –Ņ—Ä–Ķ–ł–ľ—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –Ĺ–Ķ–Ī–Ķ–Ľ–ļ–ĺ–≤—É—é –Ņ—Ä–ł—Ä–ĺ–ī—É, –Ī–ĺ–Ľ—Ć—ą–ĺ–Ļ —Ä–į–∑–ľ–Ķ—Ä –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ –ł¬†—Ä–Ķ–≥—É–Ľ—Ź—Ä–Ĺ–ĺ –Ņ–ĺ–≤—ā–ĺ—Ä—Ź—é—Č–ł–Ķ—Ā—Ź —ć–Ņ–ł—ā–ĺ–Ņ—č, –≤—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł–Ķ —á–Ķ–≥–ĺ —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ—č –≤—č–∑—č–≤–į—ā—Ć –ļ–Ľ–į—Ā—ā–Ķ—Ä–ł–∑–į—Ü–ł—é –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤—č—Ö —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–ĺ–≤ –Ĺ–į¬†–Ņ–ĺ–≤–Ķ—Ä—Ö–Ĺ–ĺ—Ā—ā–ł –í-–ļ–Ľ–Ķ—ā–ĺ–ļ. –í¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ —Ą–ĺ—Ä–ľ–ł—Ä—É—é—Č–ł–Ļ—Ā—Ź —Ā–ł–≥–Ĺ–į–Ľ –ī–ĺ—Ā—ā–ł–≥–į–Ķ—ā —Ā–ł–Ľ—č, –ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ–Ļ –ī–Ľ—Ź –į–ļ—ā–ł–≤–į—Ü–ł–ł –Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–į, –ł¬†–≤—ā–ĺ—Ä–ĺ–≥–ĺ —Ā–ł–≥–Ĺ–į–Ľ–į –ĺ—ā¬†–Ę-–ļ–Ľ–Ķ—ā–ļ–ł –Ĺ–Ķ¬†—ā—Ä–Ķ–Ī—É–Ķ—ā—Ā—Ź (—ć—ā–ĺ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ—Ź–Ķ—ā –Ę-–Ĺ–Ķ–∑–į–≤–ł—Ā–ł–ľ–ĺ—Ā—ā—Ć –ĺ—ā–≤–Ķ—ā–į). –Ę–Ě-2-–į–≥–Ķ–Ĺ—ā—č –≤¬†–ĺ—ā–Ľ–ł—á–ł–Ķ –ĺ—ā¬†–Ę–Ě-1-–į–≥–Ķ–Ĺ—ā–ĺ–≤ —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ—č –į–ļ—ā–ł–≤–ł—Ä–ĺ–≤–į—ā—Ć –í-–ļ–Ľ–Ķ—ā–ļ–ł —ā–ĺ–Ľ—Ć–ļ–ĺ —Ā–Ņ–Ķ—Ü–ł—Ą–ł—á–Ķ—Ā–ļ–ł, –Ņ–ĺ—ć—ā–ĺ–ľ—É –Ĺ–į¬†–Ĺ–ł—Ö –ĺ—ā–≤–Ķ—á–į—é—ā –Ņ—Ä–Ķ–ł–ľ—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ B1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č, —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ—č–Ķ –ļ¬†–Ę–Ě-–ĺ—ā–≤–Ķ—ā—É [1].
IgA¬†‚Äď –ĺ—Ā–Ĺ–ĺ–≤–Ĺ–ĺ–Ļ –ł–∑–ĺ—ā–ł–Ņ –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤ –≤¬†—Ā–Ľ–ł–∑–ł—Ā—ā—č—Ö –ĺ–Ī–ĺ–Ľ–ĺ—á–ļ–į—Ö, –ł–≥—Ä–į—é—Č–ł–Ļ –ļ–Ľ—é—á–Ķ–≤—É—é —Ä–ĺ–Ľ—Ć –≤¬†–Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ–ľ —Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–ł—Ä–ĺ–≤–į–Ĺ–ł–ł –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į. –í¬†—á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, IgA –Ņ—Ä–Ķ–Ņ—Ź—ā—Ā—ā–≤—É–Ķ—ā –Ņ—Ä–ĺ–Ĺ–ł–ļ–Ĺ–ĺ–≤–Ķ–Ĺ–ł—é –Ņ–į—ā–ĺ–≥–Ķ–Ĺ–ĺ–≤ –≤¬†–ĺ—Ä–≥–į–Ĺ–ł–∑–ľ —á–Ķ—Ä–Ķ–∑ —Ā–Ľ–ł–∑–ł—Ā—ā—č–Ķ –ĺ–Ī–ĺ–Ľ–ĺ—á–ļ–ł. S. Wolbank –ł¬†—Ā–ĺ–į–≤—ā. —É—Ā—ā–į–Ĺ–ĺ–≤–ł–Ľ–ł, —á—ā–ĺ –į–Ĺ—ā–ł—ā–Ķ–Ľ–į 2F5IgA, 2F5IgM –ł¬†2G12IgM —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ –ł–Ĺ–≥–ł–Ī–ł—Ä—É—é—ā –Ņ–Ķ—Ä–≤—č–Ķ —Ā—ā–į–ī–ł–ł –Ņ—Ä–ĺ–Ĺ–ł–ļ–Ĺ–ĺ–≤–Ķ–Ĺ–ł—Ź –í–ė–ß —á–Ķ—Ä–Ķ–∑ —Ā–Ľ–ł–∑–ł—Ā—ā—č–Ķ –ĺ–Ī–ĺ–Ľ–ĺ—á–ļ–ł [2].
–Ď–ĺ–Ľ—Ć—ą–į—Ź —á–į—Ā—ā—Ć IgA –≤—č—Ä–į–Ī–į—ā—č–≤–į–Ķ—ā—Ā—Ź –≤¬†—Ā–Ľ–ł–∑–ł—Ā—ā–ĺ–Ļ –ĺ–Ī–ĺ–Ľ–ĺ—á–ļ–Ķ –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į, –Ņ—Ä–Ķ–ł–ľ—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ B1-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł. –ě–Ī—Ä–į–∑–ĺ–≤–į–Ĺ–ł–Ķ –í1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–į–ľ–ł IgA –≤¬†—ā–ĺ–Ĺ–ļ–ĺ–Ļ –ļ–ł—ą–ļ–Ķ —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤—É–Ķ—ā –嬆—ā–ĺ–ľ, —á—ā–ĺ –≤¬†–Ę–Ě-–í1-–ļ–Ľ–Ķ—ā–ļ–į—Ö –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ –ł–∑–ĺ—ā–ł–Ņ–į —Ā¬†IgM –Ĺ–į¬†–ł–∑–ĺ—ā–ł–Ņ IgA [3]. –ü—Ä–ł –ĺ—ā–≤–Ķ—ā–Ķ –Ĺ–į¬†–Ę-–∑–į–≤–ł—Ā–ł–ľ—č–Ķ (–Ę–ó) –į–Ĺ—ā–ł–≥–Ķ–Ĺ—č –ī–Ľ—Ź –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł—Ź –ł–∑–ĺ—ā–ł–Ņ–ĺ–≤ –í2-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā –ī–ĺ–Ľ–∂–Ķ–Ĺ –Ņ–ĺ–Ľ—É—á–ł—ā—Ć –≤—ā–ĺ—Ä–ĺ–Ļ —Ā–ł–≥–Ĺ–į–Ľ –ĺ—ā¬†–Ę-–ļ–Ľ–Ķ—ā–ļ–ł. –ß–Ķ–ľ –≤¬†–ĺ—ā—Ā—É—ā—Ā—ā–≤–ł–Ķ –ļ–Ľ–į—Ā—Ā–ł—á–Ķ—Ā–ļ–ĺ–Ļ –Ņ–ĺ–ľ–ĺ—Č–ł –Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ –ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–ł–≤–į–Ķ—ā—Ā—Ź –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ –ļ–Ľ–į—Ā—Ā–ĺ–≤ –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤ –≤¬†–í1-–ļ–Ľ–Ķ—ā–ļ–į—Ö –ł¬†–ļ–į–ļ –ĺ–Ĺ–ĺ –ĺ—Ā—É—Č–Ķ—Ā—ā–≤–Ľ—Ź–Ķ—ā—Ā—Ź, –≤¬†–Ĺ–į—Ā—ā–ĺ—Ź—Č–Ķ–Ķ –≤—Ä–Ķ–ľ—Ź –Ĺ–Ķ¬†–ł–∑–≤–Ķ—Ā—ā–Ĺ–ĺ. –ü—Ä–Ķ–ī–Ņ–ĺ–Ľ–į–≥–į–Ķ—ā—Ā—Ź, —á—ā–ĺ –≤¬†—ć—ā–ĺ–ľ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–Ķ –ľ–ĺ–≥—É—ā —É—á–į—Ā—ā–≤–ĺ–≤–į—ā—Ƭ†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–ł [4‚Äď6].
