ааЗаМаЕаНаЕаНаИб б аАбаАаКбаЕбаИббаИаК аДаЕаЛббаА-аКаОаЛаЕаБаАаНаИаЙ аИ аБаЕбаА-баИбаМаА аНаА баЛаЕаКббаОбаНбаЕбаАаЛаОаГбаАаМаМаЕ аПбаИТ баАаМаОаПбаОаИаЗаВаОаЛбаНаОаМ аПбаОаБбаЖаДаЕаНаИаИ аИаЗТ аВбаОбаОаЙ ббаАаДаИаИ аДаНаЕаВаНаОаГаО баНаА бТ аПаОбаЛаЕаДбббаИаМ аВбаПаОаЛаНаЕаНаИаЕаМ аПбаИб аОаМаОбаОбаНаОаЙ аДаЕббаЕаЛбаНаОббаИ баАаЗаНаОаЙ аДаЛаИбаЕаЛбаНаОббаИ
- ааНаНаОбаАбаИб
- аЁбаАббб
- аЁббаЛаКаИ
- English


ааВаЕаДаЕаНаИаЕ
ааКбаИаВаАбаИб аМаОаЗаГаА, аКаОбаОбаАб ббаАббаВбаЕб аВТ баЕаГбаЛббаИаИ баНаА аВТ баОаОбаВаЕбббаВаИаИ бТ аВаНбббаЕаНаНаИаМаИ аПаОббаЕаБаНаОбббаМаИ аИТ аВаНаЕбаНаИаМ аВаОаЗаДаЕаЙббаВаИаЕаМ, баВаЛбаЕббб аВаАаЖаНбаМ аОаБбаЕаКбаОаМ аИббаЛаЕаДаОаВаАаНаИаЙ аВТ ббаЛаОаВаИбб ббаИаЛаИаВаАббаИб бб баОбаИаАаЛбаНбб ббаЕаБаОаВаАаНаИаЙ, аНаАбббаАббаИб аЕббаЕббаВаЕаНаНаОаЕ баЕбаЕаНаИаЕ баИаКаЛаА ТЋбаОаНТ т аБаОаДбббаВаОаВаАаНаИаЕТЛ. ааНаА аОаБаЕбаПаЕбаИаВаАаЕб аОаБбаАбаИаМаОббб баНаА аИТ аОаПбаЕаДаЕаЛаЕаНаНбаЙ ббаОаВаЕаНб аБаДаИбаЕаЛбаНаОббаИ, ббаО аПаОаЗаВаОаЛбаЕб баПббаЕаМб аАаДаАаПбаИбаОаВаАбббб аКТ аВаНаЕбаНаЕаЙ ббаЕаДаЕ [1]. аТ баО аЖаЕ аВбаЕаМб аАаКбаИаВаАбаИаОаНаНбаЕ аПбаОбаЕббб аВаОаВаЛаЕбаЕаНб аВТ аПаАбаОбаИаЗаИаОаЛаОаГаИб аНаАбббаЕаНаИаЙ баНаА [2]. аб Т аВаОаЗаНаИаКаНаОаВаЕаНаИаЕ аИТ ббаАбаЕаНаИаЕ аМаОаГбб баВаИаДаЕбаЕаЛбббаВаОаВаАбб аОТ ббаАаГаМаЕаНбаАбаИаИ аИТ аНаЕббаАаБаИаЛбаНаОббаИ баНаА, баАббаМаАббаИаВаАбббб аКаАаК аМаАбаКаЕб аКаАбаЕббаВаА баНаА аИТ аИаНбаОаМаНаИаИТ [3]. ааАТ баЛаЕаКббаОбаНбаЕбаАаЛаОаГбаАаМаМаЕ (ааа) аОаПбаЕаДаЕаЛббббб аБбббббаЕ баДаВаИаГаИ баАббаОбаНаОаГаО баОббаАаВаА ааа аВТ ббаОбаОаНб баЕбаА-, аАаЛббаА- аИаЛаИ аБаЕбаА-баИбаМаОаВ, аАТ баАаКаЖаЕ аВббаОаКаОаАаМаПаЛаИббаДаНбаЕ аКаОаЛаЕаБаАаНаИб (аДаЕаЛббаА-аПаОаДаОаБаНбаЕ аВаОаЛаНб аИаЛаИ а-аКаОаМаПаЛаЕаКбб), аКаОбаОббаЕ аОббаАаЖаАбб аОбаОаБбб аАаКбаИаВаАбаИб, баОаПбаОаВаОаЖаДаАбббббб баОбаМаОаЗаНбаМаИ аПбаОбаЕббаАаМаИ аВТ аМаОаЗаГаЕ [1, 2, 4].
аТ ббаДаЕ баАаБаОб аПаОаКаАаЗаАаНаА аАббаОбаИаАбаИб аБаЕбаА-баИбаМаА аВТ аМаОбаОбаНаОаЙ аКаОбаЕ бТ аПаОбаВаЛаЕаНаИаЕаМ а-аКаОаМаПаЛаЕаКбаА, аЗаА аКаОбаОббаМ баЛаЕаДбаЕб баПаИаЗаОаД аАаКбаИаВаАбаИаИ [4, 5]. аТ аИббаЛаЕаДаОаВаАаНаИбб бТ аОаДаНаОаВбаЕаМаЕаНаНаОаЙ баЕаГаИбббаАбаИаЕаЙ аКаОбаКаОаВаОаЙ аИТ аВаНбббаИаКаОбаКаОаВаОаЙ ааа бТ аПаАбаИаЕаНбаОаВ бТ баПаИаЛаЕаПбаИаЕаЙ аВаНаЕ аПаЕбаИаОаДаА аОаБаОбббаЕаНаИб аВббаВаЛаЕаНаА баВбаЗб баЕаАаКбаИаЙ аАаКбаИаВаАбаИаИ баОТ ббаАаДаИбаМаИ баНаА, аОаБаЛаАбббаМаИ аОбаВаЕаДаЕаНаИб аБаИаОбаЛаЕаКббаИбаЕбаКаОаЙ аАаКбаИаВаНаОббаИ, аАТ баАаКаЖаЕ баИаПаОаМ аПбаОаБбаЖаДаЕаНаИбТ т баАаМаОаПбаОаИаЗаВаОаЛбаНбаМ аИаЛаИ аВбаЗаВаАаНаНбаМ аВаНаЕбаНаИаМ аВаОаЗаДаЕаЙббаВаИаЕаМ [6, 7]. аТ баО аЖаЕ аВбаЕаМб баАббаОбаНбаЕ б аАбаАаКбаЕбаИббаИаКаИ ааа аПбаИ баПаОаНбаАаНаНаОаЙ аАаКбаИаВаАбаИаИ аДаОббаАбаОбаНаО аИаНбаОбаМаАбаИаВаНб, ббаОаБб аНаАТ аОбаНаОаВаАаНаИаИ аБаОаЛббаИб аДаАаНаНбб аИТ аИбаКббббаВаЕаНаНаОаГаО аИаНбаЕаЛаЛаЕаКбаА баАаЗаДаЕаЛаИбб аЗаАаПаИбаИ аЗаДаОбаОаВбб ббаБбаЕаКбаОаВ аИТ аПаАбаИаЕаНбаОаВ бТ б баОаНаИбаЕбаКаОаЙ аБаЕббаОаНаНаИбаЕаЙ аИТ аНаАбббаЕаНаИаЕаМ аВаОбаПбаИббаИб баОббаОбаНаИб баНаА. аЃТ аПаОбаЛаЕаДаНаИб аПаОТ ббаАаВаНаЕаНаИб баОТ аЗаДаОбаОаВбаМаИ аЛаИбаАаМаИ аМаЕаНаЕаЕ аВббаАаЖаЕаНб аИаЗаМаЕаНаЕаНаИб аВТ аДаЕаЛббаА- аИТ аБаОаЛббаЕ аВТ баЕбаА-, аАаЛббаА- аИТ аБаЕбаА-аДаИаАаПаАаЗаОаНаАб [8].
