ąźą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ąĖ ąŁąŁąō ą┐čĆąĖ ą▓ąŠąĘą▒čāąČą┤ąĄąĮąĖčÅčģ ą╝ąŠąĘą│ą░ ą▓ąŠ┬Āą▓čĆąĄą╝čÅ ą▓č鹊čĆąŠą╣ čüčéą░ą┤ąĖąĖ ą┤ąĮąĄą▓ąĮąŠą│ąŠ čüąĮą░, čüąŠą┐čĆąŠą▓ąŠąČą┤ą░čÄčēąĖčģčüčÅ čüą░ą╝ąŠą┐čĆąŠąĖąĘą▓ąŠą╗čīąĮčŗą╝ąĖ ą╝ąĖą║čĆąŠą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅą╝ąĖ ąĖ┬Āą╝ąĖąĮąĖą╝ą░ą╗čīąĮąŠą╣ ą┐ąŠą▓ąĄą┤ąĄąĮč湥čüą║ąŠą╣ ą░ą║čéąĖą▓ąĮąŠčüčéčīčÄ ą▓ čüąĖčéčāą░čåąĖąĖ ą▓čŗą┐ąŠą╗ąĮąĄąĮąĖčÅ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░
- ąÉąĮąĮąŠčéą░čåąĖčÅ
- ąĪčéą░čéčīčÅ
- ąĪčüčŗą╗ą║ąĖ
- English

ąÆą▓ąĄą┤ąĄąĮąĖąĄ
ąśąĘčāč湥ąĮąĖąĄ ą▓ąŠąĘą▒čāąČą┤ąĄąĮąĖą╣ ą╝ąŠąĘą│ą░ ą▓┬ĀčüąŠčüč鹊čÅąĮąĖąĖ čüąĮą░, ą║ąŠč鹊čĆčŗąĄ ąĘą░ą║ą░ąĮčćąĖą▓ą░čÄčéčüčÅ ą┤ą▓ąĖą│ą░č鹥ą╗čīąĮąŠą╣ ą░ą║čéąĖą▓ąĮąŠčüčéčīčÄ ąĖ┬Āą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅą╝ąĖ, ą▓ą░ąČąĮąŠ ą┤ą╗čÅ ąŠčåąĄąĮą║ąĖ ąĮąĄčüčéą░ą▒ąĖą╗čīąĮąŠčüčéąĖ čŹč鹊ą│ąŠ čüąŠčüč鹊čÅąĮąĖčÅ ą▓┬Āčāčüą╗ąŠą▓ąĖčÅčģ ą▓ą╗ąĖčÅąĮąĖčÅ ąĮą░┬ĀąĮąĄą│ąŠ čüąŠčåąĖą░ą╗čīąĮčŗčģ ąĖ┬Ā菹║ąŠą╗ąŠą│ąĖč湥čüą║ąĖčģ čäą░ą║č鹊čĆąŠą▓ [1], ą┐čĆąĖ ąĖčüčüą╗ąĄą┤ąŠą▓ą░ąĮąĖąĖ čĆą░čüčüčéčĆąŠą╣čüčéą▓ ą▓ąŠąĘą▒čāąČą┤ąĄąĮąĖą╣ ą▓ąŠ ą▓čĆąĄą╝čÅ čüąĮą░, ą║ąŠč鹊čĆčŗąĄ ą┐čĆąĖą▓ąŠą┤čÅčé ą║┬ĀąĮąĄą┐čĆąŠąĖąĘą▓ąŠą╗čīąĮąŠą╣ ą┤ą▓ąĖą│ą░č鹥ą╗čīąĮąŠą╣ ą░ą║čéąĖą▓ąĮąŠčüčéąĖ čĆą░ąĘą╗ąĖčćąĮąŠą╣ ą┤ą╗ąĖč鹥ą╗čīąĮąŠčüčéąĖ ąĖ┬Āčüą╗ąŠąČąĮąŠčüčéąĖ┬Ā[2].┬Ā ą×ąĮąŠ čéą░ą║ąČąĄ ą╝ąŠąČąĄčé ą▒čŗčéčī ą┐ąŠą╗ąĄąĘąĮąŠ čüą┐ąĄčåąĖą░ą╗ąĖčüčéą░ą╝ ą┐čĆąĖ ąŠčåąĄąĮą║ąĄ čĆąĖčüą║ąŠą▓, ą║ąŠč鹊čĆčŗąĄ ą▓čŗąĘčŗą▓ą░čÄčé čüąŠąĘąĮą░č鹥ą╗čīąĮčŗąĄ čĆąĄą░ą║čåąĖąĖ čüčāą▒čŖąĄą║čéą░ (ą▓┬Āč鹊ą╝ čćąĖčüą╗ąĄ čüąĮąŠą▓ąĖą┤ąĄąĮąĖčÅ) ą▓ąŠ ą▓čĆąĄą╝čÅ ą░ąĮąĄčüč鹥ąĘąĖąĖ ąĖ┬Āą┐ąŠčüą╗ąĄ ąŠą┐ąĄčĆą░čåąĖąĖ┬Ā[3].
ąÆ┬ĀąĖčüčüą╗ąĄą┤ąŠą▓ą░ąĮąĖčÅčģ ą▓ąĮčāčéčĆąĖą╝ąŠąĘą│ąŠą▓čŗčģ ąĘą░ą┐ąĖčüąĄą╣ ą▒ąĖąŠčŹą╗ąĄą║čéčĆąĖč湥čüą║ąŠą╣ ą░ą║čéąĖą▓ąĮąŠčüčéąĖ ą┐ą░čåąĖąĄąĮč鹊ą▓ čü┬Ā菹┐ąĖą╗ąĄą┐čüąĖąĄą╣ ą▓┬Āą┐ąĄčĆąĖąŠą┤ čĆąĄą╝ąĖčüčüąĖąĖ ą┐ąŠą║ą░ąĘą░ąĮąŠ, čćč鹊 ą┐ąĄčĆąĄčģąŠą┤čŗ ą║ąŠčĆčŗ ąŠčé┬ĀčüąĮą░ ą║┬Āą▒ąŠą┤čĆčüčéą▓ąŠą▓ą░ąĮąĖčÄ ąĮąŠčüčÅčé ą┐čĆąŠą┤ąŠą╗ąČąĖč鹥ą╗čīąĮčŗą╣ čģą░čĆą░ą║č鹥čĆ ąĖ┬Āą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ ą╝ąŠąČąĮąŠ čĆą░čüčüą╝ą░čéčĆąĖą▓ą░čéčī ą║ą░ą║ čĆą░ąĘąĮčŗąĄ čüąŠčüč鹊čÅąĮąĖčÅ ą╝ąŠąĘą│ą░. ąóą░ą╗ą░ą╝ąĖč湥čüą║ą░čÅ ą░ą║čéąĖą▓ąĮąŠčüčéčī ąĄą┤ąĖąĮąŠąŠą▒čĆą░ąĘąĮą░ ąĖ┬ĀčüąŠąŠčéą▓ąĄčéčüčéą▓čāąĄčé čüąŠčüč鹊čÅąĮąĖčÄ ą╝ąĄąČą┤čā ą▒ąŠą┤čĆčüčéą▓ąŠą▓ą░ąĮąĖąĄą╝ ąĖ┬ĀčüąĮąŠą╝. ą¤čĆąĖ čŹč鹊ą╝ ą░ą║čéąĖą▓ą░čåąĖčÅ ą║ąŠčĆčŗ ąĮąĄąŠą┤ąĮąŠčĆąŠą┤ąĮą░ ąĖ┬ĀąŠą▒čāčüą╗ąŠą▓ą╗ąĄąĮą░ čĆą░ąĘąĮčŗą╝ąĖ čäą░ą║č鹊čĆą░ą╝ąĖ čĆąĄą│čāą╗čÅčåąĖąĖ. ąĢąĄ čüą┐ąĄą║čéčĆą░ą╗čīąĮčŗą╣ čüąŠčüčéą░ą▓ ąĘą░ą▓ąĖčüąĖčé ąŠčé┬Āą╝ąŠą┤ą░ą╗čīąĮąŠčüčéąĖ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ (čüą┐ąŠąĮčéą░ąĮąĮą░čÅ ąĖą╗ąĖ ąĖąĮą┤čāčåąĖčĆąŠą▓ą░ąĮąĮą░čÅ),┬Āą│ąŠą╝ąĄąŠčüčéą░ąĘą░, čüčéą░ą┤ąĖąĖ čüąĮą░ ąĖ┬ĀąŠą▒ą╗ą░čüčéąĖ ąŠčéą▓ąĄą┤ąĄąĮąĖčÅ [4, 5]. ąŁčéąĖ čüąŠčüč鹊čÅąĮąĖčÅ čüąŠą┐čĆąŠą▓ąŠąČą┤ą░čÄčéčüčÅ čéą░ą║ąĖą╝ čÅą▓ą╗ąĄąĮąĖąĄą╝, ą║ą░ą║ ą╗ąŠą║ą░ą╗čīąĮąŠąĄ ą▒ąŠą┤čĆčüčéą▓ąŠą▓ą░ąĮąĖąĄ, ą║ąŠą│ą┤ą░ ąĮą░┬Āč乊ąĮąĄ ą░ą║čéąĖą▓ąĮąŠčüčéąĖ, ą┐ąŠčģąŠąČąĄą╣ ąĮą░┬ĀčüąŠąĮ, ąĮą░ą▒ą╗čÄą┤ą░ąĄą╝ąŠą╣ ą▓┬Āą▒ąŠą╗čīčłąĖąĮčüčéą▓ąĄ ą║ąŠčĆą║ąŠą▓čŗčģ ąŠą▒ą╗ą░čüč鹥ą╣ ą╝ąŠąĘą│ą░, ą┐ąŠčÅą▓ą╗čÅčÄčéčüčÅ ą┐ą░čéč鹥čĆąĮčŗ, čüąŠąŠčéą▓ąĄčéčüčéą▓čāčÄčēąĖąĄ čüąŠčüč鹊čÅąĮąĖčÄ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ [6]. ąÆ┬ĀčŹčéąĖčģ ą┐ą░čéč鹥čĆąĮą░čģ ąŠčéą╝ąĄčćą░ąĄčéčüčÅ ą▒ąŠą╗ąĄąĄ ą▓čŗčĆą░ąČąĄąĮąĮąŠąĄ ą┐čĆąĖčüčāčéčüčéą▓ąĖąĄ ą▓čŗčüąŠą║ąŠčćą░čüč鹊čéąĮčŗčģ ą║ąŠą╗ąĄą▒ą░ąĮąĖą╣ 菹╗ąĄą║čéčĆąŠčŹąĮčåąĄčäą░ą╗ąŠą│čĆą░ą╝ą╝čŗ (ąŁąŁąō), ą║ąŠč鹊čĆčŗąĄ ą╗ąĖą▒ąŠ ąĘą░ą╝ąĄčēą░čÄčé ąĮąĖąĘą║ąŠčćą░čüč鹊čéąĮčŗąĄ, ą╗ąĖą▒ąŠ ąĮą░ą║ą╗ą░ą┤čŗą▓ą░čÄčéčüčÅ ąĮą░┬ĀąĮąĖčģ [7]. ąŁč鹊 čÅą▓ą╗ąĄąĮąĖąĄ ą╝ąŠąČąĄčé ąŠčéčĆą░ąČą░čéčī čüąĄąĮčüąŠą╝ąŠč鹊čĆąĮčāčÄ ąŠą▒čĆą░ą▒ąŠčéą║čā ą▓ąĮąĄčłąĮąĖčģ čĆą░ąĘą┤čĆą░ąČąĖč鹥ą╗ąĄą╣ ąĖ┬Āą│ąŠč鹊ą▓ąĖčéčī ą╝ąŠąĘą│ ą║┬Āą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÄ [8] ąĖą╗ąĖ ąČąĄ čüąŠą┐čĆąŠą▓ąŠąČą┤ą░čéčī ą┤ą▓ąĖą│ą░č鹥ą╗čīąĮčāčÄ ą░ą║čéąĖą▓ąĮąŠčüčéčī ą▒ąĄąĘ ąĄąĄ ąŠčüąŠąĘąĮą░ą▓ą░ąĮąĖčÅ [2, 9, 10].
