–≠–≤–Њ–ї—О—Ж–Є—П —Б–љ–∞ –Њ—В –њ—А–Њ—Б—В—Л—Е —Д–Њ—А–Љ –і–Њ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е
- –Р–љ–љ–Њ—В–∞—Ж–Є—П
- –°—В–∞—В—М—П
- –°—Б—Л–ї–Ї–Є
- English

–£ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж —Б–Њ–љ —Б–Њ—Б—В–Њ–Є—В –Є–Ј¬†–і–≤—Г—Е –њ–Њ–і—Б—В–∞–і–Є–є¬†вАУ –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–≥–Њ —Б–љ–∞ (–Ь–°) –Є¬†–њ–∞—А–∞–і–Њ–Ї—Б–∞–ї—М–љ–Њ–≥–Њ —Б–љ–∞ (–Я–°) (–Є–ї–Є –±—Л—Б—В—А–Њ–≥–Њ —Б–љ–∞, –Є–ї–Є —Б—В–∞–і–Є–Є –±—Л—Б—В—А—Л—Е –і–≤–Є–ґ–µ–љ–Є–є¬†–≥–ї–∞–Ј). –†–∞–Ј–љ—Л–µ –≤–Є–і—Л –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –њ—А–Њ–≤–Њ–і—П—В –≤–Њ —Б–љ–µ –Њ—В¬†–і–≤—Г—Е –і–Њ 20¬†—З–∞—Б–Њ–≤ –≤¬†—Б—Г—В–Ї–Є, –∞¬†–њ—В–Є—Ж—Л¬†вАУ –Њ—В¬†–њ—П—В–Є –і–Њ 17¬†—З–∞—Б–Њ–≤. –Я–Њ–њ—Л—В–Ї–Є —Г—Б—В–∞–љ–Њ–≤–Є—В—М –Ї–Њ—А—А–µ–ї—П—Ж–Є–Є –њ–∞—А–∞–Љ–µ—В—А–Њ–≤ —Б–љ–∞ —Б¬†–∞–љ–∞—В–Њ–Љ–Њ-—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ–Є —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–∞–Љ–Є –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж, –∞¬†—В–∞–Ї–ґ–µ —Б¬†—Г—Б–ї–Њ–≤–Є—П–Љ–Є —Б–љ–∞ –і–Њ —Б–Є—Е –њ–Њ—А –±—Л–ї–Є –љ–µ¬†–Њ—З–µ–љ—М —Г—Б–њ–µ—И–љ—Л–Љ–Є [1вАУ6]. –Т–Њ –Љ–љ–Њ–≥–Њ–Љ —Н—В–Њ –Њ–њ—А–µ–і–µ–ї—П–µ—В—Б—П —В–µ–Љ, —З—В–Њ –і–∞–љ–љ—Л–µ –Њ¬†–њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Б–љ–∞ –Є¬†–µ–≥–Њ —Б—В–∞–і–Є–є –њ–Њ–ї—Г—З–µ–љ—Л –≤¬†–Њ—Б–љ–Њ–≤–љ–Њ–Љ –≤¬†–ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е. –Ю–љ–Є –љ–µ¬†—Г—З–Є—В—Л–≤–∞—О—В –≤—Б–µ –Љ–љ–Њ–≥–Њ–Њ–±—А–∞–Ј–Є–µ –≤–љ–µ—И–љ–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤, –≤–ї–Є—П—О—Й–Є—Е –љ–∞¬†—Б–Њ–љ –ґ–Є–≤–Њ—В–љ—Л—Е –≤¬†–њ—А–Є—А–Њ–і–µ. –Ъ–Њ—А—А–µ–ї—П—Ж–Є–Њ–љ–љ—Л–µ –Њ—В–љ–Њ—И–µ–љ–Є—П –љ–µ¬†–≤—Б–µ–≥–і–∞ —Г–Ї–∞–Ј—Л–≤–∞—О—В –Є¬†–љ–∞ –њ—А–Є—З–Є–љ–љ–Њ-—Б–ї–µ–і—Б—В–≤–µ–љ–љ—Л–µ —Б–≤—П–Ј–Є. –Я—А–Є—Е–Њ–і–Є—В—Б—П –њ—А–Є–Ј–љ–∞—В—М, —З—В–Њ —Д—Г–љ–Ї—Ж–Є—П –Є¬†–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–∞—П —А–Њ–ї—М —Б–љ–∞ –і–Њ –Ї–Њ–љ—Ж–∞ –љ–µ¬†–њ–Њ–љ—П—В–љ—Л. –Ч–∞–і–∞—З–∞ —Н—В–Њ–≥–Њ –Њ–±–Ј–Њ—А–∞¬†вАУ –Њ–±–Њ–±—Й–Є—В—М —А–µ–Ј—Г–ї—М—В–∞—В—Л –љ–∞–Є–±–Њ–ї–µ–µ –Є–љ—В–µ—А–µ—Б–љ—Л—Е —Б—А–∞–≤–љ–Є—В–µ–ї—М–љ–Њ-—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Б–љ–∞, –≤—Л–њ–Њ–ї–љ–µ–љ–љ—Л—Е –≤¬†–њ–Њ—Б–ї–µ–і–љ–Є–µ –і–≤–∞ –і–µ—Б—П—В–Є–ї–µ—В–Є—П. –Ф–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ–∞—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П –Љ–Њ–ґ–µ—В –±—Л—В—М –љ–∞–є–і–µ–љ–∞ –≤¬†–Њ—А–Є–≥–Є–љ–∞–ї—М–љ—Л—Е —А–∞–±–Њ—В–∞—Е –Є¬†–і—А—Г–≥–Є—Е –Њ–±–Ј–Њ—А–∞—Е, —Б—Б—Л–ї–Ї–Є –љ–∞¬†–Ї–Њ—В–Њ—А—Л–µ –љ–∞—Е–Њ–і—П—В—Б—П –≤¬†—Б–њ–Є—Б–Ї–µ —Ж–Є—В–Є—А—Г–µ–Љ–Њ–є –ї–Є—В–µ—А–∞¬≠—В—Г—А—Л.
–°–Њ–љ —Г¬†–±–µ—Б–њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е
–Э–∞¬†—Б–µ–≥–Њ–і–љ—П—И–љ–Є–є –і–µ–љ—М —Б–Њ—Б—В–Њ—П–љ–Є–µ –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞ –Є–Ј—Г—З–µ–љ–Њ —Г¬†–љ–µ—Б–Ї–Њ–ї—М–Ї–Є—Е –і–µ—Б—П—В–Ї–Њ–≤ –≤–Є–і–Њ–≤ –±–µ—Б–њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е [7вАУ9]. –£¬†–≤—Б–µ—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ—Л—Е –≤–Є–і–Њ–≤ –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–є —Б–Њ–љ –Є–Љ–µ–µ—В —В–∞–Ї–Є–µ –ґ–µ –њ—А–Є–Ј–љ–∞–Ї–Є, –Ї–∞–Ї —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж, –≤–Ї–ї—О—З–∞—П –љ–µ–њ–Њ–і–≤–Є–ґ–љ–Њ—Б—В—М, —В–Є–њ–Є—З–љ—Г—О –њ–Њ–Ј—Г, —Б–љ–Є–ґ–µ–љ–љ—Г—О —А–µ–∞–Ї—Ж–Є—О –љ–∞¬†—А–∞–Ј–і—А–∞–ґ–Є—В–µ–ї–Є. –Я—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М —Б–љ–∞ —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В—Б—П –њ–Њ—Б–ї–µ –љ–∞–≤—П–Ј–∞–љ–љ–Њ–є –і–≤–Є–≥–∞—В–µ–ї—М–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є (–њ—А–Є–Ј–љ–∞–Ї¬†–≥–Њ–Љ–µ–Њ—Б—В–∞—В–Є—З–µ—Б–Ї–Њ–є —А–µ–≥—Г–ї—П—Ж–Є–Є). –Я—А–µ–њ–∞—А–∞—В—Л —Б¬†—Б–µ–і–∞—В–Є–≤–љ—Л–Љ —Н—Д—Д–µ–Ї—В–Њ–Љ —Г–і–ї–Є–љ—П—О—В –≤—А–µ–Љ—П —Б–љ–∞, –∞¬†—Б—В–Є–Љ—Г–ї—П—В–Њ—А—Л, –љ–∞–Њ–±–Њ—А–Њ—В, —Б–Њ–Ї—А–∞—Й–∞—О—В. –Ъ–∞–Ї –Є¬†—Г –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –љ–∞–Є–±–Њ–ї—М—И–µ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ —Б–љ–∞ —Г¬†–±–µ—Б–њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е –Њ—В–Љ–µ—З–∞–µ—В—Б—П –љ–∞¬†—А–∞–љ–љ–Є—Е —Н—В–∞–њ–∞—Е —А–∞–Ј–≤–Є—В–Є—П –Є¬†–њ–Њ—Б—В–µ–њ–µ–љ–љ–Њ —Б–љ–Є–ґ–∞–µ—В—Б—П —Б¬†–≤–Њ–Ј—А–∞—Б—В–Њ–Љ [8вАУ13]. –§–Њ—А–Љ–∞–ї—М–љ—Л–µ –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–µ —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є —Б–љ–∞ –љ–∞–±–ї—О–і–∞—О—В—Б—П –і–∞–ґ–µ —Г¬†–Љ–µ–і—Г–Ј¬†вАУ –њ—А–Њ—Б—В—Л—Е —Д–Њ—А–Љ —Б¬†–і–µ—Ж–µ–љ—В—А–∞–ї–Є–Ј–Њ–≤–∞–љ–љ–Њ–є –љ–µ—А–≤¬≠–љ–Њ–є —Б–Є—Б—В–µ–Љ–Њ–є, –Ї–Њ—В–Њ—А—Л–µ —Б—Г—Й–µ—Б—В–≤—Г—О—В –љ–∞¬†–Ч–µ–Љ–ї–µ —Г–ґ–µ –±–Њ–ї–µ–µ 600 –Љ–ї–љ –ї–µ—В [14]. –°–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ, –Њ—Б–љ–Њ–≤–љ—Л–µ –њ—А–Є–Ј–љ–∞–Ї–Є –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞ —Г¬†–ґ–Є–≤–Њ—В–љ—Л—Е —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–Є—Б—М –≤¬†—Н–≤–Њ–ї—О—Ж–Є–Є –µ—Й–µ –і–Њ –њ–Њ—П–≤–ї–µ–љ–Є—П —Ж–µ–љ—В—А–∞–ї–Є–Ј–Њ–≤–∞–љ–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л.
–£¬†–љ–µ—Б–Ї–Њ–ї—М–Ї–Є—Е –≤–Є–і–Њ–≤ –±–µ—Б–њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е –±—Л–ї–Є –Ј–∞—А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–љ—Л —В–∞–Ї–ґ–µ —Н–ї–µ–Ї—В—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –њ—А–Є–Ј–љ–∞–Ї–Є —Б–љ–∞. –Э–∞–њ—А–Є–Љ–µ—А, —Б–љ–Є–ґ–µ–љ–Є–µ –і–≤–Є–≥–∞—В–µ–ї—М–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –Є¬†—А–Њ—Б—В –њ–Њ—А–Њ–≥–Њ–≤ –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П —Г¬†—Д—А—Г–Ї—В–Њ–≤–Њ–є –Љ—Г—И–Ї–Є –њ—А–Њ–Є—Б—Е–Њ–і–Є–ї–Є –њ–∞—А–∞–ї–ї–µ–ї—М–љ–Њ —Г–Љ–µ–љ—М—И–µ–љ–Є—О –∞–Љ–њ–ї–Є—В—Г–і—Л —Н–ї–µ–Ї—В—А–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є (–ї–Њ–Ї–∞–ї—М–љ—Л–µ –њ–Њ—В–µ–љ—Ж–Є–∞–ї—Л –≤¬†–і–Є–∞–њ–∞–Ј–Њ–љ–µ 11вАУ40 –У—Ж)¬†–≥—А–Є–±–Њ–≤–Є–і–љ–Њ–≥–Њ —В–µ–ї–∞ –Є¬†–Ј–∞–Љ–µ–і–ї–µ–љ–Є—О —Г—А–Њ–≤–љ—П –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞. –Ф–µ–њ—А–Є–≤–∞—Ж–Є—П —Б–љ–∞ –њ—А–Є–≤–Њ–і–Є–ї–∞ –Ї¬†–Њ—В–і–∞—З–µ¬†вАУ —Б–љ–Є–ґ–µ–љ–Є—О –∞–Љ–њ–ї–Є—В—Г–і—Л —Н–ї–µ–Ї—В—А–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є¬†–≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ –Є¬†—А–Њ—Б—В—Г –њ–Њ—А–Њ–≥–Њ–≤ –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П [15вАУ17]. –£¬†—А–µ—З–љ–Њ–≥–Њ —А–∞–Ї–∞ —А–∞–Ј–≤–Є—В–Є–µ –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞ –њ—А–Њ–Є—Б—Е–Њ–і–Є–ї–Њ, –љ–∞–Њ–±–Њ—А–Њ—В, –њ–∞—А–∞–ї–ї–µ–ї—М–љ–Њ —Б¬†—А–Њ—Б—В–Њ–Љ –∞–Љ–њ–ї–Є—В—Г–і—Л —Н–ї–µ–Ї—В—А–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є (–≤¬†–і–Є–∞–њ–∞–Ј–Њ–љ–µ 8вАУ9 –У—Ж)¬†–≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ [18]. –Я–Њ¬†—Д–Њ—А–Љ–∞–ї—М–љ—Л–Љ –њ—А–Є–Ј–љ–∞–Ї–∞–Љ —Н—В–Њ —Б–Њ—Б—В–Њ—П–љ–Є–µ –љ–∞–њ–Њ–Љ–Є–љ–∞–µ—В –Ь–° —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г –≤¬†–Њ–±–Њ–Є—Е —Б–ї—Г—З–∞—П—Е –µ—Б—В—М –њ—А–Є–Ј–љ–∞–Ї–Є —Г–≤–µ–ї–Є—З–µ–љ–Є—П –Ї–Њ–≥–µ—А–µ–љ—В–љ–Њ—Б—В–Є —Н–ї–µ–Ї—В—А–Є—З–µ—Б–Ї–Є—Е –њ–Њ—В–µ–љ—Ж–Є–∞–ї–Њ–≤ –Љ–Њ–Ј–≥–∞. –£¬†–њ—З–µ–ї—Л —Б–њ–Њ–љ—В–∞–љ–љ–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –Ј—А–Є—В–µ–ї—М–љ—Л—Е –Є–љ—В–µ—А–љ–µ–є—А–Њ–љ–Њ–≤ –Љ–Њ–Ј–≥–∞ –і–љ–µ–Љ –≤¬†–њ–µ—А–Є–Њ–і –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –±—Л–ї–∞ –≤—Л—И–µ, —З–µ–Љ –≤¬†–њ–µ—А–Є–Њ–і –њ–Њ–Ї–Њ—П –≤¬†–љ–Њ—З–љ–Њ–µ –≤—А–µ–Љ—П. –С–Њ–ї–µ–µ —В–Њ–≥–Њ, –µ—Б–ї–Є –≤¬†–і–љ–µ–≤–љ–Њ–µ –≤—А–µ–Љ—П —Н—В–Є –љ–µ–є—А–Њ–љ—Л —А–µ–∞–≥–Є—А–Њ–≤–∞–ї–Є –љ–∞¬†–Ї–∞–ґ–і–Њ–µ –њ—А–µ–і—К—П–≤–ї–µ–љ–Є–µ –Ј—А–Є—В–µ–ї—М–љ–Њ–≥–Њ —Б—В–Є–Љ—Г–ї–∞, —В–Њ –≤¬†–љ–Њ—З–љ–Њ–µ –≤—А–µ–Љ—П –±—Л–ї–Є –њ–µ—А–Є–Њ–і—Л, –Ї–Њ–≥–і–∞ —А–µ–∞–Ї—Ж–Є—П –љ–∞¬†—Б—В–Є–Љ—Г–ї—Л –Њ—В—Б—Г—В—Б—В–≤–Њ–≤–∞–ї–∞ [19].
–°–Њ—Б—В–Њ—П–љ–Є–µ —Б–љ–∞ —Г¬†–±–µ—Б–њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е¬†вАУ –љ–µ¬†–Њ–і–љ–Њ—А–Њ–і–љ—Л–є –њ—А–Њ—Ж–µ—Б—Б. –Т¬†—Н—В–Њ–Љ —В–Њ–ґ–µ –µ—Б—В—М —Б—Е–Њ–і—Б—В–≤–Њ —Б–Њ¬†—Б–љ–Њ–Љ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж. –Э–∞–њ—А–Є–Љ–µ—А, —Г¬†–њ—З–µ–ї—Л –≤¬†—В–µ—З–µ–љ–Є–µ –≤—Б–µ–≥–Њ –љ–Њ—З–љ–Њ–≥–Њ –њ–µ—А–Є–Њ–і–∞ –њ–Њ–Ї–Њ—П —В–Њ–љ—Г—Б —И–µ–є–љ—Л—Е –Љ—Л—И—Ж –Є–Ј–Љ–µ–љ—П–µ—В—Б—П —Ж–Є–Ї–ї–Є—З–µ—Б–Ї–Є, –≤—А–µ–Љ–µ–љ–∞–Љ–Є –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –і–Њ—Б—В–Є–≥–∞—П –љ—Г–ї—П [10]. –£¬†–і—А–Њ–Ј–Њ—Д–Є–ї—Л –≤–µ–ї–Є—З–Є–љ–∞ –њ–Њ—А–Њ–≥–Њ–≤ –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П –Ј–∞–≤–Є—Б–Є—В –Њ—В¬†–і–ї–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Н–њ–Є–Ј–Њ–і–∞ —Б–љ–∞ [16].