аббаЕаКбаИаВаНаОббб аВаОбббаАаНаОаВаЛаЕаНаИб аКаОаГаНаИбаИаВаНбб ббаНаКбаИаЙ аПбаИ аАаКбаИаВаАбаИаИ аИаЗббаАаЛаАбб аВТ аИббаЛаЕаДаОаВаАаНаИбб баНаА аВТ аЗаНаАбаИбаЕаЛбаНаО аМаЕаНббаЕаЙ ббаЕаПаЕаНаИ, баЕаМ аИб аКаЛаИаНаИбаЕбаКаИаЙ аАбаПаЕаКб. ааОаКаАаЗаАаНаА аКаЛббаЕаВаАб баОаЛб баЕаАаКбаИаВаНаОббаИ аМаОаЗаГаА аИТ аПбаОаДаОаЛаЖаИбаЕаЛбаНаОббаИ аПбаОаБбаЖаДаЕаНаИб аПаОбаЛаЕ аНаАбаАаЛаА аПаАббаЕбаНаА аАаКбаИаВаАбаИаИ аВТ баОбаМаИбаОаВаАаНаИаИ аПаАаМббаИ аОТ баОаДаЕбаЖаАаНаИаИ баНаОаВаИаДаЕаНаИаЙ [9]. абаИ баЕаЗбаЛббаАбб баОаГаЛаАбббббб бТ аДаАаНаНбаМаИ аИббаЛаЕаДаОаВаАаНаИаЙ аАаКбаИаВаНаОббаИТ аГаИаПаПаОаКаАаМаПаА аИТ аОбаБаИбаОббаОаНбаАаЛбаНаОаЙ аКаОбб, аВбаЗаВаАаНаНаОаЙ баПаОаНбаАаНаНаОаЙ аАаКбаИаВаАбаИаЕаЙ бТ аПаОбаЛаЕаДбббаИаМ аПбаОаБбаЖаДаЕаНаИаЕаМ. аТ аИббаЛаЕаДаОаВаАаНаИбб аОбаМаЕбаАаЛбб аПбаОаГбаЕббаИббббаИаЙ аПбаОбаЕбб, аМаОаДбаЛаИббаЕаМбаЙ ббаАаДаИаЕаЙ баНаА, аАТ аВ аЛаОаБаНаОаЙ аКаОбаЕТ т аПаЕбаВаОаНаАбаАаЛбаНбаМ ббаОаВаНаЕаМ аАаКбаИаВаАбаИаИ [7]. абаИ баАаКбб аМаОаГбб аОаБбббаНаИбб аБаОаЛаЕаЕ ббаПаЕбаНбаЕ аВаОбаПаОаМаИаНаАаНаИб аОТ баНаОаВаИаДаЕаНаИбб аПаОбаЛаЕ аДаЛаИбаЕаЛбаНаОаГаО аПбаОаБбаЖаДаЕаНаИб аИаЗТ REM-баНаА, аПаОбаКаОаЛбаКб аАаКбаИаВаНаОбббТ аГаИаПаПаОаКаАаМаПаА аПбаИаБаЛаИаЖаАаЕббб аКТ ббаОаВаНб аПбаИ аБаОаДбббаВаОаВаАаНаИаИ аИТ аПаОаЗаВаОаЛбаЕб аВаОбббаАаНаОаВаИбб ббаНаКбаИаИ аКаОаДаИбаОаВаАаНаИб аИТ аКаОаНбаОаЛаИаДаАбаИаИ аПаАаМббаИТ [7].
ааАбаИ аИббаЛаЕаДаОаВаАаНаИб баАаМаОаПбаОаИаЗаВаОаЛбаНаОаЙ аАаКбаИаВаАбаИаИ аВаО аВбаЕаМб баПаИаЗаОаДаА аДаНаЕаВаНаОаГаО баНаА, аЕаГаО аВбаОбаОаЙ ббаАаДаИаИ (N2), аИТ баЛаЕаДбббаЕаГаО аЗаА аНаИаМ аПаЕбаИаОаДаА аПбаОаБбаЖаДаЕаНаИб, аВаКаЛббаАаВбаЕаГаО аПбаИаПаОаМаИаНаАаНаИаЕ аИаНббббаКбаИаИ аИТ аПаОаДаГаОбаОаВаКб аКТ аПбаИб аОаМаОбаОбаНаОаЙ аДаЕббаЕаЛбаНаОббаИ, аПаОаКаАаЗаАаЛаИ баЛаЕаДбббаЕаЕ. ааЛббаА-баИбаМб аПбаИ аПбаОаБбаЖаДаЕаНаИаИ баАббаО аПбаЕаДбаЕббаВбаЕб аПаОбаВаЛаЕаНаИаЕ а-аКаОаМаПаЛаЕаКбаОаВ баАаЗаНаОаЙ ббаЕаПаЕаНаИ аВббаАаЖаЕаНаНаОббаИ, аНаАТ аКаОбаОббаЕ аНаАаКаЛаАаДбаВаАбббб аАаЛббаА-аКаОаЛаЕаБаАаНаИб. ааОаДаОаБаНбаЙ аПаАббаЕбаН, аВаЕбаОббаНаО, аОббаАаЖаАаЕб аДаЕаЙббаВаИаЕ баНаИаВаЕббаАаЛбаНаОаГаО баАаЛаАаМаО-аКаОббаИаКаАаЛбаНаОаГаО аАаКбаИаВаИббббаЕаГаО аМаЕб аАаНаИаЗаМаА. ааГаО аПаОбаВаЛаЕаНаИаЕ аНаЕТ аЗаАаВаИбаИб аОбТ баОаГаО, аЗаАаВаЕббаИббб аЛаИ аКаОбаОбаКаИаЙ аПаЕбаИаОаД аАаКбаИаВаАбаИаИ аПаОаГббаЖаЕаНаИаЕаМ аВТ баОаН, аИаЛаИ аПаОбаЛаЕаДбаЕб аПбаОаБбаЖаДаЕаНаИаЕ [10].
аІаЕаЛб аДаАаНаНаОаЙ баАаБаОббТ т аИаЗббаИбб б аАбаАаКбаЕбаИббаИаКаИ ааа аПаОбаЛаЕ баАаМаОаПбаОаИаЗаВаОаЛбаНаОаЙ аАаКбаИаВаАбаИаИ аМаОаЗаГаА аВаО аВбаЕаМб N2 аДаНаЕаВаНаОаГаО баНаА, аКаОбаОбаАб аЗаАаВаЕббаАаЕббб баПаИаЗаОаДаАаМаИ аПбаОаБбаЖаДаЕаНаИб баАаЗаНаОаЙ аДаЛаИбаЕаЛбаНаОббаИ аИТ баОаПбаОаВаОаЖаДаАаЕббб аВбаПаОаЛаНаЕаНаИаЕаМ аПбаИб аОаМаОбаОбаНаОаГаО баЕббаА (аааЂ) [11]. аТ аЗаАаДаАбаИ аИббаЛаЕаДаОаВаАаНаИб аВб аОаДаИаЛаА аОбаЕаНаКаА:
- баОаОбаНаОбаЕаНаИб аНаИаЗаКаИб (аДаЕаЛббаА) аИТ аВббаОаКаИб (аБаЕбаА) аДаИаАаПаАаЗаОаНаОаВ баПаЕаКббаА ааа аВаО аВбаЕаМб баПаИаЗаОаДаА аАаКбаИаВаАбаИаИ бТ аПаОбаЛаЕаДбббаИаМ аПбаОаБбаЖаДаЕаНаИаЕаМ баАаЗаНаОаЙ аДаЛаИбаЕаЛбаНаОббаИ, аКаОбаОбаОаЕ баОаПбаОаВаОаЖаДаАаЕббб аааЂ,Т т аДаЛб ббаМаМаАбаНаОаЙ ааа аИТ аОбаДаЕаЛбаНаО аПаОТ аОаБаЛаАбббаМ аОбаВаЕаДаЕаНаИб;
- баОаОбаНаОбаЕаНаИб ббаИб баАббаОб аВТ аНаАбаАаЛаЕ аВбаПаОаЛаНаЕаНаИб аааЂ аВаО аВбаЕаМб аПбаОаБбаЖаДаЕаНаИб баАаЗаНаОаЙ аДаЛаИбаЕаЛбаНаОббаИ аВТ баАаЗаНбаЕ баАаЗб аВбаПаОаЛаНаЕаНаИб аааЂ: ббаЕб аВТ баМаЕ бТ баИаНб баОаНаНбаМаИ аНаАаЖаАбаИбаМаИ (аПаЕбаВаАб баАаЗаА) аИТ баОаЛбаКаО ббаЕб аВТ баМаЕ (аВбаОбаАб баАаЗаА).
ааАбаЕбаИаАаЛ аИТ аМаЕбаОаДб
аТ аИббаЛаЕаДаОаВаАаНаИаИ ббаАббаВаОаВаАаЛаИ 34 аПбаАаКбаИбаЕбаКаИ аЗаДаОбаОаВбб бббаДаЕаНбаА (26 аЖаЕаНбаИаН аИТ 8 аМбаЖбаИаН, аВбаЕ аПбаАаВбаИ) аВТ аВаОаЗбаАббаЕ аОбТ 18 аДаО 22 аЛаЕб. аббаЛаЕаДаОаВаАаНаИаЕ аБбаЛаО аОаДаОаБбаЕаНаО ббаИбаЕбаКаОаЙ аКаОаМаИббаИаЕаЙ ааНббаИбббаА аВбббаЕаЙ аНаЕбаВаНаОаЙ аДаЕббаЕаЛбаНаОббаИ аИТ аНаЕаЙбаОбаИаЗаИаОаЛаОаГаИаИ а аОббаИаЙбаКаОаЙ аАаКаАаДаЕаМаИаИ аНаАбаК (аПбаОбаОаКаОаЛ тТ 2 аОбТ 3 аИбаНб 2019Т аГ.) аИТ баОаОбаВаЕбббаВаОаВаАаЛаО ббаИбаЕбаКаИаМ аНаОбаМаАаМ аЅаЕаЛббаИаНаКбаКаОаЙ аДаЕаКаЛаАбаАбаИаИ абаЕаМаИбаНаОаЙ аМаЕаДаИбаИаНбаКаОаЙ аАббаОбаИаАбаИаИ ТЋабаИбаЕбаКаИаЕ аПбаИаНбаИаПб аПбаОаВаЕаДаЕаНаИб аНаАббаНбб аМаЕаДаИбаИаНбаКаИб аИббаЛаЕаДаОаВаАаНаИаЙ бТ ббаАббаИаЕаМ баЕаЛаОаВаЕаКаАТЛ бТ аПаОаПбаАаВаКаАаМаИ аОбТ 2000Т аГ.