–ď–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č —Ā—ā–į–Ľ–ł –į–ļ—ā–ł–≤–Ĺ–ĺ –ł–∑—É—á–į—ā—Ć—Ā—Ź –Ĺ–Ķ–ī–į–≤–Ĺ–ĺ. –Ě–į¬†–ī–į–Ĺ–Ĺ—č–Ļ –ľ–ĺ–ľ–Ķ–Ĺ—ā –≤—č—Ź–≤–Ľ–Ķ–Ĺ–į –ł—Ö —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ–ĺ—Ā—ā—Ć —Ä–į—Ā–Ņ–ĺ–∑–Ĺ–į–≤–į—ā—Ć –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č–Ķ –į–Ĺ—ā–ł–≥–Ķ–Ĺ—č –ł¬†–≤—č—Ā—ā—É–Ņ–į—ā—Ć –≤¬†—Ä–ĺ–Ľ–ł –į–Ĺ—ā–ł–≥–Ķ–Ĺ–Ņ—Ä–Ķ–∑–Ķ–Ĺ—ā–ł—Ä—É—é—Č–ł—Ö –ļ–Ľ–Ķ—ā–ĺ–ļ (–ź–ü–ö). –ö—Ä–ĺ–ľ–Ķ —ā–ĺ–≥–ĺ, –ĺ–Ĺ–ł –ĺ–Ī–Ľ–į–ī–į—é—ā —Ü–ł—ā–ĺ—ā–ĺ–ļ—Ā–ł—á–Ķ—Ā–ļ–ł–ľ–ł —Ā–≤–ĺ–Ļ—Ā—ā–≤–į–ľ–ł, –Ņ–ĺ—ć—ā–ĺ–ľ—É —Ä–į—Ā—Ā–ľ–į—ā—Ä–ł–≤–į—é—ā—Ā—Ź –ļ–į–ļ –Ņ–ĺ—ā–Ķ–Ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–Ķ —ā–Ķ—Ä–į–Ņ–Ķ–≤—ā–ł—á–Ķ—Ā–ļ–ł–Ķ –ľ–ł—ą–Ķ–Ĺ–ł [7].
–ď–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č, —ā–į–ļ –∂–Ķ –ļ–į–ļ B1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č, –ĺ–Ī—Ä–į–∑—É—é—ā—Ā—Ź –≤¬†–ĺ–Ĺ—ā–ĺ–≥–Ķ–Ĺ–Ķ–∑–Ķ —Ä–į–Ĺ—Ć—ą–Ķ –ī—Ä—É–≥–ł—Ö –Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤. –ě–Ĺ–ł –Ľ–ĺ–ļ–į–Ľ–ł–∑—É—é—ā—Ā—Ź –Ņ—Ä–Ķ–ł–ľ—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –Ĺ–į¬†–ļ–ĺ–∂–Ķ –ł¬†—Ā–Ľ–ł–∑–ł—Ā—ā—č—Ö –ĺ–Ī–ĺ–Ľ–ĺ—á–ļ–į—Ö, –≤¬†—ā–ĺ–ľ —á–ł—Ā–Ľ–Ķ –≤¬†–Ľ–ł–ľ—Ą–ĺ–ł–ī–Ĺ–ĺ–Ļ —ā–ļ–į–Ĺ–ł, –į—Ā—Ā–ĺ—Ü–ł–ł—Ä—É—é—Č–Ķ–Ļ—Ā—Ź —Ā–嬆—Ā–Ľ–ł–∑–ł—Ā—ā–ĺ–Ļ –ĺ–Ī–ĺ–Ľ–ĺ—á–ļ–ĺ–Ļ –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į.
–Ě–Ķ–ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ—Ā—ā—Ć –Ņ–ĺ–Ņ—É–Ľ—Ź—Ü–ł–ł –ł–Ĺ—ā—Ä–į—ć–Ņ–ł—ā–Ķ–Ľ–ł–į–Ľ—Ć–Ĺ—č—Ö¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ –ľ–ĺ–∂–Ķ—ā –ĺ–Ī—É—Ā–Ľ–ĺ–≤–ł—ā—Ć —Ä–į–∑–≤–ł—ā–ł–Ķ –Ĺ–Ķ–ļ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö —ć–Ĺ—ā–Ķ—Ä–ĺ–ļ–ĺ–Ľ–ł—ā–ĺ–≤ [8], —á—ā–ĺ —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤—É–Ķ—ā –嬆—Ä–Ķ–≥—É–Ľ—Ź—ā–ĺ—Ä–Ĺ—č—Ö —Ā–≤–ĺ–Ļ—Ā—ā–≤–į—Ö —ć—ā–ł—Ö –ļ–Ľ–Ķ—ā–ĺ–ļ.
–°–Ņ–ĺ—Ā–ĺ–Ī–Ĺ–ĺ—Ā—ā—Ƭ†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ –≤–Ľ–ł—Ź—ā—Ć –Ĺ–į¬†–ĺ–Ī—Ä–į–∑–ĺ–≤–į–Ĺ–ł–Ķ –į–Ĺ—ā–ł—ā–Ķ–Ľ –í1-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł –Ņ—Ä—Ź–ľ–ĺ–Ļ —ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ–ĺ–Ļ –Ņ—Ä–ĺ–≤–Ķ—Ä–ļ–Ķ –Ĺ–Ķ¬†–Ņ–ĺ–ī–≤–Ķ—Ä–≥–į–Ľ–į—Ā—Ć, –ł¬†–≤–ĺ–Ņ—Ä–ĺ—Ā –ĺ–Ī¬†–ł—Ö —Ä–ĺ–Ľ–ł –≤¬†–Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–ł —Ā–ł–Ĺ—ā–Ķ–∑–į —Ā¬†IgM –Ĺ–į¬†IgA –≤¬†–í1-–ļ–Ľ–Ķ—ā–ļ–į—Ö –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į –Ņ–ĺ-–Ņ—Ä–Ķ–∂–Ĺ–Ķ–ľ—É –ĺ—Ā—ā–į–Ķ—ā—Ā—Ź –ĺ—ā–ļ—Ä—č—ā—č–ľ. –ě–ī–Ĺ–į–ļ–ĺ –ł–ľ–Ķ–Ķ—ā—Ā—Ź –ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ–Ķ –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–ĺ —Ā–ĺ–ĺ–Ī—Č–Ķ–Ĺ–ł–Ļ –嬆–≤–Ľ–ł—Ź–Ĺ–ł–ł –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö —Ā—É–Ī–Ņ–ĺ–Ņ—É–Ľ—Ź—Ü–ł–Ļ¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ –Ĺ–į¬†—É—Ä–ĺ–≤–Ķ–Ĺ—Ć –Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ—č—Ö –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤ –≤¬†–ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–Ķ [9].
–ė–∑—É—á–Ķ–Ĺ–ł–Ķ –ĺ—Ā–ĺ–Ī–Ķ–Ĺ–Ĺ–ĺ—Ā—ā–Ķ–Ļ –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł—Ź –Ĺ–į¬†IgA B1-–ļ–Ľ–Ķ—ā–ĺ–ļ –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į –Ņ–ĺ–∑–≤–ĺ–Ľ–ł—ā —Ä–į–∑—Ä–į–Ī–ĺ—ā–į—ā—Ć –Ĺ–ĺ–≤—č–Ķ —ā–Ķ—Ä–į–Ņ–Ķ–≤—ā–ł—á–Ķ—Ā–ļ–ł–Ķ –Ņ–ĺ–ī—Ö–ĺ–ī—č –ļ¬†–Ľ–Ķ—á–Ķ–Ĺ–ł—é –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į –ł–Ĺ—Ą–Ķ–ļ—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–Ļ –ł¬†–Ĺ–Ķ–ł–Ĺ—Ą–Ķ–ļ—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–Ļ –Ņ—Ä–ł—Ä–ĺ–ī—č. –£—á–ł—ā—č–≤–į—Ź –≤—č—Ā–ĺ–ļ—É—é –∑–į—ā—Ä–į—ā–Ĺ–ĺ—Ā—ā—Ć —ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ—č—Ö –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ, –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ—Ź–Ľ–ĺ—Ā—Ć –Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ—č–ľ —Ā—É–∑–ł—ā—Ć –ļ—Ä—É–≥ –∑–į–ī–į—á –ł¬†–ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į—ā—Ć –Ī–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č–Ķ –ľ–Ķ—ā–ĺ–ī—č –į–Ĺ–į–Ľ–ł–∑–į.
–¶–Ķ–Ľ—Ć—é –ī–į–Ĺ–Ĺ–ĺ–Ļ —Ä–į–Ī–ĺ—ā—č —Ā—ā–į–Ľ–ĺ –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ –Ī–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–≥–ĺ –į–Ĺ–į–Ľ–ł–∑–į –ī–Ľ—Ź –≤—č—Ź–≤–Ľ–Ķ–Ĺ–ł—Ź –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č—Ö –Ņ—É—ā–Ķ–Ļ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź B1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤ —Ā¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ľ–ĺ–≥—É—ā –ł–ľ–Ķ—ā—Ć –ĺ—ā–Ĺ–ĺ—ą–Ķ–Ĺ–ł–Ķ –ļ¬†–≥—É–ľ–ĺ—Ä–į–Ľ—Ć–Ĺ–ĺ–ľ—É –ł–ľ–ľ—É–Ĺ–Ĺ–ĺ–ľ—É –ĺ—ā–≤–Ķ—ā—É –Ĺ–į¬†–Ę–Ě-2-–į–≥–Ķ–Ĺ—ā—č, –į¬†—ā–į–ļ–∂–Ķ —Ä–ĺ–Ľ–ł –ľ–į–Ľ—č—Ö –†–Ě–ö (miRNA) –≤¬†–Ņ—Ä–ĺ–ī—É–ļ—Ü–ł–ł –į–Ĺ—ā–ł—ā–Ķ–Ľ, –į–ļ—ā–ł–≤–į—Ü–ł–ł –ł¬†–Ņ—Ä–ĺ–Ľ–ł—Ą–Ķ—Ä–į—Ü–ł–ł B1-–ļ–Ľ–Ķ—ā–ĺ–ļ.