–Т¬†–ї–Є—В–µ—А–∞—В—Г—А–µ –≤—Б—В—А–µ—З–∞—О—В—Б—П —Г–њ–Њ–Љ–Є–љ–∞–љ–Є—П –Њ¬†—Б—В–µ—А–µ–Њ—В–Є–њ–љ—Л—Е –і–≤–Є–ґ–µ–љ–Є—П—Е¬†–≥–ї–∞–Ј —Г¬†–љ–∞—Е–Њ–і—П—Й–Є—Е—Б—П –≤¬†—Б–Њ—Б—В–Њ—П–љ–Є–Є –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞ –Ї–∞—А–∞–Ї–∞—В–Є—Ж [20], –∞¬†—В–∞–Ї–ґ–µ –њ–Њ–і–µ—А–≥–Є–≤–∞–љ–Є–Є –∞–љ—В–µ–љ–љ –Є¬†–Ї–Њ–љ–µ—З–љ–Њ—Б—В–µ–є —Г¬†–њ—З–µ–ї [21] –Є¬†–Љ—Г—А–∞–≤—М–µ–≤ [22]. –Т–Њ –≤—Б–µ—Е —А–∞–±–Њ—В–∞—Е –Њ–±—А–∞—Й–∞–µ—В—Б—П –≤–љ–Є–Љ–∞–љ–Є–µ –љ–∞¬†–љ–µ–Ї–Њ—В–Њ—А–Њ–µ —Б—Е–Њ–і—Б—В–≤–Њ —Н—В–Є—Е –і–≤–Є–ґ–µ–љ–Є–є —Б–Њ¬†–≤–Ј–і—А–∞–≥–Є–≤–∞–љ–Є—П–Љ–Є —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –≤¬†–Я–°. –Ю–і–љ–∞–Ї–Њ –њ–Њ–і–Њ–±–љ–Њ–µ –њ–Њ–≤–µ–і–µ–љ–Є–µ —Г¬†–±–µ—Б–њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е –љ–Є–Ї–Њ–≥–і–∞ –і–µ—В–∞–ї—М–љ–Њ –љ–µ¬†–∞–љ–∞–ї–Є–Ј–Є—А–Њ–≤–∞–ї–Њ—Б—М.
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –њ–Њ¬†–њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–Љ –Є¬†–љ–µ–Ї–Њ—В–Њ—А—Л–Љ —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ –Ї—А–Є—В–µ—А–Є—П–Љ —Б–Њ—Б—В–Њ—П–љ–Є–µ —Б–љ–∞ –µ—Б—В—М —Г¬†–≤—Б–µ—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ—Л—Е –±–µ—Б–њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е, –≤–Ї–ї—О—З–∞—П –Њ—А–≥–∞–љ–Є–Ј–Љ—Л —Б¬†–і–µ—Ж–µ–љ—В—А–∞–ї–Є–Ј–Њ–≤–∞–љ–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ–Њ–є. –Я–Њ¬†—Д–Њ—А–Љ–∞–ї—М–љ—Л–Љ –њ—А–Є–Ј–љ–∞–Ї–∞–Љ —Н—В–Њ —Б–Њ—Б—В–Њ—П–љ–Є–µ –Є–Љ–µ–µ—В —Б—Е–Њ–і—Б—В–≤–Њ —Б¬†–Ь–° –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж.
–°–Њ–љ —Г¬†—А—Л–±
–≠–њ–Є–Ј–Њ–і—Л –љ–µ–њ–Њ–і–≤–Є–ґ–љ–Њ—Б—В–Є –Њ–њ—А–µ–і–µ–ї–µ–љ–љ–Њ–є –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В–Є (–њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–є —Б–Њ–љ) —Г¬†—А—Л–± —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—В—Б—П —Г—Е—Г–і—И–µ–љ–Є–µ–Љ —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В–Є —А–µ–∞–≥–Є—А–Њ–≤–∞—В—М –љ–∞¬†–≤–љ–µ—И–љ–Є–µ —А–∞–Ј–і—А–∞–ґ–Є—В–µ–ї–Є. –Я–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–є —Б–Њ–љ —А—Л–± –Є–Љ–µ–µ—В –њ—А–Є–Ј–љ–∞–Ї–Є¬†–≥–Њ–Љ–µ–Њ—Б—В–∞—В–Є—З–µ—Б–Ї–Њ–є –Є¬†—Ж–Є—А–Ї–∞–і–Є–∞–љ–љ–Њ–є —А–µ–≥—Г–ї—П—Ж–Є–Є. –Х–≥–Њ —Б—Г–Љ–Љ–∞—А–љ–∞—П –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В—Б—П –њ–Њ–і –і–µ–є—Б—В–≤–Є–µ–Љ –Љ–µ–ї–∞—В–Њ–љ–Є–љ–∞ –Є¬†–і–Є–∞–Ј–µ–њ–∞–Љ–∞ –Є¬†—Г–Љ–µ–љ—М—И–∞–µ—В—Б—П –њ–Њ–і –і–µ–є—Б—В–≤–Є–µ–Љ¬†–≥–Є–њ–Њ–Ї—А–µ—В–Є–љ–∞. –Э–∞–Є–±–Њ–ї—М—И–∞—П –Њ–±—Й–∞—П –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М —Н–њ–Є–Ј–Њ–і–Њ–≤ —Б–љ–∞ —Г¬†—А—Л–± –љ–∞–±–ї—О–і–∞–µ—В—Б—П –љ–∞¬†—А–∞–љ–љ–Є—Е —Н—В–∞–њ–∞—Е –њ–Њ—Б—В–љ–∞—В–∞–ї—М–љ–Њ–≥–Њ —А–∞–Ј–≤–Є—В–Є—П –Є¬†–њ–Њ—Б—В–µ–њ–µ–љ–љ–Њ —Б–љ–Є–ґ–∞–µ—В—Б—П —Б¬†–≤–Њ–Ј—А–∞—Б—В–Њ–Љ [23вАУ25].
–Ф–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞ —Г¬†—А—Л–± –≤–∞—А—М–Є—А—Г–µ—В—Б—П –≤¬†–±–Њ–ї—М—И–Њ–Љ –і–Є–∞–њ–∞–Ј–Њ–љ–µ. –Э–∞–њ—А–Є–Љ–µ—А, —Г¬†–Њ–±–Є—В–∞—О—Й–Є—Е –≤¬†–њ–µ—Й–µ—А–љ—Л—Е –≤–Њ–і–Њ–µ–Љ–∞—Е –Љ–µ–Ї—Б–Є–Ї–∞–љ—Б–Ї–Є—Е —А—Л–±–Њ–Ї (Astyanax mexicanus) –љ–∞¬†–њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–є —Б–Њ–љ –њ—А–Є—Е–Њ–і–Є—В—Б—П –≤¬†–њ—П—В—М —А–∞–Ј –Љ–µ–љ—М—И–µ –≤—А–µ–Љ–µ–љ–Є, —З–µ–Љ —Г¬†—А—Л–± —В–Њ–≥–Њ –ґ–µ –≤–Є–і–∞, –ґ–Є–≤—Г—Й–Є—Е –≤¬†—А–µ–Ї–∞—Е [26]. –Ь–µ–љ—М—И–∞—П –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М —Б–љ–∞ —Г¬†—А—Л–± –њ–µ—Й–µ—А–љ—Л—Е –њ–Њ–њ—Г–ї—П—Ж–Є–є –Њ–њ—А–µ–і–µ–ї—П–µ—В—Б—П –±–Њ–ї—М—И–Є–Љ —З–Є—Б–ї–Њ–Љ¬†–≥–Є–њ–Њ–Ї—А–µ—В–Є–љ—Б–Њ–і–µ—А–ґ–∞—Й–Є—Е –љ–µ–є—А–Њ–љ–Њ–≤ –≤¬†–≥–Є–њ–Њ—В–∞–ї–∞–Љ—Г—Б–µ. –Ґ–Њ—А–Љ–Њ–ґ–µ–љ–Є–µ –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –љ–µ–є—А–Њ–љ–Њ–≤ –њ—А–Є–≤–Њ–і–Є—В –Ї¬†—Г–≤–µ–ї–Є—З–µ–љ–Є—О –≤—А–µ–Љ–µ–љ–Є —Б–љ–∞ [27]. –†–∞–Ј—А—Г—И–µ–љ–Є–µ —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –±–Њ–Ї–Њ–≤–Њ–є –ї–Є–љ–Є–Є, –љ–∞–Њ–±–Њ—А–Њ—В, –њ–Њ–≤—Л—И–∞–µ—В –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М —Б–љ–∞ —Г¬†–њ–µ—Й–µ—А–љ—Л—Е —Д–Њ—А–Љ –і–Њ —Г—А–Њ–≤–љ—П —А–µ—З–љ—Л—Е —А—Л–±–Њ–Ї, —З—В–Њ —Г–Ї–∞–Ј—Л–≤–∞–µ—В –љ–∞¬†–≤–∞–ґ–љ—Г—О —А–Њ–ї—М —Б–µ–љ—Б–Њ—А–љ–Њ–є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є –≤¬†—А–µ–≥—Г–ї—П—Ж–Є–Є –Ї–Њ–ї–Є—З–µ—Б—В–≤–∞ —Б–љ–∞ [8]. –†–µ–Ј—Г–ї—М—В–∞—В—Л —Н—В–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —А–∞—Б–Ї—А—Л–≤–∞—О—В –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ–Њ-–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є–µ –Є¬†–љ–µ–є—А–Њ–љ–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л —А–µ–≥—Г–ї—П—Ж–Є–Є –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Б–љ–∞, –∞¬†—В–∞–Ї–ґ–µ –Ї–Њ–љ—Б–µ—А–≤–∞—В–Є–≤–љ–Њ—Б—В—М –љ–µ–є—А–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Є—Е —Б–Є—Б—В–µ–Љ –Љ–Њ–Ј–≥–∞, —А–µ–≥—Г–ї–Є—А—Г—О—Й–Є—Е —Д–µ–љ–Њ—В–Є–њ —Б–љ–∞ —Г¬†—А–∞–Ј–љ—Л—Е –ґ–Є–≤–Њ—В–љ—Л—Е. –≠–≤–Њ–ї—О—Ж–Є—П —Б–љ–∞ –њ–µ—Й–µ—А–љ–Њ–є —Д–Њ—А–Љ—Л –Љ–µ–Ї—Б–Є–Ї–∞–љ—Б–Ї–Њ–є —А—Л–±–Ї–Є, –Ї–Њ—В–Њ—А–∞—П –њ—А–Њ–Є—Б—Е–Њ–і–Є–ї–∞ –≤¬†—В–µ—З–µ–љ–Є–µ –њ—А–Є–Љ–µ—А–љ–Њ 2вАУ3 –Љ–ї–љ –ї–µ—В,¬†вАУ –њ—А–µ–Ї—А–∞—Б–љ—Л–є –њ—А–Є–Љ–µ—А –≤–ї–Є—П–љ–Є—П —Н–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤ (–і–µ—Д–Є—Ж–Є—В –Ї–Њ—А–Љ–∞, –Њ–±–Є—В–∞–љ–Є–µ –≤¬†—В–µ–Љ–љ–Њ—В–µ) –љ–∞¬†–њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М —Б–љ–∞ –Є¬†–µ–≥–Њ —Б—Г—В–Њ—З–љ—Г—О –і–Є–љ–∞¬≠–Љ–Є–Ї—Г.
–Ф–∞–љ–љ—Л–µ –Њ–±¬†—Н–ї–µ–Ї—В—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –∞—Б–њ–µ–Ї—В–∞—Е —Б–љ–∞ —Г¬†—А—Л–± –њ—А–Њ—В–Є–≤–Њ—А–µ—З–Є–≤—Л. –Ь–µ–і–ї–µ–љ–љ—Л–µ –≤–Њ–ї–љ—Л –Є¬†–Њ—Б—В—А—Л–µ —Б–њ–∞–є–Ї–Є –±—Л–ї–Є –Ј–∞—Д–Є–Ї—Б–Є—А–Њ–≤–∞–љ—Л –≤–Њ –≤—А–µ–Љ—П –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞ —Г¬†—Б–Њ–Љ–∞ [28]. –Т¬†–і—А—Г–≥–Є—Е —А–∞–±–Њ—В–∞—Е –Њ–і–љ–Њ–љ–∞–њ—А–∞–≤–ї–µ–љ–љ—Л—Е –Є–Ј–Љ–µ–љ–µ–љ–Є–є —Н–ї–µ–Ї—В—А–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –љ–µ¬†—А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–ї–Њ—Б—М [7]. –Т¬†–Њ–і–љ–Њ–Љ –Є–Ј¬†–њ–µ—А–≤—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Б–љ–∞ —Г¬†—А—Л–± –±—Л–ї–Є –Њ–њ–Є—Б–∞–љ—Л —Б—В–µ—А–µ–Њ—В–Є–њ–љ—Л–µ –і–≤–Є–ґ–µ–љ–Є—П¬†–≥–ї–∞–Ј –Є¬†—Б–і–µ–ї–∞–љ–Њ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–µ–љ–Є–µ –Њ¬†–љ–∞–ї–Є—З–Є–Є —Г¬†–љ–Є—Е –Я–° [29]. –Ю–і–љ–∞–Ї–Њ –њ–Њ–Ј–ґ–µ —Н—В–Є —А–µ–Ј—Г–ї—М—В–∞—В—Л –љ–µ¬†–њ–Њ–і—В–≤–µ—А–і–Є–ї–Є—Б—М [28, 30]. –Р–љ–∞—В–Њ–Љ–Є—З–µ—Б–Ї–Є–µ –Є¬†–Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ–Ї–∞–Ј–∞–ї–Є –±–Њ–ї—М—И–Њ–µ —Б—Е–Њ–і—Б—В–≤–Њ –≤¬†–Њ—А–≥–∞–љ–Є–Ј–∞—Ж–Є–Є –∞–Ї—В–Є–≤–Є—А—Г—О—Й–Є—Е (—Е–Њ–ї–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Њ–є, –Љ–Њ–љ–Њ–∞–Љ–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Њ–є,¬†–≥–Є–њ–Њ–Ї—А–µ—В–Є–љ–Њ–≤–Њ–є) –Є¬†—Б–Њ–Љ–љ–Њ–≥–µ–љ–љ—Л—Е (–Љ–µ–ї–∞—В–Њ–љ–Є–љ–Њ–≤–Њ–є) —Б–Є—Б—В–µ–Љ –Љ–Њ–Ј–≥–∞ —Г¬†—А—Л–± –Є¬†–њ–Њ–Ј–≤–Њ–љ–Њ—З–љ—Л—Е [24, 31].
–°–Њ–љ —Г¬†–∞–Љ—Д–Є–±–Є–є
–Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Б–љ–∞ —Г¬†–∞–Љ—Д–Є–±–Є–є –Ї—А–∞–є–љ–µ –Љ–∞–ї–Њ. –£¬†–Њ–і–љ–Є—Е –≤–Є–і–Њ–≤ –њ—А–Є–Ј–љ–∞–Ї–Њ–≤ –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞ –Њ–±–љ–∞—А—Г–ґ–µ–љ–Њ –љ–µ¬†–±—Л–ї–Њ. –£¬†–і—А—Г–≥–Є—Е —В–∞–Ї–Є–µ –њ—А–Є–Ј–љ–∞–Ї–Є –±—Л–ї–Є, –љ–Њ¬†–љ–µ –±—Л–ї–Њ —Н–ї–µ–Ї—В—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Ї–Њ—А—А–µ–ї—П—В–Њ–≤. –Э–∞–Ї–Њ–љ–µ—Ж, –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–µ –њ—А–Є–Ј–љ–∞–Ї–Є —Б–љ–∞ –њ–∞—А–∞–ї–ї–µ–ї—М–љ–Њ —Б¬†—Е–∞—А–∞–Ї—В–µ—А–љ—Л–Љ–Є –≤–Њ–ї–љ–∞–Љ–Є-—Б–њ–∞–є–Ї–∞–Љ–Є –≤¬†–≠–≠–У –±—Л–ї–Є –Њ–њ–Є—Б–∞–љ—Л —Г¬†–ґ–∞–±—Л –Є¬†–ї—П–≥—Г—И–Ї–Є [7, 28, 32]. –Ш–Љ–µ—О—Й–Є–µ—Б—П –і–∞–љ–љ—Л–µ —Б–Ї–Њ—А–µ–µ —Г–Ї–∞–Ј—Л–≤–∞—О—В –љ–∞¬†–љ–∞–ї–Є—З–Є–µ –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞ —Г¬†–∞–Љ—Д–Є–±–Є–є, —В–Њ–≥–і–∞ –Ї–∞–Ї –і–∞–љ–љ—Л—Е –Њ–±¬†—Н–ї–µ–Ї—В—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Ї–Њ—А—А–µ–ї—П—В–∞—Е –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ.
–°–Њ–љ —Г —А–µ–њ—В–Є–ї–Є–є
–Т—Л—Б–Њ–Ї–Њ–∞–Љ–њ–ї–Є—В—Г–і–љ—Л–µ —Б–њ–∞–є–Ї–Є –љ–∞¬†—Н–ї–µ–Ї—В—А–Њ—Н–љ—Ж–µ—Д–∞–ї–Њ–≥—А–∞–Љ–Љ–µ (–≠–≠–У), –љ–∞–њ–Њ–Љ–Є–љ–∞—О—Й–Є–µ –Љ–µ–і–ї–µ–љ–љ—Л–µ –≤–Њ–ї–љ—Л —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –±—Л–ї–Є –Ј–∞—А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–љ—Л –љ–∞¬†—Д–Њ–љ–µ –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞ —Г¬†–Љ–љ–Њ–≥–Є—Е –≤–Є–і–Њ–≤ –њ—А–µ—Б–Љ—Л–Ї–∞—О—Й–Є—Е—Б—П, –≤–Ї–ї—О—З–∞—П –Ї–∞–є–Љ–∞–љ–∞, –Є–≥—Г–∞–љ—Г, —З–µ—А–µ–њ–∞—Е—Г –Є¬†–Ј–Љ–µ—О [7, 32]. –§–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ—А–Њ–і–µ–Љ–Њ–љ—Б—В—А–Є—А–Њ–≤–∞–ї–Є, —З—В–Њ —А–µ–∞–Ї—Ж–Є—П –≠–≠–У-—Б–њ–∞–є–Ї–Њ–≤ —Г¬†—А–µ–њ—В–Є–ї–Є–є –љ–∞¬†—Б–Њ–Љ–љ–Њ–≥–µ–љ–љ—Л–µ –њ—А–µ–њ–∞—А–∞—В—Л —В–∞–Ї–∞—П –ґ–µ, –Ї–∞–Ї –Є¬†–≥–Є–њ–њ–Њ–Ї–∞–Љ–њ–∞–ї—М–љ—Л—Е —Б–њ–∞–є–Ї–Њ–≤ (–∞–љ–∞–ї–Њ–≥–Њ–≤ –Ї–Њ—А–Ї–Њ–≤–Њ–є –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є –≠–≠–У) —Г¬†–Ї–Њ—И–Ї–Є. –І–∞—Б—В–Њ—В–∞ –≠–≠–У-—Б–њ–∞–є–Ї–Њ–≤ —Г¬†—А–µ–њ—В–Є–ї–Є–є —В–∞–Ї–ґ–µ —Г–≤–µ–ї–Є—З–Є–≤–∞–ї–∞—Б—М –њ–Њ—Б–ї–µ –љ–∞–≤—П–Ј–∞–љ–љ–Њ–≥–Њ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П [33].