ąÆ┬ĀąĖčüčüą╗ąĄą┤ąŠą▓ą░ąĮąĖčÅčģ ąŁąŁąō ą▓ąŠ ą▓čĆąĄą╝čÅ čŹą┐ąĖąĘąŠą┤ąŠą▓ čüą┐ąŠąĮčéą░ąĮąĮčŗčģ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖą╣ ą▓ąŠ ą▓čĆąĄą╝čÅ ąĮąŠčćąĮąŠą│ąŠ čüąĮą░, ą║ąŠč鹊čĆčŗąĄ ąĖčüą┐čŗčéčāąĄą╝čŗąĄ ą┐ąŠą┤čéą▓ąĄčƹȹ┤ą░ą╗ąĖ ąĮą░ąČą░čéąĖčÅą╝ąĖ ąĮą░┬Āą║ąĮąŠą┐ą║čā, ą▒čŗą╗ąŠ ą┐ąŠą║ą░ąĘą░ąĮąŠ, čćč鹊 ą▓┬Āą▒ąŠą╗čīčłąĖąĮčüčéą▓ąĄ čüą╗čāčćą░ąĄą▓ čŹčéąĖą╝ ąĮą░ąČą░čéąĖčÅą╝ ą┐čĆąĄą┤čłąĄčüčéą▓ąŠą▓ą░ą╗ ą┐ąĄčĆąĖąŠą┤ ą┐ąŠčÅą▓ą╗ąĄąĮąĖčÅ ą▓┬Āą▒ąĖąŠčŹą╗ąĄą║čéčĆąĖč湥čüą║ąŠą╣ ą░ą║čéąĖą▓ąĮąŠčüčéąĖ ą░ą╗čīčäą░-čĆąĖčéą╝ą░ [11]. ą×čüąŠą▒ąĄąĮąĮąŠčüčéčī ąĮą░čłąĄą╣ čĆą░ą▒ąŠčéčŗ čüąŠčüč鹊ąĖčé ą▓┬Āč鹊ą╝, čćč鹊 ą╝čŗ ąĖčüčüą╗ąĄą┤čāąĄą╝ ą░ą║čéąĖą▓ą░čåąĖąĖ ą╝ąŠąĘą│ą░ čü┬Āą┐ąŠčüą╗ąĄą┤čāčÄčēąĖą╝ ą║ąŠčĆąŠčéą║ąĖą╝ 菹┐ąĖąĘąŠą┤ąŠą╝ čüą░ą╝ąŠą┐čĆąŠąĖąĘą▓ąŠą╗čīąĮąŠą│ąŠ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ, čüąŠą┐čĆąŠą▓ąŠąČą┤ą░čÄčēąĄą│ąŠčüčÅ ą▓ąŠčüčüčéą░ąĮąŠą▓ą╗ąĄąĮąĖąĄą╝ ą▓čŗą┐ąŠą╗ąĮąĄąĮąĖčÅ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░, ą┐čĆąĄčĆčŗą▓ą░ąĄą╝ąŠą│ąŠ čüąĮąŠą╝. ą£čŗ ą┐čĆąĄą┤ą┐ąŠą╗ą░ą│ą░ąĄą╝, čćč鹊 čĆą░ąĘą╗ąĖčćąĖčÅ ą▓┬Āą┐ąŠą▓ąĄą┤ąĄąĮąĖąĖ (ą▓┬ĀčćąĖčüą╗ąĄ ąĮą░ąČą░čéąĖą╣ ąĮą░┬Āą║ąĮąŠą┐ą║čā) ąĖ┬ĀąŁąŁąō, ą║ąŠč鹊čĆą░čÅ ą┐čĆąĄą┤čłąĄčüčéą▓čāąĄčé ąĖ┬ĀčüąŠą┐čĆąŠą▓ąŠąČą┤ą░ąĄčé čŹč鹊 ą┐ąŠą▓ąĄą┤ąĄąĮąĖąĄ, ą┐ąŠąĘą▓ąŠą╗čÅčé ąŠčåąĄąĮąĖčéčī čüąŠčüč鹊čÅąĮąĖąĄ čüčāą▒čŖąĄą║čéą░ ąĖ┬ĀąĖąĘą╝ąĄąĮąĄąĮąĖčÅ čāčĆąŠą▓ąĮčÅ čüąŠąĘąĮą░ąĮąĖčÅ ą▓┬ĀčŹčéąĖčģ ąŠą▒čüč鹊čÅč鹥ą╗čīčüčéą▓ą░čģ┬Ā[12]. ąĀą░ąĮąĄąĄ ąĮą░ą╝ąĖ ą▒čŗą╗ąĖ ą┐čĆąĖą▓ąĄą┤ąĄąĮčŗ ą┤ąŠą║ą░ąĘą░č鹥ą╗čīčüčéą▓ą░, ą║ąŠč鹊čĆčŗąĄ ą┐ąŠąĘą▓ąŠą╗čÅčÄčé ą┐čĆąĄą┤ą┐ąŠą╗ąŠąČąĖčéčī, čćč鹊 ą┤ą░ąČąĄ ą┐čĆąĖ ąĮąĄą▒ąŠą╗čīčłąŠą╝ čćąĖčüą╗ąĄ ąĮą░ąČą░čéąĖą╣ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮą░čÅ ą░ą║čéąĖą▓ąĮąŠčüčéčī ą▓┬ĀčŹč鹊čé ą┐ąĄčĆąĖąŠą┤ ąĮąĄ┬ĀčÅą▓ą╗čÅąĄčéčüčÅ ąĮąĄąŠčüąŠąĘąĮą░ą▓ą░ąĄą╝ąŠą╣, ą░┬ĀčüąŠą┐čĆąŠą▓ąŠąČą┤ą░ąĄčéčüčÅ čüąĮąĖąČąĄąĮąĮčŗą╝ čāčĆąŠą▓ąĮąĄą╝ čüąŠąĘąĮą░ąĮąĖčÅ [13]. ą£čŗ čĆąĄčłąĖą╗ąĖ ą┐čĆąŠą┤ąŠą╗ąČąĖčéčī ąĮą░čłčā čĆą░ą▒ąŠčéčā: ąĖčüčüą╗ąĄą┤ąŠą▓ą░čéčī ąŁąŁąō čüčāą▒čŖąĄą║č鹊ą▓ ą▓ąŠ ą▓čĆąĄą╝čÅ ą╝ąĖą║čĆąŠą┐čĆąŠą▒čāąČą┤ąĄąĮąĖą╣, ą║ąŠč鹊čĆčŗąĄ čüąŠą┐čĆąŠą▓ąŠąČą┤ą░čÄčéčüčÅ ąĄą┤ąĖąĮąĖčćąĮčŗą╝ąĖ ąĮą░ąČą░čéąĖčÅą╝ąĖ, ąĖ┬ĀčüąŠą┐ąŠčüčéą░ą▓ąĖčéčī ąĄąĄ čü┬ĀąŁąŁąō č鹥čģ ąČąĄ čüčāą▒čŖąĄą║č鹊ą▓ ą▓┬ĀčüąĖčéčāą░čåąĖąĖ čü┬Āą║ąŠčĆąŠčéą║ąĖą╝ ą┐ą░čéč鹥čĆąĮąŠą╝ ąĮą░ąČą░čéąĖą╣ (2ŌĆō6).
ą£ą░č鹥čĆąĖą░ą╗ ąĖ┬Āą╝ąĄč鹊ą┤čŗ
ąśčüčüą╗ąĄą┤ąŠą▓ą░ą╗ąĖ 19 ą▓ąĘčĆąŠčüą╗čŗčģ ąĘą┤ąŠčĆąŠą▓čŗčģ ąĖčüą┐čŗčéčāąĄą╝čŗčģ (čéčĆąŠąĄ ą╝čāąČčćąĖąĮ ąĖ┬Ā16 ąČąĄąĮčēąĖąĮ, čüčéčāą┤ąĄąĮčéčŗ, ą┐čĆą░ą▓čłąĖ, čüčĆąĄą┤ąĮąĖą╣ ą▓ąŠąĘčĆą░čüčé┬ĀŌĆō 21,26 ┬▒ 0,89┬Āą│ąŠą┤ą░) ą▒ąĄąĘ ą┤ąĖą░ą│ąĮąŠčüčéąĖčĆąŠą▓ą░ąĮąĮčŗčģ ąĮąĄą▓čĆąŠą╗ąŠą│ąĖč湥čüą║ąĖčģ, ą┐čüąĖčģąĖą░čéčĆąĖč湥čüą║ąĖčģ ąĘą░ą▒ąŠą╗ąĄą▓ą░ąĮąĖą╣ ąĖ┬ĀąĮą░čĆčāčłąĄąĮąĖą╣ čüąĮą░. ąÆčüąĄčģ ąĖčüą┐čŗčéčāąĄą╝čŗčģ ą┐čĆąŠčüąĖą╗ąĖ ąĮąĄ┬Āčāą┐ąŠčéčĆąĄą▒ą╗čÅčéčī ą░ą╗ą║ąŠą│ąŠą╗čī ą▓┬Āč鹥č湥ąĮąĖąĄ čüčāč鹊ą║, ą░┬Āčéą░ą║ąČąĄ čćą░ą╣ ąĖ┬Āą║ąŠč乥 ą▓┬Āč鹥č湥ąĮąĖąĄ čłąĄčüčéąĖ čćą░čüąŠą▓ ą┐ąĄčĆąĄą┤ ąŠą┐čŗč鹊ą╝. ąśčüčüą╗ąĄą┤ąŠą▓ą░ąĮąĖąĄ ą▒čŗą╗ąŠ ąŠą┤ąŠą▒čĆąĄąĮąŠ čŹčéąĖč湥čüą║ąŠą╣ ą║ąŠą╝ąĖčüčüąĖąĄą╣ ąśąĮčüčéąĖčéčāčéą░ ą▓čŗčüčłąĄą╣ ąĮąĄčĆą▓ąĮąŠą╣ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ ąĖ┬ĀąĮąĄą╣čĆąŠčäąĖąĘąĖąŠą╗ąŠą│ąĖąĖ ąĀąÉąØ (ą┐čĆąŠč鹊ą║ąŠą╗ Ōä¢┬Ā2 ąŠčé┬Ā3 ąĖčÄąĮčÅ 2019┬Āą│.) ąĖ┬ĀčüąŠąŠčéą▓ąĄčéčüčéą▓ąŠą▓ą░ą╗ąŠ čŹčéąĖč湥čüą║ąĖą╝ ąĮąŠčĆą╝ą░ą╝ ąźąĄą╗čīčüąĖąĮą║čüą║ąŠą╣ ą┤ąĄą║ą╗ą░čĆą░čåąĖąĖ ąÆčüąĄą╝ąĖčĆąĮąŠą╣ ą╝ąĄą┤ąĖčåąĖąĮčüą║ąŠą╣ ą░čüčüąŠčåąĖą░čåąĖąĖ ┬½ąŁčéąĖč湥čüą║ąĖąĄ ą┐čĆąĖąĮčåąĖą┐čŗ ą┐čĆąŠą▓ąĄą┤ąĄąĮąĖčÅ ąĮą░čāčćąĮčŗčģ ą╝ąĄą┤ąĖčåąĖąĮčüą║ąĖčģ ąĖčüčüą╗ąĄą┤ąŠą▓ą░ąĮąĖą╣ čü┬Āčāčćą░čüčéąĖąĄą╝ č湥ą╗ąŠą▓ąĄą║ą░┬╗ čü┬Āą┐ąŠą┐čĆą░ą▓ą║ą░ą╝ąĖ 2000┬Āą│.┬ĀąĪąŠčüč鹊čÅąĮąĖąĄ ą┐ąĄčĆąĄą┤ ąŠą┐čŗč鹊ą╝ ąŠčåąĄąĮąĖą▓ą░ą╗ąŠčüčī čü┬Āą┐ąŠą╝ąŠčēčīčÄ ą┤ąĮąĄą▓ąĮąĖą║ą░ čüąĮą░ [14] ąĖ┬ĀąŠą┐čĆąŠčüąĮąĖą║ą░ čāčĆąŠą▓ąĮčÅ čüąŠąĮą╗ąĖą▓ąŠčüčéąĖ KSS. ąĪčāą▒čŖąĄą║čéčŗ čüą┐ą░ą╗ąĖ ą▓┬ĀąĮąŠčćčī ą┐ąĄčĆąĄą┤ čüąĮąŠą╝ ą▓┬ĀčüčĆąĄą┤ąĮąĄą╝ ą£ = 6,46 ┬▒ 0,46 čćą░čüą░. ąÜą░č湥čüčéą▓ąŠ čüąĮą░ ąĖ┬Āčüą░ą╝ąŠčćčāą▓čüčéą▓ąĖąĄ ą┐čĆąĖ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖąĖ ą┐ąŠ┬Āą┐čÅčéąĖą▒ą░ą╗ą╗čīąĮąŠą╣ čłą║ą░ą╗ąĄ (5┬ĀŌĆō ┬½ąŠčéą╗ąĖčćąĮąŠąĄ┬╗): ą£ = 4,0 ┬▒ 0,19 ąĖ┬Āą£ = 3,32 ┬▒ 0,27 čüąŠąŠčéą▓ąĄčéčüčéą▓ąĄąĮąĮąŠ. ą×čéą╝ąĄčéąĖą╝ ą▓čŗčüąŠą║ąĖą╣ čāčĆąŠą▓ąĄąĮčī čüąŠąĮą╗ąĖą▓ąŠčüčéąĖ: ą£ = 5,42 ┬▒ 0,41 (ą┐ąŠ┬ĀčüąĄą╝ąĖą▒ą░ą╗ą╗čīąĮąŠą╣ čłą║ą░ą╗ąĄ, 7┬ĀŌĆō ┬½ąŠč湥ąĮčī ą▓čŗčüąŠą║ąĖą╣┬╗).