–ú–į—ā–Ķ—Ä–ł–į–Ľ –ł¬†–ľ–Ķ—ā–ĺ–ī—č
–í¬†–Ī–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–ľ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–ł –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–į –Ņ—Ä–ĺ–≥—Ä–į–ľ–ľ–į Pathway Studio¬ģ –ł¬†—Ä–Ķ—Ą–Ķ—Ä–į—ā–ł–≤–Ĺ–į—Ź –Ī–į–∑–į –ī–į–Ĺ–Ĺ—č—Ö ResNet¬ģ –ļ–ĺ–ľ–Ņ–į–Ĺ–ł–ł Ariadne Genomics (–°–®–ź).
–ě–Ī—ä–Ķ–ļ—ā–į–ľ–ł –ł–∑—É—á–Ķ–Ĺ–ł—Ź —Ā—ā–į–Ľ–ł –į–Ĺ–Ĺ–ĺ—ā–į—Ü–ł–ł –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö –ĺ–Ī—ä–Ķ–ļ—ā–ĺ–≤ (–≤¬†—á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, –Ī–Ķ–Ľ–ļ–ĺ–≤, –ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č—Ö –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤ –ł¬†–Ī–ĺ–Ľ–Ķ–∑–Ĺ–Ķ–Ļ), –į¬†—ā–į–ļ–∂–Ķ —Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–į–Ľ—Ć–Ĺ—č–Ķ —Ā–≤—Ź–∑–ł –ľ–Ķ–∂–ī—É –Ĺ–ł–ľ–ł, —Ā—Ą–ĺ—Ä–ľ–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č–Ķ –≤¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –ĺ–Ī—Ä–į–Ī–ĺ—ā–ļ–ł —ā–Ķ–ļ—Ā—ā–ĺ–≤–ĺ–≥–ĺ –ľ–į—Ā—Ā–ł–≤–į –Ņ–ĺ–Ľ–Ĺ–ĺ—ā–Ķ–ļ—Ā—ā–ĺ–≤—č—Ö —Ā—ā–į—ā–Ķ–Ļ –ł¬†–į–Ī—Ā—ā—Ä–į–ļ—ā–ĺ–≤, –ł–Ĺ–ī–Ķ–ļ—Ā–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č—Ö –≤¬†MEDLINE.
–í¬†–ļ–į—á–Ķ—Ā—ā–≤–Ķ –ľ–ĺ–ī–Ķ–Ľ–ł —Ā–Ľ—É—á–į–Ļ–Ĺ–ĺ–≥–ĺ –Ņ–Ķ—Ä–Ķ—Ā–Ķ—á–Ķ–Ĺ–ł—Ź –ľ–Ĺ–ĺ–∂–Ķ—Ā—ā–≤ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ľ–ĺ—Ā—Ƭ†–≥–ł–Ņ–Ķ—Ä–≥–Ķ–ĺ–ľ–Ķ—ā—Ä–ł—á–Ķ—Ā–ļ–ĺ–Ķ —Ä–į—Ā–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–ł–Ķ. –†–į—Ā–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–ł–Ķ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤ –Ņ–嬆–Ņ—Ä–ł–ĺ—Ä–ł—ā–Ķ—ā–į–ľ –Ņ—Ä–ĺ–≤–ĺ–ī–ł–Ľ–ĺ—Ā—Ć –Ņ—Ä–ĺ–≥—Ä–į–ľ–ľ–ĺ–Ļ. –ß–Ķ–ľ –ľ–Ķ–Ĺ—Ć—ą–Ķ –∑–Ĺ–į—á–Ķ–Ĺ–ł–Ķ p, —ā–Ķ–ľ –Ī–ĺ–Ľ—Ć—ą–Ķ –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ—Ā—ā—Ć, —á—ā–嬆–≥–Ķ–Ĺ—č, –Ņ–ĺ–Ņ–į–≤—ą–ł–Ķ –≤¬†–ļ–ĺ–Ĺ–ļ—Ä–Ķ—ā–Ĺ—č–Ļ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā, –ĺ—ā–Ĺ–Ķ—Ā–Ķ–Ĺ—č –ļ¬†–Ĺ–Ķ–ľ—É –Ĺ–Ķ¬†—Ā–Ľ—É—á–į–Ļ–Ĺ–ĺ. –†–Ķ–∑—É–Ľ—Ć—ā–į—ā —Ā—á–ł—ā–į–Ľ—Ā—Ź –∑–Ĺ–į—á–ł–ľ—č–ľ –Ņ—Ä–ł p = 0,01. –°¬†—É–≤–Ķ–Ľ–ł—á–Ķ–Ĺ–ł–Ķ–ľ¬†–≥–Ķ–Ĺ–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ā–ĺ–≤–ĺ–ļ—É–Ņ–Ĺ–ĺ—Ā—ā–ł –∑–Ĺ–į—á–Ķ–Ĺ–ł–Ķ p —É–ľ–Ķ–Ĺ—Ć—ą–į–Ķ—ā—Ā—Ź, —ā–ĺ –Ķ—Ā—ā—Ć –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ—Ā—ā—Ć —ā–ĺ–≥–ĺ, —á—ā–ĺ –≤¬†–Ņ—Ä–ĺ–ł–∑–≤–ĺ–Ľ—Ć–Ĺ–ĺ–Ļ –≤—č–Ī–ĺ—Ä–ļ–Ķ –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–ĺ –ĺ—ā–ľ–Ķ—á–Ķ–Ĺ–Ĺ—č—Ö –ĺ–Ī—ä–Ķ–ļ—ā–ĺ–≤ –Ĺ–Ķ¬†—Ā–Ľ—É—á–į–Ļ–Ĺ–ĺ, –≤–ĺ–∑—Ä–į—Ā—ā–į–Ķ—ā.
–†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –ł¬†–ł—Ö –ĺ–Ī—Ā—É–∂–ī–Ķ–Ĺ–ł–Ķ
–Ě–į–ľ–ł –Ņ—Ä–ĺ–į–Ĺ–į–Ľ–ł–∑–ł—Ä–ĺ–≤–į–Ĺ —Ā–Ņ–ł—Ā–ĺ–ļ¬†–≥–Ķ–Ĺ–ĺ–≤ (–Ī–į–∑–į –ī–į–Ĺ–Ĺ—č—Ö ResNet¬ģ), –ł–ľ–Ķ—é—Č–ł—Ö –ĺ—ā–Ĺ–ĺ—ą–Ķ–Ĺ–ł–Ķ –ļ¬†–í1- –ł¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–į–ľ, —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł—Ź –ļ–ĺ—ā–ĺ—Ä—č—Ö –ī–ĺ—Ā—ā–ĺ–≤–Ķ—Ä–Ĺ–ĺ —Ä–Ķ–≥—É–Ľ–ł—Ä—É–Ķ—ā—Ā—Ź –≤–Ĺ—É—ā—Ä–ł–ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č–ľ–ł —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ—č–ľ–ł –ļ–į—Ā–ļ–į–ī–į–ľ–ł, –ĺ–Ņ–ł—Ā–į–Ĺ–Ĺ—č–ľ–ł –≤¬†–Ľ–ł—ā–Ķ—Ä–į—ā—É—Ä–Ķ.
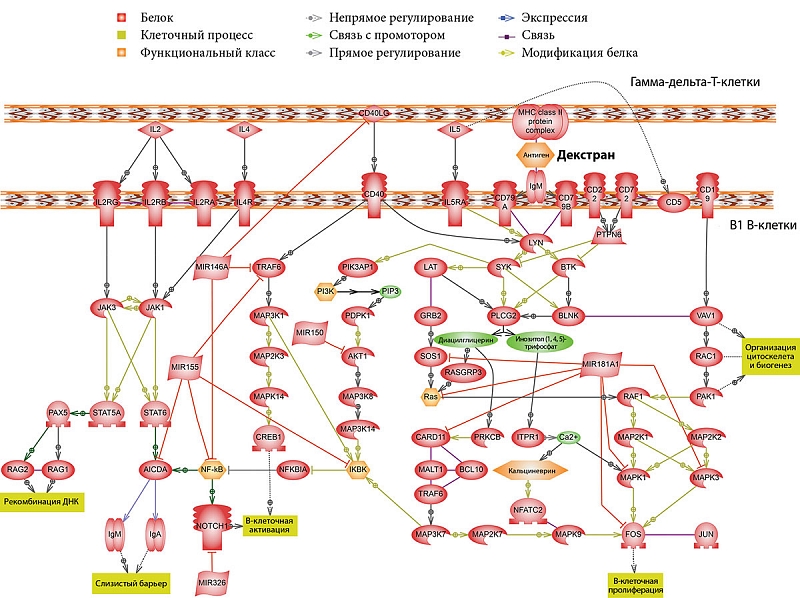
–†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–Ĺ–ĺ–Ļ —Ä–į–Ī–ĺ—ā—č –Ņ–ĺ–ī—ā–≤–Ķ—Ä–ī–ł–Ľ–ł, —á—ā–嬆–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–ł —É—á–į—Ā—ā–≤—É—é—ā –≤¬†—Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–ł—Ä–ĺ–≤–į–Ĺ–ł–ł –í1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤. –Ě–į¬†—Ä–ł—Ā—É–Ĺ–ļ–Ķ 1 –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ—č –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č–Ķ –Ņ—É—ā–ł –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź —ć—ā–ł—Ö —Ā—É–Ī–Ņ–ĺ–Ņ—É–Ľ—Ź—Ü–ł–Ļ.