–Я—А–Є–Ј–љ–∞–Ї–Є –Я–° —Г¬†—А–µ–њ—В–Є–ї–Є–є –≤¬†—А–∞–љ–љ–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П—Е –Њ—В–Љ–µ—З–∞–ї–Є—Б—М —А–µ–і–Ї–Њ. –Ю–і–љ–Є–Љ –Є–Ј¬†–∞—А–≥—Г–Љ–µ–љ—В–Њ–≤ –≤¬†–њ–Њ–ї—М–Ј—Г –Њ—В—Б—Г—В—Б—В–≤–Є—П —Н—В–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞ —Г¬†—А–µ–њ—В–Є–ї–Є–є –±—Л–ї–Њ –Є¬†–љ–∞—И–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –і–Є–љ–∞–Љ–Є–Ї–Є —А–∞–Ј—А—П–і–∞ –Є–Ј–Њ–ї–Є—А–Њ–≤–∞–љ–љ—Л—Е –љ–µ–є—А–Њ–љ–Њ–≤ –≤¬†—Ж–Є–Ї–ї–µ ¬Ђ—Б–Њ–љ¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬ї —Г¬†—З–µ—А–µ–њ–∞—Е. –Э–∞–Љ –љ–µ¬†—Г–і–∞–ї–Њ—Б—М –Њ–±–љ–∞—А—Г–ґ–Є—В—М –њ—А–Є–Ј–љ–∞–Ї–Є –њ–Њ–≤—В–Њ—А—П—О—Й–µ–є—Б—П –∞–Ї—В–Є–≤–∞—Ж–Є–Є –љ–µ–є—А–Њ–љ–Њ–≤ —А–µ—В–Є–Ї—Г–ї—П—А–љ–Њ–є —Д–Њ—А–Љ–∞—Ж–Є–Є¬†вАУ –Ї–ї—О—З–µ–≤–Њ–≥–Њ –њ—А–Є–Ј–љ–∞–Ї–∞ –Я–° —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е вАТ —Г¬†—З–µ—А–µ–њ–∞—Е –≤–Њ –≤—А–µ–Љ—П –њ–µ—А–Є–Њ–і–Њ–≤ –µ—Б—В–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Б–љ–∞ [34]. –Ґ–µ–Љ –љ–µ¬†–Љ–µ–љ–µ–µ –Ї–∞–Ї –Љ–Є–љ–Є–Љ—Г–Љ —Г¬†–і–≤—Г—Е –≤–Є–і–Њ–≤ —З–µ—А–µ–њ–∞—Е —Б–Њ–љ –±—Л–ї —А–∞–Ј–і–µ–ї–µ–љ –љ–∞¬†–і–≤–µ —Б—В–∞–і–Є–Є¬†вАУ –Ь–° –Є¬†–Я–° [35, 36]. –Э–µ–і–∞–≤–љ–Њ —Г¬†–і–≤—Г—Е –≤–Є–і–Њ–≤ —П—Й–µ—А–Є—Ж –±—Л–ї–Є –Ј–∞—А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–љ—Л –њ–Њ–≤—В–Њ—А—П—О—Й–Є–µ—Б—П –і–≤–Є–ґ–µ–љ–Є—П¬†–≥–ї–∞–Ј –≤–Њ –≤—А–µ–Љ—П –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞. –≠–њ–Є–Ј–Њ–і—Л –љ–∞–±–ї—О–і–∞–ї–Є—Б—М —Б¬†–Є–љ—В–µ—А–≤–∞–ї–Њ–Љ 80 —Б–µ–Ї—Г–љ–і¬†–Є —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–ї–Є—Б—М —Б–љ–Є–ґ–µ–љ–Є–µ–Љ –∞–Љ–њ–ї–Є—В—Г–і—Л –≠–≠–У-—Б–њ–∞–є–Ї–Њ–≤. –§–ї—Г–Њ–Ї—Б–µ—В–Є–љ (–Є–љ–≥–Є–±–Є—В–Њ—А –Њ–±—А–∞—В–љ–Њ–≥–Њ —В—А–∞–љ—Б–њ–Њ—А—В–∞ —Б–µ—А–Њ—В–Њ–љ–Є–љ–∞, —Г–Ї–Њ—А–∞—З–Є–≤–∞–µ—В –≤—А–µ–Љ—П –Я–° —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е) —Г–Љ–µ–љ—М—И–∞–ї –∞–Љ–њ–ї–Є—В—Г–і—Г –≠–≠–У-—Б–њ–∞–є–Ї–Њ–≤ [37, 38]. –Я–Њ¬†–Љ–љ–µ–љ–Є—О –∞–≤—В–Њ—А–Њ–≤, —В–∞–Ї–Є–µ —Н–њ–Є–Ј–Њ–і—Л –Є–Љ–µ—О—В –њ—А–Є–Ј–љ–∞–Ї–Є –Я–° –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е. –Я—А–Є—А–Њ–і–∞ —А–µ–≥—Г–ї—П—А–љ–Њ–≥–Њ —З–µ—А–µ–і–Њ–≤–∞–љ–Є—П –і–≤—Г—Е —Б—В–∞–і–Є–є —Б–љ–∞ —Б¬†–њ–Њ–і–Њ–±–љ—Л–Љ –Ї–Њ—А–Њ—В–Ї–Є–Љ –њ–µ—А–Є–Њ–і–Њ–Љ –љ–µ¬†–њ–Њ–љ—П—В–љ–∞ –Є¬†–љ–µ –Є–Љ–µ–µ—В –∞–љ–∞–ї–Њ–≥–Њ–≤ —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж.
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, —Г¬†—А–µ–њ—В–Є–ї–Є–є, –њ–Њ¬†–≤—Б–µ–є –≤–Є–і–Є–Љ–Њ—Б—В–Є, –µ—Б—В—М —Б—В–∞–і–Є—П —Б–љ–∞, –љ–∞–њ–Њ–Љ–Є–љ–∞—О—Й–∞—П –њ–Њ¬†–њ–∞—А–∞–Љ–µ—В—А–∞–Љ –Ь–° –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е. –£–±–µ–і–Є—В–µ–ї—М–љ—Л—Е –і–∞–љ–љ—Л—Е –Њ¬†—Б—Г—Й–µ—Б—В–≤–Њ–≤–∞–љ–Є–Є —Г¬†—А–µ–њ—В–Є–ї–Є–є –Я–° –љ–µ—В. –†–µ–њ—В–Є–ї–Є–Є –і–∞–ї–Є –љ–∞—З–∞–ї–Њ –Њ–±—Й–µ–Љ—Г –њ—А–µ–і–Ї—Г –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж. –Х—Б–ї–Є –Я–° —Г¬†—А–µ–њ—В–Є–ї–Є–є –Њ—В—Б—Г—В—Б—В–≤—Г–µ—В, –Ј–љ–∞—З–Є—В, –Њ–љ –њ–Њ—П–≤–Є–ї—Б—П –≤¬†—Н–≤–Њ–ї—О—Ж–Є–Є —Н—В–Є—Е –і–≤—Г—Е –≥—А—Г–њ–њ –љ–µ–Ј–∞–≤–Є—Б–Є–Љ–Њ.
–°–Њ–љ —Г¬†–њ—В–Є—Ж
–≠–ї–µ–Ї—В—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ –Љ–µ—В–Њ–і–Њ–Љ —Б–Њ–љ –Є–Ј—Г—З–µ–љ –њ—А–Є–Љ–µ—А–љ–Њ —Г¬†30¬†–≤–Є–і–Њ–≤ –Ї–ї–∞—Б—Б–∞ –њ—В–Є—Ж, –±–Њ–ї—М—И–Є–љ—Б—В–≤–Њ –Є–Ј¬†–Ї–Њ—В–Њ—А—Л—Е –њ—А–Є–љ–∞–і–ї–µ–ґ–∞—В –Ї¬†–љ–∞–і–Њ—В—А—П–і—Г –љ–Њ–≤–Њ–љ—С–±–љ—Л—Е (Neognathae), –Є¬†–ї–Є—И—М –і–≤–∞ –≤–Є–і–∞ (–∞—Д—А–Є–Ї–∞–љ—Б–Ї–Є–є —Б—В—А–∞—Г—Б –Є¬†—В–Є–љ–∞–Љ—Г) –Њ—В–љ–Њ—Б—П—В—Б—П –Ї–Њ¬†–≤—В–Њ—А–Њ–Љ—Г –љ–∞–і–Њ—В—А—П–і—Г¬†вАУ –±–µ—Б–Ї–Є–ї–µ–≤—Л—Е, –Є–ї–Є –і—А–µ–≤–љ–µ–љ—С–±–љ—Л—Е (Palaeognathae). –Э–Њ–≤–Њ–љ—С–±–љ—Л–µ –љ–∞—Б—З–Є—В—Л–≤–∞—О—В –±–Њ–ї–µ–µ 10 —В—Л—Б. –≤–Є–і–Њ–≤ —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л—Е –њ—В–Є—Ж, –∞¬†–±–µ—Б–Ї–Є–ї–µ–≤—Л–µ, –Є–ї–Є –і—А–µ–≤–љ–µ–љ—С–±–љ—Л–µ,¬†вАУ –±–∞–Ј–Њ–≤–∞—П¬†–≥—А—Г–њ–њ–∞, –≤–Ї–ї—О—З–∞—О—Й–∞—П –≤—Б–µ–≥–Њ 60 –≤–Є–і–Њ–≤ —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л—Е –њ—В–Є—Ж. –Ш—Е¬†–њ—А–µ–і–Ї–Є –Њ—В–і–µ–ї–Є–ї–Є—Б—М –Њ—В¬†–Њ—Б–љ–Њ–≤–љ–Њ–є –≤–µ—В–≤–Є, –і–∞–≤—И–µ–є –љ–∞—З–∞–ї–Њ –љ–Њ–≤–Њ–љ—С–±–љ—Л–Љ –њ—В–Є—Ж–∞–Љ, –њ—А–Є–Љ–µ—А–љ–Њ 110¬†–Љ–ї–љ –ї–µ—В –љ–∞–Ј–∞–і [39].
–Ґ–Њ–ї—М–Ї–Њ —Г¬†–њ—В–Є—Ж –Є¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е —Б–Њ–љ —Б–Њ—Б—В–Њ–Є—В –Є–Ј¬†–і–≤—Г—Е –њ–Њ–і—Б—В–∞–і–Є–є¬†вАУ –Ь–° –Є¬†–Я–°. –У–ї–∞–≤–љ—Л–є –њ—А–Є–Ј–љ–∞–Ї –Ь–° —Г¬†–њ—В–Є—Ж¬†вАУ —Е–∞—А–∞–Ї—В–µ—А–љ—Л–µ –Љ–µ–і–ї–µ–љ–љ—Л–µ –Ї–Њ–ї–µ–±–∞–љ–Є—П –≤¬†–≠–≠–У –≤¬†–і–Є–∞–њ–∞–Ј–Њ–љ–µ 1вАУ4 –У—Ж. –Ф—А—Г–≥–Є–µ —А–Є—В–Љ—Л (–љ–∞–њ—А–Є–Љ–µ—А, —Б–Њ–љ–љ—Л–µ –≤–µ—А–µ—В–µ–љ–∞) —Г¬†–њ—В–Є—Ж –Њ–±—Л—З–љ–Њ –љ–µ¬†–≤—Л–і–µ–ї—П—О—В—Б—П. –£¬†–±–Њ–ї—М—И–Є–љ—Б—В–≤–∞ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ—Л—Е –≤–Є–і–Њ–≤ –Љ–µ–і–ї–µ–љ–љ—Л–µ –≤–Њ–ї–љ—Л –≤¬†–≠–≠–У —А–∞–Ј–≤–Є–≤–∞—О—В—Б—П –≤¬†–і–≤—Г—Е –њ–Њ–ї—Г—И–∞—А–Є—П—Е —Б–Є–љ—Е—А–Њ–љ–љ–Њ (–±–Є–ї–∞—В–µ—А–∞–ї—М–љ–Њ-—Б–Є–Љ–Љ–µ—В—А–Є—З–љ—Л–є –Ь–°). –Ъ–∞–Ї –Љ–Є–љ–Є–Љ—Г–Љ —Г¬†—Б–µ–Љ–Є –≤–Є–і–Њ–≤ –≤¬†–Ь–° –±—Л–ї–Є –Ј–∞—А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–љ—Л —Н–њ–Є–Ј–Њ–і—Л –Љ–µ–ґ–њ–Њ–ї—Г—И–∞—А–љ–Њ–є –∞—Б–Є–Љ–Љ–µ—В—А–Є–Є –≠–≠–У, –Є–ї–Є –Њ–і–љ–Њ–њ–Њ–ї—Г—И–∞—А–љ—Л–є —Б–Њ–љ. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М —Н—В–Є—Е —Н–њ–Є–Ј–Њ–і–Њ–≤, –Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ, —Б–Њ—Б—В–∞–≤–ї—П–µ—В –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —Б–µ–Ї—Г–љ–і [3, 40вАУ44] –Є¬†—В–Њ–ї—М–Ї–Њ —Г¬†–љ–µ–Ї–Њ—В–Њ—А—Л—Е –≤–Є–і–Њ–≤ –±–Њ–ї–µ–µ –Њ–і–љ–Њ–є –Љ–Є–љ—Г—В—Л [43, 45]. –≠–њ–Є–Ј–Њ–і—Л –Љ–µ–ґ–њ–Њ–ї—Г—И–∞—А–љ–Њ–є –∞—Б–Є–Љ–Љ–µ—В—А–Є–Є –≠–≠–У —Г¬†–њ—В–Є—Ж —З–∞—Б—В–Њ —Б–Њ—З–µ—В–∞—О—В—Б—П —Б¬†–∞—Б–Є–Љ–Љ–µ—В—А–Є—З–љ—Л–Љ —Б–Њ—Б—В–Њ—П–љ–Є–µ–Љ¬†–≥–ї–∞–Ј. –Э–∞–њ—А–Є–Љ–µ—А, —Г—В–Ї–Є, –Ї–Њ—В–Њ—А—Л–µ —Б–њ—П—В –љ–∞¬†–Ї—А–∞—О¬†–≥—А—Г–њ–њ—Л, –њ—А–Њ–≤–Њ–і—П—В –±–Њ–ї—М—И–µ –≤—А–µ–Љ–µ–љ–Є —Б¬†–Њ–і–љ–Є–Љ –Њ—В–Ї—А—Л—В—Л–Љ¬†–≥–ї–∞–Ј–Њ–Љ –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–њ—В–Є—Ж–∞–Љ–Є, –Ї–Њ—В–Њ—А—Л–µ —Б–њ—П—В –≤¬†—Ж–µ–љ—В—А–µ¬†–≥—А—Г–њ–њ—Л –≤¬†–Њ–Ї—А—Г–ґ–µ–љ–Є–Є –і—А—Г–≥–Є—Е –Њ—Б–Њ–±–µ–є. –Я—А–Є —Н—В–Њ–Љ¬†–≥–ї–∞–Ј, –Ї–Њ—В–Њ—А—Л–є –љ–∞–њ—А–∞–≤–ї–µ–љ –Њ—В¬†—Ж–µ–љ—В—А–∞¬†–≥—А—Г–њ–њ—Л, –љ–∞—Е–Њ–і–Є—В—Б—П –≤¬†–Њ—В–Ї—А—Л—В–Њ–Љ —Б–Њ—Б—В–Њ—П–љ–Є–Є –±–Њ–ї—М—И–µ –≤—А–µ–Љ–µ–љ–Є, —З–µ–Љ¬†–≥–ї–∞–Ј, –Њ–±—А–∞—Й–µ–љ–љ—Л–є –љ–∞¬†–і—А—Г–≥–Є—Е –њ—В–Є—Ж. –Р—Б–Є–Љ–Љ–µ—В—А–Є—З–љ–Њ–µ —Б–Њ—Б—В–Њ—П–љ–Є–µ¬†–≥–ї–∞–Ј –Ї–Њ—А—А–µ–ї–Є—А—Г–µ—В —Б¬†—Н–њ–Є–Ј–Њ–і–∞–Љ–Є –Љ–µ–ґ–њ–Њ–ї—Г—И–∞—А–љ–Њ–є –∞—Б–Є–Љ–Љ–µ—В—А–Є–Є –≠–≠–У: –≤¬†–њ–Њ–ї—Г—И–∞—А–Є–Є, –њ—А–Њ—В–Є–≤–Њ–њ–Њ–ї–Њ–ґ–љ–Њ–Љ –Ј–∞–Ї—А—Л—В–Њ–Љ—Г¬†–≥–ї–∞–Ј—Г, –Љ–µ–і–ї–µ–љ–љ—Л–µ –≤–Њ–ї–љ—Л –≠–≠–У –Є–Љ–µ–ї–Є –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї—Г—О –∞–Љ–њ–ї–Є—В—Г–і—Г, —З–µ–Љ –≤¬†–њ–Њ–ї—Г—И–∞—А–Є–Є, –њ—А–Њ—В–Є–≤–Њ–њ–Њ–ї–Њ–ґ–љ–Њ–Љ –Њ—В–Ї—А—Л—В–Њ–Љ—Г¬†–≥–ї–∞–Ј—Г [44]. –§—А–µ–≥–∞—В—Л –≤–Њ –≤—А–µ–Љ—П –њ–Њ–ї–µ—В–Њ–≤ –љ–∞–і –Њ–Ї–µ–∞–љ–Њ–Љ –љ–∞¬†–Ї–Њ—А–Љ–Њ–≤—Л–µ –њ–Њ–ї—П —Б–њ—П—В –≤¬†–≤–Њ–Ј–і—Г—Е–µ, –њ–ї–∞–љ–Є—А—Г—П –њ–Њ¬†—Б–њ–Є—А–∞–ї–Є. –Т–Љ–µ—Б—В–µ —Б¬†—В–µ–Љ –њ–Њ–ї—Г—И–∞—А–Є–µ –Љ–Њ–Ј–≥–∞, –њ—А–Њ—В–Є–≤–Њ–њ–Њ–ї–Њ–ґ–љ–Њ–µ –љ–∞–њ—А–∞–≤–ї–µ–љ–Є—О –≤—А–∞—Й–µ–љ–Є—П, –±–Њ–ї—М—И–µ –≤—А–µ–Љ–µ–љ–Є –љ–∞—Е–Њ–і–Є—В—Б—П –≤¬†—Б–Њ—Б—В–Њ—П–љ–Є–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є–ї–Є –љ–Є–Ј–Ї–Њ–∞–Љ–њ–ї–Є—В—Г–і–љ–Њ–≥–Њ –Ь–°. –Ъ–Њ—Б–≤–µ–љ–љ–Њ —Н—В–Њ¬†–≥–Њ–≤–Њ—А–Є—В –Њ¬†—В–Њ–Љ, —З—В–Њ –≤–Њ –≤—А–µ–Љ—П —Б–љ–∞ –њ—В–Є—Ж—Л –Њ—В–Ї—А—Л–≤–∞—О—В¬†–≥–ї–∞–Ј, –Ї–Њ—В–Њ—А—Л–є –љ–∞–њ—А–∞–≤–ї–µ–љ –≤¬†—Б—В–Њ—А–Њ–љ—Г –≤—А–∞—Й–µ–љ–Є—П [45]. –Я–Њ—Н—В–Њ–Љ—Г —Д—Г–љ–Ї—Ж–Є—П –Ь–° —Б¬†–Љ–µ–ґ–њ–Њ–ї—Г—И–∞—А–љ–Њ–є –∞—Б–Є–Љ–Љ–µ—В—А–Є–µ–є –≠–≠–У —Г¬†–њ—В–Є—Ж —Б–Њ—Б—В–Њ–Є—В, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –≤¬†–Њ–±—А–∞–±–Њ—В–Ї–µ –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є, –њ–Њ—Б—В—Г–њ–∞—О—Й–µ–є –Њ—В¬†–Њ—В–Ї—А—Л—В–Њ–≥–Њ –≤¬†—Б—В–Њ—А–Њ–љ—Г –Њ–њ–∞—Б–љ–Њ—Б—В–Є¬†–≥–ї–∞–Ј–∞, –Є, —Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ, –њ–Њ–і–і–µ—А–ґ–∞–љ–Є–Є –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є –≤–Њ –≤—А–µ–Љ—П —Б–љ–∞ [40вАУ43]. –°—З–Є—В–∞–µ—В—Б—П, —З—В–Њ –Њ—Б–љ–Њ–≤–љ—Л–µ –њ—А–Є–Ј–љ–∞–Ї–Є –Я–° –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е (–±—Л—Б—В—А—Л–µ –і–≤–Є–ґ–µ–љ–Є—П¬†–≥–ї–∞–Ј, –Љ—Л—И–µ—З–љ—Л–µ –≤–Ј–і—А–∞–≥–Є–≤–∞–љ–Є—П, —В–Њ—А–Љ–Њ–ґ–µ–љ–Є–µ —Б–Ї–µ–ї–µ—В–љ–Њ–є –Љ—Г—Б–Ї—Г–ї–∞—В—Г—А—Л) –µ—Б—В—М –Є¬†—Г –њ—В–Є—Ж. –Ю–і–љ–∞–Ї–Њ —Г¬†–±–Њ–ї—М—И–Є–љ—Б—В–≤–∞ –≤–Є–і–Њ–≤ –њ—В–Є—Ж –Я–° –љ–µ¬†—Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–µ—В—Б—П –Ј–∞–Љ–µ—В–љ—Л–Љ —Б–љ–Є–ґ–µ–љ–Є–µ–Љ –Љ—Л—И–µ—З–љ–Њ–≥–Њ —В–Њ–љ—Г—Б–∞ –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–Ь–°. –£¬†–≤—Б–µ—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ—Л—Е –≤–Є–і–Њ–≤ –њ—В–Є—Ж, –Ї—А–Њ–Љ–µ –∞—Д—А–Є–Ї–∞–љ—Б–Ї–Њ–≥–Њ —Б—В—А–∞—Г—Б–∞, —Н–њ–Є–Ј–Њ–і—Л –Я–° –і–ї—П—В—Б—П –≤—Б–µ–≥–Њ –ї–Є—И—М –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —Б–µ–Ї—Г–љ–і [3, 40вАУ42]. –Я—А–Є–Љ–µ–љ—П—П —В—А–∞–і–Є—Ж–Є–Њ–љ–љ—Л–µ –њ–Њ–ї–Є–≥—А–∞—Д–Є—З–µ—Б–Ї–Є–µ –Ї—А–Є—В–µ—А–Є–Є, —Б–ї–Њ–ґ–љ–Њ —В–Њ—З–љ–Њ –Њ–њ—А–µ–і–µ–ї–Є—В—М¬†–≥—А–∞–љ–Є—Ж—Л —В–∞–Ї–Є—Е —Н–њ–Є–Ј–Њ–і–Њ–≤ –Я–°.
–°–Њ–љ –∞—Д—А–Є–Ї–∞–љ—Б–Ї–Њ–≥–Њ —Б—В—А–∞—Г—Б–∞ –Њ—В–ї–Є—З–∞–µ—В—Б—П –Њ—В¬†—Б–љ–∞ –≤—Б–µ—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ—Л—Е –њ—В–Є—Ж [46], –≤–Ї–ї—О—З–∞—П –µ—Й–µ –Њ–і–љ–Њ–≥–Њ –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї—П –і—А–µ–≤–љ–µ–љ—С–±–љ—Л—Е –њ—В–Є—Ж¬†вАУ —В–Є–љ–∞–Љ—Г [47]. –£¬†–≤–Ј—А–Њ—Б–ї—Л—Е —Б—В—А–∞—Г—Б–Њ–≤ —Н–њ–Є–Ј–Њ–і—Л –Я–° –Љ–Њ–≥—Г—В –і–Њ—Б—В–Є–≥–∞—В—М –њ—П—В–Є –Љ–Є–љ—Г—В, –±–Њ–ї–µ–µ —З–µ–Љ –≤¬†–і–µ—Б—П—В—М —А–∞–Ј –њ—А–µ–≤—Л—И–∞—П –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ—Г—О –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М —Н–њ–Є–Ј–Њ–і–∞ –Я–° —Г¬†–і—А—Г–≥–Є—Е –њ—В–Є—Ж. –Я—А–Є —Н—В–Њ–Љ –љ–∞¬†–Я–° —Г¬†—Б—В—А–∞—Г—Б–Њ–≤ –њ—А–Є—Е–Њ–і–Є—В—Б—П –≤¬†—Б—А–µ–і–љ–µ–Љ 24% –Њ—В¬†–≤—А–µ–Љ–µ–љ–Є –љ–Њ—З–љ–Њ–≥–Њ –њ–µ—А–Є–Њ–і–∞, –Є–ї–Є 26% –Њ—В¬†–Њ–±—Й–µ–≥–Њ –≤—А–µ–Љ–µ–љ–Є —Б–љ–∞, —В–Њ –µ—Б—В—М –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –±–Њ–ї—М—И–µ, —З–µ–Љ —Г¬†–і—А—Г–≥–Є—Е –њ—В–Є—Ж [3]. –Ш–љ—В–µ—А–µ—Б–љ–Њ, —З—В–Њ —Е–∞—А–∞–Ї—В–µ—А–љ—Л–µ –і–ї—П –Я–° –≤–Ј–і—А–∞–≥–Є–≤–∞–љ–Є—П¬†–≥–ї–∞–Ј –Є¬†–≥–Њ–ї–Њ–≤—Л —А–µ–≥–Є—Б—В—А–Є—А—Г—О—В—Б—П –љ–∞¬†—Д–Њ–љ–µ –Ї–∞–Ї –љ–Є–Ј–Ї–Њ–∞–Љ–њ–ї–Є—В—Г–і–љ–Њ–є, —В–∞–Ї –Є¬†–≤—Л—Б–Њ–Ї–Њ–∞–Љ–њ–ї–Є—В—Г–і–љ–Њ–є –≠–≠–У. –Я–Њ–і–Њ–±–љ—Л–µ –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В–Є —Б–љ–∞ —Г¬†—Б—В—А–∞—Г—Б–∞ –њ–Њ—Б–ї—Г–ґ–Є–ї–Є –Њ—Б–љ–Њ–≤–∞–љ–Є–µ–Љ –і–ї—П¬†–≥–Є–њ–Њ—В–µ–Ј—Л –Њ¬†—В–Њ–Љ, —З—В–Њ –µ–≥–Њ –љ–µ–і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–љ—Л–є —Б–Њ–љ —П–≤–ї—П–µ—В—Б—П –њ—А–µ–і—И–µ—Б—В–≤–µ–љ–љ–Є–Ї–Њ–Љ ¬Ђ—Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–Њ –±–Њ–ї–µ–µ –Љ–Њ–ї–Њ–і–Њ–≥–Њ¬ї –Я–° –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж, –Ї–Њ—В–Њ—А—Л–є —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–µ—В—Б—П –∞–Ї—В–Є–≤–∞—Ж–Є–µ–є –Ї–Њ—А—Л –њ–µ—А–µ–і–љ–µ–≥–Њ –Љ–Њ–Ј–≥–∞. –Я–Њ—Н—В–Њ–Љ—Г, –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ, –Я–° —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї—Б—П –≤¬†—Н–≤–Њ–ї—О—Ж–Є–Є –њ—В–Є—Ж –Є¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –љ–µ–Ј–∞–≤–Є—Б–Є–Љ–Њ [46]. –Ю–і–љ–∞–Ї–Њ –Њ—В—Б—Г—В—Б—В–≤–Є–µ —В–∞–Ї–Њ–є –ґ–µ –љ–µ–і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–љ–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞ —Г¬†—В–Є–љ–∞–Љ—Г –Є¬†–Ї–Њ—А–Њ—В–Ї–Є–µ —Н–њ–Є–Ј–Њ–і—Л –Є—Е –Я–° (–Ї–∞–Ї —Г¬†–≤—Б–µ—Е –љ–Њ–≤–Њ–љ—С–±–љ—Л—Е –њ—В–Є—Ж) —Б—В–∞–≤—П—В —Н—В—Г¬†–≥–Є–њ–Њ—В–µ–Ј—Г –њ–Њ–і —Б–Њ–Љ–љ–µ–љ–Є–µ.
–С–Њ–ї—М—И–Є–љ—Б—В–≤–Њ –љ–µ–є—А–Њ–љ–Њ–≤ –≤¬†—А–µ—В–Є–Ї—Г–ї—П—А–љ–Њ–є —Д–Њ—А–Љ–∞—Ж–Є–Є —Б—В–≤–Њ–ї–∞ –Љ–Њ–Ј–≥–∞ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е —А–∞–Ј—А—П–ґ–∞—О—В—Б—П —Б¬†–љ–∞–Є–±–Њ–ї—М—И–µ–є —З–∞—Б—В–Њ—В–Њ–є –≤–Њ –≤—А–µ–Љ—П –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†–Я–°, —Б–љ–Є–ґ–∞—П —З–∞—Б—В–Њ—В—Г —А–∞–Ј—А—П–і–∞ –≤¬†–Ь–°. –Ф—А—Г–≥–Є–Љ–Є —Б–ї–Њ–≤–∞–Љ–Є, –∞–Ї—В–Є–≤–∞—Ж–Є—П –њ–Њ–і–Ї–Њ—А–Ї–Њ–≤—Л—Е —Б—В—А—Г–Ї—В—Г—А –Љ–Њ–Ј–≥–∞ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е¬†вАУ –Њ–і–Є–љ –Є–Ј¬†–≤–∞–ґ–љ–µ–є—И–Є—Е –њ—А–Є–Ј–љ–∞–Ї–Њ–≤ –Я–°. –Э–∞–Љ–Є –±—Л–ї–∞ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–∞ –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В—М –Є–Љ–њ—Г–ї—М—Б–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –љ–µ–є—А–Њ–љ–Њ–≤ —Б—В–≤–Њ–ї–∞ –Љ–Њ–Ј–≥–∞ –≤–Њ –≤—А–µ–Љ—П —Ж–Є–Ї–ї–∞ ¬Ђ—Б–Њ–љ¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬ї —Г¬†–і–Њ–Љ–∞—И–љ–Є—Е –Ї—Г—А. –Ю–Ї–∞–Ј–∞–ї–Њ—Б—М, —З—В–Њ –±–Њ–ї—М—И–Є–љ—Б—В–≤–Њ —В–∞–Ї–Є—Е –љ–µ–є—А–Њ–љ–Њ–≤ —А–∞–Ј—А—П–ґ–∞–ї–Є—Б—М —Б¬†–љ–∞–Є–±–Њ–ї—М—И–µ–є —З–∞—Б—В–Њ—В–Њ–є –≤–Њ –≤—А–µ–Љ—П —Н–њ–Є–Ј–Њ–і–Њ–≤ –Я–°. –Я–µ—А–Є–Њ–і—Л —А–∞–Ј—А—П–і–∞ –љ–µ–є—А–Њ–љ–Њ–≤ –љ–µ¬†–≤—Б–µ–≥–і–∞ —В–Њ—З–љ–Њ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Њ–≤–∞–ї–Є –і—А—Г–≥–Є–Љ –њ—А–Є–Ј–љ–∞–Ї–∞–Љ –Я–° [48]. –Т¬†–љ–∞–Є–±–Њ–ї—М—И–µ–є —Б—В–µ–њ–µ–љ–Є —Г—З–∞—Й–µ–љ–Є–µ —З–∞—Б—В–Њ—В—Л —А–∞–Ј—А—П–і–∞ —Б–Њ–≤–њ–∞–і–∞–ї–Њ —Б¬†–±—Л—Б—В—А—Л–Љ–Є –і–≤–Є–ґ–µ–љ–Є—П–Љ–Є¬†–≥–ї–∞–Ј –Є¬†–≤ –љ–∞–Є–Љ–µ–љ—М—И–µ–є¬†вАУ —Б¬†–Є–Ј–Љ–µ–љ–µ–љ–Є–µ–Љ –Љ—Л—И–µ—З–љ–Њ–≥–Њ —В–Њ–љ—Г—Б–∞. –Я–Њ—Н—В–Њ–Љ—Г –≤¬†—Н—В–Њ–Љ —Б–ї—Г—З–∞–µ –њ—А–∞–≤–Є–ї—М–љ–µ–µ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞—В—М –љ–∞–Ј–≤–∞–љ–Є–µ ¬Ђ—Б–Њ–љ —Б¬†–±—Л—Б—В—А—Л–Љ–Є –і–≤–Є–ґ–µ–љ–Є—П–Љ–Є¬†–≥–ї–∞–Ј¬ї, —В–∞–Ї –Ї–∞–Ї –Є–Љ–µ–љ–љ–Њ —Н—В–Њ—В –њ–∞—А–∞–Љ–µ—В—А —П–≤–ї—П–µ—В—Б—П –Њ–њ—А–µ–і–µ–ї—П—О—Й–Є–Љ –і–ї—П —Н—В–Њ–≥–Њ —Б–Њ—Б—В–Њ—П–љ–Є—П —Г¬†–њ—В–Є—Ж. –Э–µ–Ї–Њ—В–Њ—А—Л–µ —Н–њ–Є–Ј–Њ–і—Л –±—Л—Б—В—А—Л—Е –і–≤–Є–ґ–µ–љ–Є–є¬†–≥–ї–∞–Ј —Г¬†–Ї—Г—А —А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–ї–Є—Б—М –љ–∞¬†—Д–Њ–љ–µ –Љ–µ–і–ї–µ–љ–љ—Л—Е –≤–Њ–ї–љ –≤¬†–≠–≠–У, —В–Њ –µ—Б—В—М –±–µ–Ј –∞–Ї—В–Є–≤–∞—Ж–Є–Є –њ–µ—А–µ–і–љ–µ–≥–Њ –Љ–Њ–Ј–≥–∞ (—А–Є—Б.¬†1). –≠—В–Њ –Њ–Ј–љ–∞—З–∞–µ—В, —З—В–Њ –і–∞–ґ–µ —Г¬†–Ї—Г—А¬†вАУ —В–Є–њ–Є—З–љ—Л—Е –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–µ–є –љ–Њ–≤–Њ–љ—С–±–љ—Л—Е –Я–° –Љ–µ–љ–µ–µ –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ, —З–µ–Љ —Г¬†–љ–∞–Ј–µ–Љ–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, —Б–ї–Њ–ґ–Є–≤—И–µ–µ—Б—П –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є–µ –Њ¬†—В–Њ–Љ, —З—В–Њ —Б—Г–Љ–Љ–∞—А–љ–∞—П –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М –Я–°/—Б–љ–∞ —Б¬†–±—Л—Б—В—А—Л–Љ–Є –і–≤–Є–ґ–µ–љ–Є—П–Љ–Є¬†–≥–ї–∞–Ј —Г¬†–њ—В–Є—Ж –Љ–µ–љ—М—И–µ, —З–µ–Љ —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ, –Њ—И–Є–±–Њ—З–љ–Њ, –µ—Б–ї–Є –≤¬†–і–Њ–њ–Њ–ї–љ–µ–љ–Є–µ –Ї¬†–њ–Њ–ї–Є–≥—А–∞—Д–Є—З–µ—Б–Ї–Є–Љ –Ї—А–Є—В–µ—А–Є—П–Љ –њ—А–Є–Љ–µ–љ—П—В—М –њ–∞—А–∞–Љ–µ—В—А –∞–Ї—В–Є–≤–∞—Ж–Є–Є –љ–µ–є—А–Њ–љ–Њ–≤ —А–µ—В–Є–Ї—Г–ї—П—А–љ–Њ–є —Д–Њ—А–Љ–∞—Ж–Є–Є.