ąŁą║čüą┐ąĄčĆąĖą╝ąĄąĮčé ąĮą░čćąĖąĮą░ą╗ąĖ ą▓┬Āą┐ąĄčĆąĖąŠą┤ 12:00ŌĆō13:00 čćą░čüąŠą▓, ą┤ą╗ąĖč鹥ą╗čīąĮąŠčüčéčī ąĄą│ąŠ┬ĀŌĆō 1,5ŌĆō2 čćą░čüą░. ąÆ┬Āč鹥č湥ąĮąĖąĄ ą▓čüąĄą│ąŠ ąŠą┐čŗčéą░ ąĖčüą┐čŗčéčāąĄą╝čŗą╣ ąĮą░čģąŠą┤ąĖą╗čüčÅ ą▓┬ĀąĘą░č鹥ą╝ąĮąĄąĮąĮąŠą╝ ąĘą▓čāą║ąŠąĘą░ą│ą╗čāčłą░čÄčēąĄą╝ ą┐ąŠą╝ąĄčēąĄąĮąĖąĖ, ąĮą░┬Āą║čāčłąĄčéą║ąĄ, čü┬ĀąĘą░ą║čĆčŗčéčŗą╝ąĖ┬Āą│ą╗ą░ąĘą░ą╝ąĖ. ąĪ┬Āą┐ąŠą╝ąŠčēčīčÄ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░ [12] ą▓┬ĀąĄą│ąŠ čāąĮąĖą╝ą░ąĮčāą░ą╗čīąĮąŠą╣ ą╝ąŠą┤ąĖčäąĖą║ą░čåąĖąĖ ą┐ąŠą╗čāčćą░ą╗ąĖ čĆčÅą┤ ą┐ąŠčüą╗ąĄą┤ąŠą▓ą░č鹥ą╗čīąĮčŗčģ ą┐ąĄčĆąĖąŠą┤ąŠą▓ ąĘą░čüčŗą┐ą░ąĮąĖčÅ┬ĀŌĆō ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ. ąóąĄčüčé čüąŠčüč鹊ąĖčé ąĖąĘ┬Āą┤ą▓čāčģ č湥čĆąĄą┤čāčÄčēąĖčģčüčÅ čäą░ąĘ. ąśčüą┐čŗčéčāąĄą╝čŗąĄ čüčćąĖčéą░ą╗ąĖ ą┐čĆąŠ čüąĄą▒čÅ ąŠčé┬Ā1┬Āą┤ąŠ 10, ąŠą┤ąĮąŠą▓čĆąĄą╝ąĄąĮąĮąŠ ą┐čĆąĖ ą║ą░ąČą┤ąŠą╝ ąŠčéčüč湥č鹥 ąŠąĮąĖ ąĮą░ąČąĖą╝ą░ą╗ąĖ ąĮą░┬Āą║ąĮąŠą┐ą║čā ą▒ąŠą╗čīčłąĖą╝ ą┐ą░ą╗čīčåąĄą╝ ą┐čĆą░ą▓ąŠą╣ čĆčāą║ąĖ, ąĘą░čäąĖą║čüąĖčĆąŠą▓ą░ąĮąĮčāčÄ ąĮą░┬Āčāą║ą░ąĘą░č鹥ą╗čīąĮąŠą╝ ą┐ą░ą╗čīčåąĄ č鹊ą╣ ąČąĄ čĆčāą║ąĖ (ą┐ąĄčĆą▓ą░čÅ čäą░ąĘą░). ąöą░ą╗ąĄąĄ ąŠąĮąĖ ą┐čĆąŠą┤ąŠą╗ąČą░ą╗ąĖ čüčćąĖčéą░čéčī ą┐čĆąŠ čüąĄą▒čÅ ąŠčé┬Ā1 ą┤ąŠ 10, ąĮąŠ┬ĀčāąČąĄ ą▒ąĄąĘ ąĮą░ąČą░čéąĖą╣ (ą▓č鹊čĆą░čÅ čäą░ąĘą░). ą¦ąĄčĆąĄą┤ąŠą▓ą░ąĮąĖąĄ čŹčéąĖčģ čäą░ąĘ ą┤ą╗ąĖą╗ąŠčüčī ą┤ąŠ č鹥čģ ą┐ąŠčĆ, ą┐ąŠą║ą░ ąĖčüą┐čŗčéčāąĄą╝čŗąĄ ąĮąĄ┬ĀąĘą░čüčŗą┐ą░ą╗ąĖ. ą¤čĆąĖ čüą░ą╝ąŠą┐čĆąŠąĖąĘą▓ąŠą╗čīąĮčŗčģ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅčģ ąŠąĮąĖ ą▓ąŠąĘąŠą▒ąĮąŠą▓ą╗čÅą╗ąĖ ą▓čŗą┐ąŠą╗ąĮąĄąĮąĖąĄ č鹥čüčéą░ čü┬Āą┐ąĄčĆą▓ąŠą╣ čäą░ąĘčŗ.
ąĀąĄą│ąĖčüčéčĆąĖčĆąŠą▓ą░ą╗ąĖ ąŁąŁąō ąŠčé┬Ā17 ąŠčéą▓ąĄą┤ąĄąĮąĖą╣ ą▓┬ĀčüąŠąŠčéą▓ąĄčéčüčéą▓ąĖąĖ čüąŠ┬ĀčüčģąĄą╝ąŠą╣ 10ŌĆō20, 菹╗ąĄą║čéčĆąŠąŠą║čāą╗ąŠą│čĆą░ą╝ą╝čā, 菹╗ąĄą║čéčĆąŠą╝ąĖąŠą│čĆą░ą╝ą╝čā ą║ąŠčĆąŠčéą║ąŠą╣ ą╝čŗčłčåčŗ, ąŠčéą▓ąŠą┤čÅčēąĄą╣ ą▒ąŠą╗čīčłąŠą╣ ą┐ą░ą╗ąĄčå ą║ąĖčüčéąĖ ą┐čĆą░ą▓ąŠą╣ čĆčāą║ąĖ, ąĖ┬Āą╝ąĄčģą░ąĮąŠą│čĆą░ą╝ą╝čā ąĮą░ąČą░čéąĖą╣ ąĮą░┬Āą║ąĮąŠą┐ą║čā. ąĀąĄą│ąĖčüčéčĆą░čåąĖčÄ ą┐čĆąŠą▓ąŠą┤ąĖą╗ąĖ čü┬Āą┐ąŠą╝ąŠčēčīčÄ čüąĖčüč鹥ą╝čŗ Neocortex-Pro (Neurobotics). ą¦ą░čüč鹊čéą░ ą┤ąĖčüą║čĆąĄčéąĖąĘą░čåąĖąĖ┬ĀŌĆō 250 ąōčå, ą┐ąŠą╗ąŠčüą░ ą┐čĆąŠą┐čāčüą║ą░ąĮąĖčÅ čćą░čüč鹊čé: 0,5ŌĆō70 ąōčå. ąśčüą┐ąŠą╗čīąĘąŠą▓ą░ą╗ąĖ čłą╗ąĄą╝ ąĖ┬Āčģą╗ąŠčĆčüąĄčĆąĄą▒čĆčÅąĮčŗąĄ 菹╗ąĄą║čéčĆąŠą┤čŗ, čüąŠą┐čĆąŠčéąĖą▓ą╗ąĄąĮąĖąĄ┬ĀŌĆō ą╝ąĄąĮčīčłąĄ 5 ą║ą×ą╝.
ąöą╗čÅ ą░ąĮą░ą╗ąĖąĘą░ ą▓čŗą▒ąĖčĆą░ą╗ąĖ ą▒ąĄąĘą░čĆč鹥čäą░ą║čéąĮčŗąĄ ąŠčéčĆąĄąĘą║ąĖ ą┐ąŠą╗ąĖčüąŠą╝ąĮąŠą│čĆą░ą╝ą╝, čüąŠąŠčéą▓ąĄčéčüčéą▓čāčÄčēąĖąĄ 菹┐ąĖąĘąŠą┤ą░ą╝ čüą░ą╝ąŠą┐čĆąŠąĖąĘą▓ąŠą╗čīąĮąŠą│ąŠ ą║ąŠčĆąŠčéą║ąŠą│ąŠ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ ąĖąĘ┬Āą▓č鹊čĆąŠą╣ čüčéą░ą┤ąĖąĖ ąŠčĆč鹊ą┤ąŠą║čüą░ą╗čīąĮąŠą│ąŠ čüąĮą░ (N2) čü┬Āčćą░čüčéąĖčćąĮčŗą╝ ą▓čŗą┐ąŠą╗ąĮąĄąĮąĖąĄą╝ ą┐ąĄčĆą▓ąŠą╣ čäą░ąĘčŗ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░.┬Ā ąöą╗ąĖč鹥ą╗čīąĮąŠčüčéčī N2 ą┤ąŠą╗ąČąĮą░ ą▒čŗą╗ą░ ą▒čŗčéčī ąĮąĄ┬Āą╝ąĄąĮąĄąĄ 1,5┬Āą╝ąĖąĮčāčéčŗ, ąŠčåąĄąĮą║ą░ ą┐čĆąĖąĮą░ą┤ą╗ąĄąČąĮąŠčüčéąĖ ą┐ąŠą╗ąĖčüąŠą╝ąĮąŠą│čĆą░ą╝ą╝ ą║┬ĀN2 ą┐čĆąŠą▓ąŠą┤ąĖą╗ą░čüčī ą▓ąĖąĘčāą░ą╗čīąĮąŠ ą┐ąŠ┬Āą║čĆąĖč鹥čĆąĖčÅą╝ AASM (2023┬Āą│.). ąŻ┬Āą║ą░ąČą┤ąŠą│ąŠ ąĖčüą┐čŗčéčāąĄą╝ąŠą│ąŠ ą▓čŗą▒ąĖčĆą░ą╗ąĖ ą┐ąŠ┬Āą┤ą▓ą░ ąŠčéčĆąĄąĘą║ą░ ą┐ąŠą╗ąĖčüąŠą╝ąĮąŠą│čĆą░ą╝ą╝┬ĀŌĆō čü┬ĀąĄą┤ąĖąĮąĖčćąĮčŗą╝ ąĮą░ąČą░čéąĖąĄą╝ ąĖ┬Āčü ą┐ą░čéč鹥čĆąĮąŠą╝ (2ŌĆō6 ąĮą░ąČą░čéąĖą╣). ąĢčüą╗ąĖ ąŠčéčĆąĄąĘą║ąŠą▓ čü┬Āč鹥ą╝ ąĖą╗ąĖ ąĖąĮčŗą╝ čćąĖčüą╗ąŠą╝ ąĮą░ąČą░čéąĖą╣ ą▒čŗą╗ąŠ ąĮąĄčüą║ąŠą╗čīą║ąŠ, č鹊 ą┐čĆąĄą┤ą┐ąŠčćč鹥ąĮąĖąĄ ąŠčéą┤ą░ą▓ą░ą╗ąŠčüčī ą┐ą░čĆą░ą╝, ąĮą░ąĖą▒ąŠą╗ąĄąĄ ą▒ą╗ąĖąĘą║ąĖą╝ ą┐ąŠ┬Āą▓čĆąĄą╝ąĄąĮąĖ. ą¤čĆąĖą╝ąĄčĆ ą┐ąŠą╗ąĖčüąŠą╝ąĮąŠą│čĆą░ą╝ą╝čŗ čü┬Āą┐čĆąŠą▒čāąČą┤ąĄąĮąĖąĄą╝ ąĖ┬Āčćą░čüčéąĖčćąĮčŗą╝ ą▓čŗą┐ąŠą╗ąĮąĄąĮąĖąĄą╝ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░ ą┐čĆąĖą▓ąĄą┤ąĄąĮ ą▓┬ĀčĆą░ą▒ąŠč鹥 [13].