–ě–ī–ł–Ĺ –ł–∑¬†—ā–į–ļ–ł—Ö —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ—č—Ö –Ņ—É—ā–Ķ–Ļ –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†–Ņ—Ä–ĺ–ī—É–ļ—Ü–ł–ł IgM. –ö–į–ļ –Ī—č–Ľ–ĺ –ĺ—ā–ľ–Ķ—á–Ķ–Ĺ–ĺ —Ä–į–Ĺ–Ķ–Ķ, B1-–ļ–Ľ–Ķ—ā–ļ–į–ľ –ī–Ľ—Ź –ĺ—ā–≤–Ķ—ā–į –Ĺ–į¬†–į–Ĺ—ā–ł–≥–Ķ–Ĺ –ł¬†—Ā–ł–Ĺ—ā–Ķ–∑–į/—Ā–Ķ–ļ—Ä–Ķ—Ü–ł–ł –į–Ĺ—ā–ł—ā–Ķ–Ľ –Ĺ–Ķ¬†—ā—Ä–Ķ–Ī—É–Ķ—ā—Ā—Ź –Ņ–ĺ–ľ–ĺ—Č—Ć –ļ–Ľ–į—Ā—Ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ. –Ę–Ě-2-–į–Ĺ—ā–ł–≥–Ķ–Ĺ –į–Ľ—Ć—Ą–į (1‚Üí3) –ī–Ķ–ļ—Ā—ā—Ä–į–Ĺ –Ī–Ľ–į–≥–ĺ–ī–į—Ä—Ź —Ā–≤–ĺ–Ķ–Ļ —Ā—ā—Ä—É–ļ—ā—É—Ä–Ķ —Ā¬†—Ä–Ķ–≥—É–Ľ—Ź—Ä–Ĺ–ĺ –Ņ–ĺ–≤—ā–ĺ—Ä—Ź—é—Č–ł–ľ–ł—Ā—Ź –ĺ–ī–ł–Ĺ–į–ļ–ĺ–≤—č–ľ–ł —ć–Ņ–ł—ā–ĺ–Ņ–į–ľ–ł –≤—č–∑—č–≤–į–Ķ—ā –Ņ–Ķ—Ä–Ķ–ļ—Ä–Ķ—Ā—ā–Ĺ–ĺ–Ķ —Ā–≤—Ź–∑—č–≤–į–Ĺ–ł–Ķ (–ļ–Ľ–į—Ā—ā–Ķ—Ä–ł–∑–į—Ü–ł—é) –Ĺ–Ķ—Ā–ļ–ĺ–Ľ—Ć–ļ–ł—Ö –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ļ—Ā–ĺ–≤ BCR –ł¬†—ā–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ –ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–ł–≤–į–Ķ—ā —Ā–ł–≥–Ĺ–į–Ľ, –ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ—č–Ļ –ī–Ľ—Ź –į–ļ—ā–ł–≤–į—Ü–ł–ł –í1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤. –ü–Ķ—Ä–≤—č–ľ–ł –ĺ–Ī—Ä–į–∑—É—é—ā—Ā—Ź IgM-–į–Ĺ—ā–ł—ā–Ķ–Ľ–į.
–í–į–∂–Ĺ—É—é —Ą—É–Ĺ–ļ—Ü–ł—é –≤¬†–Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–ł –ł–∑–ĺ—ā–ł–Ņ–ĺ–≤ –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤ –í-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł —Ā¬†IgM –Ĺ–į¬†–ī—Ä—É–≥–ł–Ķ –ļ–Ľ–į—Ā—Ā—č –≤—č–Ņ–ĺ–Ľ–Ĺ—Ź–Ķ—ā –ł–Ĺ–ī—É—Ü–ł—Ä–ĺ–≤–į–Ĺ–Ĺ–į—Ź –į–ļ—ā–ł–≤–į—Ü–ł–Ķ–Ļ —Ü–ł—ā–ł–ī–ł–Ĺ –ī–Ķ–∑–į–ľ–ł–Ĺ–į–∑–į (AID). AID —É—á–į—Ā—ā–≤—É–Ķ—ā –≤¬†—Ā–ĺ–ľ–į—ā–ł—á–Ķ—Ā–ļ–ĺ–ľ¬†–≥–ł–Ņ–Ķ—Ä–ľ—É—ā–ł—Ä–ĺ–≤–į–Ĺ–ł–ł (SHM), –ļ–ĺ–Ĺ–≤–Ķ—Ä—Ā–ł–ł¬†–≥–Ķ–Ĺ–ĺ–≤ –ł¬†—Ä–Ķ–ļ–ĺ–ľ–Ī–ł–Ĺ–į—Ü–ł–ł –Ņ—Ä–ł –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–ł –ł–∑–ĺ—ā–ł–Ņ–ĺ–≤ –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤ –≤¬†B-–ļ–Ľ–Ķ—ā–ļ–į—Ö –ł–ľ–ľ—É–Ĺ–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č. –í¬†–Ĺ–į—Ā—ā–ĺ—Ź—Č–Ķ–Ķ –≤—Ä–Ķ–ľ—Ź AID –Ņ—Ä–ł–∑–Ĺ–į–Ĺ–į –ĺ—Ā–Ĺ–ĺ–≤–Ĺ—č–ľ —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā–ĺ–ľ, –ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–ł–≤–į—é—Č–ł–ľ —Ä–į–∑–Ĺ–ĺ–ĺ–Ī—Ä–į–∑–ł–Ķ –ł–∑–ĺ—ā–ł–Ņ–ĺ–≤ –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤.
–£¬†–ľ—č—ą–Ķ–Ļ —Ā¬†–≤—Ä–ĺ–∂–ī–Ķ–Ĺ–Ĺ—č–ľ –ī–Ķ—Ą–ł—Ü–ł—ā–ĺ–ľ AID –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–ĺ –Ĺ–Ķ–Ņ–į—ā–ĺ–≥–Ķ–Ĺ–Ĺ—č—Ö –į–Ĺ–į—ć—Ä–ĺ–Ī–Ĺ—č—Ö –Ī–į–ļ—ā–Ķ—Ä–ł–Ļ –≤¬†—ā–ĺ–Ĺ–ļ–ĺ–Ļ –ļ–ł—ą–ļ–Ķ —É–≤–Ķ–Ľ–ł—á–Ķ–Ĺ–ĺ. –ö—Ä–ĺ–ľ–Ķ —ā–ĺ–≥–ĺ, —ɬ†–Ĺ–ł—Ö –ĺ–Ī—Ä–į–∑—É–Ķ—ā—Ā—Ź –ľ–Ĺ–ĺ–≥–ĺ –Ľ–ł–ľ—Ą–ĺ–ł–ī–Ĺ—č—Ö —Ą–ĺ–Ľ–Ľ–ł–ļ—É–Ľ—Ź—Ä–Ĺ—č—Ö —Ā—ā—Ä—É–ļ—ā—É—Ä, –≤—č—Ā—ā–ł–Ľ–į—é—Č–ł—Ö —ā–ĺ–Ĺ–ļ—É—é –ļ–ł—ą–ļ—É. –£¬†–Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ —Ā¬†–ł–ľ–ľ—É–Ĺ–ĺ–ī–Ķ—Ą–ł—Ü–ł—ā–ĺ–ľ –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–į –ļ–ł—ą–Ķ—á–Ĺ–į—Ź –ľ–ł–ļ—Ä–ĺ—Ą–Ľ–ĺ—Ä–į. –Ē–ł—Ā–Ī–į–Ľ–į–Ĺ—Ā –ľ–ł–ļ—Ä–ĺ—Ą–Ľ–ĺ—Ä—č –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į –ł¬†–ī—Ä—É–≥–ł—Ö —Ā–Ľ–ł–∑–ł—Ā—ā—č—Ö –ĺ–Ī–ĺ–Ľ–ĺ—á–Ķ–ļ —Ā–≤—Ź–∑–į–Ĺ —Ā¬†–Ī–Ľ–ĺ–ļ–ł—Ä–ĺ–≤–ļ–ĺ–Ļ AID-–ĺ–Ņ–ĺ—Ā—Ä–Ķ–ī–ĺ–≤–į–Ĺ–Ĺ–ĺ–Ļ —Ä–Ķ–ļ–ĺ–ľ–Ī–ł–Ĺ–į—Ü–ł–ł –Ņ—Ä–ł –Ņ–Ķ—Ä–Ķ–ļ–Ľ—é—á–Ķ–Ĺ–ł–ł –ł–∑–ĺ—ā–ł–Ņ–į (–ī–Ľ—Ź –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ–ł—Ź IgA) –≤¬†–í-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–į—Ö [10].
–ú–Ķ—Ö–į–Ĺ–ł–∑–ľ –ī–Ķ–Ļ—Ā—ā–≤–ł—Ź AID –∑–į–ļ–Ľ—é—á–į–Ķ—ā—Ā—Ź –≤¬†–∑–į–ľ–Ķ–Ĺ–Ķ —Ü–ł—ā–ł–ī–ł–Ĺ–į –Ĺ–į¬†—É—Ä–ł–ī–ł–Ĺ –≤¬†–ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–Ķ –Ē–Ě–ö, —á—ā–ĺ –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†—ā–ĺ—á–Ķ—á–Ĺ—č–ľ –ľ—É—ā–į—Ü–ł—Ź–ľ –ł¬†—Ä–Ķ–ļ–ĺ–ľ–Ī–ł–Ĺ–į—Ü–ł–ł¬†–≥–Ķ–Ĺ–ĺ–≤ –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤.
–†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –Ī–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–≥–ĺ –į–Ĺ–į–Ľ–ł–∑–į –Ņ—Ä–ĺ–ī–Ķ–ľ–ĺ–Ĺ—Ā—ā—Ä–ł—Ä–ĺ–≤–į–Ľ–ł, —á—ā–ĺ AID –ł–≥—Ä–į–Ķ—ā –Ī–ĺ–Ľ—Ć—ą—É—é —Ä–ĺ–Ľ—Ć –≤¬†—Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–ł—Ä–ĺ–≤–į–Ĺ–ł–ł –í1-–ļ–Ľ–Ķ—ā–ĺ–ļ.