–Ь–љ–Њ–≥–Є–µ –њ—В–Є—Ж—Л —Б–Њ–≤–µ—А—И–∞—О—В —Б–µ–Ј–Њ–љ–љ—Л–µ –Љ–Є–≥—А–∞—Ж–Є–Є, –∞¬†—В–∞–Ї–ґ–µ –њ–Њ–ї–µ—В—Л –љ–∞¬†–Ї–Њ—А–Љ–Њ–≤—Л–µ –њ–Њ–ї—П, –њ—А–Є—З–µ–Љ –Њ–љ–Є –љ–∞—Е–Њ–і—П—В—Б—П –≤¬†–≤–Њ–Ј–і—Г—Е–µ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –і–љ–µ–є –Є–ї–Є –і–∞–ґ–µ –љ–µ–і–µ–ї—М. –Я—В–Є—Ж—Л –Љ–Њ–≥—Г—В —Б–њ–∞—В—М –≤¬†–њ–Њ–ї–µ—В–µ, –љ–Њ¬†–Њ–±—Й–∞—П –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М —Б–љ–∞ –њ—А–Є —Н—В–Њ–Љ, —Б–Ї–Њ—А–µ–µ –≤—Б–µ–≥–Њ, –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –Љ–µ–љ—М—И–µ —В–∞–Ї–Њ–≤–Њ–є –љ–∞¬†–Ј–µ–Љ–ї–µ. –Э–∞–њ—А–Є–Љ–µ—А, —Г¬†—Д—А–µ–≥–∞—В–Њ–≤ –Њ–±—Й–µ–µ –≤—А–µ–Љ—П —Б–љ–∞ –≤¬†–њ–Њ–ї–µ—В–µ —Б–Њ—Б—В–∞–≤–ї—П–µ—В –Љ–µ–љ–µ–µ 7% –Њ—В¬†–њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Б–љ–∞ –≤¬†–≥–љ–µ–Ј–і–∞—Е [45]. –≠—В–Њ –Њ–і–Є–љ –Є–Ј¬†–њ—А–Є–Љ–µ—А–Њ–≤ –∞–і–∞–њ—В–Є–≤–љ–Њ–≥–Њ —Б–Њ–Ї—А–∞—Й–µ–љ–Є—П –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Б–љ–∞ –≤¬†—Б–Є—В—Г–∞—Ж–Є–Є, –Ї–Њ–≥–і–∞ —Б–Њ–љ –њ—А–µ–њ—П—В—Б—В–≤—Г–µ—В —А–µ–∞–ї–Є–Ј–∞—Ж–Є–Є –±–Њ–ї–µ–µ –≤–∞–ґ–љ–Њ–є —Б—В—А–∞—В–µ–≥–Є–Є –њ–Њ–≤–µ–і–µ–љ–Є—П, –∞¬†–Є–Љ–µ–љ–љ–Њ –Ї–Њ—А–Љ–ї–µ–љ–Є—О –Є¬†–Њ–±–µ—Б–њ–µ—З–µ–љ–Є—О –±–µ–Ј–Њ–њ–∞—Б–љ–Њ—Б—В–Є. –Т¬†–і—А—Г–≥–Є—Е —Б–ї—Г—З–∞—П—Е –њ—В–Є—Ж—Л —Б–Њ–Ї—А–∞—Й–∞—О—В –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ —Б–љ–∞ –≤¬†–њ–µ—А–Є–Њ–і —Б–њ–∞—А–Є–≤–∞–љ–Є—П. –Т¬†—А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–∞–Љ—Ж—Л, –Ї–Њ—В–Њ—А—Л–µ —Б–њ—П—В –Љ–µ–љ—М—И–µ, —Б–њ–∞—А–Є–≤–∞—О—В—Б—П —Б¬†–±–Њ–ї—М—И–Є–Љ —З–Є—Б–ї–Њ —Б–∞–Љ–Њ–Ї. –°—Г–і—П –њ–Њ¬†—З–∞—Б—В–Њ—В–µ –≤—Б—В—А–µ—З–∞–µ–Љ–Њ—Б—В–Є –њ—В–Є—Ж –Є¬†–Є—Е –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В–Є, —Б–Њ–Ї—А–∞—Й–µ–љ–Є–µ –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Б–љ–∞ –≤¬†–њ–µ—А–Є–Њ–і —А–∞–Ј–Љ–љ–Њ–ґ–µ–љ–Є—П –љ–µ¬†–Є–Љ–µ–µ—В –њ–Њ—Б–ї–µ–і—Б—В–≤–Є–є –і–ї—П –Є—Е —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ —Б–Њ—Б—В–Њ—П–љ–Є—П –Є¬†–Ј–і–Њ—А–Њ–≤—М—П [49].
–°–Њ–љ —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е
–Э–∞¬†–Ч–µ–Љ–ї–µ –Њ–±–Є—В–∞–µ—В –њ—А–Є–Љ–µ—А–љ–Њ 3,5 —В—Л—Б. –≤–Є–і–Њ–≤ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –≤–Ї–ї—О—З–∞—П –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–µ–є –њ–ї–∞—Ж–µ–љ—В–∞—А–љ—Л—Е, —Б—Г–Љ—З–∞—В—Л—Е –Є¬†–Њ–і–љ–Њ–њ—А–Њ—Е–Њ–і–љ—Л—Е. –°–Њ–љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ –±–Њ–ї–µ–µ —З–µ–Љ —Г¬†100 –≤–Є–і–Њ–≤. –°—Г—Й–µ—Б—В–≤–Њ–≤–∞–љ–Є–µ –Ь–° —Г¬†–Њ–і–љ–Њ–њ—А–Њ—Е–Њ–і–љ—Л—Е (–µ—Е–Є–і–љ—Л, —Г—В–Ї–Њ–љ–Њ—Б–∞) –љ–µ¬†–≤—Л–Ј—Л–≤–∞–µ—В —Б–Њ–Љ–љ–µ–љ–Є–є. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П —Б–Є—В—Г–∞—Ж–Є—П —Б¬†–Я–° —Г¬†—Н—В–Є—Е –ґ–Є–≤–Њ—В–љ—Л—Е –љ–µ¬†–њ–Њ–љ—П—В–љ–∞. –Ю–і–љ–Є —Б—З–Є—В–∞—О—В, —З—В–Њ –Я–° —Г¬†–µ—Е–Є–і–љ—Л –љ–µ—В [50]. –Ф—А—Г–≥–Є–µ –њ—А–Є–і–µ—А–ґ–Є–≤–∞—О—В—Б—П –Љ–љ–µ–љ–Є—П, —З—В–Њ –Я–° —Г¬†–µ—Е–Є–і–љ—Л –µ—Б—В—М [51]. –Ґ—А–µ—В—М–Є, –Њ—Б–љ–Њ–≤—Л–≤–∞—П—Б—М –љ–∞¬†–Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В—П—Е —А–∞–Ј—А—П–і–∞ –љ–µ–є—А–Њ–љ–Њ–≤ –≤¬†—Б—В–≤–Њ–ї–µ –Љ–Њ–Ј–≥–∞, –њ—А–µ–і–њ–Њ–ї–∞–≥–∞—О—В, —З—В–Њ —Г¬†–µ—Е–Є–і–љ—Л –µ—Б—В—М –љ–µ–і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–љ–∞—П —Б—В–∞–і–Є—П —Б–љ–∞, —Б–Њ—З–µ—В–∞—О—Й–∞—П –њ—А–Є–Ј–љ–∞–Ї–Є –Ь–° –Є¬†–Я–° –њ–ї–∞—Ж–µ–љ—В–∞—А–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е [52]. –Ш–љ—В–µ—А–µ—Б–љ–Њ, —З—В–Њ —Г¬†—Г—В–Ї–Њ–љ–Њ—Б–∞ –Љ—Л—И–µ—З–љ—Л–µ –≤–Ј–і—А–∞–≥–Є–≤–∞–љ–Є—П –Є¬†–њ–Њ–і–µ—А–≥–Є–≤–∞–љ–Є—П —А–µ–≥–Є—Б—В—А–Є—А—Г—О—В—Б—П –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ —Б¬†–≤—Л—Б–Њ–Ї–Њ–≤–Њ–ї—М—В–љ–Њ–є –≠–≠–У. –Э–∞¬†–Њ—Б–љ–Њ–≤–∞–љ–Є–Є —Н—В–Є—Е –і–∞–љ–љ—Л—Е –±—Л–ї–∞ –≤—Л–і–µ–ї–µ–љ–∞ —Б—В–∞–і–Є—П —Б–њ–Њ–Ї–Њ–є–љ–Њ–≥–Њ —Б–љ–∞ (–≥–Њ–Љ–Њ–ї–Њ–≥–Є—З–љ–∞—П –Ь–° –љ–∞–Ј–µ–Љ–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е) –Є¬†—Б—В–∞–і–Є—П, –Ї–Њ—В–Њ—А–∞—П —Б–Њ—З–µ—В–∞–µ—В –њ—А–Є–Ј–љ–∞–Ї–Є –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–љ–Њ–≥–Њ –Ь–° –Є¬†–Я–° –њ–ї–∞—Ж–µ–љ—В–∞—А–љ—Л—Е [52, 53]. –Ю—З–µ–≤–Є–і–љ—Л–µ –њ—А–Є–Ј–љ–∞–Ї–Є –љ–µ–і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–љ–Њ–≥–Њ –Я–° –µ—Б—В—М –Є¬†—Г —Б—Г–Љ—З–∞—В—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е. –Э–∞–њ—А–Є–Љ–µ—А, —Г¬†–Њ–њ–Њ—Б—Б—Г–Љ–∞ —Е–∞—А–∞–Ї—В–µ—А–љ—Л–µ –і–ї—П –Я–° –±—Л—Б—В—А—Л–µ –і–≤–Є–ґ–µ–љ–Є—П¬†–≥–ї–∞–Ј –Є¬†–Љ—Л—И–µ—З–љ—Л–µ –≤–Ј–і—А–∞–≥–Є–≤–∞–љ–Є—П —Д–Є–Ї—Б–Є—А—Г—О—В—Б—П –љ–∞¬†—Д–Њ–љ–µ –љ–µ¬†—В–Њ–ї—М–Ї–Њ –∞–Ї—В–Є–≤–Є—А–Њ–≤–∞–љ–љ–Њ–≥–Њ –њ–∞—В—В–µ—А–љ–∞ –≤¬†–≠–≠–У, –љ–Њ¬†–Є –љ–Є–Ј–Ї–Њ–∞–Љ–њ–ї–Є—В—Г–і–љ—Л—Е –Љ–µ–і–ї–µ–љ–љ—Л—Е –≤–Њ–ї–љ [54]. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, —Б–Њ–љ (–≤¬†–њ–µ—А–≤—Г—О –Њ—З–µ—А–µ–і—М –Я–°) —Г¬†—Н–≤–Њ–ї—О—Ж–Є–Њ–љ–љ–Њ ¬Ђ–њ—А–Є–Љ–Є—В–Є–≤–љ—Л—Е¬ї –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е (–Њ–і–љ–Њ–њ—А–Њ—Е–Њ–і–љ—Л—Е, —Б—Г–Љ—З–∞—В—Л—Е) –Є–Љ–µ–µ—В –њ—А–Є–Ј–љ–∞–Ї–Є –љ–µ–і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–љ–Њ—Б—В–Є, –Ї–∞–Ї –Є¬†—Г –∞—Д—А–Є–Ї–∞–љ—Б–Ї–Њ–≥–Њ —Б—В—А–∞—Г—Б–∞.
–Т—Б–µ –љ–∞–Ј–µ–Љ–љ—Л–µ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є–µ –≤–Њ –≤—А–µ–Љ—П —Б–љ–∞ –љ–µ–њ–Њ–і–≤–Є–ґ–љ—Л,¬†–≥–ї–∞–Ј–∞ –њ—А–Є —Н—В–Њ–Љ –Ј–∞–Ї—А—Л—В—Л, –∞¬†–і—Л—Е–∞–љ–Є–µ —А–µ–≥—Г–ї—П—А–љ–Њ. –Ь–µ–і–ї–µ–љ–љ—Л–µ –≤–Њ–ї–љ—Л –≤¬†–≠–≠–У –і–≤—Г—Е –њ–Њ–ї—Г—И–∞—А–Є–є –Ї–Њ—А—Л –Љ–Њ–Ј–≥–∞ —А–∞–Ј–≤–Є–≤–∞—О—В—Б—П –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ. –Я–° –Њ–±–љ–∞—А—Г–ґ–µ–љ —Г¬†–≤—Б–µ—Е –љ–∞–Ј–µ–Љ–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е. –Ю–±—Л—З–љ–Њ –Њ–љ —А–∞–Ј–≤–Є–≤–∞–µ—В—Б—П –њ–Њ—Б–ї–µ –Ь–°. –Ш–Ј–±–Є—А–∞—В–µ–ї—М–љ–∞—П –і–µ–њ—А–Є–≤–∞—Ж–Є—П –Я–° –њ—А–Є–≤–Њ–і–Є—В –Ї¬†—Г–≤–µ–ї–Є—З–µ–љ–Є—О —З–Є—Б–ї–∞ –њ–Њ–њ—Л—В–Њ–Ї –Ј–∞—Б–љ—Г—В—М –Я–°, —В–Њ –µ—Б—В—М –њ—А–µ–і–њ–Њ–ї–∞–≥–∞–µ—В—Б—П, —З—В–Њ –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М –Я–° —А–µ–≥—Г–ї–Є—А—Г–µ—В—Б—П¬†–≥–Њ–Љ–µ–Њ—Б—В–∞—В–Є—З–µ—Б–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ. –Ф–µ–њ—А–Є–≤–∞—Ж–Є—П –Я–° —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–µ—В—Б—П –љ–∞—А—Г—И–µ–љ–Є–µ–Љ –Ї–Њ–≥–љ–Є—В–Є–≤–љ—Л—Е —Д—Г–љ–Ї—Ж–Є–є, –Љ–Њ–ґ–µ—В –Є–Љ–µ—В—М –њ–Њ—Б–ї–µ–і—Б—В–≤–Є—П –і–ї—П –Ј–і–Њ—А–Њ–≤—М—П –Є¬†–і–∞–ґ–µ –њ—А–Є–≤–Њ–і–Є—В—М –Ї¬†—Б–Љ–µ—А—В–Є [55].
–°—А–µ–і–Є –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –љ–∞–Є–±–Њ–ї—М—И–µ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –Я–° —А–µ–≥–Є—Б—В—А–Є—А—Г–µ—В—Б—П —Г¬†–Њ–і–љ–Њ–њ—А–Њ—Е–Њ–і–љ—Л—Е –Є¬†—Б—Г–Љ—З–∞—В—Л—Е, —В–Њ –µ—Б—В—М —Г¬†–ґ–Є–≤–Њ—В–љ—Л—Е —Б¬†–љ–∞–Є–±–Њ–ї–µ–µ –њ—А–Є–Љ–Є—В–Є–≤–љ–Њ —Г—Б—В—А–Њ–µ–љ–љ—Л–Љ –Љ–Њ–Ј–≥–Њ–Љ. –Т¬†—Ж–µ–ї–Њ–Љ –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М –Я–° –±–Њ–ї—М—И–µ —Г¬†–љ–µ–Ј—А–µ–ї–Њ—А–Њ–ґ–і–∞—О—Й–Є—Е—Б—П –≤–Є–і–Њ–≤, —З—В–Њ¬†–≥–Њ–≤–Њ—А–Є—В –Њ¬†–µ–≥–Њ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ–є —А–Њ–ї–Є –≤¬†—А–∞–Ј–≤–Є—В–Є–Є –Љ–Њ–Ј–≥–∞. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, —Б—Г—Й–µ—Б—В–≤—Г–µ—В –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–∞—П –Ї–Њ—А—А–µ–ї—П—Ж–Є—П –Љ–µ–ґ–і—Г –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ–Љ –Я–° —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†—Б—В–µ–њ–µ–љ—М—О –Є—Е –Ј–∞—Й–Є—Й–µ–љ–љ–Њ—Б—В–Є –≤–Њ –≤—А–µ–Љ—П —Б–љ–∞ [3вАУ6]. –С–µ–Ј—Г—Б–ї–Њ–≤–љ–Њ, –љ—Г–ґ–љ–Њ —Г—З–Є—В—Л–≤–∞—В—М, —З—В–Њ –љ–∞—И–Є –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є—П –Њ¬†—Б–љ–µ –ґ–Є–≤–Њ—В–љ—Л—Е —Б–ї–Њ–ґ–Є–ї–Є—Б—М –≤¬†–њ–µ—А–≤—Г—О –Њ—З–µ—А–µ–і—М –њ–Њ¬†—А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є, –≤—Л–њ–Њ–ї–љ–µ–љ–љ—Л—Е –≤¬†–ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е. –Т¬†–њ—А–Є—А–Њ–і–µ –ґ–Є–≤–Њ—В–љ—Л–µ –љ–µ¬†—Б–њ—П—В –Ї–∞–ґ–і—Л–є –і–µ–љ—М –≤¬†–Њ–і–љ–Њ –Є¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –Є¬†–Я–° [56, 57]. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, —Б–љ–Є–ґ–µ–љ–Є–µ –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В–Є –Я–°¬†вАУ –љ–µ¬†–µ–і–Є–љ—Б—В–≤–µ–љ–љ—Л–є —Б–њ–Њ—Б–Њ–± —Б–Њ–Ї—А–∞—Й–µ–љ–Є—П –≤—А–µ–Љ–µ–љ–Є –њ—А–µ–±—Л–≤–∞–љ–Є—П –≤¬†–љ–∞–Є–±–Њ–ї–µ–µ¬†–≥–ї—Г–±–Њ–Ї–Њ–є (—Б—Г–і—П –њ–Њ¬†–њ–Њ—А–Њ–≥–∞–Љ –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П) —Б—В–∞–і–Є–Є —Б–љ–∞. –Ф—А—Г–≥–Њ–є —Б–њ–Њ—Б–Њ–±¬†вАУ –Є–Ј–Љ–µ–љ–µ–љ–Є–µ —Ж–Є—А–Ї–∞–і–љ–Њ–є –Њ—А–≥–∞–љ–Є–Ј–∞—Ж–Є–Є –њ–∞—В—В–µ—А–љ–∞ —Б–љ–∞ –Є¬†—Б–Љ–µ—Й–µ–љ–Є–µ –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –љ–∞¬†–≤—А–µ–Љ—П, –Ї–Њ–≥–і–∞ —Б—В–µ–њ–µ–љ—М –Њ–њ–∞—Б–љ–Њ—Б—В–Є –љ–Є–ґ–µ [58].
