đÉđŻđ░đ╗đŞđĚ đŞđĚđ╝đÁđŻđÁđŻđŞđ╣ đ╝đżĐëđŻđżĐüĐéđŻĐőĐů Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║ ĐŹđ╗đÁđ║ĐéĐÇđżĐŹđŻĐćđÁĐäđ░đ╗đżđ│ĐÇđ░đ╝đ╝Đő đ┐ĐÇđŞ đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆĐů đ▓đż┬áđ▓ĐÇđÁđ╝ĐĆ đ┤đŻđÁđ▓đŻđżđ│đż ĐüđŻđ░ đ┐đżĐüđ╗đÁ Đüđ░đ╝đżđ┐ĐÇđżđŞđĚđ▓đżđ╗ĐîđŻĐőĐů đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞđ╣, Đüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░đÁđ╝ĐőĐů đ╝đżđŻđżĐéđżđŻđŻđżđ╣ đ┐ĐüđŞĐůđżđ╝đżĐéđżĐÇđŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéĐîĐÄ: đ▓ đŻđ░Đçđ░đ╗đÁ đżđ┐ĐőĐéđ░ đŞ┬áđ┐đżĐüđ╗đÁ ĐŹđ┐đŞđĚđżđ┤đżđ▓ đ▓ĐéđżĐÇđżđ╣ ĐüĐéđ░đ┤đŞđŞ ĐüđŻđ░
- đÉđŻđŻđżĐéđ░ĐćđŞĐĆ
- đíĐéđ░ĐéĐîĐĆ
- đíĐüĐőđ╗đ║đŞ
- English
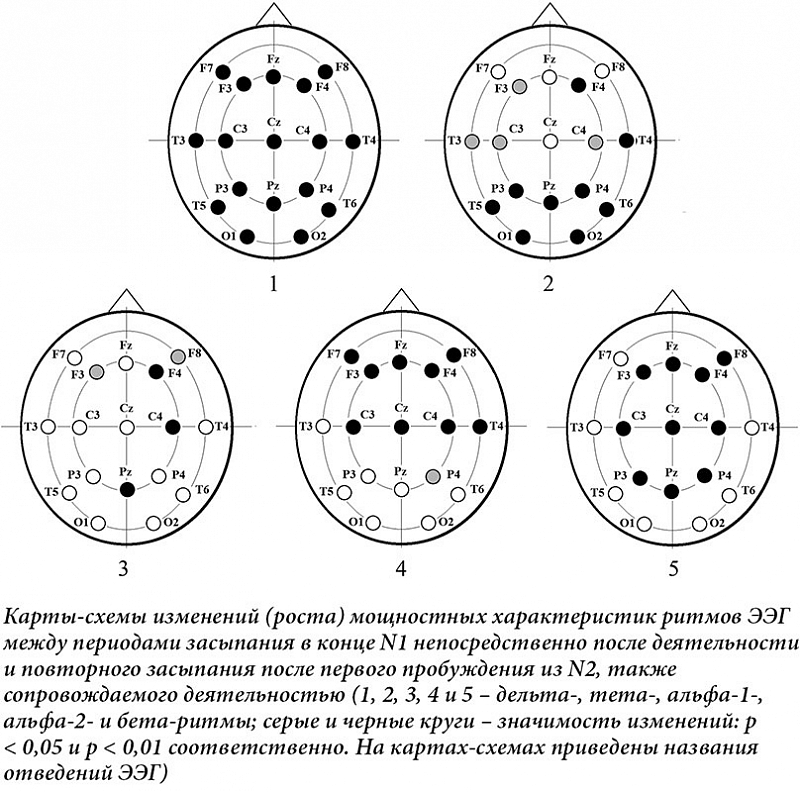

đúĐůĐâđ┤ĐłđÁđŻđŞđÁ đ║đ░ĐçđÁĐüĐéđ▓đ░ ĐüđŻđ░, đ▓┬áĐçđ░ĐüĐéđŻđżĐüĐéđŞ đÁđ│đż ĐäĐÇđ░đ│đ╝đÁđŻĐéđ░ĐćđŞĐĆ ĐÇđ░đĚđŻđżđ╣ ĐŹĐéđŞđżđ╗đżđ│đŞđŞ, đ┤đżĐüĐéđ░ĐéđżĐçđŻđż ĐÇđ░Đüđ┐ĐÇđżĐüĐéĐÇđ░đŻđÁđŻđŻđżđÁ ĐĆđ▓đ╗đÁđŻđŞđÁ [1], đ║đżĐéđżĐÇđżđÁ đ┤đ╗ĐĆ đ▒đżđ╗đÁđÁ ĐâĐüđ┐đÁĐłđŻđżđ╣ ĐéđÁĐÇđ░đ┐đŞđŞ ĐéĐÇđÁđ▒ĐâđÁĐé đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ đ╝đżđĚđ│đżđ▓ĐőĐů đ║đżĐÇĐÇđÁđ╗ĐĆĐéđżđ▓ [2] đ┤đ╗ĐĆ đżĐćđÁđŻđ║đŞ đŻđÁđ╣ĐÇđżđŻđŻĐőĐů đ╝đÁĐůđ░đŻđŞđĚđ╝đżđ▓, đ║đżđ│đŻđŞĐéđŞđ▓đŻđżđ╣ đ┤đŞđŻđ░đ╝đŞđ║đŞ đŞ┬áđ║đ╗đŞđŻđŞĐçđÁĐüđ║đŞĐů đ┐đżĐüđ╗đÁđ┤ĐüĐéđ▓đŞđ╣┬á[3]. đčĐÇđ░đ║ĐéđŞĐçđÁĐüđ║đŞ đ▓ĐüđÁ đŻđ░ĐâĐçđŻĐőđÁ ĐÇđ░đ▒đżĐéĐő đ▓┬áĐŹĐéđżđ╣ đżđ▒đ╗đ░ĐüĐéđŞ ĐüđżĐüĐÇđÁđ┤đżĐéđżĐçđÁđŻĐő đŻđ░┬áđ┐đÁĐÇđŞđżđ┤đÁ đŻđ░Đçđ░đ╗đ░ ĐüđŻđ░ (sleep onset, ĐüĐéđ░đ┤đŞĐĆ ĐüđŻđ░ N1). đčđżđ▓ĐéđżĐÇđŻĐőđÁ đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ đ▓┬áĐüđ╗ĐâĐçđ░đÁ đ┐ĐÇđÁĐÇĐőđ▓đ░đŻđŞĐĆ ĐüđŻđ░ Đüđ░đ╝đżđ┐ĐÇđżđŞđĚđ▓đżđ╗ĐîđŻĐőđ╝đŞ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆđ╝đŞ đżĐüĐéđ░ĐÄĐéĐüĐĆ đ╝đ░đ╗đż đŞđĚĐâĐçđÁđŻđŻĐőđ╝đŞ. đťĐő đ┐đżđ┐ĐőĐéđ░đ╗đŞĐüĐî đ▓đżĐüđ┐đżđ╗đŻđŞĐéĐî ĐŹĐéđżĐé đ┐ĐÇđżđ▒đÁđ╗.
đŁđÁđ╣ĐÇđżĐäđŞđĚđŞđżđ╗đżđ│đŞĐçđÁĐüđ║đŞđÁ đ║đżĐÇĐÇđÁđ╗ĐĆĐéĐő đ┐ĐÇđżĐćđÁĐüĐüđ░ đŻđ░Đçđ░đ╗ĐîđŻđżđ│đż đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ ĐůđżĐÇđżĐłđż đŞđĚĐâĐçđÁđŻĐő [4, 5]. đşĐéđżĐé đ┐ĐÇđżĐćđÁĐüĐü đŻđ░┬áĐŹđ╗đÁđ║ĐéĐÇđżĐŹđŻĐćđÁĐäđ░đ╗đżđ│ĐÇđ░đ╝đ╝đÁ (đşđşđô) Đüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░đÁĐéĐüĐĆ đ║ĐÇđ░Đéđ║đżđ▓ĐÇđÁđ╝đÁđŻđŻĐőđ╝đŞ (đŻđÁĐüđ║đżđ╗Đîđ║đż ĐüđÁđ║ĐâđŻđ┤) đ┐đÁĐÇđÁĐůđżđ┤đ░đ╝đŞ Đü┬áđ▓ĐőĐüđżđ║đżđ╣ Đçđ░ĐüĐéđżĐéđżđ╣ đŞ┬áđŻđŞđĚđ║đżđ╣ đ░đ╝đ┐đ╗đŞĐéĐâđ┤đżđ╣ đ║┬áđŻđŞđĚđ║đżĐçđ░ĐüĐéđżĐéđŻĐőđ╝, đ▓ĐőĐüđżđ║đżđ░đ╝đ┐đ╗đŞĐéĐâđ┤đŻĐőđ╝ đ╝đÁđ┤đ╗đÁđŻđŻĐőđ╝ đ▓đżđ╗đŻđ░đ╝ đŞ┬áđżđ▒ĐÇđ░ĐéđŻđż, ĐçĐéđż Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞđĚĐâđÁĐé đ║đżđ╗đÁđ▒đ░đŻđŞĐĆ đ▒đżđ┤ĐÇĐüĐéđ▓đżđ▓đ░đŻđŞĐĆ đŞ┬á┬źđŞđ╝đÁđÁĐé ĐéđÁđŻđ┤đÁđŻĐćđŞĐÄ ĐüđŞĐüĐéđÁđ╝đ░ĐéđŞĐçđÁĐüđ║đŞ đ┐đżđ┤đ░đ▓đ╗ĐĆĐéĐî đ░đ╗ĐîĐäđ░-ĐÇđŞĐéđ╝ đ▓┬áđ┐đżđ╗ĐîđĚĐâ đ┐ĐÇđżđ│ĐÇđÁĐüĐüđŞđ▓đŻđżđ│đż đĚđ░đ╝đÁđ┤đ╗đÁđŻđŞĐĆ đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ đşđşđô┬╗┬á[4]. đŚđ░ĐüĐőđ┐đ░đŻđŞđÁ đ┐ĐÇđÁđ┤ĐüĐéđ░đ▓đ╗ĐĆđÁĐé Đüđżđ▒đżđ╣ đ┐đżĐüĐéđÁđ┐đÁđŻđŻĐőđ╣ đŞ┬áđŻđÁđżđ┤đŻđżĐÇđżđ┤đŻĐőđ╣ đ┐ĐÇđżĐćđÁĐüĐü đ║đ░đ║ Đü┬áđ┐ĐÇđżĐüĐéĐÇđ░đŻĐüĐéđ▓đÁđŻđŻđżđ╣, Đéđ░đ║┬áđŞ┬áĐü đ▓ĐÇđÁđ╝đÁđŻđŻßŻ╣đ╣ ĐéđżĐçđ║đŞ đĚĐÇđÁđŻđŞĐĆ [5]. đĺ┬áĐÇđ░đĚđŻĐőĐů đżđ▒đ╗đ░ĐüĐéĐĆĐů đ║đżĐÇĐő đ▓đż đ▓ĐÇđÁđ╝ĐĆ đŻđ░ĐüĐéĐâđ┐đ╗đÁđŻđŞĐĆ ĐüđŻđ░ đŻđ░đ▒đ╗ĐÄđ┤đ░ĐÄĐéĐüĐĆ đ╗đżđ║đ░đ╗ĐîđŻĐőđÁ đ┐đ░ĐéĐéđÁĐÇđŻĐő ĐüđŞđŻĐůĐÇđżđŻđŞđĚđ░ĐćđŞđŞ đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ đşđşđô, đ┐đżđ┤ĐÇđ░đĚĐâđ╝đÁđ▓đ░ĐÄĐëđŞđÁ đ▓ĐÇđÁđ╝đÁđŻđŻđżđÁ ĐüđżĐüĐâĐëđÁĐüĐéđ▓đżđ▓đ░đŻđŞđÁ đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ, đ┐đżđ┤đżđ▒đŻđżđ╣ đ▒đżđ┤ĐÇĐüĐéđ▓đżđ▓đ░đŻđŞĐÄ, đŞ┬áđ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ, đ┐đżđ┤đżđ▒đŻđżđ╣ ĐüđŻĐâ. đčĐÇđŞ ĐŹĐéđżđ╝ ĐÇđ░đĚđŻĐőđÁ đżđ▒đ╗đ░ĐüĐéđŞ đ║đżĐÇĐő đ┤đÁđ╝đżđŻĐüĐéĐÇđŞĐÇĐâĐÄĐé ĐüđŻđŞđÂđÁđŻđŞđÁ đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ đŻđÁ┬áđżđ┤đŻđżđ▓ĐÇđÁđ╝đÁđŻđŻđż, đ┐ĐÇđŞĐçđÁđ╝ đ▓┬áĐÇđ░đĚđŻĐőĐů Đçđ░ĐüĐéđżĐéđŻĐőĐů đ┤đŞđ░đ┐đ░đĚđżđŻđ░Đů [6].
đĺ┬áđŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆĐů đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ đŻđ░ĐÇĐĆđ┤Đâ Đü┬áđŻđÁđ╣ĐÇđżĐäđŞđĚđŞđżđ╗đżđ│đŞĐçđÁĐüđ║đŞđ╝đŞ đ║đżĐÇĐÇđÁđ╗ĐĆĐéđ░đ╝đŞ đŞĐüđ┐đżđ╗ĐîđĚĐâĐÄĐéĐüĐĆ đŞ┬áđ┐đżđ▓đÁđ┤đÁđŻĐçđÁĐüđ║đŞđÁ đ╝đ░ĐÇđ║đÁĐÇĐő. đóđ░đ║, đŻđÁđ┐ĐÇđÁĐÇĐőđ▓đŻĐőđÁ đŻđ░đÂđ░ĐéđŞĐĆ đŻđ░┬áđ║đŻđżđ┐đ║Đâ┬á[7], ĐüđÂđŞđ╝đ░đŻđŞĐĆ đ╝ĐĆĐçđŞđ║đ░ đ┤đ╗ĐĆ ĐüđŻĐĆĐéđŞĐĆ ĐüĐéĐÇđÁĐüĐüđ░, đŻđ░đ┐đżđ╗đŻđÁđŻđŻđżđ│đż┬áđ│đÁđ╗đŞđÁđ╝, ĐüđŞđŻĐůĐÇđżđŻđŞđĚđŞĐÇđżđ▓đ░đŻđŻĐőđÁ Đü┬áđ┤ĐőĐůđ░đŻđŞđÁđ╝ [8], đŞ┬áĐçđÁĐÇđÁđ┤đżđ▓đ░đŻđŞđÁ đŻđ░đÂđ░ĐéđŞđ╣ đŻđ░┬áđ║đŻđżđ┐đ║Đâ đ┐ĐÇđ░đ▓đżđ╣ đŞ┬áđ╗đÁđ▓đżđ╣ ĐÇĐâđ║đżđ╣ đ┐đż┬áđ┤đÁĐüĐĆĐéĐî ĐÇđ░đĚ Đü┬áĐüđŞđŻĐůĐÇđżđŻđŻĐőđ╝ ĐüĐçđÁĐéđżđ╝ đ▓┬áĐâđ╝đÁ (đ┐ĐüđŞĐůđżđ╝đżĐéđżĐÇđŻĐőđ╣ ĐéđÁĐüĐé, đčđťđó) [9] ĐůđżĐÇđżĐłđż ĐÇđÁđ░đ│đŞĐÇĐâĐÄĐé đŻđ░┬áđŞđĚđ╝đÁđŻđÁđŻđŞĐĆ đ▓┬áĐüđżĐüĐéđżĐĆđŻđŞđŞ đżđ▒Đüđ╗đÁđ┤ĐâđÁđ╝đżđ│đż. đťđżđŻđżĐéđżđŻđŻđ░ĐĆ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéĐî đ┐đżđ│ĐÇĐâđÂđ░đÁĐé đ▒đżđ╗ĐîĐłđŞđŻĐüĐéđ▓đż ĐüĐâđ▒ĐŐđÁđ║Đéđżđ▓ đ▓┬áĐüđżđŻ đ║đ░đ║ đ▓┬áđŻđ░Đçđ░đ╗đÁ đżđ┐ĐőĐéđ░, Đéđ░đ║ đŞ┬áđ┐đżđĚđÂđÁ, đ┐đżĐüđ╗đÁ Đüđ┐đżđŻĐéđ░đŻđŻĐőĐů đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞđ╣ đŞ┬áđ▓đżđĚđżđ▒đŻđżđ▓đ╗đÁđŻđŞĐĆ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ, đ┐ĐÇđÁĐÇđ▓đ░đŻđŻđżđ╣ ĐüđŻđżđ╝. đáđ░Đüđ┐ĐÇđÁđ┤đÁđ╗đÁđŻđŞđÁ ĐŹđ┐đŞđĚđżđ┤đżđ▓ đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ đŞ┬áđ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ đ▓đż đ▓ĐÇđÁđ╝ĐĆ ĐüđŻđ░, đŞĐů đ┐ĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻđżĐüĐéĐî, đ░┬áđ▓ đ┐đżĐüđ╗đÁđ┤đŻđÁđ╝ Đüđ╗ĐâĐçđ░đÁ đŞ┬áđ┐ĐÇđ░đ▓đŞđ╗ĐîđŻđżĐüĐéĐî đ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐĆ đ┐ĐÇđÁđ┤đ┐đŞĐüđ░đŻđŻđżđ╣ đŞđŻĐüĐéĐÇĐâđ║ĐćđŞđÁđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đ┐đżđĚđ▓đżđ╗ĐĆĐÄĐé đżĐćđÁđŻđŞĐéĐî ĐäĐâđŻđ║ĐćđŞđżđŻđ░đ╗ĐîđŻđżđÁ ĐüđżĐüĐéđżĐĆđŻđŞđÁ ĐüĐâđ▒ĐŐđÁđ║Đéđ░ đŞ┬áĐâĐÇđżđ▓đÁđŻĐî đÁđ│đż ĐüđżđĚđŻđ░đŻđŞĐĆ đ▓┬áĐŹĐéđŞ đ┐đÁĐÇđŞđżđ┤Đő.