–Ě–į–ľ–ł —É—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ĺ, —á—ā–ĺ —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł–ĺ–Ĺ–Ĺ—č–Ļ —Ą–į–ļ—ā–ĺ—Ä –ļ–į–Ņ–Ņ–į-–Ī–ł (NF-őļB) —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É–Ķ—ā –ł–Ĺ–ī—É–ļ—Ü–ł–ł AID –≤¬†–í1-–ļ–Ľ–Ķ—ā–ļ–į—Ö, –į¬†—ā–į–ļ–∂–Ķ –ł–Ĺ—ā–Ķ—Ä–Ľ–Ķ–Ļ–ļ–ł–Ĺ–ĺ–≤ (IL) 2 –ł¬†4 –Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–į–ľ–ł. IL-2 –≤–Ľ–ł—Ź–Ķ—ā –Ĺ–į¬†–ī–ł—Ą—Ą–Ķ—Ä–Ķ–Ĺ—Ü–ł—Ä–ĺ–≤–ļ—É –ł¬†–Ņ—Ä–ĺ–Ľ–ł—Ą–Ķ—Ä–į—Ü–ł—é –Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤, –≤—č—Ä–į–Ī–į—ā—č–≤–į–Ķ—ā—Ā—Ź –Ę-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł –≤¬†–ĺ—ā–≤–Ķ—ā –Ĺ–į¬†–į–Ĺ—ā–ł–≥–Ķ–Ĺ–Ĺ—É—é –ł¬†–ľ–ł—ā–ĺ–≥–Ķ–Ĺ–Ĺ—É—é —Ā—ā–ł–ľ—É–Ľ—Ź—Ü–ł—é. –Ē–Ľ—Ź –ĺ–Ī—Ä–į–∑–ĺ–≤–į–Ĺ–ł—Ź IL-2¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł —ā—Ä–Ķ–Ī—É–Ķ—ā—Ā—Ź —Ā—ā–ł–ľ—É–Ľ—Ź—Ü–ł—Ź —á–Ķ—Ä–Ķ–∑ CD28 [11]. –Ē—Ä—É–≥–ĺ–Ļ —Ü–ł—ā–ĺ–ļ–ł–Ĺ, –≤—č—Ä–į–Ī–į—ā—č–≤–į–Ķ–ľ—č–Ļ –Ę-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł,¬†‚Äď IL-4. –ě–Ĺ —Ä–Ķ–≥—É–Ľ–ł—Ä—É–Ķ—ā —Ä–ĺ—Ā—ā –ł¬†–ī–ł—Ą—Ą–Ķ—Ä–Ķ–Ĺ—Ü–ł—Ä–ĺ–≤–ļ—É –í-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤, –į¬†—ā–į–ļ–∂–Ķ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č –Ī–ł–ĺ—Ā–ł–Ĺ—ā–Ķ–∑–į –ł¬†—Ā–Ķ–ļ—Ä–Ķ—Ü–ł–ł –į–Ĺ—ā–ł—ā–Ķ–Ľ. –£—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ĺ, —á—ā–ĺ –Ķ–≥–ĺ –ľ–ĺ–≥—É—ā –≤—č—Ä–į–Ī–į—ā—č–≤–į—ā—Ƭ†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č, –≤¬†—á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, –Ņ—Ä–ł –ļ–ĺ–Ĺ—ā–į–ļ—ā–Ĺ—č—Ö –ī–Ķ—Ä–ľ–į—ā–ł—ā–į—Ö, –≤—č–∑–≤–į–Ĺ–Ĺ—č—Ö —Ā—ā–Ķ—Ä–ĺ–ł–ī–į–ľ–ł [12].
–Ę–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, –ī–į–Ĺ–Ĺ—č–Ķ, –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ–Ĺ—č–Ķ —Ā¬†–Ņ–ĺ–ľ–ĺ—Č—Ć—é –Ī–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–≥–ĺ –į–Ĺ–į–Ľ–ł–∑–į, –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź—é—ā —Ā–Ņ—Ä–ĺ–≥–Ĺ–ĺ–∑–ł—Ä–ĺ–≤–į—ā—Ć –≤–Ľ–ł—Ź–Ĺ–ł–Ķ¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ –Ĺ–į¬†B1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č –Ņ–ĺ—Ā—Ä–Ķ–ī—Ā—ā–≤–ĺ–ľ –≤—č—Ä–į–Ī–ĺ—ā–ļ–ł IL-2 –ł¬†-4.
–Ď—č–Ľ –≤—č—Ź–≤–Ľ–Ķ–Ĺ —ā–į–ļ–∂–Ķ —Ä—Ź–ī –ľ–į—Ä–ļ–Ķ—Ä–ĺ–≤, –≤–ĺ–≤–Ľ–Ķ—á–Ķ–Ĺ–Ĺ—č—Ö –≤¬†—Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–į–Ľ—Ć–Ĺ—É—é –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć –í1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤. –†–į—Ā–Ņ–ĺ–∑–Ĺ–į–≤–į–Ĺ–ł–Ķ IL-2 —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–į–ľ–ł (IL-2RG –ł¬†IL-2RB) –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†–į–ļ—ā–ł–≤–į—Ü–ł–ł —ā–ł—Ä–ĺ–∑–ł–Ĺ–ļ–ł–Ĺ–į–∑ JAK1 –ł¬†JAK3. –ź–ļ—ā–ł–≤–į—Ü–ł—Ź JAK1 –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā —ā–į–ļ–∂–Ķ –Ņ—Ä–ł –į–ļ—ā–ł–≤–į—Ü–ł–ł —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–į IL-4 (IL-4R). JAK1 –ł¬†JAK3 –ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–ł–≤–į—é—ā —Ą–ĺ—Ā—Ą–ĺ—Ä–ł–Ľ–ł—Ä–ĺ–≤–į–Ĺ–ł–Ķ STAT6 –ł¬†STAT5A, –∑–į —Ā—á–Ķ—ā —á–Ķ–≥–ĺ –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī–ł—ā –į–ļ—ā–ł–≤–į—Ü–ł—Ź AID –ł¬†RAG-–Ņ–ĺ–Ľ–ł–ľ–Ķ—Ä–į–∑—č (RAG1 –ł¬†RAG2) (RAG¬†‚Äst–≥–Ķ–Ĺ, –į–ļ—ā–ł–≤–ł—Ä—É—é—Č–ł–Ļ —Ä–Ķ–ļ–ĺ–ľ–Ī–ł–Ĺ–į—Ü–ł—é).
–Ę–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, –≤–ĺ–∑–ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ IL-2 –ł¬†-4 –Ĺ–į¬†B1-–ļ–Ľ–Ķ—ā–ļ–ł –ī–ĺ–Ľ–∂–Ĺ–ĺ –≤—č–∑—č–≤–į—ā—Ć —Ä–Ķ–ļ–ĺ–ľ–Ī–ł–Ĺ–į—Ü–ł—é V-–≥–Ķ–Ĺ–ĺ–≤ –ł–ľ–ľ—É–Ĺ–ĺ–≥–Ľ–ĺ–Ī—É–Ľ–ł–Ĺ–ĺ–≤. –†–į–Ĺ–Ķ–Ķ –Ī—č–Ľ–ĺ –Ņ–ĺ–ļ–į–∑–į–Ĺ–ĺ, —á—ā–ĺ STAT6 –ł¬†NF-kB –į—Ā—Ā–ĺ—Ü–ł–ł—Ä–ĺ–≤–į–Ĺ—č —Ā¬†–ł–Ĺ–ī—É–ļ—Ü–ł–Ķ–Ļ —Ä—Ź–ī–į¬†–≥–Ķ–Ĺ–ĺ–≤ [13‚Äď15] –ł¬†–ľ–Ķ–∂–ī—É —ć—ā–ł–ľ–ł —Ą–į–ļ—ā–ĺ—Ä–į–ľ–ł —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł–ł —Ā—É—Č–Ķ—Ā—ā–≤—É–Ķ—ā –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ [16]. –°–ł–Ĺ–Ķ—Ä–≥–ł—á–Ķ—Ā–ļ–ł–Ļ —ć—Ą—Ą–Ķ–ļ—ā STAT6 –ł¬†NF-kB, –Ņ–ĺ-–≤–ł–ī–ł–ľ–ĺ–ľ—É, –Ľ–Ķ–∂–ł—ā –≤¬†–ĺ—Ā–Ĺ–ĺ–≤–Ķ —Ā–≤—Ź–∑–ł –ľ–Ķ–∂–ī—É —Ä–į—Ā–Ņ–ĺ–∑–Ĺ–į–≤–į–Ĺ–ł–Ķ–ľ IL-4 –ł¬†CD40 –ł¬†–ł–Ĺ–ī—É–ļ—Ü–ł–Ķ–Ļ —ć–ļ—Ā–Ņ—Ä–Ķ—Ā—Ā–ł–ł¬†–≥–Ķ–Ĺ–į AID [17].