ąÉąĮą░ą╗ąĖąĘąĖčĆąŠą▓ą░ą╗ąĖ 40-čüąĄą║čāąĮą┤ąĮčŗąĄ ąŠčéčĆąĄąĘą║ąĖ ąĘą░ą┐ąĖčüąĖ ąŁąŁąō, ąĮą░┬ĀčüąĄčĆąĄą┤ąĖąĮčā ą║ąŠč鹊čĆčŗčģ ą┐čĆąĖčģąŠą┤ąĖą╗ąĖčüčī ąĮą░ąČą░čéąĖčÅ ąĮą░┬Āą║ąĮąŠą┐ą║čā: ą┐ąŠ┬Ā20 čüąĄą║čāąĮą┤ ą┤ąŠ ąĖ┬Āą┐ąŠčüą╗ąĄ ąĮą░čćą░ą╗ą░ ąĮą░ąČą░čéąĖą╣. ąöą╗čÅ ąŠčåąĄąĮą║ąĖ ą╝ąŠčēąĮąŠčüčéąĮčŗčģ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ ą▒ąĖąŠčŹą╗ąĄą║čéčĆąĖč湥čüą║ąŠą╣ ą░ą║čéąĖą▓ąĮąŠčüčéąĖ ą║ąŠčĆčŗ ą┐čĆąŠą▓ąŠą┤ąĖą╗ąĖ ąĮąĄą┐čĆąĄčĆčŗą▓ąĮąŠąĄ ą▓ąĄą╣ą▓ą╗ąĄčé-ą┐čĆąĄąŠą▒čĆą░ąĘąŠą▓ą░ąĮąĖąĄ ąĮą░┬ĀąŠčüąĮąŠą▓ąĄ ą╝ą░č鹥čĆąĖąĮčüą║ąŠą│ąŠ ą║ąŠą╝ą┐ą╗ąĄą║čüąĮąŠą│ąŠ Morlet-ą▓ąĄą╣ą▓ą╗ąĄčéą░ (Matlab 78.01). ąÜą░čĆčéčŗ čĆą░čüą┐čĆąĄą┤ąĄą╗ąĄąĮąĖčÅ ąĘąĮą░č湥ąĮąĖą╣ ą╝ąŠą┤čāą╗čÅ ą║ąŠčŹčäčäąĖčåąĖąĄąĮčéą░ ą▓ąĄą╣ą▓ą╗ąĄčé-ą┐čĆąĄąŠą▒čĆą░ąĘąŠą▓ą░ąĮąĖčÅ (ąÜąÆą¤) čüčéčĆąŠąĖą╗ąĖ ą▓┬Āą┐ąŠą╗ąŠčüąĄ 0,5ŌĆō40 ąōčå čü┬Āčłą░ą│ąŠą╝ 0,5 ąōčå ąĖ┬ĀčĆą░ąĘčĆąĄčłąĄąĮąĖąĄą╝ ą┐ąŠ┬Āą▓čĆąĄą╝ąĄąĮąĖ 0,01 čüąĄą║čāąĮą┤čŗ. ą¤čĆąŠą▓ąŠą┤ąĖą╗ąĖ čāčüčĆąĄą┤ąĮąĄąĮąĖąĄ ąÜąÆą¤ ą┐ąŠ┬Āčćą░čüč鹊č鹥 ą▓┬Āą┤ąĖą░ą┐ą░ąĘąŠąĮą░čģ 0,5ŌĆō3,5, 4ŌĆō7,5, 8ŌĆō10,5 ąĖ┬Ā11ŌĆō14,5, 15ŌĆō20,5 ąĖ┬Ā21ŌĆō40 ąōčå (ą┤ąĄą╗čīčéą░-, č鹥čéą░-, ąĮąĖąĘą║ąŠčćą░čüč鹊čéąĮčŗą╣ ąĖ┬Āą▓čŗčüąŠą║ąŠčćą░čüč鹊čéąĮčŗą╣ ą░ą╗čīčäą░-čĆąĖčéą╝čŗ, ą▒ąĄčéą░- ąĖ┬ĀąĮąĖąĘą║ąŠčćą░čüč鹊čéąĮčŗą╣┬Āą│ą░ą╝ą╝ą░-čĆąĖčéą╝), ą┤ą░ą╗ąĄąĄ čāčüčĆąĄą┤ąĮčÅą╗ąĖ ą┐ąŠ┬Āą▓čĆąĄą╝ąĄąĮąĖ, ą▓┬Āą┤ą▓čāčģčüąĄą║čāąĮą┤ąĮčŗčģ ąĖąĮč鹥čĆą▓ą░ą╗ą░čģ.
ą¤ąŠą╗čāč湥ąĮąĮčŗąĄ ą░ą╝ą┐ą╗ąĖčéčāą┤ąĮąŠ-ą╝ąŠčēąĮąŠčüčéąĮčŗąĄ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ąĖ ąŁąŁąō ą░ąĮą░ą╗ąĖąĘąĖčĆąŠą▓ą░ą╗ąĖ čü┬Āą┐ąŠą╝ąŠčēčīčÄ ą┤ąĖčüą┐ąĄčĆčüąĖąŠąĮąĮąŠą│ąŠ ą░ąĮą░ą╗ąĖąĘą░ (ANOVA RM). ąĀą░čüčüą╝ą░čéčĆąĖą▓ą░ą╗ąĖ ą▓ą╗ąĖčÅąĮąĖąĄ ąĮą░┬ĀąĮąĖčģ čäą░ą║č鹊čĆąŠą▓: ┬½čćąĖčüą╗ąŠ ąĮą░ąČą░čéąĖą╣┬╗┬ĀŌĆō ą┤ą▓ą░ čāčĆąŠą▓ąĮčÅ (ąĄą┤ąĖąĮąĖčćąĮąŠąĄ ąĮą░ąČą░čéąĖąĄ ąĖ┬Āą┐ą░čéč鹥čĆąĮ ąĮą░ąČą░čéąĖą╣), ┬½ą┐ąŠą▓ąĄą┤ąĄąĮč湥čüą║ą░čÅ ą░ą║čéąĖą▓ąĮąŠčüčéčī┬╗┬ĀŌĆō ą┤ą▓ą░ čāčĆąŠą▓ąĮčÅ (ą┐ąĄčĆąĖąŠą┤čŗ┬ĀŌĆō ą┤ąŠ ąĮą░čćą░ą╗ą░ ąĮą░ąČą░čéąĖą╣ ąĮą░┬Āą║ąĮąŠą┐ą║čā ąĖ┬Āą┐ąŠčüą╗ąĄ) ąĖ┬Ā┬½ą▓čĆąĄą╝čÅ┬╗ (10 čāčĆąŠą▓ąĮąĄą╣┬ĀŌĆō 10┬Āą┤ą▓čāčģčüąĄą║čāąĮą┤ąĮčŗčģ ąĖąĮč鹥čĆą▓ą░ą╗ąŠą▓ ą▓čĆąĄą╝ąĄąĮąĖ). ANOVA RM ą┐čĆąŠą▓ąŠą┤ąĖą╗ąĖ ą║ą░ą║ čüčāą╝ą╝ą░čĆąĮąŠ (č鹊 ąĄčüčéčī ą┐ąŠ┬ĀčāčüčĆąĄą┤ąĮąĄąĮąĮčŗą╝ ą╝ąŠčēąĮąŠčüčéąĮčŗą╝ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ą░ą╝ ą▓čüąĄčģ ąŠčéą▓ąĄą┤ąĄąĮąĖą╣ ąŁąŁąō ą┤ą╗čÅ ą║ą░ąČą┤ąŠą│ąŠ ąŠčéčüč湥čéą░), čéą░ą║ ąĖ┬Āą┐ąŠ ąŠčéą┤ąĄą╗čīąĮąŠčüčéąĖ ą┤ą╗čÅ ą║ą░ąČą┤ąŠą│ąŠ ąŠčéą▓ąĄą┤ąĄąĮąĖčÅ ąŁąŁąō. ąĪčéą░čéąĖčüčéąĖč湥čüą║ąĖąĄ ąŠčåąĄąĮą║ąĖ ą┐čĆąŠą▓ąŠą┤ąĖą╗ąĖ čü┬Āčāč湥č鹊ą╝ ą┐ąŠą┐čĆą░ą▓ą║ąĖ ąōčĆąĖąĮčģą░čāąĘą░┬ĀŌĆō ąōąĄčüčüąĄčĆą░. ąĪ┬Āą┐ąŠą╝ąŠčēčīčÄ ą┐ą░čĆąĮąŠą│ąŠ ą║čĆąĖč鹥čĆąĖčÅ ąĪčéčīčÄą┤ąĄąĮčéą░ ą┐ąŠą╗čāč湥ąĮąĮčŗąĄ ą▓┬ĀčĆąĄąĘčāą╗čīčéą░č鹥 čāčüčĆąĄą┤ąĮąĄąĮąĖą╣ ą┐ąŠ┬Āą▓čüąĄą╝ ąŠčéą▓ąĄą┤ąĄąĮąĖčÅą╝ ąŁąŁąō ą░ą╝ą┐ą╗ąĖčéčāą┤ąĮąŠ-ą╝ąŠčēąĮąŠčüčéąĮčŗąĄ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ąĖ čüčĆą░ą▓ąĮąĖą▓ą░ą╗ąĖ ą╝ąĄąČą┤čā čüąĖčéčāą░čåąĖčÅą╝ąĖ čü┬ĀąĄą┤ąĖąĮąĖčćąĮčŗą╝ ąĮą░ąČą░čéąĖąĄą╝ ąĖ┬Āą┐ą░čéč鹥čĆąĮąŠą╝ ąĮą░ąČą░čéąĖą╣┬ĀŌĆō ą┐ąŠ┬Āą║ą░ąČą┤ąŠą╝čā ą▓čŗą┤ąĄą╗ąĄąĮąĮąŠą╝čā ą┤ą▓čāčģčüąĄą║čāąĮą┤ąĮąŠą╝čā ąĖąĮč鹥čĆą▓ą░ą╗čā ąĖ┬Āą▓ ą║ą░ąČą┤ąŠą╝ ąĖąĘ┬Āčüą┐ąĄą║čéčĆą░ą╗čīąĮčŗčģ ą┤ąĖą░ą┐ą░ąĘąŠąĮąŠą▓. ą¤ąŠ┬ĀčĆąĄąĘčāą╗čīčéą░čéą░ą╝ čŹč鹊ą│ąŠ čüčĆą░ą▓ąĮąĄąĮąĖčÅ ą▓čŗą┤ąĄą╗čÅą╗ąĖ ą▓čĆąĄą╝ąĄąĮąĮčŗąĄ ąĖąĮč鹥čĆą▓ą░ą╗čŗ čü┬ĀčĆą░ąĘą╗ąĖčćą░čÄčēąĖą╝ąĖčüčÅ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ą░ą╝ąĖ. ąÆąĮčāčéčĆąĖ čŹčéąĖčģ ąĖąĮč鹥čĆą▓ą░ą╗ąŠą▓ ą┤ą╗čÅ ą║ą░ąČą┤ąŠą│ąŠ ąŠčéą▓ąĄą┤ąĄąĮąĖčÅ ą┐čĆąŠą▓ąŠą┤ąĖą╗ąĖ ą░ąĮą░ą╗ąŠą│ąĖčćąĮąŠąĄ čüčĆą░ą▓ąĮąĄąĮąĖąĄ┬ĀŌĆō čéą░ą║ąČąĄ ą┐ąŠ┬Āą┐ą░čĆąĮąŠą╝čā ą║čĆąĖč鹥čĆąĖčÄ ąĪčéčīčÄą┤ąĄąĮčéą░. ąÆčüąĄ ą▓čŗčćąĖčüą╗ąĄąĮąĖčÅ ą┐čĆąŠą▓ąŠą┤ąĖą╗ąĖ čü┬Āą┐ąŠą╝ąŠčēčīčÄ ą┐ą░ą║ąĄčéą░ ą┐čĆąŠą│čĆą░ą╝ą╝ SPSS 13.0.
ąĀąĄąĘčāą╗čīčéą░čéčŗ
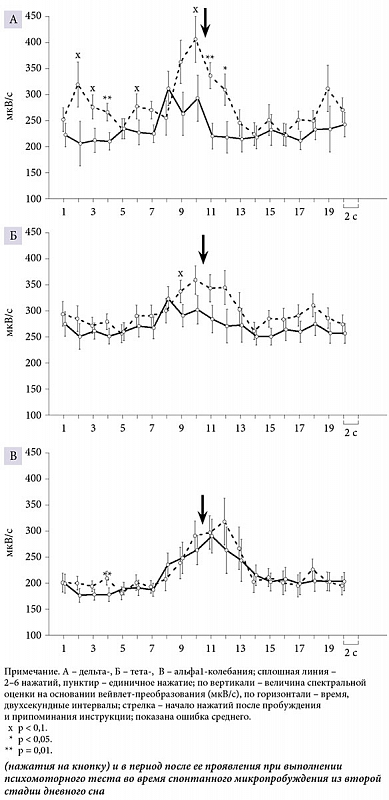
ą¤ąŠą║ą░ąĘą░ąĮąŠ ą▓ą╗ąĖčÅąĮąĖąĄ čäą░ą║č鹊čĆąŠą▓ ┬½čćąĖčüą╗ąŠ ąĮą░ąČą░čéąĖą╣┬╗ ąĖ┬Ā┬½ą▓čĆąĄą╝čÅ┬╗ ąĮą░┬Āčüčāą╝ą╝ą░čĆąĮčŗąĄ (čāčüčĆąĄą┤ąĮąĄąĮąĮčŗąĄ ą┐ąŠ┬Āą▓čüąĄą╝ ąŠčéą▓ąĄą┤ąĄąĮąĖčÅą╝) čüą┐ąĄą║čéčĆą░ą╗čīąĮčŗąĄ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ąĖ ąŁąŁąō ą┤ąĄą╗čīčéą░-ą║ąŠą╗ąĄą▒ą░ąĮąĖą╣ (F┬Ā(1; 18) = 8,01, p < 0,011 ąĖ┬ĀF (9; 10) = 3,13, p < 0,05 čüąŠąŠčéą▓ąĄčéčüčéą▓ąĄąĮąĮąŠ) ąĖ┬Āč鹥čéą░-čĆąĖčéą╝ą░ (F (1; 18) = 8,9, p < 0,01 ąĖ┬ĀF┬Ā(9;┬Ā10) = 3,89, p < 0,03 čüąŠąŠčéą▓ąĄčéčüčéą▓ąĄąĮąĮąŠ), ą░┬Āčéą░ą║ąČąĄ čäą░ą║č鹊čĆąŠą▓ ┬½ą┐ąŠą▓ąĄą┤ąĄąĮč湥čüą║ą░čÅ ą░ą║čéąĖą▓ąĮąŠčüčéčī┬╗ ąĖ┬Ā┬½ą▓čĆąĄą╝čÅ┬╗ ąĮą░┬Āą░ą╗čīčäą░1-čĆąĖčéą╝ (F (1; 18) = 5,63, p < 0,03 ąĖ┬ĀF┬Ā(9; 10) = 9,35, p < 0,001 čüąŠąŠčéą▓ąĄčéčüčéą▓ąĄąĮąĮąŠ).