đáđ░đŻđÁđÁ đ▓┬áđżđ┐ĐőĐéđ░Đů Đü┬áđ┤đŻđÁđ▓đŻĐőđ╝ ĐüđŻđżđ╝, đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆĐů đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ đ┐đżĐüđ╗đÁ Đüđ┐đżđŻĐéđ░đŻđŻĐőĐů đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞđ╣ Đü┬áđ┐đżđ╝đżĐëĐîĐÄ đčđťđó đ▓┬áđÁđ│đż ĐâđŻđŞđ╝đ░đŻĐâđ░đ╗ĐîđŻđżđ╣ đ╝đżđ┤đŞĐäđŞđ║đ░ĐćđŞđŞ (ĐçđÁĐÇđÁđ┤đżđ▓đ░đŻđŞđÁ đ┤đÁĐüĐĆĐéđŞ đŻđ░đÂđ░ĐéđŞđ╣ đŻđ░┬áđ║đŻđżđ┐đ║Đâ đ┐ĐÇđ░đ▓đżđ╣ ĐÇĐâđ║đżđ╣ Đüđż┬áĐüĐçđÁĐéđżđ╝ đ▓┬áĐâđ╝đÁ đŞ┬áĐéđżđ╗Đîđ║đż ĐüĐçđÁĐé đ▒đÁđĚ đŻđ░đÂđ░ĐéđŞđ╣) [10] đ╝Đő đ┐đżđ║đ░đĚđ░đ╗đŞ Đüđ╗đÁđ┤ĐâĐÄĐëđÁđÁ. đčĐÇđŞ đżđ▒ĐëđÁđ╣ ĐéđÁđŻđ┤đÁđŻĐćđŞđŞ ĐüđŻđŞđÂđÁđŻđŞĐĆ ĐüĐéđÁđ┐đÁđŻđŞ đ▒đżđ┤ĐÇĐüĐéđ▓đżđ▓đ░đŻđŞĐĆ đ▓đż đ▓ĐÇđÁđ╝ĐĆ đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ ĐüđżĐůĐÇđ░đŻđÁđŻđŞđÁ ĐâĐÇđżđ▓đŻĐĆ đ░đ║ĐéđŞđ▓đ░ĐćđŞđŞ đ╗đżđ▒đŻĐőĐů đżđ▒đ╗đ░ĐüĐéđÁđ╣ đżđ▒đÁĐüđ┐đÁĐçđŞđ▓đ░đÁĐé đ▓Đőđ┐đżđ╗đŻđÁđŻđŞđÁ đ▓┬áĐŹĐéđŞĐů ĐâĐüđ╗đżđ▓đŞĐĆĐů đ┐ĐüđŞĐůđżđ╝đżĐéđżĐÇđŻđżđ╣, đ║đżđ│đŻđŞĐéđŞđ▓đŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ. đĽđÁ đ┐ĐÇđÁĐÇĐőđ▓đ░đŻđŞđÁ đ▓┬áđŻđ░Đçđ░đ╗đÁ ĐüđŻđ░ (đŻđ░┬áđ┐đÁĐÇđ▓đżđ╣ ĐüĐéđ░đ┤đŞđŞ), đ▓đÁĐÇđżĐĆĐéđŻđż, đżĐüĐéđ░đ▓đ╗ĐĆđÁĐé Đüđ╗đÁđ┤đżđ▓đżđ╣ ĐŹĐäĐäđÁđ║Đé đ▓┬áđ▓đŞđ┤đÁ đ┐ĐÇđżđ┤đżđ╗đÂđ░ĐÄĐëđÁđ╣ĐüĐĆ đ░đ║ĐéđŞđ▓đ░ĐćđŞđŞ, đ║đżĐéđżĐÇđ░ĐĆ đ┐đżĐüđ╗đÁ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ đ┤đ░đÁĐé đ▓đżđĚđ╝đżđÂđŻđżĐüĐéĐî đ▒ĐőĐüĐéĐÇđż đ▓đżđĚđżđ▒đŻđżđ▓đŞĐéĐî đ▓Đőđ┐đżđ╗đŻđÁđŻđŞđÁ đĚđ░đ┤đ░đŻđŞđ╣ ĐéđÁĐüĐéđ░ [11]. đÜĐÇđżđ╝đÁ Đéđżđ│đż, đ╝Đő đżđ▒đŻđ░ĐÇĐâđÂđŞđ╗đŞ, ĐçĐéđż đ▒đżđ╗đÁđÁ đ┤đ╗đŞĐéđÁđ╗ĐîđŻđżđÁ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞđÁ đ┐đż┬áđżĐéđŻđżĐłđÁđŻđŞĐÄ đ║┬áđ▒đżđ╗đÁđÁ đ║đżĐÇđżĐéđ║đżđ╝Đâ (đżđ▒đ░ Đüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░đ╗đŞĐüĐî đ▓Đőđ┐đżđ╗đŻđÁđŻđŞđÁđ╝ đčđťđó đŞ┬áđ┤đ░đ╗đÁđÁ đ┐đÁĐÇđÁĐůđżđ┤đŞđ╗đŞ đ▓┬áđĚđ░ĐüĐőđ┐đ░đŻđŞđÁ) đ┐đżĐüđ╗đÁ ĐŹđ┐đŞđĚđżđ┤đ░ đ▓ĐéđżĐÇđżđ╣ ĐüĐéđ░đ┤đŞđŞ ĐüđŻđ░ (N2) Đüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░đÁĐéĐüĐĆ đ▒đżđ╗đÁđÁ đ▓ĐőĐÇđ░đÂđÁđŻđŻĐőđ╝ ĐłđŞĐÇđżđ║đż ĐÇđ░Đüđ┐ĐÇđżĐüĐéĐÇđ░đŻđÁđŻđŻĐőđ╝ đ┐đż┬áđ║đżĐÇđÁ đŻđŞđĚđ║đżĐçđ░ĐüĐéđżĐéđŻĐőđ╝ đ░đ╗ĐîĐäđ░-ĐÇđŞĐéđ╝đżđ╝┬á[12]. đśđŻĐőđ╝đŞ Đüđ╗đżđ▓đ░đ╝đŞ, đ╝Đő ĐâĐüĐéđ░đŻđżđ▓đŞđ╗đŞ đĚđ░đ▓đŞĐüđŞđ╝đżĐüĐéĐî Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║ đşđşđô đ▓đż đ▓ĐÇđÁđ╝ĐĆ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ đżĐé┬áđÁđ│đż đ┐ĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻđżĐüĐéđŞ.
đŽđÁđ╗ĐîĐÄ đ┤đ░đŻđŻđżđ╣ ĐÇđ░đ▒đżĐéĐő ĐĆđ▓đ╗ĐĆđÁĐéĐüĐĆ đŞđĚĐâĐçđÁđŻđŞđÁ đŞđĚđ╝đÁđŻđÁđŻđŞđ╣ đŻđÁđ╣ĐÇđżĐäđŞđĚđŞđżđ╗đżđ│đŞĐçđÁĐüđ║đŞĐů đ┐đżđ║đ░đĚđ░ĐéđÁđ╗đÁđ╣, đ║đżĐéđżĐÇĐőđÁ Đüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░ĐÄĐé đ║đżđ╗đÁđ▒đ░đŻđŞĐĆ ĐäĐâđŻđ║ĐćđŞđżđŻđ░đ╗ĐîđŻđżđ│đż ĐüđżĐüĐéđżĐĆđŻđŞĐĆ đŞĐüđ┐ĐőĐéĐâđÁđ╝đżđ│đż, đŞ┬áĐâĐÇđżđ▓đŻĐĆ đÁđ│đż ĐüđżđĚđŻđ░đŻđŞĐĆ đ┐ĐÇđŞ đ┐đżđ▓ĐéđżĐÇđŻĐőĐů đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆĐů đ┐đżĐüđ╗đÁ ĐŹđ┐đŞđĚđżđ┤đżđ▓ đ▓đżđĚđżđ▒đŻđżđ▓đ╗đÁđŻđŞĐĆ đ┐ĐüđŞĐůđżđ╝đżĐéđżĐÇđŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ┬áÔÇô đ▓┬áđŻđ░Đçđ░đ╗ĐîđŻĐőđ╣ đ┐đÁĐÇđŞđżđ┤ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐĆ ĐéđÁĐüĐéđ░ (N1) đŞ┬áđŻđ░ N2.
đĺ┬áđĚđ░đ┤đ░ĐçđŞ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ đ▓Đůđżđ┤đŞđ╗đŞ:
- đ▓ĐőĐçđŞĐüđ╗đÁđŻđŞđÁ đ╝đżĐëđŻđżĐüĐéđŻĐőĐů đ┐đżđ║đ░đĚđ░ĐéđÁđ╗đÁđ╣ đ┤đÁđ╗ĐîĐéđ░-, ĐéđÁĐéđ░-, đ░đ╗ĐîĐäđ░-1-, đ░đ╗ĐîĐäđ░-2- đŞ┬áđ▒đÁĐéđ░-ĐÇđŞĐéđ╝đżđ▓ đşđşđô đ┐ĐÇđŞ đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆĐů đ▓┬áĐéđÁĐçđÁđŻđŞđÁ đ┤đŻđÁđ▓đŻđżđ│đż ĐüđŻđ░ đ▓đż đ▓ĐÇđÁđ╝ĐĆ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐĆ đčđťđó;
- ĐüĐÇđ░đ▓đŻđÁđŻđŞđÁ đ▓ĐőĐçđŞĐüđ╗đÁđŻđŻĐőĐů đ▓đÁđ╗đŞĐçđŞđŻ:
- đŻđ░┬áĐÇđ░đĚđŻĐőĐů ĐüĐéđ░đ┤đŞĐĆĐů ĐüđŻđ░ (đ▓┬áđ║đżđŻĐćđÁ đŻđ░Đçđ░đ╗ĐîđŻđżđ│đż đ┐đÁĐÇđŞđżđ┤đ░ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐĆ ĐéđÁĐüĐéđ░ (N1), đĚđ░ đ║đżĐéđżĐÇĐőđ╝ Đüđ╗đÁđ┤ĐâđÁĐé đ▓ĐéđżĐÇđ░ĐĆ ĐüĐéđ░đ┤đŞĐĆ ĐüđŻđ░ (N2)), đŞ┬áđŻđ░ N2 đ┐đżĐüđ╗đÁ Đüđ┐đżđŻĐéđ░đŻđŻĐőĐů đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞđ╣;
- đ┐đżĐüđ╗đÁ ĐŹđ┐đŞđĚđżđ┤đżđ▓ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ ĐÇđ░đĚđŻđżđ╣ đ┐ĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻđżĐüĐéđŞ;
- đŻđ░┬áđ┤đ▓ĐâĐů đ┐đżĐüđ╗đÁđ┤đżđ▓đ░ĐéđÁđ╗ĐîđŻĐőĐů đ┤đÁĐüĐĆĐéđŞĐüđÁđ║ĐâđŻđ┤đŻĐőĐů đżĐéĐÇđÁđĚđ║đ░Đů đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ.
đťđ░ĐéđÁĐÇđŞđ░đ╗ đŞ┬áđ╝đÁĐéđżđ┤Đő
đĺ┬áđŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞđŞ đ┐ĐÇđŞđŻđŞđ╝đ░đ╗đŞ ĐâĐçđ░ĐüĐéđŞđÁ 34 ĐüĐéĐâđ┤đÁđŻĐéđ░ đ╝đżĐüđ║đżđ▓Đüđ║đŞĐů đ▓ĐâđĚđżđ▓ (26 đÂđÁđŻĐëđŞđŻ đŞ┬áđ▓đżĐüđÁđ╝Đî đ╝ĐâđÂĐçđŞđŻ) đ▓┬áđ▓đżđĚĐÇđ░ĐüĐéđÁ 19ÔÇô22 đ╗đÁĐé, đ┐ĐÇđ░đ║ĐéđŞĐçđÁĐüđ║đŞ đĚđ┤đżĐÇđżđ▓ĐőđÁ đ╗đŞĐćđ░, đ┐ĐÇđ░đ▓ĐłđŞ. đśĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞđÁ đżđ┤đżđ▒ĐÇđÁđŻđż ĐŹĐéđŞĐçđÁĐüđ║đżđ╣ đ║đżđ╝đŞĐüĐüđŞđÁđ╣ đśđŻĐüĐéđŞĐéĐâĐéđ░ đ▓ĐőĐüĐłđÁđ╣ đŻđÁĐÇđ▓đŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đŞ┬áđŻđÁđ╣ĐÇđżĐäđŞđĚđŞđżđ╗đżđ│đŞđŞ đáđÉđŁ (đ┐ĐÇđżĐéđżđ║đżđ╗ đżĐé┬á3 đŞĐÄđŻĐĆ 2019┬áđ│.┬áÔäľ 2) đŞ┬áĐüđżđżĐéđ▓đÁĐéĐüĐéđ▓đżđ▓đ░đ╗đż ĐŹĐéđŞĐçđÁĐüđ║đŞđ╝ đŻđżĐÇđ╝đ░đ╝ đąđÁđ╗ĐîĐüđŞđŻđ║Đüđ║đżđ╣ đ┤đÁđ║đ╗đ░ĐÇđ░ĐćđŞđŞ đĺĐüđÁđ╝đŞĐÇđŻđżđ╣ đ╝đÁđ┤đŞĐćđŞđŻĐüđ║đżđ╣ đ░ĐüĐüđżĐćđŞđ░ĐćđŞđŞ ┬źđşĐéđŞĐçđÁĐüđ║đŞđÁ đ┐ĐÇđŞđŻĐćđŞđ┐Đő đ┐ĐÇđżđ▓đÁđ┤đÁđŻđŞĐĆ đŻđ░ĐâĐçđŻĐőĐů đ╝đÁđ┤đŞĐćđŞđŻĐüđ║đŞĐů đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞđ╣ Đü┬áĐâĐçđ░ĐüĐéđŞđÁđ╝ ĐçđÁđ╗đżđ▓đÁđ║đ░┬╗ Đü┬áđ┐đżđ┐ĐÇđ░đ▓đ║đ░đ╝đŞ 2000┬áđ│.┬áđŁđÁđ▓ĐÇđżđ╗đżđ│đŞĐçđÁĐüđ║đŞđÁ, đ┐ĐüđŞĐůđŞđ░ĐéĐÇđŞĐçđÁĐüđ║đŞđÁ đĚđ░đ▒đżđ╗đÁđ▓đ░đŻđŞĐĆ đŞ┬áĐÇđ░ĐüĐüĐéĐÇđżđ╣ĐüĐéđ▓đ░ ĐüđŻđ░ Đâ┬áĐâĐçđ░ĐüĐéđŻđŞđ║đżđ▓ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ ĐÇđ░đŻđÁđÁ đŻđÁ┬áđ┤đŞđ░đ│đŻđżĐüĐéđŞĐÇđżđ▓đ░đ╗đŞĐüĐî, đżđŻđŞ đŻđÁ┬áđ┐ĐÇđŞđŻđŞđ╝đ░đ╗đŞ đ╗đÁđ║đ░ĐÇĐüĐéđ▓đÁđŻđŻĐőĐů đ┐ĐÇđÁđ┐đ░ĐÇđ░Đéđżđ▓. đíđżđ│đ╗đ░ĐüđŻđż đŞđŻĐüĐéĐÇĐâđ║ĐćđŞđŞ, đżđŻđŞ đ┤đżđ╗đÂđŻĐő đ▒Đőđ╗đŞ đżĐéđ║đ░đĚđ░ĐéĐîĐüĐĆ đżĐé┬áđ┐ĐÇđŞđÁđ╝đ░ đ░đ╗đ║đżđ│đżđ╗ĐĆ đĚđ░ ĐüĐâĐéđ║đŞ đ┤đż đŻđ░Đçđ░đ╗đ░ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ. đĺ┬áđ┤đÁđŻĐî ĐŹđ║Đüđ┐đÁĐÇđŞđ╝đÁđŻĐéđ░ đ▒Đőđ╗đż ĐÇđÁđ║đżđ╝đÁđŻđ┤đżđ▓đ░đŻđż đ▓đżđĚđ┤đÁĐÇđÂđ░ĐéĐîĐüĐĆ đżĐé┬áđ┐ĐÇđŞđÁđ╝đ░ ĐéđżđŻđŞđĚđŞĐÇĐâĐÄĐëđŞĐů ĐüĐÇđÁđ┤ĐüĐéđ▓, Đéđ░đ║đŞĐů đ║đ░đ║ đ║đżĐäđÁ, đ║ĐÇđÁđ┐đ║đŞđ╣ Đçđ░đ╣, Đłđżđ║đżđ╗đ░đ┤, ĐŹđŻđÁĐÇđ│đÁĐéđŞĐçđÁĐüđ║đŞđÁ đŻđ░đ┐đŞĐéđ║đŞ đŞ┬áđ┐ĐÇđżĐç. đčĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻđżĐüĐéĐî đŻđżĐçđŻđżđ│đż ĐüđŻđ░ đ┐đÁĐÇđÁđ┤ đ┤đŻđÁđ▓đŻĐőđ╝ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞđÁđ╝ đżđ┐ĐÇđÁđ┤đÁđ╗ĐĆđ╗đŞ Đü┬áđ┐đżđ╝đżĐëĐîĐÄ đżđ┐ĐÇđżĐüđŻđŞđ║đ░ ┬źđöđŻđÁđ▓đŻđŞđ║ ĐüđŻđ░┬╗ [13]. đśĐüđ┐ĐőĐéĐâđÁđ╝ĐőđÁ Đüđ┐đ░đ╗đŞ đ▓┬áĐüĐÇđÁđ┤đŻđÁđ╝ (M) 5,75 ┬▒ 2,26 Đçđ░Đüđ░,
đśĐüđ┐đżđ╗ĐîđĚđżđ▓đ░đ╗đŞ đŻđÁđ┐ĐÇđÁĐÇĐőđ▓đŻđż-đ┤đŞĐüđ║ĐÇđÁĐéđŻĐőđ╣ đčđťđó đ▓┬áĐâđŻđŞđ╝đ░đŻĐâđ░đ╗ĐîđŻđżđ╣ đ╝đżđ┤đŞĐäđŞđ║đ░ĐćđŞđŞ [10]. đśĐüđ┐ĐőĐéĐâđÁđ╝ĐőđÁ ĐüĐçđŞĐéđ░đ╗đŞ đ▓┬áĐâđ╝đÁ đżĐé┬áđżđ┤đŻđżđ│đż đ┤đż đ┤đÁĐüĐĆĐéđŞ, đżđ┤đŻđżđ▓ĐÇđÁđ╝đÁđŻđŻđż đ┐ĐÇđŞ đ║đ░đÂđ┤đżđ╝ đżĐéĐüĐçđÁĐéđÁ đżđŻđŞ đ┤đżđ╗đÂđŻĐő đ▒Đőđ╗đŞ đ▒đżđ╗ĐîĐłđŞđ╝ đ┐đ░đ╗ĐîĐćđÁđ╝ đ┐ĐÇđ░đ▓đżđ╣ ĐÇĐâđ║đŞ đŻđ░đÂđŞđ╝đ░ĐéĐî đŻđ░┬áđ║đŻđżđ┐đ║Đâ, đĚđ░ĐäđŞđ║ĐüđŞĐÇđżđ▓đ░đŻđŻĐâĐÄ đŻđ░┬áĐâđ║đ░đĚđ░ĐéđÁđ╗ĐîđŻđżđ╝ đ┐đ░đ╗ĐîĐćđÁ Đéđżđ╣ đÂđÁ ĐÇĐâđ║đŞ. đŚđ░ĐéđÁđ╝ Đüđ╗đÁđ┤đżđ▓đ░đ╗đż đ┐ĐÇđżđ┤đżđ╗đÂđŞĐéĐî ĐüĐçđÁĐé đ▓┬áĐâđ╝đÁ đżĐé┬áđżđ┤đŻđżđ│đż đ┤đż đ┤đÁĐüĐĆĐéđŞ đ▒đÁđĚ đŻđ░đÂđ░ĐéđŞđ╣. đžđÁĐÇđÁđ┤đżđ▓đ░đŻđŞđÁ ĐüĐçđÁĐéđ░ Đü┬áđŻđ░đÂđ░ĐéđŞĐĆđ╝đŞ đŞ┬áđ▒đÁđĚ đŻđ░đÂđ░ĐéđŞđ╣ ĐâĐçđ░ĐüĐéđŻđŞđ║đŞ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ đ┐ĐÇđżđ┤đżđ╗đÂđ░đ╗đŞ đ┤đż ĐéđÁĐů đ┐đżĐÇ, đ┐đżđ║đ░ đŻđÁ┬áđĚđ░ĐüĐőđ┐đ░đ╗đŞ. đčđżđ║đ░đĚđ░ĐéđÁđ╗đÁđ╝ đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ Đüđ╗ĐâđÂđŞđ╗đż đ┐ĐÇđÁđ║ĐÇđ░ĐëđÁđŻđŞđÁ đŻđ░đÂđ░ĐéđŞđ╣ đŻđ░┬áđ║đŻđżđ┐đ║Đâ. đčĐÇđŞ Đüđ┐đżđŻĐéđ░đŻđŻĐőĐů đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆĐů đŞĐüđ┐ĐőĐéĐâđÁđ╝ĐőđÁ đ┤đżđ╗đÂđŻĐő đ▒Đőđ╗đŞ đ▓đżđĚđżđ▒đŻđżđ▓đŞĐéĐî đ▓Đőđ┐đżđ╗đŻđÁđŻđŞđÁ ĐéđÁĐüĐéđ░.