–ē—Č–Ķ –ĺ–ī–ł–Ĺ —Ą–į–ļ—ā–ĺ—Ä, –ļ–ĺ—ā–ĺ—Ä—č–Ļ –Ī—č–Ľ –≤—č—Ź–≤–Ľ–Ķ–Ĺ –≤¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –Ī–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–≥–ĺ –į–Ĺ–į–Ľ–ł–∑–į,¬†‚Äď —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä CD40. –ü–ĺ—Ā–Ľ–Ķ–ī–Ĺ–ł–Ļ –Ņ—Ä–ł –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł–ł —Ā¬†CD40L (CD154) —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ķ–Ĺ –≤—č–∑—č–≤–į—ā—Ć –į–ļ—ā–ł–≤–į—Ü–ł—é –ł¬†–Ņ—Ä–ĺ–Ľ–ł—Ą–Ķ—Ä–į—Ü–ł—é B-–ļ–Ľ–Ķ—ā–ĺ–ļ. –°–Ľ–Ķ–ī—É–Ķ—ā –ĺ—ā–ľ–Ķ—ā–ł—ā—Ć, —á—ā–ĺ –Ľ–ł–≥–į–Ĺ–ī CD40¬†‚Äď CD40L —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź —Ö–į—Ä–į–ļ—ā–Ķ—Ä–Ĺ—č–ľ –ľ–į—Ä–ļ–Ķ—Ä–ĺ–ľ –ī–Ľ—Ź CD4+-–Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ –ł¬†–Ĺ–Ķ—ā–ł–Ņ–ł—á–Ĺ—č–ľ¬†‚Äď –ī–Ľ—Ź¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ĺ–ļ. –ě–ī–Ĺ–į–ļ–ĺ –≤¬†–Ľ–ł—ā–Ķ—Ä–į—ā—É—Ä–Ķ —É–Ņ–ĺ–ľ–ł–Ĺ–į–Ķ—ā—Ā—Ź –嬆CD4+-–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–į—Ö [18] –ł¬†–ĺ–Ī —ć–ļ—Ā–Ņ—Ä–Ķ—Ā—Ā–ł–ł CD40L –Ĺ–į¬†–į–ļ—ā–ł–≤–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č—Ö¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–į—Ö [19].
–ď–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–ł –ľ–ĺ–≥—É—ā –≤—č—Ā—ā—É–Ņ–į—ā—Ć –ļ–į–ļ –ź–ü–ö –ī–Ľ—Ź CD4+-T-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤ [20], —á—ā–ĺ –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź–Ķ—ā –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–ł—ā—Ć –ļ–ĺ—Ā–≤–Ķ–Ĺ–Ĺ–ĺ–Ķ –ł—Ö –≤–Ľ–ł—Ź–Ĺ–ł–Ķ –Ĺ–į¬†–ł–ľ–ľ—É–Ĺ–Ĺ—č–Ļ –ĺ—ā–≤–Ķ—ā.
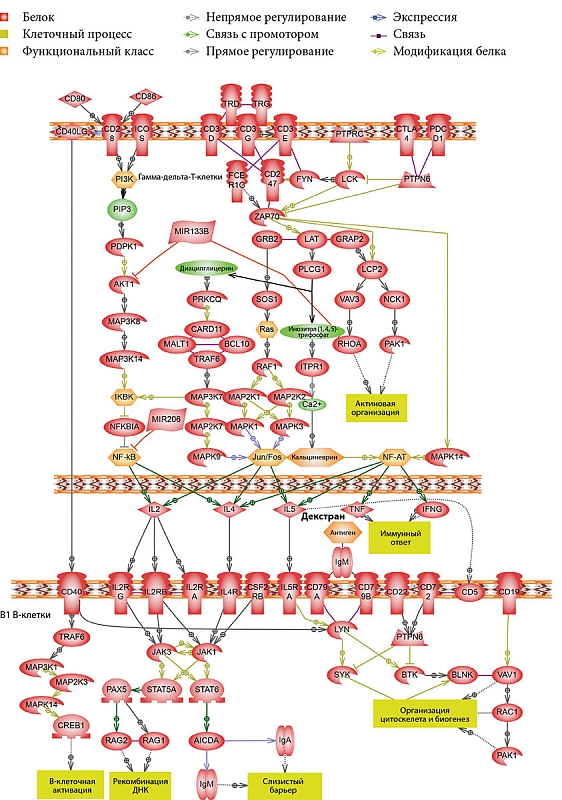
–ü–嬆–ľ–Ķ—Ä–Ķ —Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź B1-–ļ–Ľ–Ķ—ā–ļ–ł –ľ—č—ą–ł –Ņ–Ķ—Ä–Ķ—Ā—ā–į—é—ā —ć–ļ—Ā–Ņ—Ä–Ķ—Ā—Ā–ł—Ä–ĺ–≤–į—ā—Ć –≤¬†–Ī–ĺ–Ľ—Ć—ą–ł—Ö –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–į—Ö –ļ–ĺ—Ā—ā–ł–ľ—É–Ľ–ł—Ä—É—é—Č—É—é –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ—É CD86 –ł¬†—Ā—ā–į–Ĺ–ĺ–≤—Ź—ā—Ā—Ź –ľ–ĺ—Č–Ĺ—č–ľ–ł –į–ļ—ā–ł–≤–į—ā–ĺ—Ä–į–ľ–ł CD8+-T-–ļ–Ľ–Ķ—ā–ĺ–ļ [21], –į¬†—ā–į–ļ –ļ–į–ļ —á–į—Ā—ā—Ƭ†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤ –≤¬†–ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–Ķ —Ź–≤–Ľ—Ź—é—ā—Ā—Ź CD8+, –Ĺ–Ķ–Ľ—Ć–∑—Ź –ł—Ā–ļ–Ľ—é—á–į—ā—Ć –≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ—Ā—ā—Ć –Ņ—Ä—Ź–ľ–ĺ–≥–ĺ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź B1-–ļ–Ľ–Ķ—ā–ĺ–ļ —Ā¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł (—Ä–ł—Ā.¬†2).
–í¬†—Ä–į–∑–Ĺ—č—Ö –ł—Ā—ā–ĺ—á–Ĺ–ł–ļ–į—Ö –≤—Ā—ā—Ä–Ķ—á–į—é—ā—Ā—Ź –ī–į–Ĺ–Ĺ—č–Ķ –嬆—ā–ĺ–ľ, —á—ā–ĺ –í1-–ļ–Ľ–Ķ—ā–ļ–ł –ľ–ĺ–≥—É—ā –Ī—č—ā—Ć –ł¬†–Ę–ó. B B1-–ļ–Ľ–Ķ—ā–ļ–į—Ö –ĺ—ā—Ā—É—ā—Ā—ā–≤—É—é—ā —Ā—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ—č–Ķ –ī–Ķ—Ą–Ķ–ļ—ā—č, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –Ĺ–Ķ¬†–Ņ–ĺ–∑–≤–ĺ–Ľ—Ź–Ľ–ł –Ī—č –ĺ—ā–≤–Ķ—á–į—ā—Ć –Ĺ–į¬†–Ę-–ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č–Ķ —Ā—ā–ł–ľ—É–Ľ—č, –≤–ļ–Ľ—é—á–į—Ź CD40L, IL-4 –ł¬†-5 [22]. IL-5 —ā–į–ļ–∂–Ķ —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ķ–Ĺ —Ā—ā–ł–ľ—É–Ľ–ł—Ä–ĺ–≤–į—ā—Ć –Ņ—Ä–ĺ–Ľ–ł—Ą–Ķ—Ä–į—Ü–ł—é B1-–ļ–Ľ–Ķ—ā–ĺ–ļ –≤¬†–ĺ—ā—Ā—É—ā—Ā—ā–≤–ł–Ķ CD40¬†‚Äď CD40L –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź [23].
–ě–Ņ–ł—Ä–į—Ź—Ā—Ć –Ĺ–į¬†–į–Ĺ–į–Ľ–ł–∑ –Ľ–ł—ā–Ķ—Ä–į—ā—É—Ä—č –ł¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č, –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ–Ĺ—č–Ķ —Ā¬†–ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–ł–Ķ–ľ –Ī–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–≥–ĺ –į–Ĺ–į–Ľ–ł–∑–į, –ľ–ĺ–∂–Ĺ–ĺ –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–ł—ā—Ć, —á—ā–ĺ –Ņ–ĺ–ī–ĺ–Ī–Ĺ–ĺ–Ķ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ –ľ–Ķ–∂–ī—É –í1- –ł¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–į–ľ–ł –≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ.
–£—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ĺ, —á—ā–ĺ —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł—Ź –Ĺ–į¬†—É—Ä–ĺ–≤–Ĺ–Ķ miRNA –ł¬†–ī—Ä. –ļ—Ä–ł—ā–ł—á–Ĺ–į –ī–Ľ—Ź —Ä–į–∑–≤–ł—ā–ł—Ź –ł¬†–Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –ł–ľ–ľ—É–Ĺ–Ĺ–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č. –ü–ĺ–∑–ī–Ĺ–Ķ–Ķ –Ī—č–Ľ –≤—č—Ź–≤–Ľ–Ķ–Ĺ —Ä—Ź–ī miRNA, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ľ–ĺ–≥—É—ā —É—á–į—Ā—ā–≤–ĺ–≤–į—ā—Ć –Ĺ–į¬†—Ä–į–∑–Ĺ—č—Ö —Ā—ā–į–ī–ł—Ź—Ö¬†–≥–Ķ–ľ–ĺ–Ņ–ĺ—ć–∑–į [24] –ł¬†–≤–Ĺ–ĺ—Ā–ł—ā—Ć –≤–ļ–Ľ–į–ī –≤¬†–ĺ–ļ–ĺ–Ĺ—á–į—ā–Ķ–Ľ—Ć–Ĺ—É—é –ī–ł—Ą—Ą–Ķ—Ä–Ķ–Ĺ—Ü–ł—Ä–ĺ–≤–ļ—É –≤–∑—Ä–ĺ—Ā–Ľ—č—Ö –Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤ [25].