ą£ąŠčēąĮąŠčüčéčī ą┤ąĄą╗čīčéą░- ąĖ┬Āč鹥čéą░-ą░ą║čéąĖą▓ąĮąŠčüčéąĖ ą▓┬ĀčåąĄą╗ąŠą╝ ą▒čŗą╗ą░ ą▒ąŠą╗čīčłąĄ ą▓ąŠ ą▓čĆąĄą╝čÅ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ ą┐čĆąĖ ąĄą┤ąĖąĮąĖčćąĮąŠą╝ ąĮą░ąČą░čéąĖąĖ, č湥ą╝ ą┐čĆąĖ čüąĄčĆąĖąĖ ąĮą░ąČą░čéąĖą╣. ąØąĖąĘą║ąŠčćą░čüč鹊čéąĮčŗą╣ ą░ą╗čīčäą░-čĆąĖčéą╝ ą▒čŗą╗ ą▒ąŠą╗čīčłąĄ ą┐ąŠčüą╗ąĄ ąĮą░čćą░ą╗ą░ ąĮą░ąČą░čéąĖą╣, č湥ą╝ ą▓┬Āą┐ąĄčĆąĖąŠą┤ ą┤ąŠ ąĮąĖčģ. ąŚąĮą░čćąĖą╝čŗąĄ čĆą░ąĘą╗ąĖčćąĖčÅ ą╝ąĄąČą┤čā čüąĖčéčāą░čåąĖčÅą╝ąĖ čü┬ĀąĄą┤ąĖąĮąĖčćąĮčŗą╝ ąĮą░ąČą░čéąĖąĄą╝ ąĖ┬ĀčüąĄčĆąĖąĄą╣ ąĮą░ąČą░čéąĖą╣ ą┐ąŠ┬Āą▓čĆąĄą╝ąĄąĮąĮčŗą╝ ąŠčéčĆąĄąĘą║ą░ą╝ ą▒čŗą╗ąĖ ą┐ąŠą║ą░ąĘą░ąĮčŗ ą┤ą╗čÅ ą┤ąĄą╗čīčéą░-ą║ąŠą╗ąĄą▒ą░ąĮąĖą╣ ąĖ┬Āą░ą╗čīčäą░1-čĆąĖčéą╝ą░ ąĘą░ 13ŌĆō14 čüąĄą║čāąĮą┤ ą┤ąŠ ąĮą░čćą░ą╗ą░ ąĮą░ąČą░čéąĖą╣ ąĖ┬Āą┤ą╗čÅ ą┤ąĄą╗čīčéą░-ą║ąŠą╗ąĄą▒ą░ąĮąĖą╣ ą▓┬Āą┐ąĄčĆą▓čŗąĄ č湥čéčŗčĆąĄ čüąĄą║čāąĮą┤čŗ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą╣ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ (čĆąĖčü.┬Ā1).
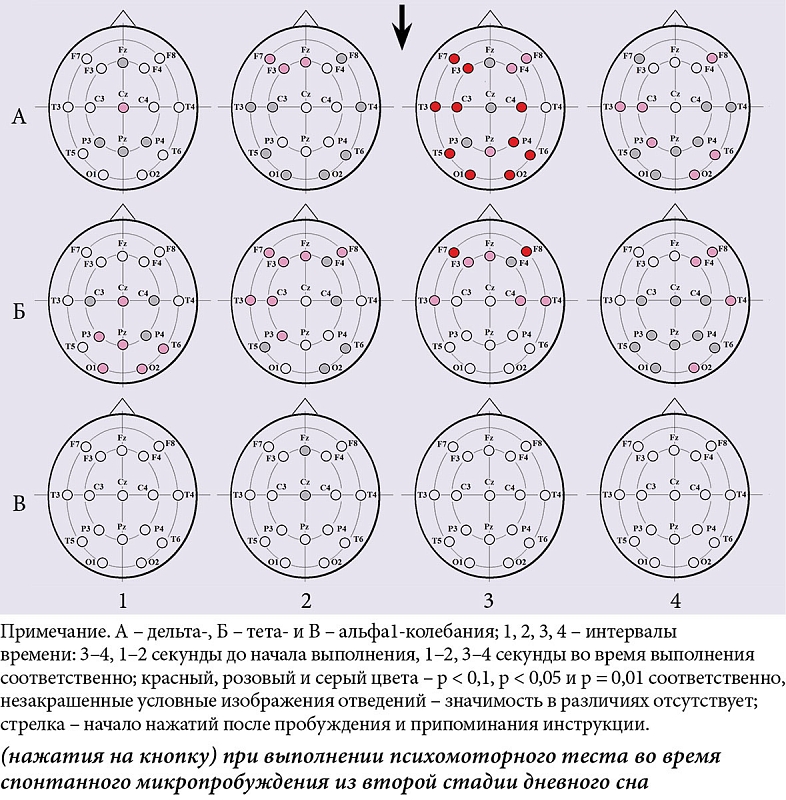
ą¤ąŠą║ą░ąĘą░ąĮąŠ┬Āą│ąĄąĮąĄčĆą░ą╗ąĖąĘąŠą▓ą░ąĮąĮąŠąĄ ą▓ą╗ąĖčÅąĮąĖąĄ ą┐ąŠ┬ĀąŠčéą▓ąĄą┤ąĄąĮąĖčÅą╝ ąŁąŁąō čäą░ą║č鹊čĆą░ ┬½čćąĖčüą╗ąŠ ąĮą░ąČą░čéąĖą╣┬╗ ąĮą░┬Āą╝ąŠčēąĮąŠčüčéčī ą┤ąĄą╗čīčéą░-ą║ąŠą╗ąĄą▒ą░ąĮąĖą╣ ąĖ┬Āč鹥čéą░-čĆąĖčéą╝ą░ (čĆąĖčü.┬Ā2ąÉ.1 ąĖ┬ĀčĆąĖčü. 2ąæ.1) ąĖ┬Āčäą░ą║č鹊čĆą░ ┬½ą┐ąŠą▓ąĄą┤ąĄąĮč湥čüą║ą░čÅ ą░ą║čéąĖą▓ąĮąŠčüčéčī┬╗ ąĮą░┬Āą╝ąŠčēąĮąŠčüčéčī ą░ą╗čīčäą░1-čĆąĖčéą╝ą░ ą▓┬Āą║ą░čāą┤ą░ą╗čīąĮčŗčģ ąŠą▒ą╗ą░čüčéčÅčģ ą║ąŠčĆčŗ (čĆąĖčü.┬Ā 2ąÆ.2).
ąÆ┬ĀąĖąĮč鹥čĆą▓ą░ą╗ąĄ 3ŌĆō4 čüąĄą║čāąĮą┤čŗ ą┤ąŠ ąĮą░čćą░ą╗ą░ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ ą╝ąĄąČą┤čā čüąĖčéčāą░čåąĖąĄą╣ čü┬ĀąĄą┤ąĖąĮąĖčćąĮčŗą╝ ąĮą░ąČą░čéąĖąĄą╝ ąĖ┬ĀčüąĖčéčāą░čåąĖąĄą╣ čü┬ĀčüąĄčĆąĖąĄą╣ ąĮą░ąČą░čéąĖą╣ ą┐ąŠą║ą░ąĘą░ąĮčŗ ąĘąĮą░čćąĖą╝čŗąĄ čĆą░ąĘą╗ąĖčćąĖčÅ ą▓┬Āą╝ąŠčēąĮąŠčüčéąĖ č鹥čéą░-čĆąĖčéą╝ą░ ą▓┬Āą║ą░čāą┤ą░ą╗čīąĮčŗčģ ąŠą▒ą╗ą░čüčéčÅčģ ą║ąŠčĆčŗ (čĆąĖčü.┬Ā3ąæ.1).┬Ā ąÆ┬ĀąĖąĮč鹥čĆą▓ą░ą╗ąĄ 1ŌĆō2 čüąĄą║čāąĮą┤čŗ, ąĮąĄą┐ąŠčüčĆąĄą┤čüčéą▓ąĄąĮąĮąŠ ą┐ąĄčĆąĄą┤ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéčīčÄ,┬ĀŌĆō čĆą░ąĘą╗ąĖčćąĖčÅ ą▓┬Āą╝ąŠčēąĮąŠčüčéąĖ ą┤ąĄą╗čīčéą░-ą║ąŠą╗ąĄą▒ą░ąĮąĖą╣ ą▓┬Āą╗ąĄą▓ąŠą╣ ą╗ąŠą▒ąĮąŠą╣ ąĖ┬Āą╗ąŠą▒ąĮąŠ-ą▓ąĖčüąŠčćąĮąŠą╣ ą║ąŠčĆąĄ (čĆąĖčü.┬Ā3ąÉ.2), ą▓┬Āą╝ąŠčēąĮąŠčüčéąĖ č鹥čéą░-čĆąĖčéą╝ą░ ą▓┬Āą╗ąŠą▒ąĮąŠą╣, ą╗ąŠą▒ąĮąŠ-ą▓ąĖčüąŠčćąĮąŠą╣, ą┐ąĄčĆąĄą┤ąĮąĄą▓ąĖčüąŠčćąĮąŠą╣, čåąĄąĮčéčĆą░ą╗čīąĮąŠą╣ ąĖ┬Āč鹥ą╝ąĄąĮąĮąŠą╣ ąŠą▒ą╗ą░čüčéčÅčģ ą╗ąĄą▓ąŠą│ąŠ ąĖ┬Āą╗ąŠą▒ąĮąŠ-ą▓ąĖčüąŠčćąĮąŠą╣ ąŠą▒ą╗ą░čüčéąĖ ą┐čĆą░ą▓ąŠą│ąŠ ą┐ąŠą╗čāčłą░čĆąĖčÅ (čĆąĖčü.┬Ā3ąæ.2). ąÆ┬ĀąĖąĮč鹥čĆą▓ą░ą╗ąĄ 1ŌĆō2┬ĀčüąĄą║čāąĮą┤čŗ, ąĮąĄą┐ąŠčüčĆąĄą┤čüčéą▓ąĄąĮąĮąŠ ą┐ąŠčüą╗ąĄ ąĮą░čćą░ą╗ą░ ąĮą░ąČą░čéąĖą╣, ą┐ąŠą║ą░ąĘą░ąĮčŗ čĆą░ąĘą╗ąĖčćąĖčÅ ą▓┬Āą╝ąŠčēąĮąŠčüčéąĖ ą┤ąĄą╗čīčéą░-ą║ąŠą╗ąĄą▒ą░ąĮąĖą╣ ą┐čĆą░ą║čéąĖč湥čüą║ąĖ ą▓ąŠ ą▓čüąĄčģ ąŠčéą▓ąĄą┤ąĄąĮąĖčÅčģ ąŁąŁąō (čĆąĖčü.┬Ā3ąÉ.3) ąĖ┬Āą▓ ą╝ąŠčēąĮąŠčüčéąĖ č鹥čéą░-čĆąĖčéą╝ą░ ą▓┬Āą┐ąĄčĆąĄą┤ąĮąĖčģ ąĖ┬Āą▓ąĖčüąŠčćąĮčŗčģ ąŠą▒ą╗ą░čüčéčÅčģ ąŠą▒ąŠąĖčģ ą┐ąŠą╗čāčłą░čĆąĖą╣ (čĆąĖčü.┬Ā3ąæ.3). ąÆąŠ ą▓čüąĄčģ čüą╗čāčćą░čÅčģ ą▓ąĄą╗ąĖčćąĖąĮčŗ čüą┐ąĄą║čéčĆą░ą╗čīąĮčŗčģ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ ą▒čŗą╗ąĖ ą▒ąŠą╗čīčłąĄ ą▓┬ĀčüąĖčéčāą░čåąĖąĖ čü┬ĀąĄą┤ąĖąĮąĖčćąĮčŗą╝ąĖ ąĮą░ąČą░čéąĖčÅą╝ąĖ.