đŁđ░Đçđ░đ╗đż ĐŹđ║Đüđ┐đÁĐÇđŞđ╝đÁđŻĐéđ░┬áÔÇô Đü┬á13.00 đ┤đż 16.00. đčĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻđżĐüĐéĐî đĚđ░đ┐đŞĐüđŞ đşđşđô┬áÔÇô đżĐé┬á55 đ╝đŞđŻĐâĐé đ┤đż 1 Đçđ░Đüđ░ 10┬áđ╝đŞđŻĐâĐé. đśĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞđÁ đ┐ĐÇđżđ▓đżđ┤đŞđ╗đżĐüĐî đ▓┬áđĚđ░ĐéđÁđ╝đŻđÁđŻđŻđżđ╝ đĚđ▓Đâđ║đżđŞđĚđżđ╗đŞĐÇĐâĐÄĐëđÁđ╝ đ┐ĐÇđżđ▓đÁĐéĐÇđŞđ▓đ░đÁđ╝đżđ╝ đ┐đżđ╝đÁĐëđÁđŻđŞđŞ, đ▓┬áđ║đżĐéđżĐÇđżđ╝ đ┐đżđ┤đ┤đÁĐÇđÂđŞđ▓đ░đ╗đ░ĐüĐî đ┐đżĐüĐéđżĐĆđŻđŻđ░ĐĆ đ║đżđ╝ĐäđżĐÇĐéđŻđ░ĐĆ ĐéđÁđ╝đ┐đÁĐÇđ░ĐéĐâĐÇđ░. đĺđż đ▓ĐÇđÁđ╝ĐĆ đĚđ░đ┐đŞĐüđŞ đşđşđô đŞĐüđ┐ĐőĐéĐâđÁđ╝ĐőđÁ đŻđ░Đůđżđ┤đŞđ╗đŞĐüĐî đ▓┬áđ┐đżđ╗đżđÂđÁđŻđŞđŞ đ╗đÁđÂđ░ (đŻđ░┬áđ║ĐâĐłđÁĐéđ║đÁ) Đü┬áđĚđ░đ║ĐÇĐőĐéĐőđ╝đŞ┬áđ│đ╗đ░đĚđ░đ╝đŞ.
đşđşđô ĐÇđÁđ│đŞĐüĐéĐÇđŞĐÇđżđ▓đ░đ╗đŞ Đü┬áđ┐đżđ╝đżĐëĐîĐÄ Đłđ╗đÁđ╝đ░ Đü┬áĐůđ╗đżĐÇĐüđÁĐÇđÁđ▒ĐÇĐĆđŻĐőđ╝đŞ ĐŹđ╗đÁđ║ĐéĐÇđżđ┤đ░đ╝đŞ Đü┬á17 đżĐéđ▓đÁđ┤đÁđŻđŞĐĆđ╝đŞ đ▓┬áĐüđżđżĐéđ▓đÁĐéĐüĐéđ▓đŞđŞ Đüđż┬áĐüĐůđÁđ╝đżđ╣ 10ÔÇô20% (ĐÇđÁĐäđÁĐÇđÁđŻĐéđŻĐőđÁ ĐŹđ╗đÁđ║ĐéĐÇđżđ┤Đő ĐÇđ░Đüđ┐đżđ╗đ░đ│đ░đ╗đŞĐüĐî đŻđ░┬áđ╝đ░ĐüĐéđżđŞđ┤đ░Đů, Đüđżđ┐ĐÇđżĐéđŞđ▓đ╗đÁđŻđŞđÁ┬áÔÇô đ┤đż 5 đ║đ×đ╝, Đçđ░ĐüĐéđżĐéđ░ đ┤đŞĐüđ║ĐÇđÁĐéđŞđĚđ░ĐćđŞđŞ┬áÔÇô 500 đôĐć, đ┐đżđ╗đżĐüđ░ đ┐ĐÇđżđ┐ĐâĐüđ║đ░đŻđŞĐĆ ĐâĐüđŞđ╗đŞĐéđÁđ╗ĐĆ┬áÔÇô 0,5ÔÇô40 đôĐć). đŚđ░đ┐đŞĐüĐőđ▓đ░đ╗đŞĐüĐî Đéđ░đ║đÂđÁ ĐŹđ╗đÁđ║ĐéĐÇđżđżđ║Đâđ╗đżđ│ĐÇđ░đ╝đ╝đ░ đŞ┬áđ╝đÁĐůđ░đŻđżđ│ĐÇđ░đ╝đ╝đ░ đŻđ░đÂđ░ĐéđŞđ╣ đŻđ░┬áđ║đŻđżđ┐đ║đŞ.
đčĐÇđŞ đ┐đÁĐÇđ▓đżđ╝ đ▓đŞđ┤đÁ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ đĚđ░ĐüĐőđ┐đ░đŻđŞđ╣ Đâ┬áđ║đ░đÂđ┤đżđ│đż đŞĐüđ┐ĐőĐéĐâđÁđ╝đżđ│đż đ▓Đőđ┤đÁđ╗ĐĆđ╗đŞ đ┤đ▓đ░ đżĐéĐÇđÁđĚđ║đ░ đ▒đÁđĚđ░ĐÇĐéđÁĐäđ░đ║ĐéđŻđżđ╣ đĚđ░đ┐đŞĐüđŞ đşđşđô┬áÔÇô đ▓┬áđ║đżđŻĐćđÁ N1 đ┐đżĐüđ╗đÁ đ┐đżĐüđ╗đÁđ┤đŻđÁđ│đż đ┐đ░ĐéĐéđÁĐÇđŻđ░ đŻđ░đÂđ░ĐéđŞđ╣, đĚđ░ đ║đżĐéđżĐÇĐőđ╝ ĐçđÁĐÇđÁđĚ đŻđÁđ┐ĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻđżđÁ đ▓ĐÇđÁđ╝ĐĆ Đüđ╗đÁđ┤đżđ▓đ░đ╗đ░ N2, đŞ┬áđ┐đż đżđ║đżđŻĐçđ░đŻđŞđŞ Đüđ┐đżđŻĐéđ░đŻđŻđżđ│đż đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ đ┐đżĐüđ╗đÁ đ┐đÁĐÇđ▓đżđ│đż ĐŹđ┐đŞđĚđżđ┤đ░ N2. đöđ╗đŞđŻđ░ đżĐéĐÇđÁđĚđ║đżđ▓┬áÔÇô 30 ĐüđÁđ║ĐâđŻđ┤. đčĐÇđŞđŻđ░đ┤đ╗đÁđÂđŻđżĐüĐéĐî ĐâĐçđ░ĐüĐéđ║đżđ▓ đşđşđô đ║┬áđ┐đÁĐÇđŞđżđ┤Đâ đŻđ░Đçđ░đ╗đ░ ĐüđŻđ░ (sleep onset) đżĐćđÁđŻđŞđ▓đ░đ╗đŞ đ▓đŞđĚĐâđ░đ╗ĐîđŻđż Đü┬áđ┐đżđ╝đżĐëĐîĐÄ ĐüĐéđ░đŻđ┤đ░ĐÇĐéđŻĐőĐů đ║ĐÇđŞĐéđÁĐÇđŞđÁđ▓ AASM [14]. đöđżđ┐đżđ╗đŻđŞĐéđÁđ╗ĐîđŻĐőđ╝ đ║ĐÇđŞĐéđÁĐÇđŞđÁđ╝ đŻđ░Đçđ░đ╗đ░ đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ Đüđ╗ĐâđÂđŞđ╗đż đżĐéĐüĐâĐéĐüĐéđ▓đŞđÁ đŻđ░┬áđ╝đÁĐůđ░đŻđżđ│ĐÇđ░đ╝đ╝đÁ đ┐đżđ▓đÁđ┤đÁđŻĐçđÁĐüđ║đŞĐů đ┐đżđ║đ░đĚđ░ĐéđÁđ╗đÁđ╣ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐĆ đčđťđó.
đčĐÇđŞ đ▓ĐéđżĐÇđżđ╝ đ▓đŞđ┤đÁ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ Đâ┬áđŞĐüđ┐ĐőĐéĐâđÁđ╝ĐőĐů đ▓Đőđ┤đÁđ╗ĐĆđ╗đŞ đ┐đż┬áĐçđÁĐéĐőĐÇđÁ đ▒đÁđĚđ░ĐÇĐéđÁĐäđ░đ║ĐéđŻĐőĐů đżĐéĐÇđÁđĚđ║đ░ đşđşđô, đ║đżĐéđżĐÇĐőđÁ ĐüđżđżĐéđ▓đÁĐéĐüĐéđ▓đżđ▓đ░đ╗đŞ đĚđ░ĐüĐőđ┐đ░đŻđŞĐÄ đ┐đżĐüđ╗đÁ ĐŹđ┐đŞđĚđżđ┤đ░ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ. đčđÁĐÇđ▓ĐőđÁ đ┤đ▓đ░┬áÔÇô đ▓┬áđŻđ░Đçđ░đ╗đÁ đżđ┐ĐőĐéđ░, đ┤đż đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ, đ║đżĐéđżĐÇđżđÁ đ┐ĐÇđŞđ▓đżđ┤đŞđ╗đż đ║┬áđ┐đżĐĆđ▓đ╗đÁđŻđŞĐÄ đ┐đÁĐÇđ▓đżđ│đż ĐŹđ┐đŞđĚđżđ┤đ░ N2. đşĐéđżđ╝Đâ đ┐ĐÇđÁđ┤ĐłđÁĐüĐéđ▓đżđ▓đ░đ╗đŞ Đüđ╗đÁđ┤ĐâĐÄĐëđŞđÁ đżđ▒ĐüĐéđżĐĆĐéđÁđ╗ĐîĐüĐéđ▓đ░. đíĐâđ▒ĐŐđÁđ║Đé đ▒đÁđĚđ┤đÁđ╣ĐüĐéđ▓đżđ▓đ░đ╗ đŻđÁ┬áđ╝đÁđŻđÁđÁ 30 ĐüđÁđ║ĐâđŻđ┤, đ┤đ░đ╗đÁđÁ đżđŻ đ▓ĐőĐůđżđ┤đŞđ╗ đŞđĚ┬áđ┤ĐÇđÁđ╝đżĐéđŻđżđ│đż ĐüđżĐüĐéđżĐĆđŻđŞĐĆ đŞ┬áđŻđ░ĐçđŞđŻđ░đ╗ đŻđ░đÂđŞđ╝đ░ĐéĐî đŻđ░┬áđ║đŻđżđ┐đ║Đâ. đĺ┬áđ┐đÁĐÇđ▓đżđ╝ Đüđ╗ĐâĐçđ░đÁ đ▒Đőđ╗ đżđ┤đŞđŻ đŻđÁđ┐đżđ╗đŻĐőđ╣ đ┐đ░ĐéĐéđÁĐÇđŻ đŻđ░đÂđ░ĐéđŞđ╣ (đżĐé┬áĐłđÁĐüĐéđŞ đ┤đż đ┤đÁđ▓ĐĆĐéđŞ), đ▓đż đ▓ĐéđżĐÇđżđ╝┬áÔÇô đŻđÁ┬áđ╝đÁđŻđÁđÁ đżđ┤đŻđżđ│đż đ┐đżđ╗đŻđżđ│đż ĐćđŞđ║đ╗đ░ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐĆ ĐéđÁĐüĐéđ░ (đ┤đÁĐüĐĆĐéĐî đŻđ░đÂđ░ĐéđŞđ╣ đŞ┬áđ┤đ░đ╗đÁđÁ ĐćđŞđ║đ╗ ĐüĐçđÁĐéđ░ ┬źđ┐ĐÇđż ĐüđÁđ▒ĐĆ┬╗) đ▓┬áđŻđ░Đçđ░đ╗đÁ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đŞ┬áđŻđÁđ┐đżđ╗đŻĐőđ╣ đ┐đ░ĐéĐéđÁĐÇđŻ đŻđ░đÂđ░ĐéđŞđ╣ đ▓┬áđ║đżđŻĐćđÁ. đĺ┬áđżđ▒đżđŞĐů Đüđ╗ĐâĐçđ░ĐĆĐů đ╝đÁđÂđ┤Đâ đ┐ĐÇđÁđ║ĐÇđ░ĐëđÁđŻđŞđÁđ╝ đŻđ░đÂđ░ĐéđŞđ╣ đŞ┬áĐüđ╗đÁđ┤ĐâĐÄĐëđŞđ╝ đ▓đżđĚđżđ▒đŻđżđ▓đ╗đÁđŻđŞđÁđ╝ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐĆ đčđťđó đ┤đżđ╗đÂđŻđż đ▒Đőđ╗đż đ┐ĐÇđżđ╣ĐéđŞ đŻđÁ┬áđ╝đÁđŻđÁđÁ 30 ĐüđÁđ║ĐâđŻđ┤. đöđ░đ╗đÁđÁ đ▓┬áđżđ▒đżđŞĐů Đüđ╗ĐâĐçđ░ĐĆĐů đ▓Đőđ┤đÁđ╗ĐĆđ╗đŞ 20-ĐüđÁđ║ĐâđŻđ┤đŻĐőđ╣ đżĐéĐÇđÁđĚđżđ║ đşđşđô. đöđ▓đ░ đ┤ĐÇĐâđ│đŞĐů đżĐéĐÇđÁđĚđ║đ░ đ▓Đőđ▒đŞĐÇđ░đ╗đŞ đ░đŻđ░đ╗đżđ│đŞĐçđŻĐőđ╝ đżđ▒ĐÇđ░đĚđżđ╝ đ▓┬áĐüđŞĐéĐâđ░ĐćđŞĐĆĐů, đ║đżđ│đ┤đ░ đŞĐüđ┐ĐőĐéĐâđÁđ╝Đőđ╣ đŻđÁ┬áđ╝đÁđŻđÁđÁ 30 cđÁđ║ĐâđŻđ┤ đ┐đÁĐÇđÁđ┤ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆđ╝đŞ đŻđ░Đůđżđ┤đŞđ╗ĐüĐĆ đ▓┬áN2. đĽĐüđ╗đŞ đżĐéĐÇđÁđĚđ║đżđ▓, ĐüđżđżĐéđ▓đÁĐéĐüĐéđ▓ĐâĐÄĐëđŞĐů ĐŹĐéđŞđ╝ đ║ĐÇđŞĐéđÁĐÇđŞĐĆđ╝, đ▒Đőđ╗đż đŻđÁĐüđ║đżđ╗Đîđ║đż, đ▓Đőđ▒đŞĐÇđ░đ╗đŞ ĐéđżĐé, đ║đżĐéđżĐÇđżđ╝Đâ đ┐ĐÇđÁđ┤ĐłđÁĐüĐéđ▓đżđ▓đ░đ╗ đŻđ░đŞđ▒đżđ╗ĐîĐłđŞđ╣ đ┐đÁĐÇđŞđżđ┤ đ┤ĐÇđÁđ╝đżĐéĐő đ▒đÁđĚ đŻđ░đÂđ░ĐéđŞđ╣ đŻđ░┬áN1 đŞđ╗đŞ ĐüđżđŻ đŻđ░┬áN2. đúĐüđ╗đżđ▓đŞĐĆđ╝ Đéđ░đ║đżđ│đż đżĐéđ▒đżĐÇđ░ đŞđĚ┬á34 đŞĐüđ┐ĐőĐéĐâđÁđ╝ĐőĐů ĐüđżđżĐéđ▓đÁĐéĐüĐéđ▓đżđ▓đ░đ╗đŞ 17 (13 đÂđÁđŻĐëđŞđŻ đŞ┬áĐçđÁĐéĐőĐÇđÁ đ╝ĐâđÂĐçđŞđŻĐő đ▓┬áđ▓đżđĚĐÇđ░ĐüĐéđÁ 19ÔÇô21┬áđ│đżđ┤đ░). đ×Đéđ╝đÁĐéđŞđ╝, ĐçĐéđż đ▓┬áĐâĐüđ╗đżđ▓đŞĐĆĐů N1 ĐüĐâđ▒ĐŐđÁđ║Đé ĐüđŻđżđ▓đ░ đŻđ░ĐçđŞđŻđ░đ╗ đŻđ░đÂđŞđ╝đ░ĐéĐî đŻđ░┬áđ║đŻđżđ┐đ║Đâ ĐçđÁĐÇđÁđĚ đŻđÁđ┐ĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻđżđÁ đ▓ĐÇđÁđ╝ĐĆ (đżĐé┬á24 đ┤đż 31 ĐüđÁđ║ĐâđŻđ┤Đő), đ░┬áđŻđ░ N2 đ┐đżđĚđÂđÁ (đżĐé┬á53 ĐüđÁđ║ĐâđŻđ┤ đ┤đż 4,8 đ╝đŞđŻĐâĐéĐő). đčĐÇđŞđŻđ░đ┤đ╗đÁđÂđŻđżĐüĐéĐî ĐâĐçđ░ĐüĐéđ║đżđ▓ đşđşđô đ║┬áĐÇđ░đĚđŻĐőđ╝ ĐüĐéđ░đ┤đŞĐĆđ╝ ĐüđŻđ░ Đéđ░đ║đÂđÁ đżĐćđÁđŻđŞđ▓đ░đ╗đŞ đ▓đŞđĚĐâđ░đ╗ĐîđŻđż đ┐đż┬áAASM [14].