–í¬†–Ī–ĺ–Ľ—Ć—ą–ł–Ĺ—Ā—ā–≤–Ķ —Ā–Ľ—É—á–į–Ķ–≤ miRNA –≤—č—Ā—ā—É–Ņ–į—é—ā –≤¬†—Ä–ĺ–Ľ–ł –ł–Ĺ–≥–ł–Ī–ł—ā–ĺ—Ä–ĺ–≤ –≤–Ĺ—É—ā—Ä–ł–ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č—Ö –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤. –Ę–į–ļ, –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–Ĺ—č–Ļ –Ĺ–į–ľ–ł –į–Ĺ–į–Ľ–ł–∑ –Ņ–ĺ–∑–≤–ĺ–Ľ–ł–Ľ –Ņ–ĺ–Ľ—É—á–ł—ā—Ć –ī–į–Ĺ–Ĺ—č–Ķ –嬆–≤–Ľ–ł—Ź–Ĺ–ł–ł miRNA155 –ł¬†miRNA146a –Ĺ–į¬†AID, –Ņ—Ä–ł–≤–ĺ–ī—Ź—Č–Ķ–ľ –ļ¬†–Ķ–Ķ —É–≥–Ĺ–Ķ—ā–Ķ–Ĺ–ł—é –ļ–į–ļ –Ĺ–į–Ņ—Ä—Ź–ľ—É—é, —ā–į–ļ –ł¬†–ĺ–Ņ–ĺ—Ā—Ä–Ķ–ī–ĺ–≤–į–Ĺ–Ĺ–ĺ —á–Ķ—Ä–Ķ–∑ –Ņ–ĺ–ī–į–≤–Ľ–Ķ–Ĺ–ł–Ķ NF-kB [26]. –†–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä Notch, —Ä–Ķ–≥—É–Ľ–ł—Ä—É—é—Č–ł–Ļ –ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—É—é –ī–ł—Ą—Ą–Ķ—Ä–Ķ–Ĺ—Ü–ł–į—Ü–ł—é, —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź —ā–ł–Ņ–ł—á–Ĺ–ĺ–Ļ –ľ–ł—ą–Ķ–Ĺ—Ć—é –ī–Ľ—Ź miRNA326 [27]. –í¬†–ī–ł—Ą—Ą–Ķ—Ä–Ķ–Ĺ—Ü–ł–į—Ü–ł–ł B-–ļ–Ľ–Ķ—ā–ĺ–ļ –Ĺ–Ķ–ľ–į–Ľ–ĺ–≤–į–∂–Ĺ—É—é —Ä–ĺ–Ľ—Ć –ł–≥—Ä–į–Ķ—ā miRNA181A [27].
–ė—Ā–Ņ–ĺ–Ľ—Ć–∑—É–Ķ–ľ—č–Ļ –Ĺ–į–ľ–ł –ľ–Ķ—ā–ĺ–ī –Ņ–ĺ–∑–≤–ĺ–Ľ–ł–Ľ –Ņ—Ä–ĺ–į–Ĺ–į–Ľ–ł–∑–ł—Ä–ĺ–≤–į—ā—Ć –Ĺ–Ķ—Ā–ļ–ĺ–Ľ—Ć–ļ–ĺ –≤–ł–ī–ĺ–≤ miRNA, –į¬†—ā–į–ļ–∂–Ķ —É—Ā—ā–į–Ĺ–ĺ–≤–ł—ā—Ć –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č–Ķ —ā–ĺ—á–ļ–ł –ł—Ö –≤–ĺ–∑–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź –Ĺ–į¬†–ĺ—Ā–Ĺ–ĺ–≤–Ĺ—č–Ķ –≤–Ĺ—É—ā—Ä–ł–ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č–Ķ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č.
–ě—Ü–Ķ–Ĺ–ļ–į –≤–ļ–Ľ–į–ī–į –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö miRNA –≤¬†—ć—ā–ł –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č –ľ–ĺ–∂–Ķ—ā —Ā–Ľ—É–∂–ł—ā—Ć –į–Ľ—Ć—ā–Ķ—Ä–Ĺ–į—ā–ł–≤–Ĺ—č–ľ –Ņ–ĺ–ī—Ö–ĺ–ī–ĺ–ľ –ī–Ľ—Ź –į–Ĺ–į–Ľ–ł–∑–į –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź B1- –ł¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤.
–Ē–į–Ĺ–Ĺ–ĺ–Ķ –Ĺ–į—É—á–Ĺ–ĺ–Ķ –Ĺ–į–Ņ—Ä–į–≤–Ľ–Ķ–Ĺ–ł–Ķ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ—Ź–Ķ—ā –ł–Ĺ—ā–Ķ—Ä–Ķ—Ā –Ĺ–Ķ¬†—ā–ĺ–Ľ—Ć–ļ–ĺ –≤¬†–ĺ–Ī–Ľ–į—Ā—ā–ł —Ą—É–Ĺ–ī–į–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł–ľ–ľ—É–Ĺ–ĺ–Ľ–ĺ–≥–ł–ł. –ė–∑–≤–Ķ—Ā—ā–Ĺ–ĺ, —á—ā–ĺ B1- –ł¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–ł —É—á–į—Ā—ā–≤—É—é—ā –≤¬†—Ä–į–∑–≤–ł—ā–ł–ł —Ä—Ź–ī–į –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ. –Ę–į–ļ, –í1-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā—č –∑–į–ī–Ķ–Ļ—Ā—ā–≤–ĺ–≤–į–Ĺ—č –≤¬†—Ä–į–∑–≤–ł—ā–ł–ł Th2-–ĺ–Ņ–ĺ—Ā—Ä–Ķ–ī–ĺ–≤–į–Ĺ–Ĺ–ĺ–≥–ĺ –ļ–ĺ–Ľ–ł—ā–į [28], –į¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–ł –ľ–ĺ–≥—É—ā –ł–≥—Ä–į—ā—Ć –Ņ—Ä–ĺ—ā–Ķ–ļ—ā–ł–≤–Ĺ—É—é —Ä–ĺ–Ľ—Ć –ī–Ľ—Ź —ć–Ņ–ł—ā–Ķ–Ľ–ł–į–Ľ—Ć–Ĺ—č—Ö –ļ–Ľ–Ķ—ā–ĺ–ļ –ł¬†—Ā–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ĺ–≤–į—ā—Ć —Ä–į–∑–≤–ł—ā–ł—é –≤–ĺ—Ā–Ņ–į–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤ [29]. –ė—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–ĺ–≤ –į–ļ—ā–ł–≤–į—Ü–ł–ł –ł–Ľ–ł —Ā—É–Ņ—Ä–Ķ—Ā—Ā–ł–ł –≤–ĺ—Ā–Ņ–į–Ľ–Ķ–Ĺ–ł—Ź –≤¬†–ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–Ķ –Ņ–ĺ–∑–≤–ĺ–Ľ–ł—ā –≤¬†–Ņ–Ķ—Ä—Ā–Ņ–Ķ–ļ—ā–ł–≤–Ķ –≤—č—Ź–≤–ł—ā—Ć –ľ–ł—ą–Ķ–Ĺ–ł –ī–Ľ—Ź —ā–Ķ—Ä–į–Ņ–Ķ–≤—ā–ł—á–Ķ—Ā–ļ–ĺ–≥–ĺ –≤–ĺ–∑–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź –ł¬†—Ä–Ķ–į–Ľ–ł–∑–ĺ–≤–į—ā—Ć –Ņ–Ķ—Ä—Ā–ĺ–Ĺ–į–Ľ–ł–∑–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č–Ļ –Ņ–ĺ–ī—Ö–ĺ–ī.
–ü–ĺ–Ľ—É—á–Ķ–Ĺ–Ĺ—č–Ķ –≤¬†–ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–ł –ī–į–Ĺ–Ĺ—č–Ķ —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤—É—é—ā, —á—ā–ĺ B1- –ł¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–ļ–Ľ–Ķ—ā–ļ–ł –ľ–ĺ–≥—É—ā –≤—č—Ā—ā—É–Ņ–į—ā—Ć –≤¬†–ļ–į—á–Ķ—Ā—ā–≤–Ķ –Ĺ–ĺ–≤—č—Ö —ā–Ķ—Ä–į–Ņ–Ķ–≤—ā–ł—á–Ķ—Ā–ļ–ł—Ö –ľ–ł—ą–Ķ–Ĺ–Ķ–Ļ.
–Ę–Ķ—Ä–į–Ņ–ł—Ź –ľ–Ĺ–ĺ–≥–ł—Ö –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ (–≤¬†—á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, –≤–ĺ—Ā–Ņ–į–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –ł¬†–ł–Ĺ—Ą–Ķ–ļ—Ü–ł–ĺ–Ĺ–Ĺ—č—Ö) —ā–Ķ—Ā–Ĺ–ĺ —Ā–≤—Ź–∑–į–Ĺ–į —Ā¬†–ľ–ĺ–Ľ–Ķ–ļ—É–Ľ—Ź—Ä–Ĺ—č–ľ–ł —ā–Ķ—Ö–Ĺ–ĺ–Ľ–ĺ–≥–ł—Ź–ľ–ł. –ü–ĺ–ī—Ö–ĺ–ī—č –ļ¬†miRNA-–ĺ–Ņ–ĺ—Ā—Ä–Ķ–ī–ĺ–≤–į–Ĺ–Ĺ–ĺ–Ļ —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł –ļ–Ľ–Ķ—ā–ĺ–ļ –∑–į–Ĺ–ł–ľ–į—é—ā –≤¬†–Ĺ–Ķ–Ļ –Ĺ–Ķ¬†–Ņ–ĺ—Ā–Ľ–Ķ–ī–Ĺ–Ķ–Ķ –ľ–Ķ—Ā—ā–ĺ. –ě–Ĺ–ł –Ņ–ĺ—ā–Ķ–Ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ –ľ–ĺ–≥—É—ā –Ī—č—ā—Ć –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ—č –ī–Ľ—Ź —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł –≤–ĺ—Ā–Ņ–į–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤ –≤¬†–ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–Ķ.