ааКбаПаЕбаИаМаЕаНб аНаАбаИаНаАаЛаИ аВТ 13.00. ааЛб аОбаЕаНаКаИ аПаАбаАаМаЕббаОаВ аНаОбаНаОаГаО баНаА, аПбаЕаДбаЕббаВаОаВаАаВбаЕаГаО аИббаЛаЕаДаОаВаАаНаИб, аИбаПбббаЕаМбаЕ аЗаАаПаОаЛаНбаЛаИ аДаНаЕаВаНаИаК баНаА [12]. ааЛб аОбаЕаНаКаИ аДаНаЕаВаНаОаЙ баОаНаЛаИаВаОббаИ аИбаПаОаЛбаЗаОаВаАаЛаИ ааАбаОаЛаИаНбаКбб баКаАаЛб баОаНаЛаИаВаОббаИ (KSS). ааО аВбаЕаМб аОаПббаА (аОаКаОаЛаО баАбаА) аИбаПбббаЕаМбаЙ аЛаЕаЖаАаЛ аНаАТ аКббаЕбаКаЕ бТ аЗаАаКббббаМаИТ аГаЛаАаЗаАаМаИ аВТ аЗаАбаЕаМаНаЕаНаНаОаМ аЗаВбаКаОаИаЗаОаЛаИббббаЕаМ аПаОаМаЕбаЕаНаИаИ. аЁТ аПаОаМаОббб аааЂ аПаОаЛббаАаЛаИ ббаД аПаОбаЛаЕаДаОаВаАбаЕаЛбаНбб аПаЕбаИаОаДаОаВ аЗаАббаПаАаНаИб аИТ аПбаОаБбаЖаДаЕаНаИб. аЂаЕбб баОббаОбаЛ аИаЗТ аДаВбб баЕбаЕаДбббаИб бб баАаЗ. абаПбббаЕаМбаЕ ббаИбаАаЛаИ ТЋаПбаО баЕаБбТЛ аОбТ аОаДаНаОаГаО аДаО аДаЕбббаИ, аОаДаНаОаВбаЕаМаЕаНаНаО аПбаИ аКаАаЖаДаОаМ аОбббаЕбаЕ аНаАаЖаИаМаАаЛаИ аНаАТ аКаНаОаПаКб, аЗаАбаИаКбаИбаОаВаАаНаНбб аНаАТ баКаАаЗаАбаЕаЛбаНаОаМ аПаАаЛббаЕ аПбаАаВаОаЙ ббаКаИ, аБаОаЛббаИаМ аПаАаЛббаЕаМ баОаЙ аЖаЕ ббаКаИ (аПаЕбаВаАб баАаЗаА). ааАаЛаЕаЕ аОаНаИ аПбаОаДаОаЛаЖаАаЛаИ ббаИбаАбб ТЋаПбаО баЕаБбТЛ аОбТ аОаДаНаОаГаО аДаО аДаЕбббаИ, аНаОТ баЖаЕ аБаЕаЗ аНаАаЖаАбаИаЙ (аВбаОбаАб баАаЗаА). аЄаАаЗб баЕбаЕаДаОаВаАаЛаИбб аДаО баЕб аПаОб, аПаОаКаА ббаАббаНаИаКаИ аИббаЛаЕаДаОаВаАаНаИб аНаЕТ аЗаАббаПаАаЛаИ. абаИ баПаОаНбаАаНаНбб аПбаОаБбаЖаДаЕаНаИбб аОаНаИ аВаОаЗаОаБаНаОаВаЛбаЛаИ аВбаПаОаЛаНаЕаНаИаЕ баЕббаА бТ аПаЕбаВаОаЙ баАаЗб. а аЕаГаИбббаИбаОаВаАаЛаИ ааа аОбТ 17 аОбаВаЕаДаЕаНаИаЙ аВТ баОаОбаВаЕбббаВаИаИ баОТ бб аЕаМаОаЙ 10т20, баЛаЕаКббаОаОаКбаЛаОаГбаАаМаМб, баЛаЕаКббаОаМаИаОаГбаАаМаМб аКаОбаОбаКаОаЙ аМбббб, аОбаВаОаДббаЕаЙ аБаОаЛббаОаЙ аПаАаЛаЕб аКаИббаИ аПбаАаВаОаЙ ббаКаИ, аИТ аМаЕб аАаНаОаГбаАаМаМб аНаАаЖаАбаИаЙ аНаАТ аКаНаОаПаКб. а аЕаГаИбббаАбаИб аОбббаЕббаВаЛбаЛаИ бТ аПаОаМаОббб баИббаЕаМб Neocortex-Pro (Neurobotics). аЇаАббаОбаА аДаИбаКбаЕбаИаЗаАбаИаИТ т 250 аб, аПаОаЛаОбаА аПбаОаПббаКаАаНаИб баАббаОбТ т 0,5т70 аб. абаПаОаЛбаЗаОаВаАаЛаИ баЛаЕаМ аИТ б аЛаОббаЕбаЕаБббаНбаЕ баЛаЕаКббаОаДб, баОаПбаОбаИаВаЛаЕаНаИаЕТ т аМаЕаНаЕаЕ 5 аКааМ. ааЛб аАаНаАаЛаИаЗаА аВбаБаИбаАаЛаИ аОббаЕаЗаКаИ аЗаАаПаИбаИ ааа, баОаОбаВаЕбббаВаОаВаАаВбаИаЕ баПаИаЗаОаДаАаМ аПаОаЛаНаОаГаО аИТ баАббаИбаНаОаГаО аВбаПаОаЛаНаЕаНаИб аааЂ аПбаИ баАаМаОаПбаОаИаЗаВаОаЛбаНаОаМ аПбаОаБбаЖаДаЕаНаИаИ аИаЗТ N2, аКаОбаОббаЙ аНаАаБаЛбаДаАаЛбб аНаЕТ аМаЕаНаЕаЕ 1,5Т аМаИаНббб. абаИаНаАаДаЛаЕаЖаНаОббб ааа аКТ N2 аОбаЕаНаИаВаАаЛаИ аВаИаЗбаАаЛбаНаО аПаОТ аКбаИбаЕбаИбаМ AASM [13]. ааОаЛаНбаМ ббаИбаАаЛаИ аВбаПаОаЛаНаЕаНаИаЕ баЕббаА, аЕбаЛаИ аИбаПбббаЕаМбаЙ аПаОбаЛаЕ аПбаОаБбаЖаДаЕаНаИб аПбаАаВаИаЛбаНаО аВбаПаОаЛаНбаЛ аЗаАаДаАаНаИаЕ аПаЕбаВаОаЙ баАаЗб баЕббаА, баО аЕббб аНаАаЖаИаМаАаЛ аДаЕбббб баАаЗ аНаАТ аКаНаОаПаКб, аИТ аЗаАбаЕаМ баЕбаЕаЗ аПбаОаМаЕаЖббаОаК, баОаИаЗаМаЕбаИаМбаЙ бТ аВбаЕаМаЕаНаЕаМ ббаЕбаА ТЋаПбаО баЕаБбТЛ аОбТ аОаДаНаОаГаО аДаО аДаЕбббаИ (аОаКаОаЛаО аДаЕбббаИ баЕаКбаНаД, аВбаОбаАб баАаЗаА баЕббаА), аПаОаЛаНаОбббб аВаОбаПбаОаИаЗаВаОаДаИаЛ аПаЕбаВбб баАаЗб. аТ баАббаИбаНаОаМ аВбаПаОаЛаНаЕаНаИаИТ аГаОаВаОбаИаЛаИ, аКаОаГаДаА аПаОбаЛаЕ аПаЕбаВаОаЙ баАаЗб, аОбббаЕббаВаЛаЕаНаНаОаЙ аНаЕТ аПаОаЛаНаОбббб (6т9 аНаАаЖаАбаИаЙ), аЖаДаАбб баЛаЕаДбббаЕаГаО аНаАаЖаАбаИб аПбаИб аОаДаИаЛаОбб аНаЕТ аМаЕаНаЕаЕ аМаИаНббб.