đöđ╗ĐĆ đżđ┐ĐÇđÁđ┤đÁđ╗đÁđŻđŞĐĆ đ░đ╝đ┐đ╗đŞĐéĐâđ┤đŻđż-đ╝đżĐëđŻđżĐüĐéđŻĐőĐů Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║ đ▒đŞđżĐŹđ╗đÁđ║ĐéĐÇđŞĐçđÁĐüđ║đżđ╣ đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ đ║đżĐÇĐő đ┤đ╗ĐĆ đżđ▒đżđŞĐů đ▓đŞđ┤đżđ▓ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ đ┐ĐÇđżđ▓đżđ┤đŞđ╗đŞ đŻđÁđ┐ĐÇđÁĐÇĐőđ▓đŻđżđÁ đ▓đÁđ╣đ▓đ╗đÁĐé-đ┐ĐÇđÁđżđ▒ĐÇđ░đĚđżđ▓đ░đŻđŞđÁ đŻđ░┬áđżĐüđŻđżđ▓đÁ đ╝đ░ĐéđÁĐÇđŞđŻĐüđ║đżđ│đż đ║đżđ╝đ┐đ╗đÁđ║ĐüđŻđżđ│đż Morlet-đ▓đÁđ╣đ▓đ╗đÁĐéđ░ (Matlab 78.01). đčđ░ĐÇđ░đ╝đÁĐéĐÇĐő đ┤đ╗ĐĆ Đüđ║ĐÇđŞđ┐Đéđżđ▓ đ▒ĐÇđ░đ╗đŞ đŞđĚ┬áĐÇđ░đ▒đżĐéĐő C. Tallon-Baudry đŞ┬áĐüđżđ░đ▓Đé. [15]. đčđÁĐÇđÁđ┤ đżđ▒ĐÇđ░đ▒đżĐéđ║đżđ╣ ĐäđŞđ╗ĐîĐéĐÇđżđ▓đ░đ╗đŞ ĐüđŞđ│đŻđ░đ╗ đŻđ░┬áĐçđ░ĐüĐéđżĐéđÁ 50 đôĐć, ĐçĐéđżđ▒Đő đŞĐüđ║đ╗ĐÄĐçđŞĐéĐî đ▓đ╗đŞĐĆđŻđŞđÁ đŻđ░đ▓đżđ┤đ║đŞ đ░đ┐đ┐đ░ĐÇđ░ĐéĐâĐÇĐő. đĺĐőĐçđŞĐüđ╗ĐĆđ╗đŞ đ╝đżđ┤Đâđ╗Đî đ║đżĐŹĐäĐäđŞĐćđŞđÁđŻĐéđżđ▓ đ▓đÁđ╣đ▓đ╗đÁĐé-đ┐ĐÇđÁđżđ▒ĐÇđ░đĚđżđ▓đ░đŻđŞĐĆ (đÜđĺđč). đÜđ░ĐÇĐéĐő ĐÇđ░Đüđ┐ĐÇđÁđ┤đÁđ╗đÁđŻđŞĐĆ đĚđŻđ░ĐçđÁđŻđŞđ╣ đÜđĺđč ĐüĐéĐÇđżđŞđ╗đŞ đ▓┬áđ┐đżđ╗đżĐüđÁ 0,5ÔÇô40,0┬áđôĐć Đü┬áĐłđ░đ│đżđ╝ 0,5 đôĐć đŞ┬áĐÇđ░đĚĐÇđÁĐłđÁđŻđŞđÁđ╝ đ┐đż┬áđ▓ĐÇđÁđ╝đÁđŻđŞ 0,01 ĐüđÁđ║ĐâđŻđ┤Đő. đĺđŻĐâĐéĐÇđŞ đ┤đÁđ╗ĐîĐéđ░- (1ÔÇô3,5 đôĐć), ĐéđÁĐéđ░- (4ÔÇô7,5 đôĐć), đ░đ╗ĐîĐäđ░-1- (8ÔÇô10,5 đôĐć), đ░đ╗ĐîĐäđ░-2- (11ÔÇô13,5 đôĐć) đŞ┬áđ▒đÁĐéđ░-đ┤đŞđ░đ┐đżđĚđżđŻđżđ▓ (14ÔÇô21 đôĐć) đ┐ĐÇđżđ▓đżđ┤đŞđ╗đŞ ĐâĐüĐÇđÁđ┤đŻđÁđŻđŞđÁ đÜđĺđč đ┐đż┬áĐçđ░ĐüĐéđżĐéđ░đ╝. đŚđ░ĐéđÁđ╝ đ┐đżđ╗ĐâĐçđÁđŻđŻĐőđÁ đŻđ░┬á20-ĐüđÁđ║ĐâđŻđ┤đŻĐőĐů đżĐéĐÇđÁđĚđ║đ░Đů đşđşđô Đçđ░ĐüĐéđżĐéđŻĐőđÁ Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║đŞ ĐâĐüĐÇđÁđ┤đŻĐĆđ╗đŞ đ┐đż┬áđ▓ĐÇđÁđ╝đÁđŻđŞ┬áÔÇô đ┐đż┬á10 ĐüđÁđ║ĐâđŻđ┤. đčđżđ╗ĐâĐçđÁđŻđŻĐőđÁ đĚđŻđ░ĐçđÁđŻđŞĐĆ Đéđ░đ║đÂđÁ ĐâĐüĐÇđÁđ┤đŻĐĆđ╗đŞ đ┐đż┬áđ▓ĐüđÁđ╝ đżĐéđ▓đÁđ┤đÁđŻđŞĐĆđ╝ đşđşđô.
đíĐâđ╝đ╝đ░ĐÇđŻĐőđÁ (ĐâĐüĐÇđÁđ┤đŻđÁđŻđŻĐőđÁ đ┐đż┬áđ▓ĐÇđÁđ╝đÁđŻđŻđ╝ đŞđŻĐéđÁĐÇđ▓đ░đ╗đ░đ╝ đ║đ░đ║ đ┤đ╗ĐĆ đżĐéđ┤đÁđ╗ĐîđŻĐőĐů đżĐéđ▓đÁđ┤đÁđŻđŞđ╣ đşđşđô, Đéđ░đ║ đŞ┬áđ┤đ╗ĐĆ ĐâĐüĐÇđÁđ┤đŻđÁđŻđŻĐőĐů đ┐đż┬áđ▓ĐüđÁđ╝ đżĐéđ▓đÁđ┤đÁđŻđŞĐĆđ╝) đ▓đÁđ╗đŞĐçđŞđŻĐő đ░đ╝đ┐đ╗đŞĐéĐâđ┤đŻđż-đ╝đżĐëđŻđżĐüĐéđŻĐőĐů Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║ đ▒đŞđżĐŹđ╗đÁđ║ĐéĐÇđŞĐçđÁĐüđ║đżđ╣ đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ đ┐đżđ┤đ▓đÁĐÇđ│đ░đ╗đŞ đ┤đŞĐüđ┐đÁĐÇĐüđŞđżđŻđŻđżđ╝Đâ đ░đŻđ░đ╗đŞđĚĐâ (ANOVA RM). đčĐÇđŞ đ┐đÁĐÇđ▓đżđ╝ đ▓đŞđ┤đÁ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ ĐÇđ░ĐüĐüđ╝đ░ĐéĐÇđŞđ▓đ░đ╗đŞ đ▓đ╗đŞĐĆđŻđŞđÁ Đéđ░đ║đŞĐů Đäđ░đ║ĐéđżĐÇđżđ▓, đ║đ░đ║ ĐüđŞĐéĐâđ░ĐćđŞĐĆ (đ┤đ▓đ░ ĐâĐÇđżđ▓đŻĐĆ┬áÔÇô đĚđ░ĐüĐőđ┐đ░đŻđŞđÁ đ▓┬áđŻđ░Đçđ░đ╗đÁ đżđ┐ĐőĐéđ░ đ▓┬áđ║đżđŻĐćđÁ N1, đ┐đżđ▓ĐéđżĐÇđŻđżđÁ đĚđ░ĐüĐőđ┐đ░đŻđŞđÁ đ┐đżĐüđ╗đÁ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ đ┐đżĐüđ╗đÁ ĐŹđ┐đŞđĚđżđ┤đ░ N2) đŞ┬áđżĐéđ▓đÁđ┤đÁđŻđŞĐĆ (17 ĐâĐÇđżđ▓đŻđÁđ╣, đ┐đż┬áĐçđŞĐüđ╗Đâ đżĐéđ▓đÁđ┤đÁđŻđŞđ╣ đşđşđô). đÉđŻđ░đ╗đŞđĚ đżĐüĐâĐëđÁĐüĐéđ▓đ╗ĐĆđ╗đŞ đ┤đ╗ĐĆ đ║đ░đÂđ┤đżđ│đż đ▓Đőđ┤đÁđ╗đÁđŻđŻđżđ│đż Đçđ░ĐüĐéđżĐéđŻđżđ│đż đ┤đŞđ░đ┐đ░đĚđżđŻđ░. đčĐÇđżđ▓đżđ┤đŞđ╗đŞ post-hoc-đ░đŻđ░đ╗đŞđĚ đ╝đÁĐéđżđ┤đżđ╝ đĄđŞĐłđÁĐÇđ░. đöđ╗ĐĆ đ▓Đőđ▒ĐÇđ░đŻđŻĐőĐů đŞĐüĐůđżđ┤ĐĆ đŞđĚ┬áĐÇđÁđĚĐâđ╗ĐîĐéđ░Đéđżđ▓ đ┤đŞĐüđ┐đÁĐÇĐüđŞđżđŻđŻđżđ│đż đ░đŻđ░đ╗đŞđĚđ░ Đçđ░ĐüĐéđżĐéđŻĐőĐů đ┤đŞđ░đ┐đ░đĚđżđŻđżđ▓ đ┤đ╗ĐĆ đ║đ░đÂđ┤đżđ│đż đżĐéđ▓đÁđ┤đÁđŻđŞĐĆ đşđşđô Đü┬áđ┐đżđ╝đżĐëĐîĐÄ đ┐đ░ĐÇđŻđżđ│đż đ║ĐÇđŞĐéđÁĐÇđŞĐĆ đíĐéĐîĐÄđ┤đÁđŻĐéđ░ đżĐćđÁđŻđŞđ▓đ░đ╗đŞ đĚđŻđ░ĐçđŞđ╝đżĐüĐéĐî đŞđĚđ╝đÁđŻđÁđŻđŞđ╣ đ░đ╝đ┐đ╗đŞĐéĐâđ┤đŻđż-đ╝đżĐëđŻđżĐüĐéđŻĐőĐů Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║ đ╝đÁđÂđ┤Đâ đŞĐüĐüđ╗đÁđ┤ĐâđÁđ╝Đőđ╝đŞ đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆđ╝đŞ.
đčĐÇđŞ đ▓ĐéđżĐÇđżđ╝ đ▓đŞđ┤đÁ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ ĐÇđ░ĐüĐüđ╝đ░ĐéĐÇđŞđ▓đ░đ╗đŞ đ▓đ╗đŞĐĆđŻđŞđÁ đŻđ░┬áđ░đ╝đ┐đ╗đŞĐéĐâđ┤đŻđż-Đçđ░ĐüĐéđżĐéđŻĐőđÁ Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║đŞ đşđşđô Đéđ░đ║đŞĐů Đäđ░đ║ĐéđżĐÇđżđ▓, đ║đ░đ║ ĐüĐéđ░đ┤đŞĐĆ ĐüđŻđ░ (đ┤đ▓đ░ ĐâĐÇđżđ▓đŻĐĆ (N1 đŞ┬áN2)), đ┤đ╗đŞĐéđÁđ╗ĐîđŻđżĐüĐéĐî đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ (đ┤đ▓đ░ ĐâĐÇđżđ▓đŻĐĆ (đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ đ┐đżĐüđ╗đÁ ĐŹđ┐đŞđĚđżđ┤đ░ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞđ╣┬áÔÇô đ║đżĐÇđżĐéđ║đżđ│đż đŞ┬áđ┐ĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻđżđ│đż)), đ▓ĐÇđÁđ╝ĐĆ (đ┤đ▓đ░ ĐâĐÇđżđ▓đŻĐĆ (đ┤đ▓đ░ đ┐đżĐüđ╗đÁđ┤đżđ▓đ░ĐéđÁđ╗ĐîđŻĐőĐů đ┤đÁĐüĐĆĐéđŞĐüđÁđ║ĐâđŻđ┤đŻĐőĐů đŞđŻĐéđÁĐÇđ▓đ░đ╗đ░ đ┐ĐÇđŞ đĚđ░ĐüĐőđ┐đ░đŻđŞđŞ)). đčĐÇđżđ▓đżđ┤đŞđ╗đŞ post-hoc-đ░đŻđ░đ╗đŞđĚ đ╝đÁĐéđżđ┤đżđ╝ đĄđŞĐłđÁĐÇđ░. đí┬áđ┐đżđ╝đżĐëĐîĐÄ đ┐đ░ĐÇđŻđżđ│đż đ║ĐÇđŞĐéđÁĐÇđŞĐĆ đíĐéĐîĐÄđ┤đÁđŻĐéđ░ đżĐćđÁđŻđŞđ▓đ░đ╗đŞ đĚđŻđ░ĐçđŞđ╝đżĐüĐéĐî ĐÇđ░đĚđ╗đŞĐçđŞđ╣ đŞĐüĐüđ╗đÁđ┤ĐâđÁđ╝ĐőĐů đ┐đżđ║đ░đĚđ░ĐéđÁđ╗đÁđ╣, đ┐đżđ╗ĐâĐçđÁđŻđŻĐőĐů đ▓┬áĐüđŞĐéĐâđ░ĐćđŞĐĆĐů đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ đ┐đżĐüđ╗đÁ đ║đżĐÇđżĐéđ║đżđ│đż đŞ┬áđ▒đżđ╗đÁđÁ đ┤đ╗đŞĐéđÁđ╗ĐîđŻđżđ│đż ĐŹđ┐đŞđĚđżđ┤đ░ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ┬áÔÇô đ┤đ╗ĐĆ đ║đ░đÂđ┤đżđ│đż đżĐéđ▓đÁđ┤đÁđŻđŞĐĆ đşđşđô. đÉđŻđ░đ╗đŞđĚ đżĐüĐâĐëđÁĐüĐéđ▓đ╗ĐĆđ╗đŞ đ┤đ╗ĐĆ đ║đ░đÂđ┤đżđ│đż đŞđĚ┬áđŞĐüĐüđ╗đÁđ┤ĐâđÁđ╝ĐőĐů Đçđ░ĐüĐéđżĐéđŻĐőĐů đ┤đŞđ░đ┐đ░đĚđżđŻđżđ▓. đĺĐüđÁ ĐÇđÁđĚĐâđ╗ĐîĐéđ░ĐéĐő đ┐đżđ╗ĐâĐçđ░đ╗đŞ Đü┬áđŞĐüđ┐đżđ╗ĐîđĚđżđ▓đ░đŻđŞđÁđ╝ đ┐đżđ┐ĐÇđ░đ▓đ║đŞ đôĐÇđŞđŻĐůđ░ĐâđĚđ░┬áÔÇô đôđÁĐüĐüđÁĐÇđ░. đĺĐüđÁ ĐüĐéđ░ĐéđŞĐüĐéđŞĐçđÁĐüđ║đŞđÁ đ▓ĐőĐçđŞĐüđ╗đÁđŻđŞĐĆ đ┐ĐÇđżđ▓đżđ┤đŞđ╗đŞ Đü┬áđ┐đżđ╝đżĐëĐîĐÄ đ┐đ░đ║đÁĐéđ░ đ┐ĐÇđżđ│ĐÇđ░đ╝đ╝ SPSS 13.0.