–°–Њ–љ –Љ–Њ—А—Б–Ї–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Њ—В–ї–Є—З–∞–µ—В—Б—П –Њ—В¬†—Б–љ–∞ –љ–∞–Ј–µ–Љ–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –њ–Њ¬†–љ–µ–Ї–Њ—В–Њ—А—Л–Љ –њ—А–Є–Ј–љ–∞–Ї–∞–Љ, –≤–Ї–ї—О—З–∞—П —В–µ, —З—В–Њ —Б—З–Є—В–∞—О—В—Б—П –Ї–ї—О—З–µ–≤—Л–Љ–Є –і–ї—П –Њ–њ—А–µ–і–µ–ї–µ–љ–Є—П —Б–∞–Љ–Њ–≥–Њ —Б–Њ—Б—В–Њ—П–љ–Є—П —Б–љ–∞. –Ю—Б–Њ–±–µ–љ–љ–Њ—Б—В–Є —Б–љ–∞ –≤–Њ–і–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е¬†вАУ –њ—А–µ–Ї—А–∞—Б–љ—Л–є –њ—А–Є–Љ–µ—А, –≤¬†–Ї–∞–Ї–Њ–є —Б—В–µ–њ–µ–љ–Є —Б–Њ–љ –ґ–Є–≤–Њ—В–љ—Л—Е –Ј–∞–≤–Є—Б–Є—В –Њ—В¬†—Г—Б–ї–Њ–≤–Є–є –Є—Е –Њ–±–Є—В–∞–љ–Є—П. –Я—А–µ–і–Ї–Є —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л—Е –≤–Њ–і–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –і–Њ–ї–ґ–љ—Л –±—Л–ї–Є ¬Ђ—А–µ—И–Є—В—М¬ї –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –≤–∞–ґ–љ—Л—Е –њ—А–Њ–±–ї–µ–Љ, –Њ–±—Г—Б–ї–Њ–≤–ї–µ–љ–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П–Љ–Є —Б–љ–∞ –≤¬†–≤–Њ–і–µ. –Т–Њ-–њ–µ—А–≤—Л—Е, –≤¬†–≤–Њ–і–µ –љ–µ–≤–Њ–Ј–Љ–Њ–ґ–љ–∞ –і–ї–Є—В–µ–ї—М–љ–∞—П –љ–µ–њ–Њ–і–≤–Є–ґ–љ–Њ—Б—В—М. –Ц–Є–≤–Њ—В–љ—Л–Љ –љ—Г–ґ–љ–Њ —А–µ–≥—Г–ї—П—А–љ–Њ –≤—Б–њ–ї—Л–≤–∞—В—М –Ї¬†–њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є –≤–Њ–і—Л –і–ї—П –і—Л—Е–∞–љ–Є—П –Є–ї–Є –њ–Њ–і–і–µ—А–ґ–Є–≤–∞—В—М¬†–≥–Њ–ї–Њ–≤—Г (–љ–Њ–Ј–і—А–Є) –љ–∞–і –≤–Њ–і–Њ–є. –Т–Њ-–≤—В–Њ—А—Л—Е, –≤¬†–≤–Њ–і–µ –љ–µ—В —Г–±–µ–ґ–Є—Й –і–ї—П —Б–љ–∞, –∞¬†–≤—Б–њ–ї—Л—В–Є–µ –Ї¬†–њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є –і–µ–ї–∞–µ—В –ґ–Є–≤–Њ—В–љ—Л—Е –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ —Г—П–Ј–≤–Є–Љ—Л–Љ–Є –і–ї—П —Е–Є—Й–љ–Є–Ї–Њ–≤ (–Ї–Њ—Б–∞—В–Њ–Ї, –∞–Ї—Г–ї). –Ч–љ–∞—З–Є—В, –≤–Њ –≤—А–µ–Љ—П —Б–љ–∞ –ґ–Є–≤–Њ—В–љ—Л–µ –і–Њ–ї–ґ–љ—Л –њ–Њ—Б—В–Њ—П–љ–љ–Њ –њ–Њ–і–і–µ—А–ґ–Є–≤–∞—В—М –≤—Л—Б–Њ–Ї–Є–є —Г—А–Њ–≤–µ–љ—М –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є. –Т-—В—А–µ—В—М–Є—Е, —В–µ–њ–ї–Њ–њ—А–Њ–≤–Њ–і–љ–Њ—Б—В—М –≤–Њ–і—Л –≤¬†–і–µ—Б—П—В–Ї–Є —А–∞–Ј –≤—Л—И–µ —В–µ–њ–ї–Њ–њ—А–Њ–≤–Њ–і–љ–Њ—Б—В–Є –≤–Њ–Ј–і—Г—Е–∞.
–Т–∞–ґ–љ–µ–є—И–Є–µ –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В–Є —Б–љ–∞ –Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л—Е¬†вАУ –Њ–і–љ–Њ–њ–Њ–ї—Г—И–∞—А–љ—Л–є –Ь–° –Є¬†—Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М —Б–њ–∞—В—М –≤–Њ –≤—А–µ–Љ—П –і–≤–Є–ґ–µ–љ–Є—П, –Ј–∞–Ї—А—Л–≤–∞—П —В–Њ–ї—М–Ї–Њ –Њ–і–Є–љ¬†–≥–ї–∞–Ј [59вАУ62]. –°—З–Є—В–∞–µ—В—Б—П, —З—В–Њ –Є–Љ–µ–љ–љ–Њ –Њ–і–љ–Њ–њ–Њ–ї—Г—И–∞—А–љ—Л–є —Б–Њ–љ (–±—Г–і—Г—З–Є –њ—А–Є —Н—В–Њ–Љ –Є¬†–Њ–і–љ–Њ–њ–Њ–ї—Г—И–∞—А–љ—Л–Љ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ–Љ) –њ–Њ–Ј–≤–Њ–ї—П–µ—В –Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л–Љ —Б–Њ—З–µ—В–∞—В—М —Б–Њ–љ –Є¬†–і–≤–Є–ґ–µ–љ–Є–µ, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—П –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –≤—Б–њ–ї—Л—В–Є—П –Ї¬†–њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є –≤–Њ–і—Л –і–ї—П –і—Л—Е–∞–љ–Є—П –±–µ–Ј –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П. –Ф–≤–Є–ґ–µ–љ–Є–µ —В–∞–Ї–ґ–µ —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В –Љ—Л—И–µ—З–љ–Њ–Љ—Г —В–µ—А–Љ–Њ–≥–µ–љ–µ–Ј—Г, –њ—А–µ–њ—П—В—Б—В–≤—Г—П –њ–µ—А–µ–Њ—Е–ї–∞–ґ–і–µ–љ–Є—О –≤¬†—Е–Њ–ї–Њ–і–љ–Њ–є –≤–Њ–і–µ. –Т—В–Њ—А–∞—П —Д—Г–љ–Ї—Ж–Є—П –Њ–і–љ–Њ–њ–Њ–ї—Г—И–∞—А–љ–Њ–≥–Њ —Б–љ–∞ —Г¬†–Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л—Е —Б–Њ—Б—В–Њ–Є—В –≤¬†–≤–Є–Ј—Г–∞–ї—М–љ–Њ–Љ –Ї–Њ–љ—В—А–Њ–ї–µ –Ј–∞ —Б–Њ—Б—В–Њ—П–љ–Є–µ–Љ –Њ–Ї—А—Г–ґ–∞—О—Й–µ–є —Б—А–µ–і—Л —Б¬†–њ–Њ–Љ–Њ—Й—М—О –Њ–і–љ–Њ–≥–Њ –Њ—В–Ї—А—Л—В–Њ–≥–Њ –≤–Њ –≤—А–µ–Љ—П —Б–љ–∞¬†–≥–ї–∞–Ј–∞. –≠—В–Њ —Б–љ–Є–ґ–∞–µ—В —А–Є—Б–Ї¬†–љ–µ–Њ–ґ–Є–і–∞–љ–љ–Њ–≥–Њ –љ–∞–њ–∞–і–µ–љ–Є—П —Е–Є—Й–љ–Є–Ї–Њ–≤ –Є¬†–њ–Њ–Љ–Њ–≥–∞–µ—В –Ї–Њ–Њ—А–і–Є–љ–Є—А–Њ–≤–∞—В—М –і–≤–Є–ґ–µ–љ–Є–µ —Б¬†–і—А—Г–≥–Є–Љ–Є –Њ—Б–Њ–±—П–Љ–Є –≤¬†–≥—А—Г–њ–њ–µ (–љ–∞–њ—А–Є–Љ–µ—А, –Љ–∞—В–µ—А–µ–є —Б¬†–і–µ—В–µ–љ—Л—И–∞–Љ–Є [60вАУ63]).
–Я—А–Є—З–Є–љ—Л –Њ—В—Б—Г—В—Б—В–≤–Є—П –Я–° —Г¬†–Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л—Е (–≤–Њ –≤—Б—П–Ї–Њ–Љ —Б–ї—Г—З–∞–µ –≤¬†—В–Њ–Љ –≤¬†–≤–Є–і–µ, –≤¬†–Ї–∞–Ї–Њ–Љ –Њ–љ —А–µ–≥–Є—Б—В—А–Є—А—Г–µ—В—Б—П —Г¬†–љ–∞–Ј–µ–Љ–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е) –љ–µ¬†–њ–Њ–љ—П—В–љ—Л [61, 62]. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П —Г¬†–Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л—Е –µ—Б—В—М —Н–ї–µ–Љ–µ–љ—В—Л –њ–Њ–≤–µ–і–µ–љ–Є—П (–≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є, –≤–Ј–і—А–∞–≥–Є–≤–∞–љ–Є—П –≤–Њ –≤—А–µ–Љ—П –Ь–°), –Ї–Њ—В–Њ—А—Л–µ –љ–∞–њ–Њ–Љ–Є–љ–∞—О—В –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–µ –њ—А–Є–Ј–љ–∞–Ї–Є –Я–° —Г¬†–љ–∞–Ј–µ–Љ–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е. –Э–µ¬†–Є—Б–Ї–ї—О—З–µ–љ–Њ, —З—В–Њ –Я–° —Г¬†–і–µ–ї—М—Д–Є–љ–Њ–≤ –Є¬†–Ї–Є—В–Њ–≤ –Љ–Њ–ґ–µ—В –њ—А–Њ—В–µ–Ї–∞—В—М –≤¬†–Љ–Њ–і–Є—Д–Є—Ж–Є—А–Њ–≤–∞–љ–љ–Њ–є —Д–Њ—А–Љ–µ (–љ–∞–њ—А–Є–Љ–µ—А, –≤¬†–≤–Є–і–µ –Ї–Њ—А–Њ—В–Ї–Є—Е —Н–њ–Є–Ј–Њ–і–Њ–≤ –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М—О –≤—Б–µ–≥–Њ –≤¬†–љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —Б–µ–Ї—Г–љ–і), —З—В–Њ –Ј–∞—В—А—Г–і–љ—П–µ—В –µ–≥–Њ –Є–і–µ–љ—В–Є—Д–Є–Ї–∞—Ж–Є—О. –Ф–∞–ґ–µ –≤¬†—Н—В–Њ–Љ —Б–ї—Г—З–∞–µ –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М –Я–° —Г¬†–Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л—Е –±—Л–ї–∞ –±—Л –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –Љ–µ–љ—М—И–µ, —З–µ–Љ —Г¬†–љ–∞–Ј–µ–Љ–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж. –°—З–Є—В–∞–µ—В—Б—П, —З—В–Њ –Я–° –≤—Л–њ–Њ–ї–љ—П–µ—В –≤–Є—В–∞–ї—М–љ—Г—О (–ґ–Є–Ј–љ–µ–њ–Њ–і–і–µ—А–ґ–Є–≤–∞—О—Й—Г—О) —Д—Г–љ–Ї—Ж–Є—О. –Т¬†—З–µ–Љ –±—Л —Н—В–∞ —Д—Г–љ–Ї—Ж–Є—П –љ–Є¬†—Б–Њ—Б—В–Њ—П–ї–∞, –Њ–љ–∞ –Њ–Ї–∞–Ј—Л–≤–∞–µ—В—Б—П –Љ–µ–љ–µ–µ –≤–∞–ґ–љ–Њ–є –і–ї—П –Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л—Е –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†—В–µ–Љ–Є —А–Є—Б–Ї–∞–Љ–Є, –Ї–Њ—В–Њ—А—Л–µ –≤–Њ–Ј–љ–Є–Ї–∞—О—В –њ—А–Є —А–∞–Ј–≤–Є—В–Є–Є –і–ї–Є–љ–љ—Л—Е —Н–њ–Є–Ј–Њ–і–Њ–≤ –Я–°.
–Ы–∞—Б—В–Њ–љ–Њ–≥–Є–µ¬†вАУ¬†–≥—А—Г–њ–њ–∞ –њ–Њ–ї—Г–≤–Њ–і–љ—Л—Е –ґ–Є–≤–Њ—В–љ—Л—Е (–Њ—В—А—П–і —Е–Є—Й–љ—Л—Е), –Ї–Њ—В–Њ—А—Л–µ –Љ–Њ–≥—Г—В —Б–њ–∞—В—М –Є¬†–љ–∞ —Б—Г—И–µ, –Є¬†–≤ –≤–Њ–і–µ. –°–Њ–љ –ї–∞—Б—В–Њ–љ–Њ–≥–Є—Е —Б–Њ—З–µ—В–∞–µ—В –њ—А–Є–Ј–љ–∞–Ї–Є —Б–љ–∞ –љ–∞–Ј–µ–Љ–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л—Е. –Ю–і–Є–љ –Є–Ј¬†—В–Є–њ–Є—З–љ—Л—Е –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–µ–є –ї–∞—Б—В–Њ–љ–Њ–≥–Є—Е, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є —Б–µ–Љ–µ–є—Б—В–≤–∞ —Г—И–∞—Б—В—Л—Е —В—О–ї–µ–љ–µ–є, –Є¬†–Њ–і–љ–∞ –Є–Ј¬†–Љ–Њ–і–µ–ї–µ–є –і–ї—П –Є–Ј—Г—З–µ–љ–Є—П –Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤ —Б–љ–∞ —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е¬†вАУ —Б–µ–≤–µ—А–љ—Л–є –Љ–Њ—А—Б–Ї–Њ–є –Ї–Њ—В–Є–Ї. –Ь–Њ—А—Б–Ї–Є–µ –Ї–Њ—В–Є–Ї–Є –Њ–±–Є—В–∞—О—В –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –≤¬†—Г–Љ–µ—А–µ–љ–љ–Њ–є –Є¬†—Б—Г–±—В—А–Њ–њ–Є—З–µ—Б–Ї–Њ–є –Ї–ї–Є–Љ–∞—В–Є—З–µ—Б–Ї–Є—Е –Ј–Њ–љ–∞—Е. –Т–Њ –≤—А–µ–Љ—П –ї–µ—В–љ–µ–≥–Њ –њ–µ—А–Є–Њ–і–∞ –Њ–љ–Є –њ—А–Њ–≤–Њ–і—П—В –±–Њ–ї—М—И—Г—О —З–∞—Б—В—М –≤—А–µ–Љ–µ–љ–Є –љ–∞¬†—Б—Г—И–µ,¬†–≥–і–µ —А–∞–Ј–Љ–љ–Њ–ґ–∞—О—В—Б—П –Є¬†–ї–Є–љ—П—О—В. –Т¬†–Ј–Є–Љ–љ–Є–є –њ–µ—А–Є–Њ–і –Ї–Њ—В–Є–Ї–Є –Љ–Є–≥—А–Є—А—Г—О—В –љ–∞¬†—А–∞—Б—Б—В–Њ—П–љ–Є–µ –і–Њ 2000¬†–Ї–Љ –Є¬†–љ–µ –≤—Л—Е–Њ–і—П—В –љ–∞¬†—Б—Г—И—Г –і–Њ –і–µ—Б—П—В–Є –Љ–µ—Б—П—Ж–µ–≤. –Т¬†–≤–Њ–і–µ –Њ–љ–Є —Б–њ—П—В –љ–∞¬†–њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є –≤¬†—Е–∞—А–∞–Ї—В–µ—А–љ–Њ–є –њ–Њ–Ј–µ, –њ–Њ–і–і–µ—А–ґ–Є–≤–∞—П¬†–≥–Њ–ї–Њ–≤—Г –љ–∞–і –≤–Њ–і–Њ–є –Є¬†—Б–Њ–≤–µ—А—И–∞—П¬†–≥—А–µ–±–Ї–Њ–≤—Л–µ –і–≤–Є–ґ–µ–љ–Є—П –Њ–і–љ–Є–Љ –њ–µ—А–µ–і–љ–Є–Љ –ї–∞—Б—В–Њ–Љ (—А–Є—Б.¬†2). –Ґ–∞–Ї–∞—П –њ–Њ–Ј–∞ –њ–Њ–Ј–≤–Њ–ї—П–µ—В —Б–љ–Є–Ј–Є—В—М –њ–Њ—В–µ—А–Є —В–µ–њ–ї–∞ (—З–µ—А–µ–Ј –љ–∞—Е–Њ–і—П—Й–Є–µ—Б—П –≤¬†–≤–Њ–Ј–і—Г—Е–µ –ї–∞—Б—В—Л) –Є¬†–Њ–±–µ—Б–њ–µ—З–Є–≤–∞–µ—В —А–µ–≥—Г–ї—П—А–љ–Њ–µ –і—Л—Е–∞–љ–Є–µ. –Т–Њ –≤—А–µ–Љ—П —Б–љ–∞ –≤¬†–≤–Њ–і–µ –Ї–Њ—В–Є–Ї–Є, –Ї–∞–Ї –Є¬†–Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л–µ, –Ј–∞–Ї—А—Л–≤–∞—О—В –љ–∞¬†–њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ–µ –≤—А–µ–Љ—П —В–Њ–ї—М–Ї–Њ –Њ–і–Є–љ¬†–≥–ї–∞–Ј, —В–Њ–≥–і–∞ –Ї–∞–Ї –≤—В–Њ—А–Њ–є –љ–∞–њ—А–∞–≤–ї–µ–љ –≤¬†—Б—В–Њ—А–Њ–љ—Г –≤–Њ–і—Л, —З—В–Њ –њ–Њ–≤—Л—И–∞–µ—В –≤–µ—А–Њ—П—В–љ–Њ—Б—В—М –Њ–±–љ–∞—А—Г–ґ–µ–љ–Є—П —Е–Є—Й–љ–Є–Ї–Њ–≤ [63вАУ65].