абаБаИбаАаЛаИ аИбаПбббаЕаМбб , бТ аКаОбаОббб б аОбб аБб аПаОТ аОаДаНаОаМб баАаЗб баЕаГаИбббаИбаОаВаАаЛаОбб аПаОаЛаНаОаЕ аИТ баАббаИбаНаОаЕ аВбаПаОаЛаНаЕаНаИаЕ баЕббаА. аЂаАаКаИб аОаКаАаЗаАаЛаОбб 17 (13 аЖаЕаНбаИаН аИТ баЕбаВаЕбаО аМбаЖбаИаН). аЁбаЕаДаНбб аПбаОаДаОаЛаЖаИбаЕаЛбаНаОббб баНаА аВТ аНаОбб аПаЕбаЕаД аОаПббаОаМ аВТ аДаАаНаНаОаЙТ аГббаПаПаЕ баОббаАаВаИаЛаАТ 5,9 ТБ 2,7 баАбаА, баАаМаОббаВббаВаИаЕ аПбаИ аПбаОаБбаЖаДаЕаНаИаИ аИТ аКаАбаЕббаВаО баНаАТ т 3,2 ТБ 1,2 аИТ 3,8 ТБ 1,3 аБаАаЛаЛаА баОаОбаВаЕбббаВаЕаНаНаО (аПаОТ аПббаИаБаАаЛаЛбаНаОаЙ баКаАаЛаЕ,Т аГаДаЕ 5Т т аОбаЛаИбаНаО), ббаЕаДаНаИаЙ ббаОаВаЕаНб баОаНаЛаИаВаОббаИ аПаЕбаЕаД аОаПббаОаМТ т 5,1 ТБ 1,7 аБаАаЛаЛаА (аПаОТ баЕаМаИаБаАаЛаЛбаНаОаЙ баКаАаЛаЕ,Т аГаДаЕ 1Т т аОбаЛаИбаНаО). абаЛаИ аПаОаЛаНбб аИТ баАббаИбаНбб баПаИаЗаОаДаОаВ аВбаПаОаЛаНаЕаНаИб аааЂ аБбаЛаО аНаЕбаКаОаЛбаКаО, баО аВбаБаИбаАаЛаИ аПаОТ аОаДаНаОаМб бТ аМаАаКбаИаМаАаЛбаНаОаЙ аПбаОаДаОаЛаЖаИбаЕаЛбаНаОбббб. аТ аОаПбаЕаДаЕаЛаЕаНаНаОаЙ ббаЕаПаЕаНаИ ббаО аПаОаЗаВаОаЛаИаЛаО баБаЛаИаЗаИбб ббаОаВаНаИ аАаКбаИаВаАбаИаИ ббаБбаЕаКбаА аВТ баПаИаЗаОаДаАб аВбаПаОаЛаНаЕаНаИб аДаЕббаЕаЛбаНаОббаИ бТ баАаЗаНаОаЙ бббаЕаКбаИаВаНаОбббб аПаОбаЛаЕ аПбаОаБбаЖаДаЕаНаИб.
ааНаАаЛаИаЗаИбаОаВаАаЛаИ 40-баЕаКбаНаДаНбаЕ аОббаЕаЗаКаИ аЗаАаПаИбаИ ааа, аНаАТ баЕбаЕаДаИаНб аКаОбаОббб аПбаИб аОаДаИаЛаОбб аВаОаЗаОаБаНаОаВаЛаЕаНаИаЕ аНаАаЖаАбаИаЙ аНаАТ аКаНаОаПаКб аПаОбаЛаЕ N2: аПаОТ 20 баЕаКбаНаД аДаО аИТ аПаОбаЛаЕ аВаОаЗаОаБаНаОаВаЛаЕаНаИб аНаАаЖаАбаИаЙ. ааЛб аОбаЕаНаКаИ аМаОбаНаОббаНбб б аАбаАаКбаЕбаИббаИаК аБаИаОбаЛаЕаКббаИбаЕбаКаОаЙ аАаКбаИаВаНаОббаИ аКаОбб аПбаОаВаОаДаИаЛаИ аНаЕаПбаЕббаВаНаОаЕ аВаЕаЙаВаЛаЕб-аПбаЕаОаБбаАаЗаОаВаАаНаИаЕ аНаАТ аОбаНаОаВаЕ аМаАбаЕбаИаНбаКаОаГаО аКаОаМаПаЛаЕаКбаНаОаГаО morlet-аВаЕаЙаВаЛаЕбаА (Matlab 78.01). ааАббб баАбаПбаЕаДаЕаЛаЕаНаИб аЗаНаАбаЕаНаИаЙ аМаОаДбаЛб аКаОбббаИбаИаЕаНбаА аВаЕаЙаВаЛаЕб-аПбаЕаОаБбаАаЗаОаВаАаНаИб (ааа) бббаОаИаЛаИ аВТ аПаОаЛаОбаЕ 0,5т40 аб бТ баАаГаОаМ 0,5 аб аИТ баАаЗбаЕбаЕаНаИаЕаМ аПаОТ аВбаЕаМаЕаНаИ 0,01 б. аТ аДаЕаЛббаА- (1т3,5 аб) аИТ аБаЕбаА-аДаИаАаПаАаЗаОаНаАб (14т21 аб) бббаЕаДаНбаЛаИ ааа аПаОТ баАббаОбаЕ; аДаАаЛаЕаЕ аПаОТ аВбаЕаМаЕаНаИ, аВТ аДаВбб баЕаКбаНаДаНбб аИаНбаЕбаВаАаЛаАб (аПаОТ аДаЕбббб аДаО аИТ аПаОбаЛаЕ аНаАбаАаЛаА аНаАаЖаАбаИаЙ). ааОаЛббаЕаНаНбаЕ аАаМаПаЛаИббаДаНаО-аМаОбаНаОббаНбаЕ б аАбаАаКбаЕбаИббаИаКаИ ааа аОбаЕаНаИаВаАаЛаИ бТ аПаОаМаОббб аДаИбаПаЕббаИаОаНаНаОаГаО аАаНаАаЛаИаЗаА (ANOVA RM). а аАббаМаАббаИаВаАаЛаИ аВаЛаИбаНаИаЕ аНаАТ аНаИб баАаКбаОбаОаВ, баАаКаИб аКаАаК бббаЕаКбаИаВаНаОббб аДаЕббаЕаЛбаНаОббаИТ т аДаВаА ббаОаВаНб (аПаОаЛаНаОаЕ аИТ баАббаИбаНаОаЕ аВбаПаОаЛаНаЕаНаИаЕ аааЂ) аИТ аПбаИб аОаМаОбаОбаНаАб аАаКбаИаВаНаОбббТ т аДаВаА ббаОаВаНб (аПаЕбаИаОаДб аДаО аНаАбаАаЛаА аНаАаЖаАбаИаЙ аНаАТ аКаНаОаПаКб аИТ аПаОбаЛаЕ). ааЛб аПаОаВбаОбаНбб аИаЗаМаЕбаЕаНаИаЙ аИбаПаОаЛбаЗаОаВаАаЛаИ баАаКбаОб аВбаЕаМаЕаНаИ (аДаЕбббб ббаОаВаНаЕаЙ, аДаЕбббб аДаВбб баЕаКбаНаДаНбб аИаНбаЕбаВаАаЛаОаВ). ANOVA RM аПбаОаВаОаДаИаЛаИ аКаАаК ббаМаМаАбаНаО (аПаОТ бббаЕаДаНаЕаНаНбаМ аМаОбаНаОббаНбаМ б аАбаАаКбаЕбаИббаИаКаАаМ аВбаЕб аОбаВаЕаДаЕаНаИаЙ ааа аДаЛб аКаАаЖаДаОаГаО аОбббаЕбаА), баАаК аИТ аПаО аОбаДаЕаЛбаНаОббаИ аДаЛб аКаАаЖаДаОаГаО аОбаВаЕаДаЕаНаИб ааа. аЁбаАбаИббаИбаЕбаКбб аОаБбаАаБаОбаКб аОбббаЕббаВаЛбаЛаИ бТ ббаЕбаОаМ аПаОаПбаАаВаКаИ абаИаНб аАбаЗаАТ т ааЕббаЕбаА.
ааОаЛббаЕаНаНбаЕ аВТ баЕаЗбаЛббаАбаЕ бббаЕаДаНаЕаНаИаЙ аАаМаПаЛаИббаДаНаО-аМаОбаНаОббаНбаЕ б аАбаАаКбаЕбаИббаИаКаИ ааа бТ аПаОаМаОббб аПаАбаНаОаГаО аКбаИбаЕбаИб аЃаИаЛаКаОаКбаОаНаА ббаАаВаНаИаВаАаЛаИ аПбаИ аПаОаЛаНаОаМ аИТ баАббаИбаНаОаМ аВбаПаОаЛаНаЕаНаИаИ аааЂТ т аПаОТ аКаАаЖаДаОаМб аВбаДаЕаЛаЕаНаНаОаМб аДаВбб баЕаКбаНаДаНаОаМб аИаНбаЕбаВаАаЛб аИТ аВ аКаАаЖаДаОаМ аИаЗТ баПаЕаКббаАаЛбаНбб аДаИаАаПаАаЗаОаНаОаВ.