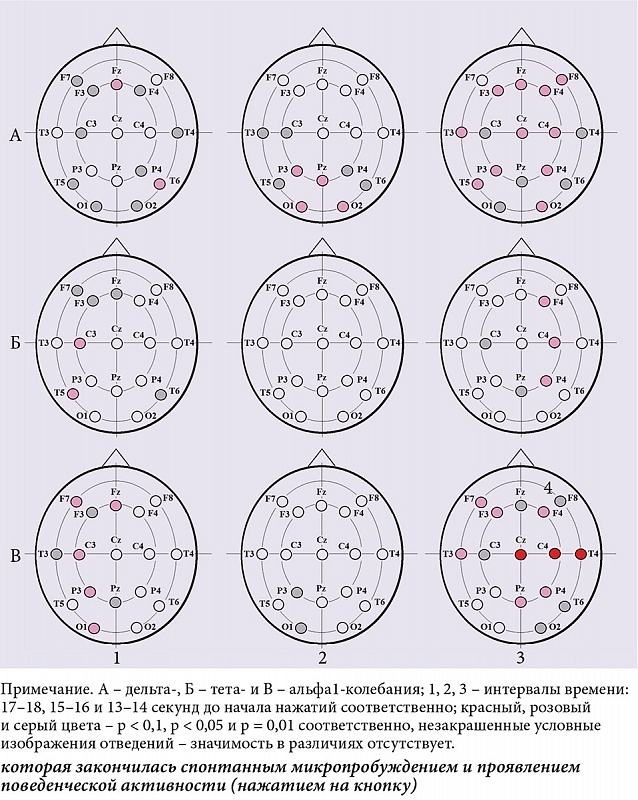
ąÆ┬ĀąĮą░čćą░ą╗ąĄ ą░ą║čéąĖą▓ą░čåąĖąĖ, ą║ąŠč鹊čĆą░čÅ ąĘą░ą║ąŠąĮčćąĖą╗ą░čüčī ą╝ąĖą║čĆąŠą┐čĆąŠą▒čāąČą┤ąĄąĮąĖąĄą╝ ąĖ┬Ā菹┐ąĖąĘąŠą┤ąŠą╝ ą┐ąŠą▓ąĄą┤ąĄąĮč湥čüą║ąŠą╣ ą░ą║čéąĖą▓ąĮąŠčüčéąĖ ą▓┬ĀąĖąĮč鹥čĆą▓ą░ą╗ąĄ 13ŌĆō14 čüąĄą║čāąĮą┤ ą┤ąŠ ąĮą░čćą░ą╗ą░ ąĮą░ąČą░čéąĖą╣ ą╝ąĄąČą┤čā čüąĖčéčāą░čåąĖąĄą╣ čü┬ĀąĄą┤ąĖąĮąĖčćąĮčŗą╝ ąĮą░ąČą░čéąĖąĄą╝ ąĖ┬ĀčüąĖčéčāą░čåąĖąĄą╣ čü┬ĀčüąĄčĆąĖąĄą╣ ąĮą░ąČą░čéąĖą╣, ą┐ąŠą║ą░ąĘą░ąĮčŗ ąĘąĮą░čćąĖą╝čŗąĄ čĆą░ąĘą╗ąĖčćąĖčÅ ą▓┬Āą╝ąŠčēąĮąŠčüčéąĖ ą┤ąĄą╗čīčéą░-ą║ąŠą╗ąĄą▒ą░ąĮąĖą╣ ą┐čĆą░ą║čéąĖč湥čüą║ąĖ ą▓ąŠ ą▓čüąĄą╣ ąŠą▒ą╗ą░čüčéąĖ ąŠčéą▓ąĄą┤ąĄąĮąĖčÅ ąŁąŁąō (čĆąĖčü.┬Ā4ąÉ.3) ąĖ┬Āą▓ ą╝ąŠčēąĮąŠčüčéąĖ ą░ą╗čīčäą░1-čĆąĖčéą╝ą░ ą▓┬Āą┐ąĄčĆąĄą┤ąĮąĖčģ ąŠą▒ą╗ą░čüčéčÅčģ ą╗ąĄą▓ąŠą│ąŠ ąĖ┬Āą│ąĄąĮąĄčĆą░ą╗ąĖąĘąŠą▓ą░ąĮąĮąŠ┬ĀŌĆō ą▓┬Āą┐čĆą░ą▓ąŠą╝ ą┐ąŠą╗čāčłą░čĆąĖąĖ (čĆąĖčü.┬Ā4ąÆ.3).
ą×ą▒čüčāąČą┤ąĄąĮąĖąĄ
ą¤ąĄčĆąĖąŠą┤čŗ ą░ą║čéąĖą▓ą░čåąĖąĖ, ą║ąŠč鹊čĆčŗąĄ ąĘą░ą║ą░ąĮčćąĖą▓ą░čÄčéčüčÅ ą╝ąĖą║čĆąŠą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅą╝ąĖ čü┬ĀąĄą┤ąĖąĮąĖčćąĮčŗą╝ ąĮą░ąČą░čéąĖąĄą╝ ąĮą░┬Āą║ąĮąŠą┐ą║čā, ąŠą▒čāčüą╗ąŠą▓ą╗ąĄąĮąĮčŗą╝ ąĮąĄąŠą▒čģąŠą┤ąĖą╝ąŠčüčéčīčÄ ą▓čŗą┐ąŠą╗ąĮąĄąĮąĖčÅ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░, čüąŠą┐čĆąŠą▓ąŠąČą┤ą░čÄčéčüčÅ ąŁąŁąō čü┬Āą▒ąŠą╗ąĄąĄ ą▓čŗčĆą░ąČąĄąĮąĮčŗą╝ąĖ ą┤ąĄą╗čīčéą░-ą║ąŠą╗ąĄą▒ą░ąĮąĖčÅą╝ąĖ ąĖ┬Āč鹥čéą░-čĆąĖčéą╝ąŠą╝ ą▒ąŠą╗čīčłąĄą╣ ą╝ąŠčēąĮąŠčüčéąĖ, č湥ą╝ ą▓┬ĀčüąĖčéčāą░čåąĖąĖ čü┬ĀčüąĄčĆąĖąĄą╣ ąĮą░ąČą░čéąĖą╣. ąŁč鹊čé čäą░ą║čé ą┐ąŠąĘą▓ąŠą╗čÅąĄčé ą┐čĆąĄą┤ą┐ąŠą╗ąŠąČąĖčéčī, čćč鹊 ą▒ąŠą╗ąĄąĄ ą║ąŠčĆąŠčéą║ąĖą╣ ą┐ąĄčĆąĖąŠą┤ ą╝ąŠč鹊čĆąĮąŠą╣ ą░ą║čéąĖą▓ąĮąŠčüčéąĖ ą┐čĆąŠąĖčüčģąŠą┤ąĖčé ą┐čĆąĖ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖąĖ ąĖąĘ┬Āą▒ąŠą╗ąĄąĄ ą▓čŗčĆą░ąČąĄąĮąĮąŠą╣ ą▓č鹊čĆąŠą╣ čüčéą░ą┤ąĖąĖ ąŠčĆč鹊ą┤ąŠą║čüą░ą╗čīąĮąŠą│ąŠ čüąĮą░. ąÆ┬ĀčŹč鹊ą╝ čüą╗čāčćą░ąĄ ą▒ąŠą╗ąĄąĄ ą▓čŗčüąŠą║ąĖąĄ ą░ą╝ą┐ą╗ąĖčéčāą┤čŗ ąÜ-ą║ąŠą╝ą┐ą╗ąĄą║čüąŠą▓, ą║ąŠč鹊čĆčŗąĄ ąĮąĄą┐ąŠčüčĆąĄą┤čüčéą▓ąĄąĮąĮąŠ ą┐čĆąĄą┤čłąĄčüčéą▓čāčÄčé ąĄą┤ąĖąĮąĖčćąĮąŠą╝čā ąĮą░ąČą░čéąĖčÄ, ą╝ąŠąČąĮąŠ čĆą░čüčüą╝ą░čéčĆąĖą▓ą░čéčī ą║ą░ą║ ą┐čĆąŠčÅą▓ą╗ąĄąĮąĖąĄ ą░ą║čéąĖą▓ą░čåąĖąŠąĮąĮčŗčģ ą┐čĆąŠčåąĄčüčüąŠą▓, čéčĆąĄą▒čāčÄčēąĖčģčüčÅ ą┤ą╗čÅ ąŠą║ąŠąĮčćą░č鹥ą╗čīąĮąŠą│ąŠ ą▓čŗą▓ąĄą┤ąĄąĮąĖčÅ čüčāą▒čŖąĄą║čéą░ ąĖąĘ┬Āą▒ąŠą╗ąĄąĄ┬Āą│ą╗čāą▒ąŠą║ąŠą│ąŠ čüąŠąĮąĮąŠą│ąŠ čüąŠčüč鹊čÅąĮąĖčÅ. ąĀąŠą╗čī ąÜ-ą║ąŠą╝ą┐ą╗ąĄą║čüąŠą▓ ą▓┬Āą░ą║čéąĖą▓ą░čåąĖčÅčģ ą╝ąŠąĘą│ą░, ą║ąŠč鹊čĆčŗąĄ ą┐čĆąĖą▓ąŠą┤čÅčé ą║┬Āą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÄ, ą┐ąŠą║ą░ąĘą░ąĮą░ ą▓┬ĀčĆą░ą▒ąŠčéą░čģ [8, 15]. ąóą░ą║ąČąĄ ą▒čŗą╗ąĖ ą┐ąŠą╗čāč湥ąĮčŗ ą┤ąŠą║ą░ąĘą░č鹥ą╗čīčüčéą▓ą░, čćč鹊 ą╝ąŠčēąĮąŠčüčéčī ąÜ-ą║ąŠą╝ą┐ą╗ąĄą║čüąŠą▓ ą┤ąŠą╗ąČąĮą░ ą┤ąŠčüčéąĖą│ą░čéčī ąŠą┐čĆąĄą┤ąĄą╗ąĄąĮąĮąŠą│ąŠ čāčĆąŠą▓ąĮčÅ, čćč鹊ą▒čŗ ąĘą░ą┐čāčüčéąĖčéčī čŹč鹊čé ą┐čĆąŠčåąĄčüčü ąĖ┬Āą┐ąŠą┤ą▓ąĄčüčéąĖ čüčāą▒čŖąĄą║čéą░ ą║┬Āą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ [16]. ąÆą╝ąĄčüč鹥 čü┬Āč鹥ą╝ ą▒ąŠą╗ąĄąĄ┬Āą│ą╗čāą▒ąŠą║ąŠąĄ čüąŠąĮąĮąŠąĄ čüąŠčüč鹊čÅąĮąĖąĄ, ą┐čĆąĄą┤čłąĄčüčéą▓čāčÄčēąĄąĄ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ, ą┐čĆąĄą┤ą┐ąŠą╗ą░ą│ą░ąĄčé ą▒ąŠą╗ąĄąĄ čüąĖą╗čīąĮąŠąĄ ą▓ą╗ąĖčÅąĮąĖąĄ ąĮą░┬ĀąĮąĄąĄ čéą░ą║ąŠą│ąŠ čÅą▓ą╗ąĄąĮąĖčÅ, ą║ą░ą║ ąĖąĮąĄčĆčåąĖčÅ čüąĮą░ [17]. ąØą░┬Āą▒ąŠą╗ąĄąĄ ą▓čŗčĆą░ąČąĄąĮąĮąŠąĄ ą▓ą╗ąĖčÅąĮąĖąĄ ąĖąĮąĄčĆčåąĖąĖ čüąĮą░, ą║ąŠč鹊čĆąŠąĄ, ą║ą░ą║ ą╝čŗ ą┐čĆąĄą┤ą┐ąŠą╗ą░ą│ą░ąĄą╝, čüąŠą║čĆą░čéąĖą╗ąŠ ą┐ąŠą▓ąĄą┤ąĄąĮąĖąĄ ą┤ąŠ ąĄą┤ąĖąĮąĖčćąĮąŠą│ąŠ ąĮą░ąČą░čéąĖčÅ ąĮą░┬Āą║ąĮąŠą┐ą║čā, čāą║ą░ąĘčŗą▓ą░čÄčé ą▒ąŠą╗ąĄąĄ ą▓čŗčüąŠą║ąĖąĄ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ąĖ ą┤ąĄą╗čīčéą░- ąĖ┬Āč鹥čéą░-ą░ą║čéąĖą▓ąĮąŠčüčéąĖ ą▓ąŠ ą▓čĆąĄą╝čÅ ąĖ┬Āą┐ąŠčüą╗ąĄ ąĄą┤ąĖąĮąĖčćąĮąŠą│ąŠ ąĮą░ąČą░čéąĖčÅ, č湥ą╝ ą▓┬ĀčüąĖčéčāą░čåąĖąĖ čüąĄčĆąĖąĖ ąĮą░ąČą░čéąĖą╣. ą×čéą╝ąĄčéąĖą╝, čćč鹊 ą▓┬Āą┤čĆčāą│ąĖčģ ąŠą▒čüč鹊čÅč鹥ą╗čīčüčéą▓ą░čģ, ą░┬ĀąĖą╝ąĄąĮąĮąŠ ąĮą░┬ĀąĮą░čćą░ą╗čīąĮąŠą╣ čüčéą░ą┤ąĖąĖ ąĖčüčüą╗ąĄą┤čāąĄą╝ąŠą╣ ąĮą░ą╝ąĖ ą┐čĆąŠą┤ąŠą╗ąČąĖč鹥ą╗čīąĮąŠą╣ ą░ą║čéąĖą▓ą░čåąĖąĖ ą╝ąŠąĘą│ą░ (ąĘą░ 13ŌĆō14 čüąĄą║čāąĮą┤ ą┤ąŠ ąĄą┤ąĖąĮąĖčćąĮąŠą│ąŠ ąĮą░ąČą░čéąĖčÅ), ą┐ąŠčÅą▓ą╗ąĄąĮąĖąĄ ąÜ-ą║ąŠą╝ą┐ą╗ąĄą║čüąŠą▓ čüčāčēąĄčüčéą▓ąĄąĮąĮąŠ ą╝ąĄąĮčīčłąĄą╣ ą░ą╝ą┐ą╗ąĖčéčāą┤čŗ ąĮąĄ┬Āą┐čĆąĖą▓ąŠą┤ąĖą╗ąŠ ą║┬Āą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÄ ąĖ┬Āą┤ą▓ąĖą│ą░č鹥ą╗čīąĮąŠą╣ ą░ą║čéąĖą▓ąĮąŠčüčéąĖ.