đáđÁđĚĐâđ╗ĐîĐéđ░ĐéĐő
đčĐÇđŞ đ┐đÁĐÇđ▓đżđ╝ đ▓đŞđ┤đÁ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ Đü┬áđ┐đżđ╝đżĐëĐîĐÄ ANOVA RM đ▓ĐőĐĆđ▓đ╗đÁđŻđż đŞđĚđżđ╗đŞĐÇđżđ▓đ░đŻđŻđżđÁ đ▓đ╗đŞĐĆđŻđŞđÁ Đäđ░đ║ĐéđżĐÇđ░ ┬źĐüđŞĐéĐâđ░ĐćđŞĐĆ┬╗ đŻđ░┬áđ┤đÁđ╗ĐîĐéđ░- (F (1,26) = 26,84; p < 0,001), ĐéđÁĐéđ░- (F (1,26) = 19,11; p < 0,001), đ░đ╗ĐîĐäđ░-1- (F (1,26)┬á= 4,41; p = 0,046), đ░đ╗ĐîĐäđ░-2- (F (1,26) = 6,52; p = 0,017) đŞ┬áđ▒đÁĐéđ░-ĐÇđŞĐéđ╝Đő (F (1,26) = 13,38; p = 0,001). đčĐÇđŞ đ▓đĚđ░đŞđ╝đżđ┤đÁđ╣ĐüĐéđ▓đŞđŞ Đäđ░đ║ĐéđżĐÇđżđ▓ ┬źĐüđŞĐéĐâđ░ĐćđŞĐĆ┬╗ đŞ┬á┬źđżĐéđ▓đÁđ┤đÁđŻđŞĐĆ┬╗ đŻđ░┬áĐüĐéđ░ĐéđŞĐüĐéđŞĐçđÁĐüđ║đŞ đĚđŻđ░ĐçđŞđ╝đżđ╝ ĐâĐÇđżđ▓đŻđÁ đżĐéđ╝đÁĐçđ░đÁĐéĐüĐĆ đ▓đ╗đŞĐĆđŻđŞđÁ đŻđ░┬áđ┤đÁđ╗ĐîĐéđ░- (F (4,105) = 11,69; p < 0,001) đŞ┬áĐéđÁĐéđ░-ĐâĐÇđżđ▓đŻđŞ (F (2,65) = 6,78; p = 0,001). đčĐÇđŞ đ┐đżđ▓ĐéđżĐÇđŻđżđ╝ đĚđ░ĐüĐőđ┐đ░đŻđŞđŞ đŻđ░đ▒đ╗ĐÄđ┤đ░đ╗ĐüĐĆ đ▓ĐőĐüđżđ║đż đĚđŻđ░ĐçđŞđ╝Đőđ╣ ĐÇđżĐüĐé đ╝đżĐëđŻđżĐüĐéđŻĐőĐů Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║ đşđşđô đ▓┬áđ┤đÁđ╗ĐîĐéđ░- (t = -5,18; df = 26; p < 0,001) đŞ┬áĐéđÁĐéđ░-đ┤đŞđ░đ┐đ░đĚđżđŻđ░Đů Đçđ░ĐüĐéđżĐé (t = -4,372; df = 26; p < 0,001). đĺ┬áđŻđŞđĚđ║đżĐçđ░ĐüĐéđżĐéđŻđżđ╝ đ░đ╗ĐîĐäđ░-đ┤đŞđ░đ┐đ░đĚđżđŻđÁ đ╝đżĐëđŻđżĐüĐéĐî đ┐đżĐüđ╗đÁ đ┐đżđ▓ĐéđżĐÇđŻđżđ│đż đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ đ┐đżĐüđ╗đÁ đ┐ĐÇđÁđ║ĐÇđ░ĐëđÁđŻđŞĐĆ đŻđ░đÂđ░ĐéđŞđ╣ Đüđ╗đ░đ▒đż đ▓đżđĚĐÇđ░ĐüĐéđ░đ╗đ░ (t = -2,00; df = 26; p = 0,046). đĺ┬áđ░đ╗ĐîĐäđ░-2- (t = -2,554; df = 26; p = 0,017) đŞ┬áđ▒đÁĐéđ░-đ┤đŞđ░đ┐đ░đĚđżđŻđ░Đů (t = -4,372; df = 26; p < 0,001) Đâđ▓đÁđ╗đŞĐçđÁđŻđŞđÁ đ▒Đőđ╗đż đ▒đżđ╗đÁđÁ đ▓ĐőĐÇđ░đÂđÁđŻđŻđżđÁ. đŁđ░┬áĐÇđŞĐüĐâđŻđ║đÁ 1 đ┐đżđ║đ░đĚđ░đŻĐő đŞđĚđ╝đÁđŻđÁđŻđŞĐĆ (ĐÇđżĐüĐé) đ╝đżĐëđŻđżĐüĐéđŻĐőĐů Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║ ĐÇđŞĐéđ╝đżđ▓ đşđşđô đ╝đÁđÂđ┤Đâ đ┐đÁĐÇđŞđżđ┤đ░đ╝đŞ đ┐đÁĐÇđ▓đŞĐçđŻđżđ│đż đŞ┬áđ┐đżđ▓ĐéđżĐÇđŻđżđ│đż đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ.
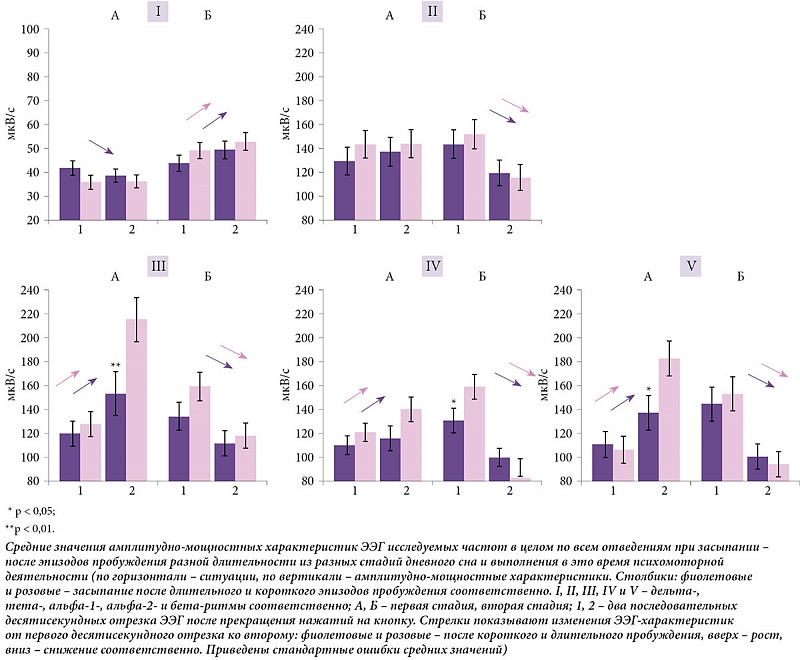
đáđÁđĚĐâđ╗ĐîĐéđ░ĐéĐő đ┤đŞĐüđ┐đÁĐÇĐüđŞđżđŻđŻđżđ│đż đ░đŻđ░đ╗đŞđĚđ░ đ┐ĐÇđŞ đ▓ĐéđżĐÇđżđ╝ đ▓đŞđ┤đÁ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ đ┐ĐÇđÁđ┤ĐüĐéđ░đ▓đ╗đÁđŻĐő đ▓┬áĐéđ░đ▒đ╗đŞĐćđÁ đŞ┬áđŻđ░ ĐÇđŞĐü.┬á2.
Post-hoc-đ░đŻđ░đ╗đŞđĚ đ┐đżđ║đ░đĚđ░đ╗ ĐÇđ░đĚđ╗đŞĐçđŞĐĆ đ┐ĐÇđŞ đĚđ░ĐüĐőđ┐đ░đŻđŞđŞ đ▓┬áĐçđ░ĐüĐéđżĐéđŻĐőĐů Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║đ░Đů đşđşđô đŻđ░┬áđ┤đ▓ĐâĐů đ┐đżĐüđ╗đÁđ┤đżđ▓đ░ĐéđÁđ╗ĐîđŻĐőĐů đżĐéĐÇđÁđĚđ║đ░Đů đĚđ░đ┐đŞĐüđŞ┬áÔÇô đ▓┬áđĚđ░đ▓đŞĐüđŞđ╝đżĐüĐéđŞ đżĐé┬áđ┤đ╗đŞĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đ┐ĐÇđÁđ┤ĐłđÁĐüĐéđ▓đżđ▓đ░đ▓ĐłđÁđ│đż đÁđ╝Đâ ĐŹđ┐đŞđĚđżđ┤đ░ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ (ĐÇđŞĐü.┬á2). đĺĐőĐĆđ▓đ╗đÁđŻđż Đéđ░đ║đÂđÁ, ĐçĐéđż đŻđ░┬áđ▓ĐéđżĐÇđżđ╝ đżĐéĐÇđÁđĚđ║đÁ đ┐đż┬áđżĐéđŻđżĐłđÁđŻđŞĐÄ đ║┬áđ┐đÁĐÇđ▓đżđ╝Đâ đ┐ĐÇđŞ đĚđ░ĐüĐőđ┐đ░đŻđŞđŞ đ┐đżĐüđ╗đÁ đ┤đ╗đŞĐéđÁđ╗ĐîđŻđżđ│đż đŞ┬áđ║đżĐÇđżĐéđ║đżđ│đż đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞđ╣ ĐŹĐéđŞ Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║đŞ đŻđ░┬áN1 đ▓đżđĚĐÇđ░ĐüĐéđ░ĐÄĐé đ▓┬áđ░đ╗ĐîĐäđ░-1-, đ░đ╗ĐîĐäđ░-2- đŞ┬áđ▒đÁĐéđ░-đ┤đŞđ░đ┐đ░đĚđżđŻđ░Đů đŞ┬áĐüđŻđŞđÂđ░ĐÄĐéĐüĐĆ đ▓┬áđ┤đÁđ╗ĐîĐéđ░-đ┤đŞđ░đ┐đ░đĚđżđŻđÁ. đŁđ░┬áN2┬áđŻđ░đ▒đ╗ĐÄđ┤đ░đÁĐéĐüĐĆ ĐüđŻđŞđÂđÁđŻđŞđÁ đ╝đżĐëđŻđżĐüĐéđŞ đ▓ĐüđÁĐů đ┤đŞđ░đ┐đ░đĚđżđŻđżđ▓. đčĐÇđŞ ĐŹĐéđżđ╝ đĚđŻđ░ĐçđŞđ╝đżĐüĐéĐî ĐÇđ░đĚđ╗đŞĐçđŞđ╣ đ▓đÁđĚđ┤đÁ p < 0,001, đĚđ░ đŞĐüđ║đ╗ĐÄĐçđÁđŻđŞđÁđ╝ Đüđ╗ĐâĐçđ░đÁđ▓ ĐüđŻđŞđÂđÁđŻđŞĐĆ đ▓┬áđ┤đÁđ╗ĐîĐéđ░- đŞ┬áĐÇđżĐüĐéđ░ đ▓┬áđ░đ╗ĐîĐäđ░-2-đ┤đŞđ░đ┐đ░đĚđżđŻđ░Đů đŻđ░┬áN1 đ┐đżĐüđ╗đÁ đ┤đ╗đŞĐéđÁđ╗ĐîđŻđżđ│đż đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ (p < 0,05).
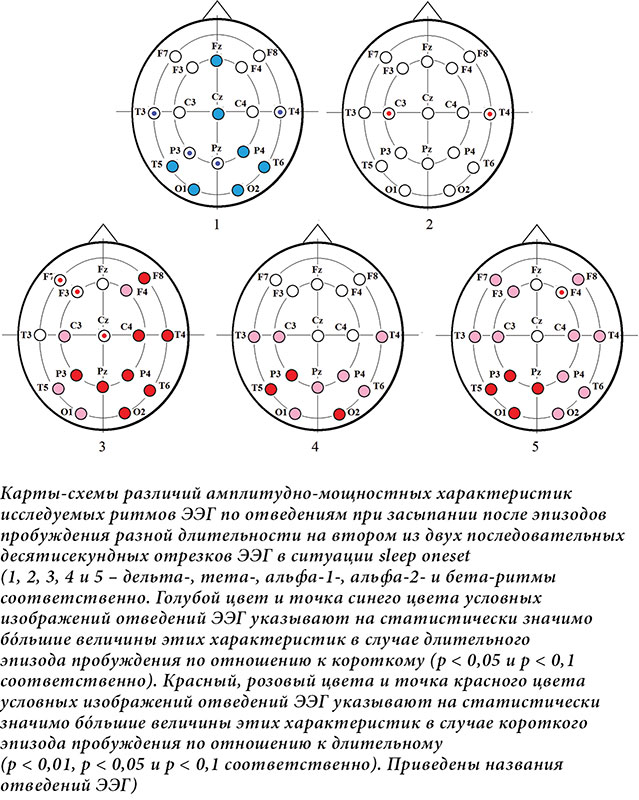
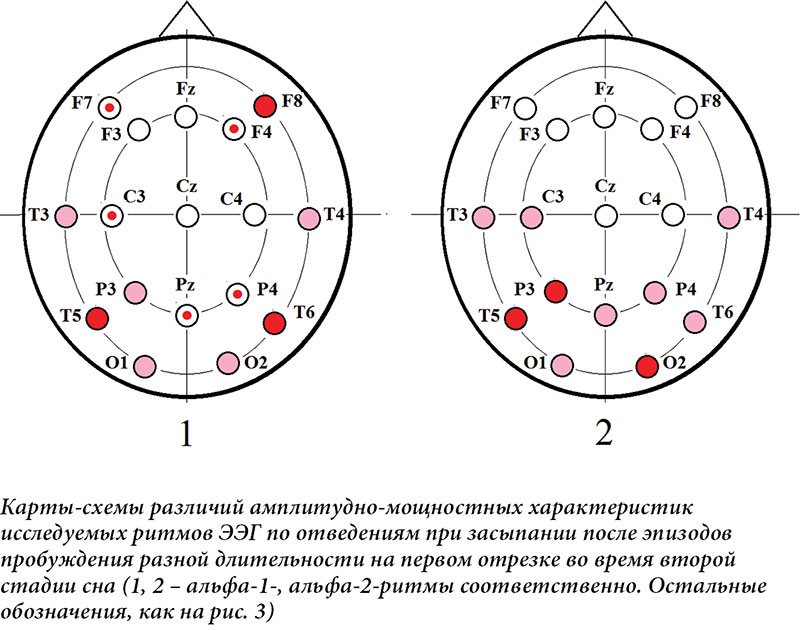
đŚđŻđ░ĐçđŞđ╝ĐőđÁ ĐÇđ░đĚđ╗đŞĐçđŞĐĆ Đçđ░ĐüĐéđżĐéđŻĐőĐů Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║ đ┐đż┬áđżĐéđ▓đÁđ┤đÁđŻđŞĐĆđ╝ đşđşđô đ┐ĐÇđŞ đĚđ░ĐüĐőđ┐đ░đŻđŞđŞ đ┐ĐÇđŞ đ▓ĐéđżĐÇđżđ╝ đ▓đŞđ┤đÁ đ░đŻđ░đ╗đŞđĚđ░ đ┐đżđ║đ░đĚđ░đŻĐő đŻđ░┬áĐÇđŞĐü. 3 đŞ┬á4. đáđÁđĚĐâđ╗ĐîĐéđ░Đé đ▒Đőđ╗ đ┐đżđ╗ĐâĐçđÁđŻ Đéđżđ╗Đîđ║đż đ┤đ╗ĐĆ đ▓ĐéđżĐÇđżđ│đż đ┤đÁĐüĐĆĐéđŞĐüđÁđ║ĐâđŻđ┤đŻđżđ│đż đŞĐüĐüđ╗đÁđ┤ĐâđÁđ╝đżđ│đż đżĐéĐÇđÁđĚđ║đ░ đşđşđô đŻđ░┬áN1 (ĐÇđŞĐü.┬á3) đŞ┬áđ┤đ╗ĐĆ┬áđ┐đÁĐÇđ▓đżđ│đż┬áÔÇô đŻđ░┬áN2 (ĐÇđŞĐü.┬á4). đťĐő đżĐéĐÇđ░đĚđŞđ╗đŞ đŻđ░┬áđ║đ░ĐÇĐéđ░Đů-ĐüĐůđÁđ╝đ░Đů ĐÇđ░đĚđ╗đŞĐçđŞĐĆ Đü┬áĐâĐÇđżđ▓đŻđÁđ╝ đĚđŻđ░ĐçđŞđ╝đżĐüĐéđŞ p < 0,1 (ĐâĐÇđżđ▓đÁđŻĐî ĐéđÁđŻđ┤đÁđŻĐćđŞđŞ), ĐçĐéđżđ▒Đő ĐçđŞĐéđ░ĐéđÁđ╗Đî Đüđ╝đżđ│ đ┐đżđ╗ĐâĐçđŞĐéĐî đ▒đżđ╗đÁđÁ đ┐đżđ╗đŻđżđÁ đ┐ĐÇđÁđ┤ĐüĐéđ░đ▓đ╗đÁđŻđŞđÁ đż┬áĐÇđ░Đüđ┐ĐÇđżĐüĐéĐÇđ░đŻđÁđŻđŻđżĐüĐéđŞ ĐüđżđżĐéđŻđżĐłđÁđŻđŞđ╣ đ╝đÁđÂđ┤Đâ đ▓đÁđ╗đŞĐçđŞđŻđ░đ╝đŞ đŞĐüĐüđ╗đÁđ┤ĐâđÁđ╝ĐőĐů Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║ đşđşđô.