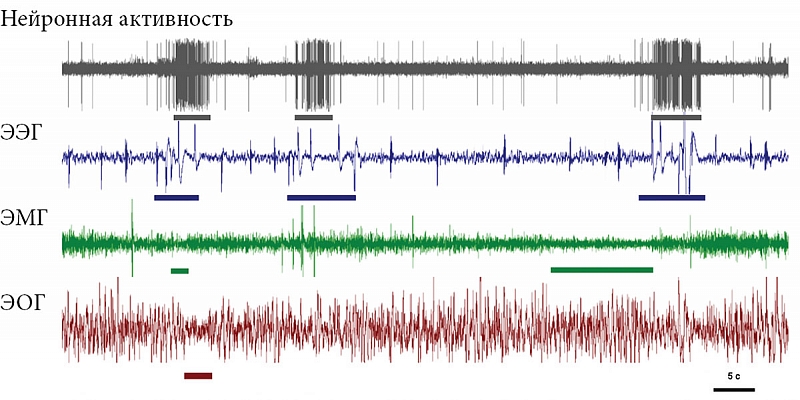
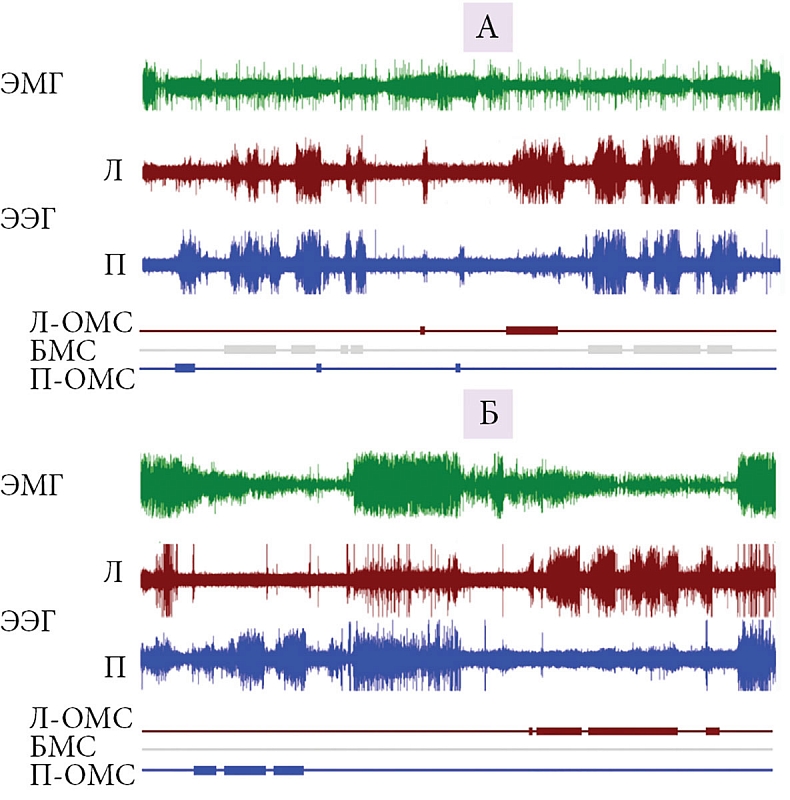
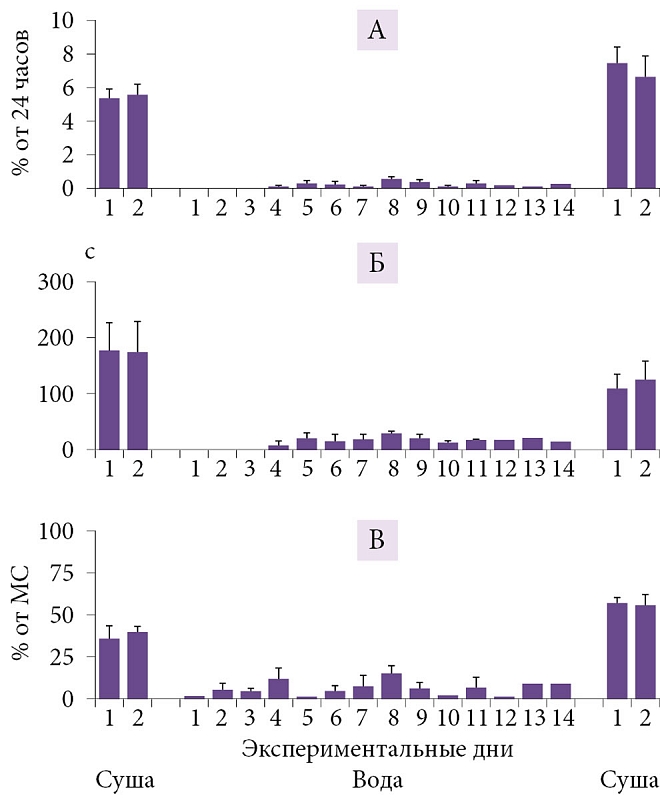
–Э–∞—И–Є –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ –њ–µ—А–µ—Е–Њ–і –Њ—В¬†–љ–∞–Ј–µ–Љ–љ–Њ–≥–Њ –Ї¬†–≤–Њ–і–љ–Њ–Љ—Г –Њ–±—А–∞–Ј—Г –ґ–Є–Ј–љ–Є —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–µ—В—Б—П —Г¬†–Ї–Њ—В–Є–Ї–Њ–≤ ¬Ђ–њ–µ—А–µ–Ї–ї—О—З–µ–љ–Є–µ–Љ¬ї —Б¬†¬Ђ—В–Є–њ–Є—З–љ–Њ–≥–Њ –љ–∞–Ј–µ–Љ–љ–Њ–≥–Њ¬ї (–±–Є–ї–∞—В–µ—А–∞–ї—М–љ–Њ-—Б–Є–Љ–Љ–µ—В—А–Є—З–љ—Л–є –Ь–°, –љ–µ–њ–Њ–і–≤–Є–ґ–љ–Њ—Б—В—М, –Я–°) –љ–∞¬†—Е–∞—А–∞–Ї—В–µ—А–љ—Л–є –і–ї—П –Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л—Е ¬Ђ–≤–Њ–і–љ—Л–є¬ї —В–Є–њ —Б–љ–∞ (–Њ–і–љ–Њ–њ–Њ–ї—Г—И–∞—А–љ—Л–є —Б–Њ–љ, —Б–Њ—Е—А–∞–љ–µ–љ–Є–µ –і–≤–Є–≥–∞—В–µ–ї—М–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є, –њ—А–µ—А—Л–≤–Є—Б—В—Л–є —В–Є–њ –і—Л—Е–∞–љ–Є—П, –Њ—В—Б—Г—В—Б—В–≤–Є–µ –Я–°) (—А–Є—Б.¬†3 –Є¬†4) [65, 66]). –°–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –Ї–Њ—В–Є–Ї–Њ–≤ –µ—Б—В–µ—Б—В–≤–µ–љ–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ —Д–∞–Ї—В–Є—З–µ—Б–Ї–Є –њ–Њ–ї–љ–Њ—Б—В—М—О –Є—Б–Ї–ї—О—З–Є—В—М –Я–° –Є–Ј¬†–ґ–Є–Ј–љ–Є –љ–∞¬†–Ї–∞–Ї –Љ–Є–љ–Є–Љ—Г–Љ –Њ–і–љ—Г-–і–≤–µ –љ–µ–і–µ–ї–Є –±–µ–Ј –Ї–∞–Ї–Є—Е –±—Л —В–Њ –љ–Є¬†–±—Л–ї–Њ –Њ—З–µ–≤–Є–і–љ—Л—Е –њ–Њ—Б–ї–µ–і—Б—В–≤–Є–є –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В –±–Њ–ї—М—И–Њ–є –Є–љ—В–µ—А–µ—Б –і–ї—П –њ–Њ–љ–Є–Љ–∞–љ–Є—П —Д—Г–љ–Ї—Ж–Є–Є –Є¬†–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є —А–Њ–ї–Є –Я–°. –Ш—Б—З–µ–Ј–љ–Њ–≤–µ–љ–Є–µ –Я–° –њ—А–Є –њ–µ—А–µ—Е–Њ–і–µ –Њ—В¬†—Б–љ–∞ –љ–∞¬†—Б—Г—И–µ –Ї–Њ¬†—Б–љ—Г –≤¬†–≤–Њ–і–µ¬†–≥–Њ–≤–Њ—А–Є—В –Њ¬†–≤–∞–ґ–љ–Њ—Б—В–Є —Н–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤ –≤¬†—А–µ–≥—Г–ї—П—Ж–Є–Є –Я–° —Г¬†—Н—В–Њ–≥–Њ –≤–Є–і–∞. –Я—А–Є —Н—В–Њ–Љ –Є—Б—З–µ–Ј–љ–Њ–≤–µ–љ–Є–µ –Я–° –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е, –Ї–Њ–≥–і–∞ –ґ–Є–≤–Њ—В–љ—Л–Љ –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ –њ–Њ–і–і–µ—А–ґ–Є–≤–∞—В—М –≤—Л—Б–Њ–Ї–Є–є —Г—А–Њ–≤–µ–љ—М –і–≤–Є–≥–∞—В–µ–ї—М–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –Є¬†–±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є. –Э–∞—Е–Њ–і—П—Б—М –≤¬†–≤–Њ–і–µ –і–≤–µ –љ–µ–і–µ–ї–Є, –Љ–Њ—А—Б–Ї–Є–µ –Ї–Њ—В–Є–Ї–Є –љ–∞–Ї–∞–њ–ї–Є–≤–∞—О—В –Њ–≥—А–Њ–Љ–љ—Л–є –і–µ—Д–Є—Ж–Є—В –Я–° –≤¬†–Ї–Њ–ї–Є—З–µ—Б—В–≤–µ 15вАУ20 —З–∞—Б–Њ–≤. –Я–Њ—Б–ї–µ –≤–Њ–Ј–≤—А–∞—Й–µ–љ–Є—П –љ–∞¬†—Б—Г—И—Г —Г¬†–љ–Є—Е –љ–µ¬†–љ–∞–±–ї—О–і–∞–µ—В—Б—П –Ј–љ–∞—З–Є–Љ–Њ–≥–Њ –њ—А–µ–≤—Л—И–µ–љ–Є—П –Я–° –љ–∞–і –Ї–Њ–љ—В—А–Њ–ї—М–љ—Л–Љ–Є –Ј–љ–∞—З–µ–љ–Є—П–Љ–Є –љ–∞¬†—Б—Г—И–µ. –Я–Њ–ї—Г—З–µ–љ–љ—Л–µ –і–∞–љ–љ—Л–µ —Б—В–∞–≤—П—В –њ–Њ–і —Б–Њ–Љ–љ–µ–љ–Є–µ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є—П –Њ¬†–≥–Њ–Љ–µ–Њ—Б—В–∞—В–Є—З–µ—Б–Ї–Њ–є —А–µ–≥—Г–ї—П—Ж–Є–Є –Я–°. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П –Њ–љ–Є —Е–Њ—А–Њ—И–Њ —Б–Њ–≥–ї–∞—Б—Г—О—В—Б—П —Б¬†—А–∞–љ–µ–µ –≤—Л—Б–Ї–∞–Ј–∞–љ–љ–Њ–є¬†–≥–Є–њ–Њ—В–µ–Ј–Њ–є, —З—В–Њ –Я–° –љ–µ–Њ–±—Е–Њ–і–Є–Љ –і–ї—П —Б–Љ—П–≥—З–µ–љ–Є—П –≤–Њ–Ј–Љ–Њ–ґ–љ—Л—Е –љ–µ–≥–∞—В–Є–≤–љ—Л—Е –њ–Њ—Б–ї–µ–і—Б—В–≤–Є–є¬†–≥–ї—Г–±–Њ–Ї–Њ–≥–Њ –±–Є–ї–∞—В–µ—А–∞–ї—М–љ–Њ-—Б–Є–Љ–Љ–µ—В—А–Є—З–љ–Њ–≥–Њ –Ь–° (–љ–∞–њ—А–Є–Љ–µ—А, —Б–љ–Є–ґ–µ–љ–Є—П —В–µ–Љ–њ–µ—А–∞—В—Г—А—Л –Љ–Њ–Ј–≥–∞ [67]). –≠—В–Њ –Љ–Њ–≥–ї–Њ –±—Л –Њ–±—К—П—Б–љ–Є—В—М –љ–µ¬†—В–Њ–ї—М–Ї–Њ —Б–Њ–Ї—А–∞—Й–µ–љ–Є–µ –Я–° –≤¬†–≤–Њ–і–µ —Г¬†–Ї–Њ—В–Є–Ї–Њ–≤ (–њ–∞—А–∞–ї–ї–µ–ї—М–љ–Њ —Б¬†–Є—Б—З–µ–Ј–љ–Њ–≤–µ–љ–Є–µ–Љ –±–Є–ї–∞—В–µ—А–∞–ї—М–љ–Њ-—Б–Є–Љ–Љ–µ—В—А–Є—З–љ–Њ–≥–Њ –Ь–°), –љ–Њ¬†–Є –Њ—В—Б—Г—В—Б—В–≤–Є–µ –Я–° —Г¬†–і–µ–ї—М—Д–Є–љ–Њ–≤ (—Г¬†–Ї–Њ—В–Њ—А—Л—Е –≤—Л—Б–Њ–Ї–Њ–∞–Љ–њ–ї–Є—В—Г–і–љ—Л–є –±–Є–ї–∞—В–µ—А–∞–ї—М–љ–Њ-—Б–Є–Љ–Љ–µ—В—А–Є—З–љ—Л–є –Ь–° –љ–µ¬†—А–µ–≥–Є—Б—В—А–Є—А—Г–µ—В—Б—П). –Ю—Б–Њ–±–µ–љ–љ–Њ—Б—В–Є —Б–љ–∞ –Љ–Њ—А—Б–Ї–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ –Ї–∞–Ї–Є–Љ-—В–Њ –Њ–±—А–∞–Ј–Њ–Љ —Б–Њ–≥–ї–∞—Б–Њ–≤–∞—В—М —Б¬†–њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є–µ–Љ –Њ¬†—А–Њ–ї–Є –Я–° –≤¬†–њ–Њ–і–і–µ—А–ґ–∞–љ–Є–Є –Ї–Њ–≥–љ–Є—В–Є–≤–љ—Л—Е —Д—Г–љ–Ї—Ж–Є–є, –Є–ї–Є —Б–ї–µ–і—Г–µ—В –њ—А–Є–Ј–љ–∞—В—М, —З—В–Њ —Н—В–∞ —Б—В–∞–і–Є—П —Б–љ–∞ –≤—Л–њ–Њ–ї–љ—П–µ—В —А–∞–Ј–ї–Є—З–љ—Л–µ —Д—Г–љ–Ї—Ж–Є–Є —Г¬†—А–∞–Ј–љ—Л—Е –≤–Є–і–Њ–≤ –Є¬†–≥—А—Г–њ–њ.