ąæąŠą╗ąĄąĄ ą▓čŗčĆą░ąČąĄąĮąĮą░čÅ ą╝ąŠčēąĮąŠčüčéčī č鹥čéą░-čĆąĖčéą╝ą░ ą┐čĆąĖ ąĄą┤ąĖąĮąĖčćąĮčŗčģ ąĮą░ąČą░čéąĖčÅčģ, ą║ą░ą║ ą╝čŗ čāąČąĄ čāą┐ąŠą╝ąĖąĮą░ą╗ąĖ, čüą▓čÅąĘčŗą▓ą░ąĄčéčüčÅ čü┬Āą┐čĆąŠčåąĄčüčüą░ą╝ąĖ ąĖąĮąĄčĆčåąĖąĖ čüąĮą░. ąÆą╝ąĄčüč鹥 čü┬Āč鹥ą╝ č鹥čéą░-čĆąĖčéą╝, ąŠčüąŠą▒ąĄąĮąĮąŠ ą▓ąŠ čäčĆąŠąĮčéą░ą╗čīąĮąŠą╣ ą║ąŠčĆąĄ, čüą▓čÅąĘčŗą▓ą░čÄčé čü┬Āą┐čĆąŠčåąĄčüčüą░ą╝ąĖ ą║ąŠą│ąĮąĖčéąĖą▓ąĮąŠą│ąŠ ą║ąŠąĮčéčĆąŠą╗čÅ [18]. ą¤čĆąĖ čŹč鹊ą╝ č鹥čéą░-čĆąĖčéą╝ čāą▓ąĄą╗ąĖčćąĖą▓ą░ąĄčéčüčÅ ąĮąĄ┬Āč鹊ą╗čīą║ąŠ ą▓ąŠ ą▓čĆąĄą╝čÅ ą▓čŗą┐ąŠą╗ąĮąĄąĮąĖčÅ ąĘą░ą┤ą░ąĮąĖą╣ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░, ąĮąŠ┬ĀąĖ ą▓┬Āą┐čĆąŠčåąĄčüčüąĄ ą┐ąŠą┤ą│ąŠč鹊ą▓ą║ąĖ ą║┬ĀąĮąĄą╝čā. ąśąĮą░č湥┬Āą│ąŠą▓ąŠčĆčÅ, čäčāąĮą║čåąĖčÅ ą║ąŠąĮčéčĆąŠą╗čÅ ą▓ą║ą╗čÄčćą░ąĄčéčüčÅ čĆą░ąĮčīčłąĄ ąĮą░čćą░ą╗ą░ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ ąĖ┬ĀąĮąŠčüąĖčé čāą┐čĆąĄąČą┤ą░čÄčēąĖą╣ čģą░čĆą░ą║č鹥čĆ. ąØą░ą▒ą╗čÄą┤ą░ąĄą╝čŗąĄ ąĮą░ą╝ąĖ ą▒├│ą╗čīčłąĖąĄ ą▓ąĄą╗ąĖčćąĖąĮčŗ ą╝ąŠčēąĮąŠčüčéąĖ č鹥čéą░-čĆąĖčéą╝ą░ ą▓┬ĀčüąĖčéčāą░čåąĖąĖ ąĄą┤ąĖąĮąĖčćąĮąŠą│ąŠ ąĮą░ąČą░čéąĖčÅ ą┐ąŠ┬ĀčüčĆą░ą▓ąĮąĄąĮąĖčÄ čü┬ĀčüąĄčĆąĖąĄą╣ ąĮą░ąČą░čéąĖą╣ ą▓ąŠ čäčĆąŠąĮčéą░ą╗čīąĮčŗčģ ąŠą▒ą╗ą░čüčéčÅčģ ąŠčéą▓ąĄą┤ąĄąĮąĖčÅ ąŁąŁąō ą╝ąŠąČąĮąŠ čĆą░čüčüą╝ą░čéčĆąĖą▓ą░čéčī ą║ą░ą║ čāčüąĖą╗ąĄąĮąĖąĄ čäčāąĮą║čåąĖąĖ ą║ąŠą│ąĮąĖčéąĖą▓ąĮąŠą│ąŠ ą║ąŠąĮčéčĆąŠą╗čÅ, ą║ąŠč鹊čĆčŗą╣ ąĮąĄąŠą▒čģąŠą┤ąĖą╝ ą┐čĆąĖ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą╣ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ ą▓┬Āčāčüą╗ąŠą▓ąĖčÅčģ ą┐čĆąĄąŠą┤ąŠą╗ąĄąĮąĖčÅ ą▒ąŠą╗ąĄąĄ ą▓čŗčĆą░ąČąĄąĮąĮąŠą│ąŠ ą▓ą╗ąĖčÅąĮąĖčÅ ąŠčĆč鹊ą┤ąŠą║čüą░ą╗čīąĮąŠą│ąŠ čüąĮą░.
ąŻčüąĖą╗ąĄąĮąĖąĄ ą░ą╗čīčäą░-čĆąĖčéą╝ą░ ą▓┬Āą┐ąĄčĆąĖąŠą┤ ą╝ąĖą║čĆąŠą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ ąĮąĄą┐ąŠčüčĆąĄą┤čüčéą▓ąĄąĮąĮąŠ ą┐ąĄčĆąĄą┤ ąĮą░čćą░ą╗ąŠą╝ ąĮą░ąČą░čéąĖą╣ ą╝čŗ čüą▓čÅąĘčŗą▓ą░ąĄą╝ ą▓┬ĀąĮą░čłąĖčģ ąĖčüčüą╗ąĄą┤ąŠą▓ą░ąĮąĖčÅčģ čü┬Āą┐čĆąĖą┐ąŠą╝ąĖąĮą░ąĮąĖąĄą╝ ąĖąĮčüčéčĆčāą║čåąĖąĖ ąĖ┬Āą┐ąŠą┤ą│ąŠč鹊ą▓ą║ąŠą╣ ą║┬Āą▓ąŠčüą┐čĆąŠąĖąĘą▓ąĄą┤ąĄąĮąĖčÄ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░ [13, 16]. ąŻą▓ąĄą╗ąĖč湥ąĮąĖąĄ ąĄą│ąŠ ą╝ąŠčēąĮąŠčüčéąĮčŗčģ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ ą▓┬Āą┐ąĄčĆąĖąŠą┤ ą┐ąŠčüą╗ąĄ ąĮą░čćą░ą╗ą░ ąĮą░ąČą░čéąĖą╣, ąŠčüąŠą▒ąĄąĮąĮąŠ ą▓┬Āč鹥ą╝ąĄąĮąĮąŠ-ąĘą░čéčŗą╗ąŠčćąĮčŗčģ ąŠą▒ą╗ą░čüčéčÅčģ, ąŠčéčĆą░ąČą░ąĄčé ą┐ąŠą┤ą┤ąĄčƹȹ░ąĮąĖąĄ ą░ą║čéąĖą▓ą░čåąĖąĖ ą┐čĆąĖ ą▓čŗą┐ąŠą╗ąĮąĄąĮąĖąĖ čŹč鹊ą╣ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą╣ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ, ą░┬Āą┤ą░ą╗ąĄąĄ, ą║ąŠą│ą┤ą░ ąĮą░ąČą░čéąĖčÅ ą┐čĆąĄą║čĆą░čēą░čÄčéčüčÅ, ą▓ąĄčĆąŠčÅčéąĮąŠ, ąĄąĄ čüą╗ąĄą┤ąŠą▓čŗąĄ ą┐čĆąŠčåąĄčüčüčŗ. ąóąĄą╝ąĄąĮąĮąŠ-ąĘą░čéčŗą╗ąŠčćąĮčŗą╣ ą░ą╗čīčäą░-čĆąĖčéą╝ ąĮąĄąŠą▒čģąŠą┤ąĖą╝ ą┤ą╗čÅ ą┐ąŠą┤ą┤ąĄčƹȹ░ąĮąĖčÅ čüąŠąĘąĮą░ąĮąĖčÅ [19]. ąØąĄąŠą▒čģąŠą┤ąĖą╝ąŠčüčéčī ą┐čĆąĖčüčāčéčüčéą▓ąĖčÅ čŹč鹊ą╣ čĆąĖčéą╝ąĖč湥čüą║ąŠą╣ čüąŠčüčéą░ą▓ą╗čÅčÄčēąĄą╣ ą▓┬ĀąŁąŁąō ą┤ą╗čÅ ąŠčüčāčēąĄčüčéą▓ą╗ąĄąĮąĖčÅ čüąŠąĘąĮą░č鹥ą╗čīąĮąŠą╣ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ ą┐čĆąĖ ą║ąŠčĆąŠčéą║ąĖčģ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅčģ ą┐ąŠą║ą░ąĘą░ąĮą░ ą▓┬ĀčĆą░ą▒ąŠč鹥 [11]. ąĀą░ąĮąĄąĄ ąĮą░ą╝ąĖ ąŠčéą╝ąĄčćą░ą╗ąĖčüčī čüčāčēąĄčüčéą▓ąĄąĮąĮą░čÅ čĆąŠą╗čī čŹč鹊ą│ąŠ čĆąĖčéą╝ą░ ą┐čĆąĖ ą▓čŗą┐ąŠą╗ąĮąĄąĮąĖąĖ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░ ąĖ┬ĀąĄą│ąŠ čüą▓čÅąĘčī čü┬ĀčāčĆąŠą▓ąĮąĄą╝ čüąŠąĘąĮą░ąĮąĖčÅ ą▓ąŠ ą▓čĆąĄą╝čÅ ą▒ąŠą╗ąĄąĄ ą┤ą╗ąĖč鹥ą╗čīąĮčŗčģ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖą╣ [13]. ąØą░┬ĀčŹč鹊ą╝ ąŠčüąĮąŠą▓ą░ąĮąĖąĖ ą╝ąŠąČąĮąŠ ą┐čĆąĄą┤ą┐ąŠą╗ąŠąČąĖčéčī, čćč鹊 ąĮąĄ┬Āč鹊ą╗čīą║ąŠ ą┐ą░čéč鹥čĆąĮ ąĖąĘ┬Ā2ŌĆō6 ąĮą░ąČą░čéąĖą╣, ąĮąŠ┬ĀąĖ ąĄą┤ąĖąĮąĖčćąĮčŗąĄ ąĮą░ąČą░čéąĖčÅ čÅą▓ą╗čÅčÄčéčüčÅ čüąŠąĘąĮą░č鹥ą╗čīąĮčŗą╝ąĖ ą░ą║čéą░ą╝ąĖ, ąŠą┤ąĮą░ą║ąŠ čāčĆąŠą▓ąĄąĮčī čüąŠąĘąĮą░ąĮąĖčÅ, ą▓ąĄčĆąŠčÅčéąĮąŠ, ąĘąĮą░čćąĖč鹥ą╗čīąĮąŠ čüąĮąĖąČąĄąĮ ą┐ąŠ┬ĀčüčĆą░ą▓ąĮąĄąĮąĖčÄ čü┬Āą▓čŗą┐ąŠą╗ąĮąĄąĮąĖąĄą╝ ą┤ą░ąĮąĮąŠą╣ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ ą▓┬ĀčüąŠčüč鹊čÅąĮąĖąĖ ą▒ąŠą┤čĆčüčéą▓ąŠą▓ą░ąĮąĖčÅ. ą¤čĆąĖ čŹč鹊ą╝ ąŠčéčüčāčéčüčéą▓ąĖąĄ čĆą░ąĘą╗ąĖčćąĖą╣ ą▓┬Āą╝ąŠčēąĮąŠčüčéąĮčŗčģ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ą░čģ ą░ą╗čīčäą░-čĆąĖčéą╝ą░, čüąŠą┐čĆąŠą▓ąŠąČą┤ą░čÄčēąĖčģ ąŠą▒čüčāąČą┤ą░ąĄą╝čāčÄ ą║čĆą░čéą║ąŠą▓čĆąĄą╝ąĄąĮąĮčāčÄ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéčī čĆą░ąĘąĮąŠą╣ ą┤ą╗ąĖč鹥ą╗čīąĮąŠčüčéąĖ, čüą▓ąĖą┤ąĄč鹥ą╗čīčüčéą▓čāąĄčé, čćč鹊 ą▓┬ĀąŠą▒ąŠąĖčģ čüą╗čāčćą░čÅčģ ą▓ąŠčüą┐čĆąŠąĖąĘą▓ąĄą┤ąĄąĮąĖąĄ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░ ą▓┬Āčāčüą╗ąŠą▓ąĖčÅčģ ą╝ąĖą║čĆąŠą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ ą┐čĆąŠąĖčüčģąŠą┤ąĖčé ąĮą░┬ĀčüčģąŠą┤ąĮąŠą╝ čāčĆąŠą▓ąĮąĄ.