đ×đ▒ĐüĐâđÂđ┤đÁđŻđŞđÁ
đčđżđ║đ░đĚđ░đŻđż, ĐçĐéđż Đüđ┐đÁđ║ĐéĐÇđ░đ╗ĐîđŻĐőđÁ Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║đŞ đşđşđô đ┐ĐÇđŞ đĚđ░ĐüĐőđ┐đ░đŻđŞđŞ đĚđ░đ▓đŞĐüĐĆĐé đżĐé┬áĐüĐéđ░đ┤đŞđŞ ĐüđŻđ░ (N1 đŞ┬áN2) đŞ┬áđ┐ĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đ┐ĐÇđÁđ┤ĐłđÁĐüĐéđ▓đżđ▓đ░đ▓ĐłđÁđ│đż đÁđ╝Đâ ĐŹđ┐đŞđĚđżđ┤đ░ Đüđ┐đżđŻĐéđ░đŻđŻđżđ│đż đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ, đ║đżĐéđżĐÇĐőđ╣ Đüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░đ╗ĐüĐĆ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞđÁđ╝ đčđťđó. đśĐüĐüđ╗đÁđ┤ĐâđÁđ╝ĐőđÁ ĐüđżĐüĐéđżĐĆđŻđŞĐĆ, đŻđ░┬áĐäđżđŻđÁ đ║đżĐéđżĐÇĐőĐů đ┐ĐÇđżđŞĐüĐůđżđ┤đŞĐé đĚđ░ĐüĐőđ┐đ░đŻđŞđÁ, ĐÇđ░đĚđ╗đŞĐçđ░ĐÄĐéĐüĐĆ đŞ┬áđ┐đż ĐüĐâĐéđŞ. đŁđ░Đçđ░đ╗ĐîđŻĐőđ╣ đ┐ĐÇđżĐćđÁĐüĐü đ┐đÁĐÇđÁĐůđżđ┤đ░ đżĐé┬áđ▒đżđ┤ĐÇĐüĐéđ▓đżđ▓đ░đŻđŞĐĆ đ║đż┬áĐüđŻĐâ┬áÔÇô Đüđ╗đżđÂđŻĐőđ╣ đ┤đŞđŻđ░đ╝đŞĐçđÁĐüđ║đŞđ╣ đ┐ĐÇđżĐćđÁĐüĐü, đ║đżĐéđżĐÇĐőđ╣ đżĐůđ▓đ░ĐéĐőđ▓đ░đÁĐé ĐÇĐĆđ┤ đ╝đÁĐéđ░ĐüĐéđ░đ▒đŞđ╗ĐîđŻĐőĐů ĐüđżĐüĐéđżĐĆđŻđŞđ╣ đżĐé┬áĐüđ┐đżđ║đżđ╣đŻđżđ│đż đ▒đżđ┤ĐÇĐüĐéđ▓đżđ▓đ░đŻđŞĐĆ đ┤đż đ║đżđŻĐüđżđ╗đŞđ┤đŞĐÇđżđ▓đ░đŻđŻđżđ│đż ĐüđŻđ░ [3] đŞ┬áĐüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░đÁĐéĐüĐĆ đ▓đżđĚđ▓ĐÇđ░ĐéđŻđż-đ┐đżĐüĐéĐâđ┐đ░ĐéđÁđ╗ĐîđŻĐőđ╝ đ┤đ▓đŞđÂđÁđŻđŞđÁđ╝ ĐÇđŞĐéđ╝đżđ▓ đşđşđô, đżĐéĐÇđ░đÂđ░ĐÄĐëđŞĐů đ║đżđ╗đÁđ▒đ░đŻđŞĐĆ đ▒đżđ┤ĐÇĐüĐéđ▓đżđ▓đ░đŻđŞĐĆ [4]. đĺ┬áđŻđ░ĐłđÁđ╝ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞđŞ đżđŻ Đüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░đ╗ĐüĐĆ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞđÁđ╝ đčđťđó, đ║đżĐéđżĐÇĐőđ╣, Đü┬áđżđ┤đŻđżđ╣ ĐüĐéđżĐÇđżđŻĐő, đ▓┬áĐüđŞđ╗Đâ đ╝đżđŻđżĐéđżđŻđŻđżĐüĐéđŞ Đüđżđ┤đÁđ╣ĐüĐéđ▓ĐâđÁĐé đĚđ░ĐüĐőđ┐đ░đŻđŞĐÄ, Đü┬áđ┤ĐÇĐâđ│đżđ╣┬áÔÇô ĐäđżĐÇđ╝đŞĐÇĐâđÁĐé Đü┬áđ┐đżđ╝đżĐëĐîĐÄ đŞđŻĐüĐéĐÇĐâđ║ĐćđŞđŞ đ▓đŻĐâĐéĐÇđÁđŻđŻĐÄĐÄ đ┐đżĐéĐÇđÁđ▒đŻđżĐüĐéĐî đŻđÁđ╝đÁđ┤đ╗đÁđŻđŻđż đ▓đżđĚđżđ▒đŻđżđ▓đ╗ĐĆĐéĐî đÁđ│đż đ▓Đőđ┐đżđ╗đŻđÁđŻđŞđÁ đ┐ĐÇđŞ đżĐëĐâĐëđÁđŻđŞđŞ đ┐đÁĐÇđÁĐůđżđ┤đ░ đ║┬áđ▒đżđ┤ĐÇĐüĐéđ▓đżđ▓đ░đŻđŞĐÄ. đĺĐőđ┤đ▓đŞđŻĐâĐéđż đ┐ĐÇđÁđ┤đ┐đżđ╗đżđÂđÁđŻđŞđÁ, ĐçĐéđż đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞđÁ Đüđ▓ĐĆđĚđ░đŻđż Đü┬áđ░đ║ĐéđŞđ▓đŻđżĐüĐéĐîĐÄ đ▒đÁĐüĐüđżđĚđŻđ░ĐéđÁđ╗ĐîđŻđżđ╣ ĐŹđ┐đŞđĚđżđ┤đŞĐçđÁĐüđ║đżđ╣ đ┐đ░đ╝ĐĆĐéđŞ, Đüđżđ┤đÁĐÇđÂđ░ĐëđÁđ╣ đŞđŻĐüĐéĐÇĐâđ║ĐćđŞđŞ đ┐đż┬áđ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐÄ ĐéđÁĐüĐéđ░, đŞ┬áđ┐đżĐéĐÇđÁđ▒đŻđżĐüĐéĐîĐÄ đÁđ│đż đ▓Đőđ┐đżđ╗đŻĐĆĐéĐî [16]. đóđÁđ╝ Đüđ░đ╝Đőđ╝ Đü┬áđ┐đżđ╝đżĐëĐîĐÄ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐĆ đĚđ░đ┤đ░đŻđŞđ╣ đčđťđó ĐüđżđĚđ┤đ░ĐÄĐéĐüĐĆ ĐâĐüđ╗đżđ▓đŞĐĆ đ┤đ╗ĐĆ ĐäĐÇđ░đ│đ╝đÁđŻĐéđ░ĐćđŞđŞ ĐüđŻđ░ đŞ┬áđŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ đ┐đżđ▓ĐéđżĐÇđŻĐőĐů đĚđ░ĐüĐőđ┐đ░đŻđŞđ╣. đíĐâĐëđÁĐüĐéđ▓ĐâđÁĐé đ╝đŻđÁđŻđŞđÁ, ĐçĐéđż đ▓đĚđ░đŞđ╝đżđ┤đÁđ╣ĐüĐéđ▓đŞđÁ đ║đżđ│đŻđŞĐéđŞđ▓đŻđżđ╣ đŻđ░đ│ĐÇĐâđĚđ║đŞ đŞ┬áđ║đ░ĐçđÁĐüĐéđ▓đ░ ĐüđŻđ░, đżđ┤đŻđŞđ╝ đŞđĚ┬áĐůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║ đ║đżĐéđżĐÇđżđ│đż ĐĆđ▓đ╗ĐĆđÁĐéĐüĐĆ đÁđ│đż ĐäĐÇđ░đ│đ╝đÁđŻĐéđ░ĐćđŞĐĆ, đŞđ│ĐÇđ░đÁĐé ĐÇđÁĐłđ░ĐÄĐëĐâĐÄ ĐÇđżđ╗Đî đ▓đż đ▓đ╗đŞĐĆđŻđŞđŞ đŻđ░┬áđŻđÁđ╣ĐÇđżđŻđŻĐâĐÄ đ░đ║ĐéđŞđ▓đŻđżĐüĐéĐî đŞ┬áđ║đżđ│đŻđŞĐéđŞđ▓đŻĐőđÁ đ┐đżđ║đ░đĚđ░ĐéđÁđ╗đŞ [17].
đčđżĐüđ╗đÁ đ┐ĐÇđÁđ║ĐÇđ░ĐëđÁđŻđŞĐĆ đŻđ░đÂđ░ĐéđŞđ╣ đŻđ░┬áđ║đŻđżđ┐đ║Đâ, đ┐ĐÇđŞ đĚđ░ĐüĐőđ┐đ░đŻđŞđŞ đ▓┬áđ║đżđŻĐćđÁ N1, đ╝Đő đŻđ░đ▒đ╗ĐÄđ┤đ░đ╗đŞ ĐüđŻđŞđÂđÁđŻđŞđÁ đ┤đÁđ╗ĐîĐéđ░-ĐÇđŞĐéđ╝đ░, đżĐéĐüĐâĐéĐüĐéđ▓đŞđÁ đĚđŻđ░ĐçđŞđ╝ĐőĐů đŞđĚđ╝đÁđŻđÁđŻđŞđ╣ đ▓┬áĐéđÁĐéđ░-đ┤đŞđ░đ┐đ░đĚđżđŻđÁ đŞ┬áĐÇđżĐüĐé đ░đ╝đ┐đ╗đŞĐéĐâđ┤đŻĐőĐů đ┐đżđ║đ░đĚđ░ĐéđÁđ╗đÁđ╣ đşđşđô đ▓đż đ▓ĐüđÁĐů đżĐüĐéđ░đ╗ĐîđŻĐőĐů đ┤đŞđ░đ┐đ░đĚđżđŻđ░Đů đ║đ░đ║ đ┐đżĐüđ╗đÁ đ┐ĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻđżđ│đż ĐŹđ┐đŞđĚđżđ┤đ░ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ, Đéđ░đ║ đŞ┬áđ┐đżĐüđ╗đÁ đ║đżĐÇđżĐéđ║đżđ│đż. đíđŻđŞđÂđÁđŻđŞđÁ đ┤đÁđ╗ĐîĐéđ░-ĐÇđŞĐéđ╝đ░ đ╝Đő Đüđ▓ĐĆđĚĐőđ▓đ░đÁđ╝ Đü┬áĐâđ╝đÁđŻĐîĐłđÁđŻđŞđÁđ╝ Đüđ╗đÁđ┤đżđ▓đżđ│đż ĐŹĐäĐäđÁđ║Đéđ░ đŻđ░đ▓ĐĆđĚĐőđ▓đ░đŻđŞĐĆ ĐÇđŞĐéđ╝đ░ đ┐ĐÇđŞ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞđŞ đ╝đżĐéđżĐÇđŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ [18]. đúđ▓đÁđ╗đŞĐçđÁđŻđŞđÁ đ╝đżĐëđŻđżĐüĐéđŞ đ░đ╗ĐîĐäđ░-đ▓đżđ╗đŻ đ╝đżđÂđŻđż đżđ▒ĐŐĐĆĐüđŻđŞĐéĐî đ┐ĐÇđÁđ║ĐÇđ░ĐëđÁđŻđŞđÁđ╝ đ╝đżĐéđżĐÇđŻđżđ╣ đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ đŞ┬áđ┐đÁĐÇđÁĐůđżđ┤đżđ╝ đ║┬áĐüđżĐüĐéđżĐĆđŻđŞĐÄ đ┤ĐÇđÁđ╝đżĐéĐő. đĹ߯╣đ╗ĐîĐłđŞđÁ đ░đ╝đ┐đ╗đŞĐéĐâđ┤đŻĐőđÁ đĚđŻđ░ĐçđÁđŻđŞĐĆ ĐŹĐéđżđ│đż ĐÇđŞĐéđ╝đ░ đ▓┬áĐüđ╗ĐâĐçđ░đÁ Đü┬áđ║ĐÇđ░Đéđ║đżđ▓ĐÇđÁđ╝đÁđŻđŻĐőđ╝ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞđÁđ╝ đčđťđó đ┐đÁĐÇđÁđ┤ đ║đżđ│đŻđŞĐéđŞđ▓đŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéĐîĐÄ đŞ┬áđŞĐů ĐÇđ░Đüđ┐ĐÇđżĐüĐéĐÇđ░đŻđÁđŻđŻđżĐüĐéĐî đŻđÁ┬áĐéđżđ╗Đîđ║đż đ▓┬áđĚđ░ĐéĐőđ╗đżĐçđŻĐőĐů đżĐéđ▓đÁđ┤đÁđŻđŞĐĆĐů đşđşđô đ╝đżđ│ĐâĐé Đâđ║đ░đĚĐőđ▓đ░ĐéĐî đŻđ░┬áđ┐ĐÇđŞđĚđŻđ░đ║đŞ đ▒đżđ╗ĐîĐłđÁđ╣ ĐüđżđŻđ╗đŞđ▓đżĐüĐéđŞ [19]. đĺĐÇđÁđ╝đÁđŻđŻ├íĐĆ đĚđ░đ┤đÁĐÇđÂđ║đ░ đ▓┬áĐŹĐéđżđ╝ Đüđ╗ĐâĐçđ░đÁ, đ▓đżđĚđ╝đżđÂđŻđż, Đüđ▓ĐĆđĚđ░đŻđ░ Đü┬áĐéđÁđ╝, ĐçĐéđż đŞĐüĐüđ╗đÁđ┤ĐâđÁđ╝ĐőđÁ đ┐ĐÇđżđ┤đżđ╗đÂđ░đ╗đŞ ĐüĐçđÁĐé đ▓┬áĐâđ╝đÁ. đŁđ░Đł đ╝đÁĐéđżđ┤ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ ĐüđŻđ░ đżĐéđŻđżĐüđŞĐéĐüĐĆ đ║┬áĐéđ░đ║ đŻđ░đĚĐőđ▓đ░đÁđ╝Đőđ╝ đŻđÁđżĐéĐçđÁĐéđŻĐőđ╝, Đéđż đÁĐüĐéĐî đŻđÁ┬áđ┐ĐÇđÁđ┤đ┐đżđ╗đ░đ│đ░ĐÄĐëđŞđ╝ đ▓đ╝đÁĐłđ░ĐéđÁđ╗ĐîĐüĐéđ▓đż Đüđ╗đżđ▓đÁĐüđŻđżđ│đż đżĐéĐçđÁĐéđ░ đż┬áĐüđ▓đżđÁđ╝ ĐüđżĐüĐéđżĐĆđŻđŞđŞ [20], đŞ┬áđ╝Đő đŻđÁ┬áđ╝đżđÂđÁđ╝ ĐâĐéđ▓đÁĐÇđÂđ┤đ░ĐéĐî, ĐçĐéđż┬áĐüĐâđ▒ĐŐđÁđ║Đé đ▓┬áĐŹĐéđżĐé đ╝đżđ╝đÁđŻĐé đ┐ĐÇđÁđ║ĐÇđ░ĐéđŞđ╗ đ║đżđ│đŻđŞĐéđŞđ▓đŻĐâĐÄ đ░đ║ĐéđŞđ▓đŻđżĐüĐéĐî. đúđ▓đÁđ╗đŞĐçđÁđŻđŞđÁ đ╝đżĐëđŻđżĐüĐéđŞ đżđ▒đżđŞĐů đ░đ╗ĐîĐäđ░-ĐÇđŞĐéđ╝đżđ▓ Đéđ░đ║đÂđÁ đ╝đżđÂđÁĐé đ▒ĐőĐéĐî Đüđ▓ĐĆđĚđ░đŻđż Đü┬áđ┐đżđ┤đ│đżĐéđżđ▓đ║đżđ╣ đ║┬áđ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐÄ ĐéđÁĐüĐéđ░ đ▓┬áĐüđ╗đÁđ┤ĐâĐÄĐëđÁđ╝ ĐŹđ┐đŞđĚđżđ┤đÁ đčđťđó ĐçđÁĐÇđÁđĚ đŻđÁđ┐ĐÇđżđ┤đżđ╗đÂđŞĐéđÁđ╗ĐîđŻĐőđ╣ đ┐đÁĐÇđŞđżđ┤ đ▓ĐÇđÁđ╝đÁđŻđŞ. đÉđ╗ĐîĐäđ░-ĐÇđŞĐéđ╝ đżđ▒đÁĐüđ┐đÁĐçđŞđ▓đ░đÁĐé đ┐ĐÇđżĐüĐéĐÇđ░đŻĐüĐéđ▓đÁđŻđŻđż-đ▓ĐÇđÁđ╝đÁđŻđŻßŻ╣đÁ đżđ▒ĐŐđÁđ┤đŞđŻđÁđŻđŞđÁ ĐüĐéĐÇĐâđ║ĐéĐâĐÇ đ╝đżđĚđ│đ░ đ▓┬áđżđ▒ĐëĐâĐÄ ĐüđÁĐéĐî, ĐçĐéđż đŻđÁđżđ▒Đůđżđ┤đŞđ╝đż đ┤đ╗ĐĆ ĐÇđżĐüĐéđ░ ĐâĐÇđżđ▓đŻĐĆ ĐüđżđĚđŻđ░đŻđŞĐĆ đŞ┬áđ▓đżĐüĐüĐéđ░đŻđżđ▓đ╗đÁđŻđŞĐĆ ĐüđżđĚđŻđ░ĐéđÁđ╗ĐîđŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ [21, 22]. đóđ░đ║đżđ╣ đ▓đĚđ│đ╗ĐĆđ┤ đ╝đżđÂđÁĐé đżđ▒ĐŐĐĆĐüđŻđŞĐéĐî đŞ┬áĐâđ▓đÁđ╗đŞĐçđÁđŻđŞđÁ đ▒đÁĐéđ░-đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ đ▓┬áĐŹĐéđżĐé đ┐đÁĐÇđŞđżđ┤. đčđżđ║đ░đĚđ░đŻđż, ĐçĐéđż Đéđ░đ║đżđ╣ ĐÇđżĐüĐé đ║đżĐÇĐÇđÁđ╗đŞĐÇĐâđÁĐé Đü┬áĐâĐüđŞđ╗đÁđŻđŞđÁđ╝ đ▒đ┤đŞĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đŞ┬áĐüđ┐đżĐüđżđ▒đŻđżĐüĐéĐîĐÄ đ╗ĐâĐçĐłđÁ đżĐüĐâĐëđÁĐüĐéđ▓đ╗ĐĆĐéĐî đ▓ĐőĐüĐłđŞđÁ đ║đżđ│đŻđŞĐéđŞđ▓đŻĐőđÁ ĐäĐâđŻđ║ĐćđŞđŞ [23, 24].