абаОаМаЕ баОаГаО, ббаАаВаНаИаВаАаЛаИ аИаЗаМаЕаНаЕаНаИб баПаЕаКббаАаЛбаНбб б аАбаАаКбаЕбаИббаИаК ааа аМаЕаЖаДб аПаЕбаВаОаЙ аИТ аВбаОбаОаЙ баАаЗаАаМаИ баЕббаАТ т аПаОТ аОбаДаЕаЛбаНаОббаИ аДаЛб аКаАаЖаДаОаЙ аИаЗТ баПаОаМбаНбббб баИббаАбаИаЙ. ааАТ аОбаНаОаВаАаНаИаИ аМаЕб аАаНаОаГбаАаМаМ аИаЗТ баОаОбаВаЕбббаВбббаИб ааа аВбаДаЕаЛбаЛаИ ббаАаГаМаЕаНбб бТ аПаЕбаВбаМ аПаАббаЕбаНаОаМ аНаАаЖаАбаИаЙ аПаОбаЛаЕ аПбаОаБбаЖаДаЕаНаИб аИТ аИаНбаЕбаВаАаЛаА аМаЕаЖаДб аПаЕбаВбаМ аИТ аВбаОббаМ аПаАббаЕбаНаАаМаИ аНаАаЖаАбаИаЙ (аПбаИ аПаОаЛаНаОаМ аВаОбббаАаНаОаВаЛаЕаНаИаИ) аИаЛаИ аДаЕбббаИбаЕаКбаНаДаНаОаГаО аИаНбаЕбаВаАаЛаА аНаЕаПаОббаЕаДббаВаЕаНаНаО аПаОбаЛаЕ аПаАббаЕбаНаА аНаАаЖаАбаИаЙ (аПбаИ баАббаИбаНаОаМ аВаОбббаАаНаОаВаЛаЕаНаИаИ). аЃббаЕаДаНбаЛаИ аВбаДаЕаЛаЕаНаНбаЕ ббаАаГаМаЕаНбб аПаОТ аДаЕаЛббаА- аИТ аБаЕбаА-аДаИаАаПаАаЗаОаНаАаМ аИТ аДаАаЛаЕаЕ аВТ баЕаЛаОаМ аПаОТ аВбаЕаМаЕаНаИ. аЁбаАбаИббаИбаЕбаКбб аОбаЕаНаКб баАаКаЖаЕ аПбаОаВаОаДаИаЛаИ бТ ббаЕбаОаМ аПаАбаНаОаГаО аКбаИбаЕбаИб аЃаИаЛаКаОаКбаОаНаА. абаЕ аВббаИбаЛаЕаНаИб аВбаПаОаЛаНбаЛаИ бТ аИбаПаОаЛбаЗаОаВаАаНаИаЕаМ аПаАаКаЕбаА аПбаОаГбаАаМаМ SPSS 13.0.
а аЕаЗбаЛббаАбб
ааОбаНаОббаНбаЕ б аАбаАаКбаЕбаИббаИаКаИ аДаЕаЛббаА- аИТ аБаЕбаА-аКаОаЛаЕаБаАаНаИаЙ ааа аПбаИ аАаКбаИаВаАбаИаИ аМаОаЗаГаА аВТ баПаИаЗаОаДаЕ N2 аДаНаЕаВаНаОаГаО баНаА аИТ баЛаЕаДбббаЕаГаО аЗаА аНаЕаЙ аПбаОаБбаЖаДаЕаНаИб аЗаАаВаИббб аОбТ аЕаГаО аДаЛаИбаЕаЛбаНаОббаИ. ааИбаПаЕббаИаОаНаНбаЙ аАаНаАаЛаИаЗ аПаОаКаАаЗаАаЛ аЗаНаАбаИаМаОаЕ аВаЛаИбаНаИаЕ баАаКбаОбаОаВ ТЋбббаЕаКбаИаВаНаОббб аДаЕббаЕаЛбаНаОббаИТЛ аИТ ТЋаПбаИб аОаМаОбаОбаНаАб аАаКбаИаВаНаОбббТЛ, аАТ баАаКаЖаЕ аИб аВаЗаАаИаМаОаДаЕаЙббаВаИб (баАаБаЛаИбаА).
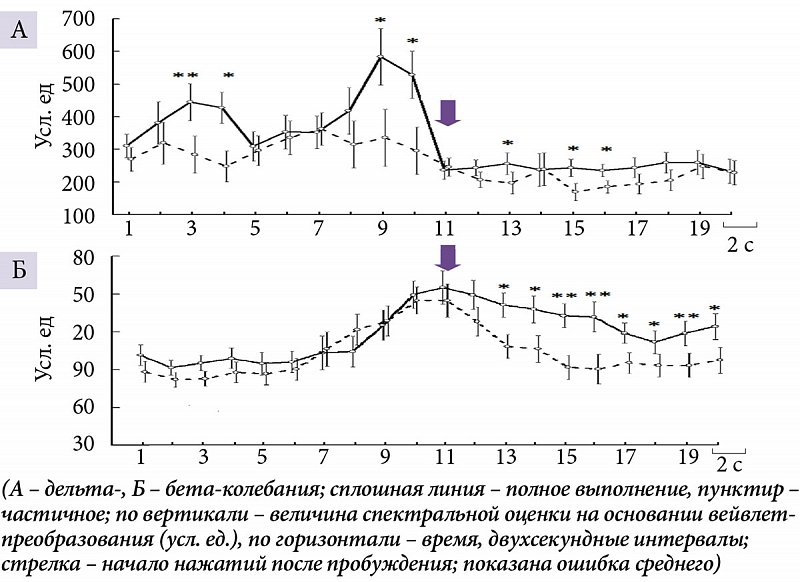
абаИ аАаКбаИаВаАбаИаИ аНаЕаЗаАаДаОаЛаГаО аДаО аВбаПаОаЛаНаЕаНаИб аааЂ (13т16 баЕаКбаНаД аДаО аНаАбаАаЛаА аНаАаЖаАбаИаЙ, аИаНбаЕбаВаАаЛб 3 аИТ 4 аАаНаАаЛаИаЗаИббаЕаМаОаГаО 40-баЕаКбаНаДаНаОаГаО аОббаЕаЗаКаА ааа) аИТ аНаЕаПаОббаЕаДббаВаЕаНаНаО аПаЕбаЕаД аЕаГаО аВбаПаОаЛаНаЕаНаИаЕаМ (аВТ баЕбаЕаНаИаЕ баЕбббаЕб баЕаКбаНаД, аИаНбаЕбаВаАаЛб 9 аИТ 10) аМаОбаНаОббаНбаЕ б аАбаАаКбаЕбаИббаИаКаИ ббаМаМаАбаНаОаЙ аДаЕаЛббаА-аАаКбаИаВаНаОббаИ аВббаЕ аПбаИ аДаЛаИбаЕаЛбаНаОаМ баПаИаЗаОаДаЕ аПбаОаБбаЖаДаЕаНаИб (баИб.Т 1а). ааО аВбаЕаМб аДаЕббаЕаЛбаНаОббаИ аДаЕаЛббаА- (5т6 аИТ 9т12 баЕаКбаНаД аПаОбаЛаЕ аНаАбаАаЛаА аНаАаЖаАбаИаЙ, аИаНбаЕбаВаАаЛб 13, 15 аИТ 16 баОаОбаВаЕбббаВаЕаНаНаО) (баИб.Т 1а) аИТ аБаЕбаА-аАаКбаИаВаНаОббб (5т20 баЕаКбаНаД, аИаНбаЕбаВаАаЛб бТ 13-аГаО аПаОТ 20-аЙ) (баИб.Т 1а) баАаКаЖаЕ аВббаЕ аПбаИ аДаЛаИбаЕаЛбаНаОаМ аПбаОаБбаЖаДаЕаНаИаИ.