ąŚą░ą║ą╗čÄč湥ąĮąĖąĄ
ąöą╗ąĖč鹥ą╗čīąĮąŠčüčéčī ą▓čŗą┐ąŠą╗ąĮąĄąĮąĖčÅ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░ ą▓┬Āčāčüą╗ąŠą▓ąĖčÅčģ ą║čĆą░čéą║ąŠą▓čĆąĄą╝ąĄąĮąĮąŠą│ąŠ ą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ ąĖąĘ┬Āą▓č鹊čĆąŠą╣ čüčéą░ą┤ąĖąĖ ąŠčĆč鹊ą┤ąŠą║čüą░ą╗čīąĮąŠą│ąŠ ą┤ąĮąĄą▓ąĮąŠą│ąŠ čüąĮą░ čüą▓čÅąĘą░ąĮą░ čü┬Āą│ą╗čāą▒ąĖąĮąŠą╣ čŹč鹊ą╣ čüčéą░ą┤ąĖąĖ, ą║ąŠč鹊čĆą░čÅ čģą░čĆą░ą║č鹥čĆąĖąĘčāąĄčéčüčÅ čĆą░ąĘąĮąŠą╣ ą▓ąĄą╗ąĖčćąĖąĮąŠą╣ ą╝ąŠčēąĮąŠčüčéąĮčŗčģ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ ą┤ąĄą╗čīčéą░-ą║ąŠą╗ąĄą▒ą░ąĮąĖą╣ ąĖ┬Āč鹥čéą░-čĆąĖčéą╝ą░. ą¤ąŠ┬ĀčüčĆą░ą▓ąĮąĄąĮąĖčÄ čü┬ĀąĮąĄčüą║ąŠą╗čīą║ąĖą╝ąĖ ąĮą░ąČą░čéąĖčÅą╝ąĖ ąĄą┤ąĖąĮąĖčćąĮčŗą╝ ąĮą░ąČą░čéąĖčÅą╝ ą┐čĆąĄą┤čłąĄčüčéą▓čāąĄčé ą┐ąĄčĆąĖąŠą┤ ą▒ąŠą╗ąĄąĄ┬Āą│ą╗čāą▒ąŠą║ąŠą│ąŠ čüąŠąĮąĮąŠą│ąŠ čüąŠčüč鹊čÅąĮąĖčÅ, ą║ąŠč鹊čĆčŗą╣ ąĘą░ą▓ąĄčĆčłą░ąĄčéčüčÅ ąÜ-ą║ąŠą╝ą┐ą╗ąĄą║čüą░ą╝ąĖ čü┬Āą▒ąŠą╗ąĄąĄ ą▓čŗčüąŠą║ąĖą╝ąĖ ą░ą╝ą┐ą╗ąĖčéčāą┤ą░ą╝ąĖ. ąÜ-ą║ąŠą╝ą┐ą╗ąĄą║čüčŗ ą▓┬ĀčŹčéąĖčģ ąŠą▒čüč鹊čÅč鹥ą╗čīčüčéą▓ą░čģ čĆą░čüčüą╝ą░čéčĆąĖą▓ą░čÄčéčüčÅ ą║ą░ą║ ą┐čĆąŠčÅą▓ą╗ąĄąĮąĖąĄ ą░ą║čéąĖą▓ą░čåąĖąŠąĮąĮčŗčģ ą┐čĆąŠčåąĄčüčüąŠą▓, čéčĆąĄą▒čāčÄčēąĖčģčüčÅ ą┤ą╗čÅ ą┐ąŠą┤ą│ąŠč鹊ą▓ą║ąĖ ą║┬Āą▓čŗą┐ąŠą╗ąĮąĄąĮąĖčÄ ąĮą░ąČą░čéąĖą╣ ąĮą░┬Āą║ąĮąŠą┐ą║čā. ą×čéčüčāčéčüčéą▓ąĖąĄ čĆą░ąĘą╗ąĖčćąĖą╣ ą▓┬Āą╝ąŠčēąĮąŠčüčéąĮčŗčģ čģą░čĆą░ą║č鹥čĆąĖčüčéąĖą║ą░čģ ą░ą╗čīčäą░-čĆąĖčéą╝ą░ ą┐čĆąĖ ą┤ąĄčÅč鹥ą╗čīąĮąŠčüčéąĖ čĆą░ąĘąĮąŠą╣ ą┤ą╗ąĖč鹥ą╗čīąĮąŠčüčéąĖ čüą▓ąĖą┤ąĄč鹥ą╗čīčüčéą▓čāąĄčé, čćč鹊 ą▓┬ĀąŠą▒ąŠąĖčģ čüą╗čāčćą░čÅčģ ą▓ąŠčüą┐čĆąŠąĖąĘą▓ąĄą┤ąĄąĮąĖąĄ ą┐čüąĖčģąŠą╝ąŠč鹊čĆąĮąŠą│ąŠ č鹥čüčéą░ ą▓┬Āčāčüą╗ąŠą▓ąĖčÅčģ ą╝ąĖą║čĆąŠą┐čĆąŠą▒čāąČą┤ąĄąĮąĖčÅ ą┐čĆąŠąĖčüčģąŠą┤ąĖčé ąĮą░┬ĀčüčģąŠą┤ąĮąŠą╝ čäčāąĮą║čåąĖąŠąĮą░ą╗čīąĮąŠą╝ ąĖ┬ĀčüąŠąĘąĮą░č鹥ą╗čīąĮąŠą╝ čāčĆąŠą▓ąĮąĄ. ą¤ąŠą╗čāč湥ąĮąĮčŗąĄ čĆąĄąĘčāą╗čīčéą░čéčŗ ą▒čāą┤čāčé ą┐ąŠą╗ąĄąĘąĮčŗ ą▓┬Āą║ą╗ąĖąĮąĖč湥čüą║ąŠą╣ ą┐čĆą░ą║čéąĖą║ąĄ ą┐čĆąĖ ąĖčüčüą╗ąĄą┤ąŠą▓ą░ąĮąĖąĖ čĆą░čüčüčéčĆąŠą╣čüčéą▓ ą▓ąŠąĘą▒čāąČą┤ąĄąĮąĖą╣ ą▓ąŠ ą▓čĆąĄą╝čÅ čüąĮą░, ą║ąŠč鹊čĆčŗąĄ ą┐čĆąĖą▓ąŠą┤čÅčé ą║┬ĀąĮąĄą┐čĆąŠąĖąĘą▓ąŠą╗čīąĮąŠą╣ ą┤ą▓ąĖą│ą░č鹥ą╗čīąĮąŠą╣ ą░ą║čéąĖą▓ąĮąŠčüčéąĖ čĆą░ąĘą╗ąĖčćąĮąŠą╣ ą┤ą╗ąĖč鹥ą╗čīąĮąŠčüčéąĖ ąĖ┬Āčüą╗ąŠąČąĮąŠčüčéąĖ.┬Ā
ąĀą░ą▒ąŠčéą░ ą▓čŗą┐ąŠą╗ąĮąĄąĮą░ ą▓┬ĀčĆą░ą╝ą║ą░čģ┬Āą│ąŠčüčāą┤ą░čĆčüčéą▓ąĄąĮąĮąŠą│ąŠ ąĘą░ą┤ą░ąĮąĖčÅ ą£ąĖąĮąĖčüč鹥čĆčüčéą▓ą░ ąŠą▒čĆą░ąĘąŠą▓ą░ąĮąĖčÅ ąĖ┬ĀąĮą░čāą║ąĖ ąĀąŠčüčüąĖą╣čüą║ąŠą╣ ążąĄą┤ąĄčĆą░čåąĖąĖ ąĮą░┬Ā2021ŌĆō2023┬Āą│ą│.
ąÜąŠąĮčäą╗ąĖą║čé ąĖąĮč鹥čĆąĄčüąŠą▓ ąŠčéčüčāčéčüčéą▓čāąĄčé.
E.A. Cheremushkin, PhD, N.E. Petrenko, V.B. Dorokhov
Institute of Higher Nervous Activity and Neurophysiology RAS
Contact person: Evgeny Al. Cheremushkin, khton@mail.ru
The recovery of different durations cognitive activity under the conditions of performing a psychomotor test during micro-awakening from stage 2 orthodox daytime sleep was investigated in 19 healthy young adults, students. We analyzed electroencephalogram (EEG) during the period of brain arousal that ended with an episode of awakening with different numbers of button presses, as well as the period of time that followed behavioral activity. The number of clicks was related to the depth of this sleep stage, which was characterized by different magnitudes of the power characteristics of delta oscillations and theta rhythm. Compared to the activity pattern (2ŌĆō6 taps), the single-tap situation is preceded by a period of deeper sleep state, which is completed by K-complexes with higher amplitudes. The K-complexes in these circumstances are considered to be a manifestation of the activation processes required to prepare for the execution of button presses. The higher power values of the theta rhythm in the situation of a single press compared to the pattern of presses in the frontal regions of the EEG can be considered as a strengthening of the function of cognitive control, which is necessary for psychomotor activity in conditions of overcoming the more pronounced influence of orthodox sleep. At the same time, the theta rhythm increases not only during the performance of psychomotor test tasks, but also in the process of preparation for it. In other words, the control function is switched on before the beginning of activity and has an anticipatory character. The absence of differences in the power characteristics of the alpha rhythm during the period of instruction recall and during activities of different duration indicates that in both cases the reproduction of the psychomotor test under conditions of micro-awakening occurs at a similar functional and conscious level. The results obtained will be useful in clinical practice in the study of disorders of arousal during sleep, which are accompanied by involuntary motor activity of varying duration and complexity.
ąŻą▓ą░ąČą░ąĄą╝čŗą╣ ą┐ąŠčüąĄčéąĖč鹥ą╗čī uMEDp!
ąŻą▓ąĄą┤ąŠą╝ą╗čÅąĄą╝ ąÆą░čü ąŠ č鹊ą╝, čćč鹊 ąĘą┤ąĄčüčī čüąŠą┤ąĄčƹȹĖčéčüčÅ ąĖąĮč乊čĆą╝ą░čåąĖčÅ, ą┐čĆąĄą┤ąĮą░ąĘąĮą░č湥ąĮąĮą░čÅ ąĖčüą║ą╗čÄčćąĖč鹥ą╗čīąĮąŠ ą┤ą╗čÅ čüą┐ąĄčåąĖą░ą╗ąĖčüč鹊ą▓ ąĘą┤čĆą░ą▓ąŠąŠčģčĆą░ąĮąĄąĮąĖčÅ.
ąĢčüą╗ąĖ ąÆčŗ ąĮąĄ čÅą▓ą╗čÅąĄč鹥čüčī čüą┐ąĄčåąĖą░ą╗ąĖčüč鹊ą╝ ąĘą┤čĆą░ą▓ąŠąŠčģčĆą░ąĮąĄąĮąĖčÅ, ą░ą┤ą╝ąĖąĮąĖčüčéčĆą░čåąĖčÅ ąĮąĄ ąĮąĄčüąĄčé ąŠčéą▓ąĄčéčüčéą▓ąĄąĮąĮąŠčüčéąĖ ąĘą░ ą▓ąŠąĘą╝ąŠąČąĮčŗąĄ ąŠčéčĆąĖčåą░č鹥ą╗čīąĮčŗąĄ ą┐ąŠčüą╗ąĄą┤čüčéą▓ąĖčÅ, ą▓ąŠąĘąĮąĖą║čłąĖąĄ ą▓ čĆąĄąĘčāą╗čīčéą░č鹥 čüą░ą╝ąŠčüč鹊čÅč鹥ą╗čīąĮąŠą│ąŠ ąĖčüą┐ąŠą╗čīąĘąŠą▓ą░ąĮąĖčÅ ąÆą░ą╝ąĖ ąĖąĮč乊čĆą╝ą░čåąĖąĖ čü ą┐ąŠčĆčéą░ą╗ą░ ą▒ąĄąĘ ą┐čĆąĄą┤ą▓ą░čĆąĖč鹥ą╗čīąĮąŠą╣ ą║ąŠąĮčüčāą╗čīčéą░čåąĖąĖ čü ą▓čĆą░č湊ą╝.
ąØą░ąČąĖą╝ą░čÅ ąĮą░ ą║ąĮąŠą┐ą║čā ┬½ąÆąŠą╣čéąĖ┬╗, ąÆčŗ ą┐ąŠą┤čéą▓ąĄčƹȹ┤ą░ąĄč鹥, čćč鹊 čÅą▓ą╗čÅąĄč鹥čüčī ą▓čĆą░č湊ą╝ ąĖą╗ąĖ čüčéčāą┤ąĄąĮč鹊ą╝ ą╝ąĄą┤ąĖčåąĖąĮčüą║ąŠą│ąŠ ą▓čāąĘą░.