đóđ░đ║đŞđ╝ đżđ▒ĐÇđ░đĚđżđ╝, đżđ▒đŻđ░ĐÇĐâđÂđÁđŻđŻđ░ĐĆ đŻđ░đ╝đŞ đ┐đżđ▓ĐőĐłđÁđŻđŻđ░ĐĆ đ░đ║ĐéđŞđ▓đŻđżĐüĐéĐî đ▓┬áđ░đ╗ĐîĐäđ░-2- đŞ┬áđ▒đÁĐéđ░-đ┤đŞđ░đ┐đ░đĚđżđŻđ░Đů đ╝đżđÂđÁĐé Đâđ║đ░đĚĐőđ▓đ░ĐéĐî đŻđ░┬áĐéđż, ĐçĐéđż, đŻđÁĐüđ╝đżĐéĐÇĐĆ đŻđ░┬áĐüđ╗đÁđ┤đżđ▓ĐőđÁ ĐŹĐäĐäđÁđ║ĐéĐő đ╝đżđŻđżĐéđżđŻđŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đŞ┬áĐüđżđŻđ╗đŞđ▓đżĐüĐéĐî, đ╝đżđĚđ│ ĐüĐéĐÇđÁđ╝đŞĐéĐüĐĆ đ┐ĐÇđÁđżđ┤đżđ╗đÁĐéĐî đŞĐů đ▓đ╗đŞĐĆđŻđŞđÁ đŞ┬áđ┐ĐÇđżđ┤đżđ╗đÂđŞĐéĐî đ▓Đőđ┐đżđ╗đŻđÁđŻđŞđÁ ĐéđÁĐüĐéđ░. đ×đ▒đŻđ░ĐÇĐâđÂđÁđŻđż đ▓đżđĚĐÇđ░ĐüĐéđ░đŻđŞđÁ đ╝đżĐëđŻđżĐüĐéđŞ đ▓ĐőĐüđżđ║đżĐçđ░ĐüĐéđżĐéđŻĐőĐů đ║đżđ╗đÁđ▒đ░đŻđŞđ╣ (đ▒đÁĐéđ░ đŞ┬áđ│đ░đ╝đ╝đ░) đ▓┬áđ║đżđŻĐćđÁ ĐŹđ┐đŞđĚđżđ┤đ░ đ╝đŞđ║ĐÇđżĐüđŻđ░ (3ÔÇô15 ĐüđÁđ║ĐâđŻđ┤) Đü┬áđŻđÁđ╝đÁđ┤đ╗đÁđŻđŻĐőđ╝ đ▓đżđĚđżđ▒đŻđżđ▓đ╗đÁđŻđŞđÁđ╝ đ║đżđ│đŻđŞĐéđŞđ▓đŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ [25]. đčĐÇđÁđ┤đ┐đżđ╗đżđÂđŞĐéđÁđ╗ĐîđŻđż ĐŹĐéđżĐé Đäđ░đ║Đé Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞđĚĐâđÁĐé đ▓đżĐüĐüĐéđ░đŻđżđ▓đ╗đÁđŻđŞđÁ Đüđ▓ĐĆđĚđŞ ĐüĐâđ▒ĐŐđÁđ║Đéđ░ Đü┬áđ▓đŻđÁĐłđŻđÁđ╣ ĐüĐÇđÁđ┤đżđ╣ đŞ┬áĐüđŞđŻĐůĐÇđżđŻđŞđĚđ░ĐćđŞĐÄ đ┐ĐÇđżĐćđÁĐüĐüđżđ▓ đ▓đŻđŞđ╝đ░đŻđŞĐĆ đŞ┬áđ┐đ░đ╝ĐĆĐéđŞ (đ▓┬áĐéđżđ╝ ĐçđŞĐüđ╗đÁ đ┐ĐÇđŞđ┐đżđ╝đŞđŻđ░đŻđŞđÁ đŞđŻĐüĐéĐÇĐâđ║ĐćđŞđŞ) đ┤đ╗ĐĆ đ▒ĐőĐüĐéĐÇđżđ│đż đ▓đżđĚđżđ▒đŻđżđ▓đ╗đÁđŻđŞĐĆ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐĆ đĚđ░đ┤đ░ĐçđŞ.
đĺ┬áđ┐đÁĐÇđ▓đżđ╝ đ▓đŞđ┤đÁ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ đ┐ĐÇđŞ đ┐đżđ▓ĐéđżĐÇđŻđżđ╝ đĚđ░ĐüĐőđ┐đ░đŻđŞđŞ đ┐đżĐüđ╗đÁ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ đŞđĚ┬áN2 đŞ┬áđ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đ▓┬áĐâĐüđ╗đżđ▓đŞĐĆĐů đŞđŻđÁĐÇĐćđŞđŞ ĐüđŻđ░, Đéđż đÁĐüĐéĐî đżĐëĐâĐéđŞđ╝đżđ╣ ĐüđżđŻđ╗đŞđ▓đżĐüĐéđŞ [26], đ┐đżđ║đ░đĚđ░đŻđż┬áđ│đÁđŻđÁĐÇđ░đ╗đŞđĚđżđ▓đ░đŻđŻđżđÁ Đâđ▓đÁđ╗đŞĐçđÁđŻđŞđÁ đ╝đżĐëđŻđżĐüĐéđŞ đ┤đÁđ╗ĐîĐéđ░- đŞ┬áĐéđÁĐéđ░-ĐÇđŞĐéđ╝đżđ▓, đ░┬áĐéđ░đ║đÂđÁ ĐÇđżĐüĐé đ░đ╗ĐîĐäđ░-ĐÇđŞĐéđ╝đ░ Đü┬áđ╗đżđ║đ░đ╗đŞđĚđ░ĐćđŞđÁđ╣ đ▓┬áđ┐đÁĐÇđÁđ┤đŻđŞĐů đżđ▒đ╗đ░ĐüĐéĐĆĐů, ĐçĐéđż Đüđ▓đŞđ┤đÁĐéđÁđ╗ĐîĐüĐéđ▓ĐâđÁĐé đż┬áđ▒đżđ╗đÁđÁ đ▓ĐőĐÇđ░đÂđÁđŻđŻđżđ╝ đ┐đż┬áđżĐéđŻđżĐłđÁđŻđŞĐÄ đ║┬áđĚđ░ĐüĐőđ┐đ░đŻđŞĐÄ đ▓┬áđ║đżđŻĐćđÁ N1 ĐâĐÇđżđ▓đŻđÁ ĐüđżđŻđ╗đŞđ▓đżĐüĐéđŞ [4, 19].
đĺđż đ▓ĐéđżĐÇđżđ╝ đ▓đŞđ┤đÁ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞĐĆ đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ đ┐đżĐüđ╗đÁ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ đŞđĚ┬áN2 đżĐéđ╝đÁĐçđ░đÁĐéĐüĐĆ ĐÇđżĐüĐé ĐâĐÇđżđ▓đŻĐĆ đ┤đÁđ╗ĐîĐéđ░-đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ đŻđ░┬áđşđşđô đżĐé┬áđ┐đÁĐÇđ▓đżđ│đż đżĐéĐÇđÁđĚđ║đ░ (đŻđÁđ┐đżĐüĐÇđÁđ┤ĐüĐéđ▓đÁđŻđŻđż đ┐đżĐüđ╗đÁ đ┐ĐÇđÁđ║ĐÇđ░ĐëđÁđŻđŞĐĆ đŻđ░đÂđ░ĐéđŞđ╣) đ║┬áđ┐đżĐüđ╗đÁđ┤ĐâĐÄĐëđÁđ╝Đâ, ĐçĐéđż đ┐ĐÇđŞđ▒đ╗đŞđÂđ░đÁĐé đ▓đżđĚđ▓ĐÇđ░ĐëđÁđŻđŞđÁ đŞĐüđ┐ĐőĐéĐâđÁđ╝đżđ│đż đ▓┬áN2 [24]. đčđżđ║đ░đĚđ░đŻđż Đéđ░đ║đÂđÁ ĐüđŻđŞđÂđÁđŻđŞđÁ đ╝đżĐëđŻđżĐüĐéđŞ đ▓ĐüđÁĐů đ▓ĐőĐüđżđ║đżĐçđ░ĐüĐéđżĐéđŻĐőĐů ĐÇđŞĐéđ╝đżđ▓ đżĐé┬áđ┐đÁĐÇđ▓đżđ│đż đżĐéĐÇđÁđĚđ║đ░ đ║┬áđ┐đżĐüđ╗đÁđ┤ĐâĐÄĐëđÁđ╝Đâ, đ░┬áĐéđ░đ║đÂđÁ đ▒߯╣đ╗ĐîĐłđŞđÁ Đüđ┐đÁđ║ĐéĐÇđ░đ╗ĐîđŻĐőđÁ đ▓đÁđ╗đŞĐçđŞđŻĐő đ░đ╗ĐîĐäđ░-1- đŞ┬áđ░đ╗ĐîĐäđ░-2-ĐÇđŞĐéđ╝đżđ▓ đŻđ░┬áđ┐đÁĐÇđ▓đżđ╝ đżĐéĐÇđÁđĚđ║đÁ đ▓┬áĐüđŞĐéĐâđ░ĐćđŞđŞ Đü┬áđ║đżĐÇđżĐéđ║đżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéĐîĐÄ. đĺ┬áđ┐đżĐüđ╗đÁđ┤đŻđÁđ╝ Đüđ╗ĐâĐçđ░đÁ ĐŹĐéđż đ╝đżđÂđÁĐé Đâđ║đ░đĚĐőđ▓đ░ĐéĐî đŻđ░┬áđ▒đżđ╗đÁđÁ đ▓ĐőĐÇđ░đÂđÁđŻđŻĐőđ╣ ĐâĐÇđżđ▓đÁđŻĐî ĐüđżđŻđ╗đŞđ▓đżĐüĐéđŞ đ▓┬áđŻđ░Đçđ░đ╗đÁ đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆ Đâ┬áĐüĐâđ▒ĐŐđÁđ║Đéđżđ▓, đ║đżĐéđżĐÇĐőđÁ đ┐đżĐüđ╗đÁ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ đŞđĚ┬áN2 Đüđ╝đżđ│đ╗đŞ đ▓Đőđ┐đżđ╗đŻđŞĐéĐî, đ┐ĐÇđ░đ▓đ┤đ░ Đçđ░ĐüĐéđŞĐçđŻđż, đ┐đÁĐÇđ▓Đőđ╣ đ┐đ░ĐéĐéđÁĐÇđŻ đŻđ░đÂđ░ĐéđŞđ╣. đíĐâĐëđÁĐüĐéđ▓đÁđŻđŻđżđÁ ĐüđŻđŞđÂđÁđŻđŞđÁ đżđ▒đżđŞĐů đ░đ╗ĐîĐäđ░- đŞ┬áđ▒đÁĐéđ░-ĐÇđŞĐéđ╝đżđ▓ đżĐé┬áđ┐đÁĐÇđ▓đżđ│đż đ║đż┬áđ▓ĐéđżĐÇđżđ╝Đâ đżĐéĐÇđÁđĚđ║Đâ đ┐đżĐüđ╗đÁ đ┐ĐÇđÁđ║ĐÇđ░ĐëđÁđŻđŞĐĆ đ┐ĐüđŞĐůđżđ╝đżĐéđżĐÇđŻđżđ╣ đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ đ║đ░đ║ đŻđ░┬áđ║đżĐÇđżĐéđ║đżđ╝ đŞđŻĐéđÁĐÇđ▓đ░đ╗đÁ đ▓ĐÇđÁđ╝đÁđŻđŞ, Đéđ░đ║ đŞ┬áđŻđ░ đ▒đżđ╗đÁđÁ đ┤đ╗đŞĐéđÁđ╗ĐîđŻđżđ╝ đ╝đżđÂđÁĐé Đüđ▓đŞđ┤đÁĐéđÁđ╗ĐîĐüĐéđ▓đżđ▓đ░ĐéĐî đż┬áĐüđŻđŞđÂđÁđŻđŞđŞ ĐâĐÇđżđ▓đŻĐĆ ĐüđżđĚđŻđ░đŻđŞĐĆ đŞ┬áđ▓ĐőĐüĐłđŞĐů đ║đżđ│đŻđŞĐéđŞđ▓đŻĐőĐů ĐäĐâđŻđ║ĐćđŞđ╣ đ▓┬áĐŹĐéđżĐé đ┐đÁĐÇđŞđżđ┤ [21ÔÇô24]. đŁđ░┬áĐäđżđŻđÁ đ┐đżđ▓đÁđ┤đÁđŻđŞĐĆ đ▓ĐüđÁĐů đżĐüĐéđ░đ╗ĐîđŻĐőĐů Đçđ░ĐüĐéđżĐé ĐüđŻđŞđÂđÁđŻđŞđÁ ĐéđÁĐéđ░-đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ đ╝Đő đżđ▒ĐŐĐĆĐüđŻĐĆđÁđ╝ đ▓┬áđ▒đżđ╗ĐîĐłđÁđ╣ ĐüĐéđÁđ┐đÁđŻđŞ đ┐ĐÇđÁđ║ĐÇđ░ĐëđÁđŻđŞđÁđ╝ đ┐ĐüđŞĐůđżđ╝đżĐéđżĐÇđŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đŞ┬áĐüđ▓ĐĆđĚđ░đŻđŻĐőđ╝ Đü┬áđŻđÁđ╣ đ║đżđ│đŻđŞĐéđŞđ▓đŻĐőđ╝ đ║đżđŻĐéĐÇđżđ╗đÁđ╝ [27], ĐçđÁđ╝ ĐüđŻđŞđÂđÁđŻđŞđÁđ╝ đ┤đ░đ▓đ╗đÁđŻđŞĐĆ ĐüđŻđ░. đĺ┬áđŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞđŞ ĐéđÁĐéđ░-đ║đżđ╗đÁđ▒đ░đŻđŞđ╣ đ▓đż đ▓ĐÇđÁđ╝ĐĆ đ▓Đőđ┐đżđ╗đŻđÁđŻđŞĐĆ đ║đżđ│đŻđŞĐéđŞđ▓đŻĐőĐů đĚđ░đ┤đ░Đç đŞ┬áđ╗đŞĐłđÁđŻđŞĐĆ ĐüđŻđ░ ĐÇđ░đĚđ╗đŞĐçđŞđ╣ đ╝đÁđÂđ┤Đâ đŻđŞđ╝đŞ đŻđÁ┬áđżđ▒đŻđ░ĐÇĐâđÂđÁđŻđż, đŻđż┬áđ┐đżđ╗ĐâĐçđÁđŻĐő đ┤đżđ║đ░đĚđ░ĐéđÁđ╗ĐîĐüĐéđ▓đ░, ĐçĐéđż ┬źđ▓┬áđżđ▒đżđŞĐů Đüđ╗ĐâĐçđ░ĐĆĐů ĐéđÁĐéđ░-đ║đżđ╗đÁđ▒đ░đŻđŞĐĆ┬áđ│đÁđŻđÁĐÇđŞĐÇĐâĐÄĐéĐüĐĆ đ║đżĐÇđ║đżđ▓Đőđ╝đŞ đżđ▒đ╗đ░ĐüĐéĐĆđ╝đŞ, đ║đżĐéđżĐÇĐőđÁ đŻđÁ┬áĐéĐÇđÁđ▒ĐâĐÄĐéĐüĐĆ đ┤đ╗ĐĆ ĐéđÁđ║ĐâĐëđÁđ│đż đ┐đżđ▓đÁđ┤đÁđŻđŞĐĆ. đčđżĐŹĐéđżđ╝Đâ, đ┐đż┬áđ║ĐÇđ░đ╣đŻđÁđ╣ đ╝đÁĐÇđÁ Đâ┬áđ╗ĐÄđ┤đÁđ╣, ĐéđÁĐéđ░-đ║đżđ╗đÁđ▒đ░đŻđŞĐĆ đ╝đżđ│ĐâĐé đżĐéĐÇđ░đÂđ░ĐéĐî đ╗đŞđ▒đż đ║đżĐÇđ║đżđ▓đżđÁ đżĐéđ║đ╗ĐÄĐçđÁđŻđŞđÁ, đ╗đŞđ▒đż ĐéđżĐÇđ╝đżđÂđÁđŻđŞđÁ┬╗ [28].