–ě–ī–Ĺ–į–ļ–ĺ –ĺ—Ā—ā–į–Ķ—ā—Ā—Ź —Ä—Ź–ī –≤–ĺ–Ņ—Ä–ĺ—Ā–ĺ–≤: —Ā–ĺ–∑–ī–į–Ĺ–ł–Ķ –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ļ—Ā–Ĺ–ĺ–≥–ĺ –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–į –ł–Ľ–ł –ľ–ĺ–Ĺ–ĺ–Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –Ī—É–ī—É—ā –ľ–Ķ–Ĺ—Ź—ā—Ć —Ä—Ź–ī –ļ–Ľ–Ķ—ā–ĺ–ļ –ł–Ľ–ł –Ī—É–ī—É—ā –Ĺ–į–Ņ—Ä–į–≤–Ľ–Ķ–Ĺ—č —ā–ĺ–Ľ—Ć–ļ–ĺ –Ĺ–į¬†–ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ—č–Ļ —ć—Ą—Ą–Ķ–ļ—ā, –į¬†—ā–į–ļ–∂–Ķ –ī–ĺ—Ā—ā–į–≤–ļ—É –Ĺ—É–ļ–Ľ–Ķ–ł–Ĺ–ĺ–≤—č—Ö –ļ–ł—Ā–Ľ–ĺ—ā (—Ä–Ķ–≥—É–Ľ—Ź—ā–ĺ—Ä–Ĺ—č—Ö –†–Ě–ö).
–ó–į–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ
–Ď–ł–ĺ–ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ĺ–Ĺ–Ĺ—č–Ļ –Ņ–ĺ–ī—Ö–ĺ–ī, –ł—Ā–Ņ–ĺ–Ľ—Ć–∑—É–Ķ–ľ—č–Ļ –Ĺ–į–ľ–ł –ī–Ľ—Ź –Ņ—Ä–ĺ–≥–Ĺ–ĺ–∑–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –Ņ—É—ā–Ķ–Ļ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź B1- –ł¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–ĺ–≤, –Ņ–ĺ–∑–≤–ĺ–Ľ–ł–Ľ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–ł—ā—Ć –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ—č–Ķ –ł–∑¬†–Ĺ–ł—Ö –ł¬†–ĺ—Ā–Ĺ–ĺ–≤–Ĺ—č–Ķ —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ—č–Ķ –ļ–į—Ā–ļ–į–ī—č, —É—á–į—Ā—ā–≤—É—é—Č–ł–Ķ –≤¬†—ā–į–ļ–ĺ–ľ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł–ł. –ö—Ä–ĺ–ľ–Ķ —ā–ĺ–≥–ĺ, –ľ–Ķ—ā–ĺ–ī –ī–į–Ľ –≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ—Ā—ā—Ć —Ā–Ņ—Ä–ĺ–≥–Ĺ–ĺ–∑–ł—Ä–ĺ–≤–į—ā—Ć —É—á–į—Ā—ā–ł–Ķ –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö –≤–ł–ī–ĺ–≤ miRNA –≤¬†—ć—ā–ł—Ö –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–į—Ö.
–Ē–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–Ķ –ł–∑—É—á–Ķ–Ĺ–ł–Ķ —Ā–≤—Ź–∑–ł –ľ–Ķ–∂–ī—É B1- –ł¬†–≥–į–ľ–ľ–į-–ī–Ķ–Ľ—Ć—ā–į-–Ę-–Ľ–ł–ľ—Ą–ĺ—Ü–ł—ā–į–ľ–ł –Ņ–ĺ–∑–≤–ĺ–Ľ–ł—ā –Ņ–ĺ-–Ĺ–ĺ–≤–ĺ–ľ—É –≤–∑–≥–Ľ—Ź–Ĺ—É—ā—Ć –Ĺ–į¬†–Ņ–į—ā–ĺ–≥–Ķ–Ĺ–Ķ–∑ —Ä—Ź–ī–į –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ, –≤¬†–ļ–ĺ—ā–ĺ—Ä—č—Ö —É—á–į—Ā—ā–≤—É—é—ā —Ā—É–Ī–Ņ–ĺ–Ņ—É–Ľ—Ź—Ü–ł–ł —ć—ā–ł—Ö –ļ–Ľ–Ķ—ā–ĺ–ļ.
O.A. Svitich, N.A. Snegireva, V.V. Sobolev, I.N. Dyakov, I.M. Korsunskaya, V.V. Zverev
I.I. Mechnikov Research Institute of Vaccines and Sera
Center for Theoretical Problems of Physicochemical Pharmacology
Contact person: Oksana Anatolyevna Svitich, svitichoa@yandex.ru
B-1 lymphocytes represent a separate subpopulation of B cells in the human body and mice, and participate in the first line of defense against infection. They are able to respond to T-independent type 2 antigens (without the involvement of T cells). It is known that B-1 lymphocytes are localized primarily in the serous cavities (abdominal and pleural) and in the lymphoid tissue of the mucous membranes. In the intestinal mucosa B-1 cells produce a significant portion of IgA. Usually, signals from T lymphocytes are required to switch immunoglobulin isotypes; however, it is known that the isotype is switched from IgM to IgA in T-independent B-1 cells which do not receive classical Th-aid in the small intestine. It is assumed that ő≥őī–Ę cells can participate in this process, which, like B-1 lymphocytes, appear earlier than other T cells in ontogenesis and have the localization in the body similar to B-1 cells. The identification of the mechanisms of B-1 and ő≥őīT lymphocytes interaction in the process of immune response to TI-2 antigens is an urgent task. To predict possible ways of such interaction bioinformatics analysis was used. As a result, data about cell markers and key signaling pathways that can participate in this interaction were obtained. Also, data have been obtained about miRNA, regulatory intracellular structures, which can affect individual components of the signaling pathways.
–£–≤–į–∂–į–Ķ–ľ—č–Ļ –Ņ–ĺ—Ā–Ķ—ā–ł—ā–Ķ–Ľ—Ć uMEDp!
–£–≤–Ķ–ī–ĺ–ľ–Ľ—Ź–Ķ–ľ –í–į—Ā –ĺ —ā–ĺ–ľ, —á—ā–ĺ –∑–ī–Ķ—Ā—Ć —Ā–ĺ–ī–Ķ—Ä–∂–ł—ā—Ā—Ź –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł—Ź, –Ņ—Ä–Ķ–ī–Ĺ–į–∑–Ĺ–į—á–Ķ–Ĺ–Ĺ–į—Ź –ł—Ā–ļ–Ľ—é—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –ī–Ľ—Ź —Ā–Ņ–Ķ—Ü–ł–į–Ľ–ł—Ā—ā–ĺ–≤ –∑–ī—Ä–į–≤–ĺ–ĺ—Ö—Ä–į–Ĺ–Ķ–Ĺ–ł—Ź.
–ē—Ā–Ľ–ł –í—č –Ĺ–Ķ —Ź–≤–Ľ—Ź–Ķ—ā–Ķ—Ā—Ć —Ā–Ņ–Ķ—Ü–ł–į–Ľ–ł—Ā—ā–ĺ–ľ –∑–ī—Ä–į–≤–ĺ–ĺ—Ö—Ä–į–Ĺ–Ķ–Ĺ–ł—Ź, –į–ī–ľ–ł–Ĺ–ł—Ā—ā—Ä–į—Ü–ł—Ź –Ĺ–Ķ –Ĺ–Ķ—Ā–Ķ—ā –ĺ—ā–≤–Ķ—ā—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ—Ā—ā–ł –∑–į –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č–Ķ –ĺ—ā—Ä–ł—Ü–į—ā–Ķ–Ľ—Ć–Ĺ—č–Ķ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź, –≤–ĺ–∑–Ĺ–ł–ļ—ą–ł–Ķ –≤ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ —Ā–į–ľ–ĺ—Ā—ā–ĺ—Ź—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–ł—Ź –í–į–ľ–ł –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł —Ā –Ņ–ĺ—Ä—ā–į–Ľ–į –Ī–Ķ–∑ –Ņ—Ä–Ķ–ī–≤–į—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –ļ–ĺ–Ĺ—Ā—É–Ľ—Ć—ā–į—Ü–ł–ł —Ā –≤—Ä–į—á–ĺ–ľ.
–Ě–į–∂–ł–ľ–į—Ź –Ĺ–į –ļ–Ĺ–ĺ–Ņ–ļ—É ¬ę–í–ĺ–Ļ—ā–ł¬Ľ, –í—č –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–į–Ķ—ā–Ķ, —á—ā–ĺ —Ź–≤–Ľ—Ź–Ķ—ā–Ķ—Ā—Ć –≤—Ä–į—á–ĺ–ľ –ł–Ľ–ł —Ā—ā—É–ī–Ķ–Ĺ—ā–ĺ–ľ –ľ–Ķ–ī–ł—Ü–ł–Ĺ—Ā–ļ–ĺ–≥–ĺ –≤—É–∑–į.