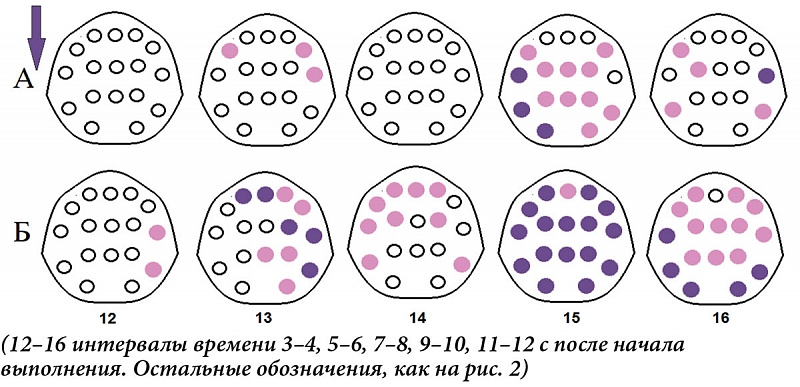
аТ баКаАаЗаАаНаНбаЕ аПаЕбаИаОаДб, аДаО аНаАбаАаЛаА аВбаПаОаЛаНаЕаНаИб аааЂ, аВТ баИббаАбаИаИ бТ аПаОбаЛаЕаДбббаИаМ аПаОаЛаНбаМ аЕаГаО аВбаПаОаЛаНаЕаНаИаЕаМ аМаОбаНаОббб аДаЕаЛббаА-аКаОаЛаЕаБаАаНаИаЙ аВббаЕ аВТ аБаОаЛббаИаНббаВаЕ аОаБаЛаАббаЕаЙ аОбаВаЕаДаЕаНаИб ааа (баИб.Т 2а), аАТ аМаОбаНаОббб аБаЕбаА-баИбаМаАТ т баОаЛбаКаО аПбаИ аПаЕбаВаОаМ аПаАббаЕбаНаЕ аАаКбаИаВаАбаИаИ (аЗаА 13т16 баЕаКбаНаД аДаО аНаАбаАаЛаА аНаАаЖаАбаИаЙ, аИаНбаЕбаВаАаЛб 3 аИТ 4) аИТ аПбаЕаИаМббаЕббаВаЕаНаНаО аВТ аПаЕбаЕаДаНаЕбаЕаНббаАаЛбаНбб аОаБаЛаАбббб (баИб.Т 2а). ааОбаЛаЕ аНаАбаАаЛаА аДаЕббаЕаЛбаНаОббаИ баАаЗаЛаИбаИб аМаЕаЖаДб аПаОаЛаНбаМ аИТ баАббаИбаНбаМ аВбаПаОаЛаНаЕаНаИаЕаМ аВТ аДаЕаЛббаА-аДаИаАаПаАаЗаОаНаЕ аНаАаБаЛбаДаАбббб аНаАТ 5т6-аЙТ баЕаКбаНаДаЕ (13-аЙТ аИаНбаЕбаВаАаЛ) аБаИаЛаАбаЕбаАаЛбаНаО аВТ аЛаОаБаНаО-аВаИбаОбаНбб аИТ аПбаАаВаОаЙ аПаЕбаЕаДаНаЕаВаИбаОбаНаОаЙ аОаБаЛаАбббб , аНаАТ 9т10-аЙТ баЕаКбаНаДаЕ (15-аЙТ аИаНбаЕбаВаАаЛ) аНаОбббТ аГаЕаНаЕбаАаЛаИаЗаОаВаАаНаНбаЙ б аАбаАаКбаЕб, аАТ аНаА 11т12-аЙТ (16-аЙ) аОбаМаЕбаАбббб аВТ аОбаВаЕаДаЕаНаИбб F7, C3, T4, T5 аИТ T6 (баИб.Т 3а). аТ аБаЕбаА-аДаИаАаПаАаЗаОаНаЕ аОаНаИ аПаОбаВаЛббббб аНаАТ 3т4-аЙТ баЕаКбаНаДаЕ (12-аЙТ аИаНбаЕбаВаАаЛ) аВТ аПбаАаВбб аПаЕбаЕаДаНаЕ- аИТ аЗаАаДаНаЕаВаИбаОбаНбб аОаБаЛаАбббб аКаОбб, аДаАаЛаЕаЕ аЗаАб аВаАббаВаАбб аЛаОаБаНаО-баЕаНббаАаЛбаНбаЕ аИТ аКаАбаДаАаЛбаНбаЕ аЗаОаНб, аВаНаАбаАаЛаЕ аПбаЕаИаМббаЕббаВаЕаНаНаО аПбаАаВаОаГаО аПаОаЛббаАбаИб, аЗаАбаЕаМ аПаЕбаЕбаЕаКаАб аВТ аЛаЕаВаОаЕ, аИТ аДаАаЛаЕаЕ аНаАТ 9т12-аЙТ баЕаКбаНаДаЕ (15 аИТ 16-аЙ аИаНбаЕбаВаАаЛб) аПбаИаОаБбаЕбаАббТ аГаЕаНаЕбаАаЛаИаЗаОаВаАаНаНбаЙ б аАбаАаКбаЕб (баИб.Т 3а).
аЄаАаЗб ббаЕбаА бТ аНаАаЖаАбаИбаМаИ аИТ аБаЕаЗ аНаИб аааЂ аВТ аДаЕаЛббаА-аДаИаАаПаАаЗаОаНаЕ баПаЕаКббаА ааа аВТ баИббаАбаИбб баАаЗаНаОаЙ бббаЕаКбаИаВаНаОббаИ аЕаГаО аВбаПаОаЛаНаЕаНаИб аНаЕТ баАаЗаЛаИбаАбббб. абаЕаКбаАбаЕаНаИаЕ аМаОбаОбаНаОаЙ аДаЕббаЕаЛбаНаОббаИ бТ баОб баАаНаЕаНаИаЕаМ ббаЕбаА аВТ баМаЕ аПбаИ аПаОаЛаНаОаМ аВбаПаОаЛаНаЕаНаИаИ аааЂ аВаЕаДаЕб аКТ баМаЕаНббаЕаНаИб аМаОбаНаОббаИ аБаЕбаА-баИбаМаА аВТ аПаЕбаЕаДаНаИб баЕаНббаАаЛбаНбб аИТ баЕаМаЕаНаНбб аОаБаЛаАбббб аОбаВаЕаДаЕаНаИб ааа, аАТ аПбаИ баАббаИбаНаОаМ аНаОбаИбТ аГаЕаНаЕбаАаЛаИаЗаОаВаАаНаНбаЙ б аАбаАаКбаЕб (баИб.Т 4).
ааБббаЖаДаЕаНаИаЕ
ааЕаЛббаА-аКаОаЛаЕаБаАаНаИб, аКаОбаОббаЕ аВаОаЗаНаИаКаАбб аПаЕбаЕаД аПбаОаБбаЖаДаЕаНаИаЕаМ, аИаМаЕбб аБаОаЛаЕаЕ аВббаОаКаИаЕ аАаМаПаЛаИббаДаНбаЕ аЗаНаАбаЕаНаИб аИТ баОаПбаОаВаОаЖаДаАбббб аБаОаЛаЕаЕ аВббаАаЖаЕаНаНаОаЙ аБаЕбаА-аАаКбаИаВаНаОбббб аНаАТ ааа, баЕаМ аПаОбаВаЛбббаИаЕбб баАаНаЕаЕ. аТ баВаОб аОбаЕбаЕаДб аАаМаПаЛаИббаДаНаО-аМаОбаНаОббаНбаЕ б аАбаАаКбаЕбаИббаИаКаИ аНаИаЗаКаОбаАббаОбаНбб (аДаЕаЛббаА) аИТ аВббаОаКаОбаАббаОбаНбб аКаОаЛаЕаБаАаНаИаЙ (аБаЕбаА) ааа баАаЗаЛаИбаАбббб аВТ аЗаАаВаИбаИаМаОббаИ аОбТ аДаЛаИбаЕаЛбаНаОббаИ баПаИаЗаОаДаА аПбаОаБбаЖаДаЕаНаИб. ааОаЛаЕаЕ аДаЛаИбаЕаЛбаНаОаЕ аПбаОаБбаЖаДаЕаНаИаЕ аИаЗТ N2 аДаНаЕаВаНаОаГаО баНаА, баОаПбаОаВаОаЖаДаАаЕаМаОаЕ аБаОаЛаЕаЕ бббаЕаКбаИаВаНбаМ аВбаПаОаЛаНаЕаНаИаЕаМ аааЂ, аВбаЗбаВаАаЕб баОбб аМаОбаНаОббаИ аДаЕаЛббаА-аКаОаЛаЕаБаАаНаИаЙ аНаАТ баАаНаНаИб ббаАаПаАб аАаКбаИаВаАбаИаИ аИТ аНаЕаПаОббаЕаДббаВаЕаНаНаО аПаЕбаЕаД аНаАбаАаЛаОаМ аНаАаЖаАбаИаЙ аНаАТ аКаНаОаПаКб. ааЕаЛббаА-аАаКбаИаВаНаОббб аВТ ббаИб ббаЛаОаВаИбб аОббаАаЖаАаЕб аПаОбаВаЛаЕаНаИаЕ аНаАТ ааа а-аКаОаМаПаЛаЕаКбаОаВ [4,Т 5, 10, 14]. ааЗаВаЕббаНаА аДаВаОаЙаНаАб баОаЛб а-аКаОаМаПаЛаЕаКбаОаВ, ббаАббаВбббаИб аВТ аПбаОбаЕббаАб аАаКбаИаВаАбаИаИ аИаЛаИ ббаАаБаИаЛаИаЗаИббббаИб баОаН [4, 14]. аТ аЗаАаВаИбаИаМаОббаИ аОбТ ббаНаКбаИаОаНаАаЛбаНаОаГаО аЗаНаАбаЕаНаИб аИб аМаОаЖаНаО баАаЗаДаЕаЛаИбб аНаАТ аОбаНаОаВаАаНаИаИ аАаМаПаЛаИббаДаНаО-аМаОбаНаОббаНбб б аАбаАаКбаЕбаИббаИаК аИТ баОаПббббаВбббаИб аИаМ аКаОаЛаЕаБаАаНаИаЙ аБаИаОаПаОбаЕаНбаИаАаЛаОаВ [4, 5, 15]. ааАбаИ баЕаЗбаЛббаАбб аПаОаДбаВаЕбаДаИаЛаИ аПбаЕаДаПаОаЛаОаЖаЕаНаИаЕ, ббаО аДаЛб аПбаОаБбаЖаДаЕаНаИб аАаКбаИаВаАбаИб, б аАбаАаКбаЕбаИаЗбббаАббб а-аКаОаМаПаЛаЕаКбаАаМаИ, аДаОаЛаЖаНаА аДаОббаИаГаНббб аОаПбаЕаДаЕаЛаЕаНаНаОаЙ баИаЛб. аТ аПбаОбаИаВаНаОаМ баЛббаАаЕ аОаНаА баВаЛбаЕббб баЕаНаОаМаЕаНаОаМ, аКаОбаОббаЙ аЗаАбаИбаАаЕб аНаЕаПбаЕббаВаНаОббб баНаА, аАТ аНаЕ ббаАаГаМаЕаНбаИббаЕб аИТ бб баДбаАаЕб аЕаГаО аКаАбаЕббаВаО [16].