–Ю—В—Б—Г—В—Б—В–≤–Є–µ/—А–µ–Ј–Ї–Њ–µ —Б–Њ–Ї—А–∞—Й–µ–љ–Є–µ –Я–° —Г¬†–і–µ–ї—М—Д–Є–љ–Њ–≤ –Є¬†–Љ–Њ—А—Б–Ї–Є—Е –Ї–Њ—В–Є–Ї–Њ–≤ –≤–Њ–≤—Б–µ –љ–µ¬†–Њ–Ј–љ–∞—З–∞–µ—В, —З—В–Њ –Я–° –љ–µ—Б–Њ–≤–Љ–µ—Б—В–Є–Љ —Б¬†–≤–Њ–і–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ –ґ–Є–Ј–љ–Є, –∞¬†–Њ–і–љ–Њ–њ–Њ–ї—Г—И–∞—А–љ—Л–є –Ь–° –љ–∞¬†—Д–Њ–љ–µ –њ–Њ—Б—В–Њ—П–љ–љ–Њ–є –і–≤–Є–≥–∞—В–µ–ї—М–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є¬†вАУ –µ–і–Є–љ—Б—В–≤–µ–љ–љ–∞—П –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –і–ї—П –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е —Б–њ–∞—В—М –≤¬†–≤–Њ–і–љ–Њ–є —Б—А–µ–і–µ. –°—Г—Й–µ—Б—В–≤—Г–µ—В –Є¬†–і—А—Г–≥–Њ–є –њ–∞—В—В–µ—А–љ (—Б—В—А–∞—В–µ–≥–Є—П) —Б–љ–∞, –Ї–Њ—В–Њ—А—Л–є —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї—Б—П –≤¬†—Н–≤–Њ–ї—О—Ж–Є–Є —Г¬†–љ–∞—Б—В–Њ—П—Й–Є—Е —В—О–ї–µ–љ–µ–є. –С–Њ–ї—М—И–Є–љ—Б—В–≤–Њ –љ–∞—Б—В–Њ—П—Й–Є—Е —В—О–ї–µ–љ–µ–є, –∞¬†—В–∞–Ї–ґ–µ –Љ–Њ—А–ґ–Є –Њ–±–Є—В–∞—О—В –≤¬†—Е–Њ–ї–Њ–і–љ—Л—Е –≤–Њ–і–∞—Е —Б—Г–±–њ–Њ–ї—П—А–љ—Л—Е –Є¬†–њ–Њ–ї—П—А–љ—Л—Е —А–∞–є–Њ–љ–Њ–≤ –≤–і–Њ–ї—М –Ї—А–Њ–Љ–Ї–Є –ї—М–і–Њ–≤ –Є–ї–Є –≤¬†–Ј–∞–Љ–µ—А–Ј–∞—О—Й–Є—Е –Њ–Ј–µ—А–∞—Е –Є¬†–≤–љ—Г—В—А–µ–љ–љ–Є—Е –Љ–Њ—А—П—Е –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е –Њ–≥—А–∞–љ–Є—З–µ–љ–љ–Њ–≥–Њ –і–Њ—Б—В—Г–њ–∞ –Ї¬†–Њ—В–Ї—А—Л—В–Њ–є –≤–Њ–і–µ. –Ч–∞–і–µ—А–ґ–Є–≤–∞—П –і—Л—Е–∞–љ–Є–µ, —В—О–ї–µ–љ–Є –Љ–Њ–≥—Г—В —Б–њ–∞—В—М –њ–Њ–і –≤–Њ–і–Њ–є –љ–∞¬†–≥–ї—Г–±–Є–љ–µ [68]. –Я–Њ–і–Њ–±–љ—Л–є –њ–∞—В—В–µ—А–љ —Б–љ–∞ –љ–µ¬†—В—А–µ–±—Г–µ—В –њ–Њ–і–і–µ—А¬≠–ґ–∞–љ–Є—П –і–≤–Є–≥–∞—В–µ–ї—М–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є. –Я–Њ—Н—В–Њ–Љ—Г –Ь–° —Г¬†—В—О–ї–µ–љ–µ–є –±–Є–ї–∞—В–µ—А–∞–ї—М–љ–Њ-—Б–Є–Љ–Љ–µ—В—А–Є—З–љ—Л–є, –Ї–∞–Ї —Г¬†–љ–µ–њ–Њ–і–≤–Є–ґ–љ–Њ —Б–њ—П—Й–Є—Е –љ–∞–Ј–µ–Љ–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е. –°–Њ—Е—А–∞–љ–µ–љ–Є–µ —В–µ–њ–ї–∞ —Г¬†—В—О–ї–µ–љ–µ–є –і–Њ—Б—В–Є–≥–∞–µ—В—Б—П –њ–Њ—Б—А–µ–і—Б—В–≤–Њ–Љ —В–Њ–ї—Б—В–Њ–≥–Њ –њ–Њ–і–Ї–Њ–ґ–љ–Њ–≥–Њ –ґ–Є—А–Њ–≤–Њ–≥–Њ —Б–ї–Њ—П. –≠—Д—Д–µ–Ї—В–Є–≤–љ–∞—П –Ј–∞—Й–Є—В–∞ –Њ—В¬†–њ–µ—А–µ–Њ—Е–ї–∞–ґ–і–µ–љ–Є—П –љ–µ¬†–њ—А–µ–њ—П—В—Б—В–≤—Г–µ—В —А–∞–Ј–≤–Є—В–Є—О –Я–°. –Я—А–Є —Н—В–Њ–Љ –Њ–љ–Є –і–Њ–ї–ґ–љ—Л –њ—А–Њ–±—Г–ґ–і–∞—В—М—Б—П –і–ї—П –і–≤–Є–ґ–µ–љ–Є—П –њ—А–Є –≤—Б–њ–ї—Л—В–Є–Є –Ї¬†–њ–Њ–≤–µ—А—Е¬≠–љ–Њ—Б—В–Є –≤–Њ–і—Л [63].
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –Є–Љ–µ—О—Й–Є–µ—Б—П –і–∞–љ–љ—Л–µ –Њ–±¬†–Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В—П—Е —Б–љ–∞ –≤–Њ–і–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е —Г–Ї–∞–Ј—Л–≤–∞—О—В –љ–∞¬†–≤–∞–ґ–љ–Њ—Б—В—М —Н–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤ –≤¬†—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–Є –њ–∞—В—В–µ—А–љ–∞ —Б–љ–∞. –Э–µ–Њ–±—Е–Њ–і–Є–Љ–Њ—Б—В—М –≤—Б–њ–ї—Л–≤–∞—В—М –Ї¬†–њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є –≤–Њ–і—Л –і–ї—П –і—Л—Е–∞–љ–Є—П, –њ–Њ—Б—В–Њ—П–љ–љ–Њ –Ї–Њ–љ—В—А–Њ–ї–Є—А–Њ–≤–∞—В—М –Њ–Ї—А—Г–ґ–∞—О—Й—Г—О —Б—А–µ–і—Г –Є¬†–Њ—Б—Г—Й–µ—Б—В–≤–ї—П—В—М —Н—Д—Д–µ–Ї—В–Є–≤–љ—Г—О —В–µ—А–Љ–Њ—А–µ–≥—Г–ї—П—Ж–Є—О¬†вАУ –Њ—Б–љ–Њ–≤–љ—Л–µ —Д–∞–Ї—В–Њ—А—Л, –Њ–њ—А–µ–і–µ–ї–Є–≤—И–Є–µ —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–µ —Б—В—А—Г–Ї—В—Г—А—Л —Б–љ–∞ —Г¬†—Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л—Е –≤–Њ–і–љ—Л—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е [61вАУ66]. –Ю–і–љ–Њ–њ–Њ–ї—Г—И–∞—А–љ—Л–є –Ь–°¬†вАУ –Њ–і–љ–∞ –Є–Ј¬†–≥–ї–∞–≤–љ—Л—Е –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В–µ–є —Б–љ–∞ –Ї–Є—В–Њ–Њ–±—А–∞–Ј–љ—Л—Е –Є¬†—Г—И–∞—Б—В—Л—Е —В—О–ї–µ–љ–µ–є —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–∞—Б—М –≤¬†—Н–≤–Њ–ї—О—Ж–Є–Є —Н—В–Є—Е¬†–≥—А—Г–њ–њ –љ–µ–Ј–∞–≤–Є—Б–Є–Љ–Њ –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е –і–µ–є—Б—В–≤–Є—П –Њ–і–љ–Є—Е –Є¬†—В–µ—Е –ґ–µ —Н–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤. –І—А–µ–Ј–≤—Л—З–∞–є–љ–Њ –Є–љ—В–µ—А–µ—Б–љ–Њ, —З—В–Њ –Њ–і–љ–Њ–њ–Њ–ї—Г—И–∞—А–љ—Л–є –Ь–° —Г¬†–Љ–Њ—А—Б–Ї–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж —Д–µ–љ–Њ—В–Є–њ–Є—З–µ—Б–Ї–Є –њ–Њ—Е–Њ–ґ, –љ–µ—Б–Љ–Њ—В—А—П –љ–∞¬†—А–∞–Ј–ї–Є—З–Є—П –≤¬†–∞–љ–∞—В–Њ–Љ–Є—З–µ—Б–Ї–Њ–є –Њ—А–≥–∞–љ–Є–Ј–∞—Ж–Є–Є –Є—Е –њ–µ—А–µ–і–љ–µ–≥–Њ –Љ–Њ–Ј–≥–∞. –Э–µ¬†–Є—Б–Ї–ї—О—З–µ–љ–Њ, —З—В–Њ –≤¬†–±—Г–і—Г—Й–µ–Љ —В–∞–Ї–∞—П —Д–Њ—А–Љ–∞ —Б–љ–∞ –Њ–±–љ–∞—А—Г–ґ–Є—В—Б—П –Є¬†—Г –і—А—Г–≥–Є—Е –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–µ–є –ґ–Є–≤–Њ—В–љ–Њ–≥–Њ –Љ–Є—А–∞, –і–ї—П –Ї–Њ—В–Њ—А—Л—Е —Г—Б–ї–Њ–≤–Є—П —Б–љ–∞ —В—А–µ–±—Г—О—В –њ–Њ–і–і–µ—А–ґ–∞–љ–Є—П –≤—Л—Б–Њ–Ї–Њ–≥–Њ —Г—А–Њ–≤–љ—П –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є –Є–ї–Є —Б–Њ—Е—А–∞–љ–µ–љ–Є—П –і–≤–Є–≥–∞—В–µ–ї—М–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –≤–Њ –≤—А–µ–Љ—П —Б–љ–∞.
–Ч–∞–Ї–ї—О—З–µ–љ–Є–µ
–Э–∞–Є–±–Њ–ї–µ–µ –Є–љ—В–µ—А–µ—Б–љ—Л–µ –Є¬†–Ј–љ–∞—З–Є–Љ—Л–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л –≤¬†–љ–µ–є—А–Њ–±–Є–Њ–ї–Њ–≥–Є–Є —Б–љ–∞ (—Б–Њ–Љ–љ–Њ–ї–Њ–≥–Є–Є) –њ–Њ—Б–ї–µ–і–љ–Є—Е 20 –ї–µ—В –±—Л–ї–Є –њ–Њ–ї—Г—З–µ–љ—Л –њ—А–Є –Є–Ј—Г—З–µ–љ–Є–Є —Б–љ–∞ —Г¬†–њ—А–Њ—Б—В—Л—Е –Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤, –∞¬†—В–∞–Ї–ґ–µ –њ—А–Є –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є —Б–љ–∞ –ґ–Є–≤–Њ—В–љ—Л—Е –≤¬†–њ—А–Є—А–Њ–і–µ. –Ю–Ї–∞–Ј–∞–ї–Њ—Б—М, —З—В–Њ —Б–Њ–љ¬†вАУ —Г–љ–Є–≤–µ—А—Б–∞–ї—М–љ–Њ–µ —Б–Њ—Б—В–Њ—П–љ–Є–µ –≤—Б–µ—Е –ґ–Є–≤–Њ—В–љ—Л—Е, –Њ—В¬†–њ—А–Њ—Б—В—Л—Е —Д–Њ—А–Љ —Б¬†–і–µ—Ж–µ–љ—В—А–∞–ї–Є–Ј–Њ–≤–∞–љ–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ–Њ–є –і–Њ –≤—Л—Б—И–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е —Б–Њ¬†—Б–ї–Њ–ґ–љ–Њ —Г—Б—В—А–Њ–µ–љ–љ—Л–Љ¬†–≥–Њ–ї–Њ–≤–љ—Л–Љ –Љ–Њ–Ј–≥–Њ–Љ. –£¬†–≤—Б–µ—Е –ґ–Є–≤–Њ—В–љ—Л—Е —Б–Њ—Б—В–Њ—П–љ–Є–µ —Б–љ–∞ —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г–µ—В—Б—П —Г–і–Є–≤–Є—В–µ–ї—М–љ–Њ –њ–Њ—Е–Њ–ґ–Є–Љ–Є –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–Љ–Є, –љ–µ–є—А–Њ–љ–љ—Л–Љ–Є, –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–Љ–Є –Є¬†—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ–Є –њ—А–Є–Ј–љ–∞–Ї–∞–Љ–Є. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П –і–≤–µ –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–љ—Л–µ —Б—В–∞–і–Є–Є —Б–љ–∞¬†вАУ –Ь–° –Є¬†–Я–° (–Є–ї–Є –±—Л—Б—В—А—Л–є —Б–Њ–љ) —Б—Г—Й–µ—Б—В–≤—Г—О—В —В–Њ–ї—М–Ї–Њ —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж. –Я–Њ—Н—В–Њ–Љ—Г —Б—А–∞–≤–љ–Є—В–µ–ї—М–љ—Л–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П —Б–љ–∞ —Г¬†–Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –Є¬†–њ—В–Є—Ж –Њ—Б–Њ–±–µ–љ–љ–Њ –≤–∞–ґ–љ—Л –і–ї—П –Њ—Б–Љ—Л—Б–ї–µ–љ–Є—П —Н–≤–Њ–ї—О—Ж–Є–Є —Б–љ–∞ –Є¬†–Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤ –µ–≥–Њ —А–µ–≥—Г–ї—П—Ж–Є–Є. –Ф–ї—П –њ–Њ–љ–Є–Љ–∞–љ–Є—П –њ—А–Є—А–Њ–і—Л –Є¬†—Д—Г–љ–Ї—Ж–Є–Є (—Д—Г–љ–Ї—Ж–Є–є) —Б–љ–∞ –Є¬†–µ–≥–Њ —Б—В–∞–і–Є–є –≤–∞–ґ–љ–Њ –Є–Ј—Г—З–∞—В—М —Б–Њ–љ –ґ–Є–≤–Њ—В–љ—Л—Е –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е, –њ—А–µ–і–µ–ї—М–љ–Њ –њ—А–Є–±–ї–Є–ґ–µ–љ–љ—Л—Е –Ї¬†–µ—Б—В–µ—Б—В–≤–µ–љ–љ—Л–Љ, –Є¬†–≤ –і–Є–Ї–Њ–є –њ—А–Є—А–Њ–і–µ. –Я—А–Є —Н—В–Њ–Љ –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ —Г—З–Є—В—Л–≤–∞—В—М –≤–ї–Є—П–љ–Є–µ –љ–∞¬†–њ–Њ–≤–µ–і–µ–љ–Є–µ –Є¬†—Б–Њ–љ –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ–≥–Њ —З–Є—Б–ї–∞ –≤–љ–µ—И–љ–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤, –і–Є–љ–∞–Љ–Є–Ї—Г –Є—Е –Є–Ј–Љ–µ–љ–µ–љ–Є–є –Є¬†–Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В–Є –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ —Ж–Є–Ї–ї–∞ –і–∞–љ–љ–Њ–≥–Њ –≤–Є–і–∞ –ґ–Є–≤–Њ—В–љ—Л—Е.¬†
–Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П —Б–љ–∞ –њ—В–Є—Ж –њ–Њ–і–і–µ—А–ґ–∞–љ—Л¬†–≥—А–∞–љ—В–Њ–Љ –†–Њ—Б—Б–Є–є—Б–Ї–Њ–≥–Њ —Д–Њ–љ–і–∞ —Д—Г–љ–і–∞–Љ–µ–љ—В–∞–ї—М–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є (18-04-01252).
–Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П —Б–љ–∞ –Љ–Њ—А—Б–Ї–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –њ–Њ–і–і–µ—А–ґ–∞–љ—Л¬†–≥—А–∞–љ—В–∞–Љ–Є –Э–∞—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–≥–Њ –љ–∞—Г—З–љ–Њ–≥–Њ —Д–Њ–љ–і–∞ –°–®–Р, –Э–∞—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–≥–Њ –Є–љ—Б—В–Є—В—Г—В–∞ –Ј–і–Њ—А–Њ–≤—М—П –°–®–Р, –†–Њ—Б—Б–Є–є—Б–Ї–Њ–≥–Њ —Д–Њ–љ–і–∞ —Д—Г–љ–і–∞–Љ–µ–љ—В–∞–ї—М–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –Є¬†–Ю–Ю–Ю¬†¬Ђ–£—В—А–Є—И—Б–Ї–Є–є –і–µ–ї—М—Д–Є–љ–∞—А–Є–є¬ї (–†–Њ—Б—Б–Є—П).
O.I. Lyamin
A.N. Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences, Moscow
University of California, Los Angeles
Contact person: Oleg Irikovich Lyamin, oilyamin@yahoo.com
Sleep and wakefulness are two basic states characteristic of all animals from simple organisms with decentralized nervous systems to placental mammals with very complex brains. In all studied animals, sleep features similar behavioral, neuronal, molecular and physiological parameters. In low vertebrates sleep has many characteristics of the mammalian slow wave sleep (SWS). At the same time, two differentiated states вАУ SWS and paradoxical (rapid eye movement of fast) sleep are present only in birds and mammals. In birds and вАШprimitiveвАЩ mammals (monotreems and marsupials) sleep is less differentiated than in placental mammals. In marine mammals (cetaceans and eared seals belonging to the group of pinnipeds) SWS may occur in one cortical hemisphere at a time (called unihemispheric sleep). Episodes of interhemispheric electroencephalogram asymmetry during SWS are also recorded in some birds. Those episodes resemble unihemispheric sleep. It is believed that unihemispheric sleep facilitates movement and vigilance in animals while they are asleep. The most recent studies have convincingly demonstrated the key role of ecological and evolutional factors in shaping the features and duration of sleep in animals.
–£–≤–∞–ґ–∞–µ–Љ—Л–є –њ–Њ—Б–µ—В–Є—В–µ–ї—М uMEDp!
–£–≤–µ–і–Њ–Љ–ї—П–µ–Љ –Т–∞—Б –Њ —В–Њ–Љ, —З—В–Њ –Ј–і–µ—Б—М —Б–Њ–і–µ—А–ґ–Є—В—Б—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П, –њ—А–µ–і–љ–∞–Ј–љ–∞—З–µ–љ–љ–∞—П –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –і–ї—П —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П.
–Х—Б–ї–Є –Т—Л –љ–µ —П–≤–ї—П–µ—В–µ—Б—М —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–Љ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П, –∞–і–Љ–Є–љ–Є—Б—В—А–∞—Ж–Є—П –љ–µ –љ–µ—Б–µ—В –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ—Б—В–Є –Ј–∞ –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ –њ–Њ—Б–ї–µ–і—Б—В–≤–Є—П, –≤–Њ–Ј–љ–Є–Ї—И–Є–µ –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–∞–Љ–Њ—Б—В–Њ—П—В–µ–ї—М–љ–Њ–≥–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Т–∞–Љ–Є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є —Б –њ–Њ—А—В–∞–ї–∞ –±–µ–Ј –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є –Ї–Њ–љ—Б—Г–ї—М—В–∞—Ж–Є–Є —Б –≤—А–∞—З–Њ–Љ.
–Э–∞–ґ–Є–Љ–∞—П –љ–∞ –Ї–љ–Њ–њ–Ї—Г ¬Ђ–Т–Њ–є—В–Є¬ї, –Т—Л –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В–µ, —З—В–Њ —П–≤–ї—П–µ—В–µ—Б—М –≤—А–∞—З–Њ–Љ –Є–ї–Є —Б—В—Г–і–µ–љ—В–Њ–Љ –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–≥–Њ –≤—Г–Ј–∞.