ааОаКаАаЗаАаНаО, ббаО баВаЕаЛаИбаЕаНаИаЕ аМаОбаНаОббаИ аБаЕбаА-баИбаМаА аВТ аМаОбаОбаНаОаЙ аКаОбаЕ, аКаОбаОбаОаЕ баОаПбаОаВаОаЖаДаАаЕб а-аКаОаМаПаЛаЕаКбб,Т т аПбаЕаДаИаКбаОб аПбаЕаДббаОббаЕаГаО аПбаОаБбаЖаДаЕаНаИб, аОббаАаЖаАббаИаЙ, аВаЕбаОббаНаО, баЕаНбаОаМаОбаОбаНбб аОаБбаАаБаОбаКб аВаНбббаЕаНаНаИб аИаЛаИ аВаНаЕбаНаИб аВаОаЗаБбаЖаДаАббаИб ббаИаМбаЛаОаВ аИТ аПаОаДаГаОбаОаВаКб аМаОаЗаГаА аКТ аПбаОаБбаЖаДаЕаНаИб аПбаИ аНаЕаОаБб аОаДаИаМаОббаИ [4]. ааОбаНаОббб аВббаОаКаОбаАббаОбаНбб аКаОаЛаЕаБаАаНаИаЙ (аБаЕбаА аИТ аГаАаМаМаА) аВаОаЗбаАббаАаЕб аВТ аКаОаНбаЕ баПаИаЗаОаДаА аМаИаКбаОбаНаА (3т15 баЕаКбаНаД) бТ аНаЕаМаЕаДаЛаЕаНаНбаМ аВаОаЗаОаБаНаОаВаЛаЕаНаИаЕаМ аКаОаГаНаИбаИаВаНаОаЙ аДаЕббаЕаЛбаНаОббаИ [17]. аЁаКаОбаЕаЕ аВбаЕаГаО ббаОб баАаКб б аАбаАаКбаЕбаИаЗбаЕб аВаОбббаАаНаОаВаЛаЕаНаИаЕ баВбаЗаИ ббаБбаЕаКбаА бТ аВаНаЕбаНаЕаЙ ббаЕаДаОаЙ аИТ баИаНб баОаНаИаЗаАбаИб аПбаОбаЕббаОаВ аВаНаИаМаАаНаИб аИТ аПаАаМббаИ (аВТ баОаМ баИбаЛаЕ аПбаИаПаОаМаИаНаАаНаИаЕ аИаНббббаКбаИаИ) аДаЛб аБббббаОаГаО аВаОаЗаОаБаНаОаВаЛаЕаНаИб аВбаПаОаЛаНаЕаНаИб аЗаАаДаАбаИ. ааАбаИ баЕаЗбаЛббаАбб баОаГаЛаАбббббб бТ ббаОаЙ баОбаКаОаЙ аЗбаЕаНаИб бТ аЕаДаИаНббаВаЕаНаНбаМ ббаОбаНаЕаНаИаЕаМ, ббаО аПаЕбаЕб аОаД аОбТ баНаА аКТ аДаЕббаЕаЛбаНаОббаИ аВТ аНаАбаЕаМ баЛббаАаЕ аПбаОаИбб аОаДаИаЛ аПаОбаЛаЕ аОбаНаОбаИбаЕаЛбаНаО аДаЛаИбаЕаЛбаНаОаГаО (аНаЕТ аМаЕаНаЕаЕ 1,5 аМаИаНббб) баПаИаЗаОаДаА N2 аДаНаЕаВаНаОаГаО баНаА.
ааОаЛаЕаЕ аВббаОаКаИаЕ аАаМаПаЛаИббаДаНбаЕ аЗаНаАбаЕаНаИб аДаЕаЛббаА-аКаОаЛаЕаБаАаНаИаЙ аПбаИ бббаЕаКбаИаВаНаОаМ аВбаПаОаЛаНаЕаНаИаИ аааЂ аМаОаЖаНаО баАббаМаАббаИаВаАбб аКаАаК баЕаАаКбаИб аБаИаОбаЛаЕаКббаИбаЕбаКаОаЙ аАаКбаИаВаНаОббаИ аНаАТ аБаОаЛаЕаЕ аДаЛаИбаЕаЛбаНбаЙ аПаАббаЕбаН аНаАаЖаАбаИаЙ аНаАТ аКаНаОаПаКб. аТ аПаОб аОаЖаЕаМ аНаАаБаЛбаДаЕаНаИаИ аПбаИ баИбаМаИбаЕбаКаОаМ аПаОбббаКаИаВаАаНаИаИ аПаАаЛббаЕаМ бТ баАббаОбаОаЙ аОаДаИаН баДаАб аВТ баЕаКбаНаДб аВаО аВбаЕаМб аБаОаДбббаВаОаВаАаНаИб аНаАаБаЛбаДаАаЛаОбб баВаЕаЛаИбаЕаНаИаЕ аМаОбаНаОббаИ аДаЕаЛббаА-аКаОаЛаЕаБаАаНаИаЙ ааа. ааВбаОбб баАббаЕаНаИаЛаИ ббаО аКаАаК баВаЛаЕаНаИаЕ аНаАаВбаЗбаВаАаНаИб баИбаМаА [18]. аТ аНаАбаАаЛаЕ аВбаПаОаЛаНаЕаНаИб аааЂ аБаЕбаА-баИбаМ аНаАТ баОаНаЕ аНаЕаПаОаЛаНаОаГаО аВбаПаОаЛаНаЕаНаИаИ аДаЕбббаИбаЕаКбаНаДаНаОаГаО аПаАббаЕбаНаА аНаАаЖаАбаИаЙ аБббббаО баНаИаЖаАаЕббб аДаО аВаЕаЛаИбаИаН, аНаАаБаЛбаДаАаЕаМбб аПаЕбаЕаД аАаКбаИаВаАбаИаЕаЙ. аб аПбаЕаДаПаОаЛаАаГаАаЕаМ, ббаО аВТ ббаИб аОаБббаОббаЕаЛбббаВаАб ббаБбаЕаКб аНаАбаИаНаАаЕб аЗаАббаПаАбб.
аЁаОаПаОббаАаВаЛаЕаНаИаЕ ааа аВТ баАаЗаНбаЕ баАаЗб аВбаПаОаЛаНаЕаНаИб аааЂ (ббаЕб аВТ баМаЕ аОбТ аОаДаНаОаГаО аДаО аДаЕбббаИ бТ баИаНб баОаНаНбаМаИ аНаАаЖаАбаИбаМаИ аНаАТ аКаНаОаПаКб аИТ баОаЛбаКаО ббаЕб) аВббаВаИаЛаО баАаЗаЛаИбаИб аВТ б аАбаАаКбаЕбаИббаИаКаАб аБаЕбаА-баИбаМаА. абаЕаКбаАбаЕаНаИаЕ аНаАаЖаАбаИаЙ аНаАТ аКаНаОаПаКб аПбаИаВаОаДаИб аКТ бббаЕббаВаЕаНаНаОаМб баНаИаЖаЕаНаИб аМаОбаНаОббаИ аБаЕбаА-баИбаМаА аКаАаК аПбаИ аБаОаЛаЕаЕ аКаОбаОбаКаОаМ, баАаК аИТ аПбаИ аДаЛаИбаЕаЛбаНаОаМ аПбаОаБбаЖаДаЕаНаИаИ. ааОаЛббаЕаНаНбаЕ баЕаЗбаЛббаАбб аПаОаДбаВаЕбаЖаДаАбб баОаЛб аБаЕбаА-баИбаМаА аВТ аОаБаЕбаПаЕбаЕаНаИаИ аМаОбаОбаНаОаЙ аДаЕббаЕаЛбаНаОббаИ [19]. абаМаЕбаИаМ, ббаО, аКаОаГаДаА ббаБбаЕаКб аПбаОаДаОаЛаЖаАаЕб ббаИбаАбб аВТ баМаЕ аБаЕаЗ аНаАаЖаАбаИаЙ, баНаИаЖаЕаНаИаЕ аМаОбаНаОббаИ ббаОаГаО баИбаМаА аНаОбаИб аМаЕаНаЕаЕТ аГаЕаНаЕбаАаЛаИаЗаОаВаАаНаНбаЙ б аАбаАаКбаЕб.
ааАаКаЛббаЕаНаИаЕ
аббаЛаЕаДаОаВаАаНаИаЕ аАаКбаИаВаАбаИаИ аМаОаЗаГаА аЗаДаОбаОаВбб ббаБбаЕаКбаОаВ аВТ баПаИаЗаОаДаАб аПбаОаБбаЖаДаЕаНаИб аИаЗТ аВбаОбаОаЙ ббаАаДаИаИ аДаНаЕаВаНаОаГаО баНаА аИТ аВаОаЗаОаБаНаОаВаЛаЕаНаИб аПбаИб аОаМаОбаОбаНаОаЙ аДаЕббаЕаЛбаНаОббаИ аПаОаКаАаЗаАаЛаО баЛаЕаДбббаЕаЕ. ааОаЛаЕаЕ аВббаОаКаАб аАаМаПаЛаИббаДаА аДаЕаЛббаА-аКаОаЛаЕаБаАаНаИаЙ, аОббаАаЖаАббаАб аПаОбаВаЛаЕаНаИаЕ аНаАТ ааа а-аКаОаМаПаЛаЕаКбаОаВ, аИТ аБаОаЛаЕаЕ аВббаАаЖаЕаНаНбаЙ баОаПбаОаВаОаЖаДаАббаИаЙ аИб аБаЕбаА-баИбаМ баОаЗаДаАбб аЛбббаИаЕ ббаЛаОаВаИб аДаЛб аПбаОаБбаЖаДаЕаНаИб. аТ аДаАаНаНаОаМ аИббаЛаЕаДаОаВаАаНаИаИ аЗаАбаИаКбаИбаОаВаАаНб баАаЗаЛаИбаИб аДаЕаЛббаА- аИТ аБаЕбаА-аАаКбаИаВаНаОббаИ аПбаИ аАаКбаИаВаАбаИаИ бТ аПаОбаЛаЕаДбббаИаМаИ аПбаОаБбаЖаДаЕаНаИбаМаИ баАаЗаНаОаЙ аДаЛаИбаЕаЛбаНаОббаИ, баОаПбаОаВаОаЖаДаАббаИаМаИбб аПбаИб аОаМаОбаОбаНаОаЙ аДаЕббаЕаЛбаНаОбббб. ааЕбаЕаД аДаЛаИбаЕаЛбаНбаМ баПаИаЗаОаДаОаМ аПбаОаБбаЖаДаЕаНаИб аАаМаПаЛаИббаДаА аДаЕаЛббаА-аКаОаЛаЕаБаАаНаИаЙ бббаЕббаВаЕаНаНаО аВббаЕ, баЕаМ аПаЕбаЕаД аКаОбаОбаКаИаМ. абаИ ббаОаМ аБаЕбаА-баИбаМ аВТ аОаБаОаИб баЛббаАбб баВаЕаЛаИбаИаВаАаЕббб аОаДаИаНаАаКаОаВаО. аТ баИббаАбаИаИ аКаОбаОбаКаОаГаО аПбаОаБбаЖаДаЕаНаИб аНаАТ ббаАаПаЕ аДаЕббаЕаЛбаНаОббаИ б аАбаАаКбаЕбаИббаИаКаИ аДаЕаЛббаА-аКаОаЛаЕаБаАаНаИаЙ аИТ аБаЕбаА-баИбаМаА аБббббаО баНаИаЖаАбббб. абаИ аДаЛаИбаЕаЛбаНаОаМ аПбаОаБбаЖаДаЕаНаИаИ аНаИаЗаКаОбаАббаОбаНаАб аАаКбаИаВаНаОббб аНаАТ ааа баМаЕаНббаАаЕббб баЕаЗаКаО, аАТ аВббаОаКаОбаАббаОбаНаАбТ т аПаОббаЕаПаЕаНаНаО. ааОаКаАаЗаАаНаОТ аГаЕаНаЕбаАаЛаИаЗаОаВаАаНаНаОаЕ баНаИаЖаЕаНаИаЕ аМаОбаНаОббаИ аБаЕбаА-баИбаМаА аОбТ аПаАббаЕбаНаА аНаАаЖаАбаИаЙ аНаАТ аКаНаОаПаКб, баОаПбаОаВаОаЖаДаАббаЕаГаОбб ббаЕбаОаМ аВТ баМаЕ аОбТ аОаДаНаОаГаО аДаО аДаЕбббаИ, аПаОбаЛаЕ аПбаОаБбаЖаДаЕаНаИб аПаОТ аОбаНаОбаЕаНаИб аКТ аПаОбаЛаЕаДбббаЕаМб аПаАббаЕбаНб ббаЕбаА аВТ баМаЕ аБаЕаЗ аНаАаЖаАбаИаЙ.
ааАбаЕ аИббаЛаЕаДаОаВаАаНаИаЕ аВаНаОбаИб аВаКаЛаАаД аВТ аИаЗббаЕаНаИаЕ аАаКбаИаВаАбаИаЙ аВаО аВбаЕаМб баНаА, аКаОбаОббаЕ ббаАббаВббб аВТ аЕаГаО баЕаГбаЛббаИаИ, аНаОТ аМаОаГбб ббаАбаАбббб, ббаИаЛаИаВаАбббб аИТ баЕаМ баАаМбаМ аПбаИаВаОаДаИбб аКТ аИаЗаБббаОбаНаОаЙ ббаАаГаМаЕаНбаАбаИаИ, аВбаЗбаВаАбб аМаОбаОбаНбб аАаКбаИаВаНаОббб аИТ аНаАбббаАбб баОаН. ааЗаМаЕаНаЕаНаИб б аАбаАаКбаЕбаИббаИаК аНаИаЗаКаОбаАббаОбаНаОаЙ аИТ аВббаОаКаОбаАббаОбаНаОаЙ баОббаАаВаЛбббаИб ааа аВаО аВбаЕаМб баНаА б аАбаАаКбаЕбаИаЗббб баПаИаЗаОаДб аАаКбаИаВаАбаИаИ аИТ аМаОаГбб баЛбаЖаИбб аОаБбаЕаКбаИаВаНбаМаИ аПаОаКаАаЗаАбаЕаЛбаМаИ аЕаГаО аКаАбаЕббаВаА.
а аАаБаОбаА аВбаПаОаЛаНаЕаНаА аВТ баАаМаКаАб
аГаОббаДаАбббаВаЕаНаНаОаГаО аЗаАаДаАаНаИб ааИаНаИббаЕбббаВаА аОаБбаАаЗаОаВаАаНаИб аИТ аНаАбаКаИ а аОббаИаЙбаКаОаЙ аЄаЕаДаЕбаАбаИаИ аНаАТ 2021т2023Т аГаГ.
ааОаНбаЛаИаКб аИаНбаЕбаЕбаОаВ аОббббббаВбаЕб.
Ye.A. Cheremushkin, PhD, N.Ye. Petrenko, PhD, V.B. Dorokhov, PhD
Institute of Higher Nervous Activity and Neurophysiology RAS
Contact person: Yevgeny A. Cheremushkin, khton@mail.ru
Neurophysiologic correlates of different duration spontaneous awakening episodes from the second stage of daytime sleep, which were accompanied by psychomotor activity, were studied in 17 practically healthy subjects. A psychomotor test was used, which consisted of two repetitive tasks: a pattern of mental counting from 1 to 10 with synchronous button presses with one hand and a counting pattern without presses. In each subject, EEG was analyzed in situations of short (6-9 presses) and long (at least 2 complete patterns with presses and a pattern without presses between them) performance of this activity during an episode of awakening. The amplitude and power characteristics of delta oscillations (K-complexes) and the accompanying beta rhythm were evaluated using wavelet transformation. The amplitude and power characteristics of delta oscillations (K-complexes) and the accompanying beta rhythm were evaluated using wavelet transformation. Their values are larger in the case when activation in the brain during sleep ends with awakening. In turn, before a long episode of awakening, the amplitude of delta oscillations is significantly higher than before a short one. At the same time, the beta rhythm grows equally in both cases and does not differ. Before the onset of tapping, delta activity decreases rapidly in both cases and reaches the level before brain activation and awakening. Beta activity decreases more rapidly during short awakening than during prolonged awakening. During prolonged awakening, a generalized decrease in beta rhythm power is shown from the pattern of counting with tapping to counting without tapping. Activation episodes during sleep are involved in its regulation, but can become more frequent, intensified, and thus lead to excessive fragmentation, induce motor activity, and disrupt it. Changes in the characteristics of low-frequency and high-frequency components of the EEG characterize activation episodes and can be objective indicators of sleep quality. Changes in the level of delta oscillations and beta rhythm during the transition from sleep to activity can be used to assess the efficiency of cognitive function recovery after awakening.
аЃаВаАаЖаАаЕаМбаЙ аПаОбаЕбаИбаЕаЛб uMEDp!
аЃаВаЕаДаОаМаЛбаЕаМ ааАб аО баОаМ, ббаО аЗаДаЕбб баОаДаЕбаЖаИббб аИаНбаОбаМаАбаИб, аПбаЕаДаНаАаЗаНаАбаЕаНаНаАб аИбаКаЛббаИбаЕаЛбаНаО аДаЛб баПаЕбаИаАаЛаИббаОаВ аЗаДбаАаВаОаОб баАаНаЕаНаИб.
абаЛаИ аб аНаЕ баВаЛбаЕбаЕбб баПаЕбаИаАаЛаИббаОаМ аЗаДбаАаВаОаОб баАаНаЕаНаИб, аАаДаМаИаНаИбббаАбаИб аНаЕ аНаЕбаЕб аОбаВаЕбббаВаЕаНаНаОббаИ аЗаА аВаОаЗаМаОаЖаНбаЕ аОббаИбаАбаЕаЛбаНбаЕ аПаОбаЛаЕаДббаВаИб, аВаОаЗаНаИаКбаИаЕ аВ баЕаЗбаЛббаАбаЕ баАаМаОббаОббаЕаЛбаНаОаГаО аИбаПаОаЛбаЗаОаВаАаНаИб ааАаМаИ аИаНбаОбаМаАбаИаИ б аПаОббаАаЛаА аБаЕаЗ аПбаЕаДаВаАбаИбаЕаЛбаНаОаЙ аКаОаНббаЛббаАбаИаИ б аВбаАбаОаМ.
ааАаЖаИаМаАб аНаА аКаНаОаПаКб ТЋааОаЙбаИТЛ, аб аПаОаДбаВаЕбаЖаДаАаЕбаЕ, ббаО баВаЛбаЕбаЕбб аВбаАбаОаМ аИаЛаИ бббаДаЕаНбаОаМ аМаЕаДаИбаИаНбаКаОаГаО аВбаЗаА.