đŚđ░đ║đ╗ĐÄĐçđÁđŻđŞđÁ
đŁđ░ĐłđÁ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞđÁ đ┐đżđ║đ░đĚđ░đ╗đż ĐÇđ░đĚđ╗đŞĐçđŞĐĆ đ▓┬áĐçđ░ĐüĐéđżĐéđŻĐőĐů Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║đ░Đů đşđşđô đ┐ĐÇđŞ đ┐đżđ▓ĐéđżĐÇđŻĐőĐů đĚđ░ĐüĐőđ┐đ░đŻđŞĐĆĐů đ┐đżĐüđ╗đÁ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞđ╣, Đüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░đÁđ╝ĐőĐů đ┐ĐüđŞĐůđżđ╝đżĐéđżĐÇđŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéĐîĐÄ, đŻđ░┬áĐÇđ░đĚđŻĐőĐů ĐüĐéđ░đ┤đŞĐĆĐů ĐüđŻđ░. đôđÁđŻđÁĐÇđ░đ╗đŞđĚđżđ▓đ░đŻđŻđżđÁ Đâđ▓đÁđ╗đŞĐçđÁđŻđŞđÁ đ╝đżĐëđŻđżĐüĐéđŞ đ┤đÁđ╗ĐîĐéđ░- đŞ┬áĐéđÁĐéđ░-ĐÇđŞĐéđ╝đżđ▓, đ░┬áĐéđ░đ║đÂđÁ ĐÇđżĐüĐé đ░đ╗ĐîĐäđ░-ĐÇđŞĐéđ╝đ░ Đü┬áđ╗đżđ║đ░đ╗đŞđĚđ░ĐćđŞđÁđ╣ đ▓┬áđ┐đÁĐÇđÁđ┤đŻđŞĐů đżđ▒đ╗đ░ĐüĐéĐĆĐů đ┐đżĐüđ╗đÁ ĐŹđ┐đŞđĚđżđ┤đ░ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ đŞđĚ┬áN2 đ┐đż┬áđżĐéđŻđżĐłđÁđŻđŞĐÄ đ║┬áđĚđ░ĐüĐőđ┐đ░đŻđŞĐÄ đ▓┬áđ║đżđŻĐćđÁ N1 Đâđ║đ░đĚĐőđ▓đ░ĐÄĐé đŻđ░┬áđ▒đżđ╗đÁđÁ đ▓ĐőĐÇđ░đÂđÁđŻđŻĐőđ╣ ĐâĐÇđżđ▓đÁđŻĐî ĐüđżđŻđ╗đŞđ▓đżĐüĐéđŞ. đ×đ▒đŻđ░ĐÇĐâđÂđÁđŻđŻĐőđ╣ đŻđ░đ╝đŞ đ║┬áđ║đżđŻĐćĐâ đĚđ░ĐüĐőđ┐đ░đŻđŞđ╣ ĐÇđżĐüĐé đ╝đżĐëđŻđżĐüĐéđŞ đ░đ╗ĐîĐäđ░-1-, đ░đ╗ĐîĐäđ░-2- đŞ┬áđ▒đÁĐéđ░-đ┤đŞđ░đ┐đ░đĚđżđŻđżđ▓ đ▓┬áđ┐đÁĐÇđŞđżđ┤ sleep onset, đżĐüđżđ▒đÁđŻđŻđż đ┐đżĐüđ╗đÁ đ║đżĐÇđżĐéđ║đżđ│đż đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ, Đüđ▓đŞđ┤đÁĐéđÁđ╗ĐîĐüĐéđ▓ĐâđÁĐé đż┬áĐéđżđ╝, ĐçĐéđż, đŻđÁĐüđ╝đżĐéĐÇĐĆ đŻđ░┬áĐüđ╗đÁđ┤đżđ▓ĐőđÁ ĐŹĐäĐäđÁđ║ĐéĐő đ╝đżđŻđżĐéđżđŻđŻđżđ╣ đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đŞ┬áđ┤ĐÇđÁđ╝đżĐéĐâ, đ╝đżđĚđ│ ĐüĐéĐÇđÁđ╝đŞĐéĐüĐĆ đ┐ĐÇđÁđżđ┤đżđ╗đÁđ▓đ░ĐéĐî đŞĐů đ▓đ╗đŞĐĆđŻđŞđÁ đŞ┬áđ▒ĐőĐéĐî┬áđ│đżĐéđżđ▓Đőđ╝ đ▓đżđĚđżđ▒đŻđżđ▓đŞĐéĐî đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéĐî. đŚđ░ĐüĐőđ┐đ░đŻđŞđÁ đ┐đżĐüđ╗đÁ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞđ╣ ĐÇđ░đĚđŻđżđ╣ đ┤đ╗đŞĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đŞđĚ┬áN2 Đüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░đÁĐéĐüĐĆ ĐüĐâĐëđÁĐüĐéđ▓đÁđŻđŻĐőđ╝ ĐÇđżĐüĐéđżđ╝ đ┤đÁđ╗ĐîĐéđ░- đŞ┬áĐüđŻđŞđÂđÁđŻđŞđÁđ╝ đ░đ╗ĐîĐäđ░-1-, đ░đ╗ĐîĐäđ░-2- đŞ┬áđ▒đÁĐéđ░-đ░đ║ĐéđŞđ▓đŻđżĐüĐéđŞ, ĐçĐéđż đ╝đżđÂđÁĐé┬áđ│đżđ▓đżĐÇđŞĐéĐî đż┬áđ▒ĐőĐüĐéĐÇđżđ╝ ĐüđŻđŞđÂđÁđŻđŞđŞ ĐâĐÇđżđ▓đŻĐĆ ĐüđżđĚđŻđ░đŻđŞĐĆ đŞ┬áđ▓ĐőĐüĐłđŞĐů đ║đżđ│đŻđŞĐéđŞđ▓đŻĐőĐů ĐäĐâđŻđ║ĐćđŞđ╣ đ▓┬áĐŹĐéđżĐé đ┐đÁĐÇđŞđżđ┤. đĄĐâđŻđ║ĐćđŞđżđŻđ░đ╗ĐîđŻđżđÁ ĐüđżĐüĐéđżĐĆđŻđŞđÁ ĐüĐâđ▒ĐŐđÁđ║Đéđ░ đ┐ĐÇđŞ đĚđ░ĐüĐőđ┐đ░đŻđŞđŞ đĚđ░đ▓đŞĐüđŞĐé đ▓┬áđ▒đżđ╗ĐîĐłđÁđ╣ ĐüĐéđÁđ┐đÁđŻđŞ đżĐé┬áĐüĐéđ░đ┤đŞđŞ ĐüđŻđ░, ĐçđÁđ╝ đżĐé┬áđ┤đ╗đŞĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đ┐ĐÇđÁđ┤ĐłđÁĐüĐéđ▓đżđ▓đ░đ▓ĐłđÁđ│đż đÁđ╝Đâ ĐŹđ┐đŞđĚđżđ┤đ░ đ┐ĐÇđżđ▒ĐâđÂđ┤đÁđŻđŞĐĆ, Đüđżđ┐ĐÇđżđ▓đżđÂđ┤đ░đÁđ╝đżđ│đż đ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéĐîĐÄ.
đŁđ░ĐüĐéđżĐĆĐëđÁđÁ đŞĐüĐüđ╗đÁđ┤đżđ▓đ░đŻđŞđÁ đ▓đŻđżĐüđŞĐé đ▓đ║đ╗đ░đ┤ đ▓┬áđ┐đżđŻđŞđ╝đ░đŻđŞđÁ đ┐đÁĐÇđÁĐůđżđ┤đ░ đżĐé┬áđ┤đÁĐĆĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đ║đż┬áĐüđŻĐâ, Đüđ┐đżĐüđżđ▒ĐüĐéđ▓ĐâđÁĐé đ╗ĐâĐçĐłđÁđ╝Đâ ĐÇđ░Đüđ┐đżđĚđŻđ░đ▓đ░đŻđŞĐÄ đ┐đÁĐÇđŞđżđ┤đ░ ĐüđżđŻđ╗đŞđ▓đżĐüĐéđŞ đŞ┬áđ┐đżđĚđ▓đżđ╗ĐĆđÁĐé┬áđ│đ╗Đâđ▒đÂđÁ đŞđĚĐâĐçđŞĐéĐî Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║đŞ đ║đżđ╗đÁđ▒đ░đŻđŞđ╣ đ▒đ┤đŞĐéđÁđ╗ĐîđŻđżĐüĐéđŞ đŞ┬áđ▓đŻđŞđ╝đ░đŻđŞĐĆ đ▓┬áĐŹĐéđżĐé đ┐đÁĐÇđŞđżđ┤. đčđżđ╗ĐâĐçđÁđŻđŻĐőđÁ đżđ▒ĐŐđÁđ║ĐéđŞđ▓đŻĐőđÁ đŻđÁđ╣ĐÇđżĐäđŞđĚđŞđżđ╗đżđ│đŞĐçđÁĐüđ║đŞđÁ Đůđ░ĐÇđ░đ║ĐéđÁĐÇđŞĐüĐéđŞđ║đŞ đĚđ░ĐüĐőđ┐đ░đŻđŞđ╣ đŻđ░┬áĐÇđ░đĚđŻĐőĐů ĐüĐéđ░đ┤đŞĐĆĐů ĐüđŻđ░ đ╝đżđ│ĐâĐé đ▒ĐőĐéĐî đŞĐüđ┐đżđ╗ĐîđĚđżđ▓đ░đŻĐő đ┤đ╗ĐĆ đ╗ĐâĐçĐłđÁđ╣ đ┤đŞđ░đ│đŻđżĐüĐéđŞđ║đŞ đŞ┬áđ╗đÁĐçđÁđŻđŞĐĆ ĐÇđ░ĐüĐüĐéĐÇđżđ╣ĐüĐéđ▓ ĐüđŻđ░ đ┐ĐÇđŞ đŻđ░ĐÇĐâĐłđÁđŻđŞĐĆĐů đÁđ│đż đ║đ░ĐçđÁĐüĐéđ▓đ░ đŞ┬áđŞđĚđ▒ĐőĐéđżĐçđŻđżđ╣ ĐäĐÇđ░đ│đ╝đÁđŻĐéđ░ĐćđŞđŞ, đ░┬áĐéđ░đ║đÂe đ┤đ╗ĐĆ đ║đżđŻĐéĐÇđżđ╗ĐĆ đŞ┬áđ┐ĐÇđżđ│đŻđżđĚđŞĐÇđżđ▓đ░đŻđŞĐĆ đŻđÁđÂđÁđ╗đ░ĐéđÁđ╗ĐîđŻĐőĐů đĚđ░ĐüĐőđ┐đ░đŻđŞđ╣ đ▓┬áđ┐đżđ▓ĐüđÁđ┤đŻđÁđ▓đŻđżđ╣ ĐÇđ░đ▒đżĐéđÁ.┬á
đáđ░đ▒đżĐéđ░ đ▓Đőđ┐đżđ╗đŻđÁđŻđ░ đ▓┬áĐÇđ░đ╝đ║đ░Đů đôđżĐüĐâđ┤đ░ĐÇĐüĐéđ▓đÁđŻđŻđżđ│đż đĚđ░đ┤đ░đŻđŞĐĆ đťđŞđŻđŞĐüĐéđÁĐÇĐüĐéđ▓đ░ đżđ▒ĐÇđ░đĚđżđ▓đ░đŻđŞĐĆ đŞ┬áđŻđ░Đâđ║đŞ đáđżĐüĐüđŞđ╣Đüđ║đżđ╣ đĄđÁđ┤đÁĐÇđ░ĐćđŞđŞ đŻđ░┬á2024ÔÇô2026┬áđ│đ│.
đÜđżđŻĐäđ╗đŞđ║Đé đŞđŻĐéđÁĐÇđÁĐüđżđ▓ đżĐéĐüĐâĐéĐüĐéđ▓ĐâđÁĐé.
Ye.A. Cheremushkin, PhD, N.Ye. Petrenko, PhD
Institute of Higher Nervous Activity and Neurophysiology of the Russian Academy of Sciences
Contact person: Yevgeny A. Cheremushkin, khton@mail.ru
In healthy young adults in the first and second stages of daytime sleep, falling asleep after episodes of spontaneous awakening of different duration accompanied by psychomotor activity was investigated. A generalized increase in the power of delta and theta EEG rhythms, as well as an increase in alpha with localization in the anterior regions after an episode of awakening from the second stage relative to falling asleep at the end of the first stage indicates a more pronounced level of sleepiness. The increase in the power of the alpha1, alpha2, and beta bands in the first stage, especially after a short awakening, that we found toward the end of falling asleep indicates that despite the trace effects of monotonous activity and drowsiness, the brain tends to overcome their influence and be ready to resume activity. Falling asleep after awakenings of different durations from the second stage is accompanied by an increase in delta and a decrease in alpha1, alpha2 and beta activity, which may indicate a rapid decline in consciousness and higher cognitive functions, during this period. The subject's functional state when falling asleep depends more on the stage of sleep than on the duration of the episode of awakening accompanied by activity that preceded it. The study contributes to the understanding of the transition from activity to sleep, helps to better recognize the period of sleepiness, and allows for a deeper study of the characteristics of vigilance and attention fluctuations during this period.
đúđ▓đ░đÂđ░đÁđ╝Đőđ╣ đ┐đżĐüđÁĐéđŞĐéđÁđ╗Đî uMEDp!
đúđ▓đÁđ┤đżđ╝đ╗ĐĆđÁđ╝ đĺđ░Đü đż Đéđżđ╝, ĐçĐéđż đĚđ┤đÁĐüĐî Đüđżđ┤đÁĐÇđÂđŞĐéĐüĐĆ đŞđŻĐäđżĐÇđ╝đ░ĐćđŞĐĆ, đ┐ĐÇđÁđ┤đŻđ░đĚđŻđ░ĐçđÁđŻđŻđ░ĐĆ đŞĐüđ║đ╗ĐÄĐçđŞĐéđÁđ╗ĐîđŻđż đ┤đ╗ĐĆ Đüđ┐đÁĐćđŞđ░đ╗đŞĐüĐéđżđ▓ đĚđ┤ĐÇđ░đ▓đżđżĐůĐÇđ░đŻđÁđŻđŞĐĆ.
đĽĐüđ╗đŞ đĺĐő đŻđÁ ĐĆđ▓đ╗ĐĆđÁĐéđÁĐüĐî Đüđ┐đÁĐćđŞđ░đ╗đŞĐüĐéđżđ╝ đĚđ┤ĐÇđ░đ▓đżđżĐůĐÇđ░đŻđÁđŻđŞĐĆ, đ░đ┤đ╝đŞđŻđŞĐüĐéĐÇđ░ĐćđŞĐĆ đŻđÁ đŻđÁĐüđÁĐé đżĐéđ▓đÁĐéĐüĐéđ▓đÁđŻđŻđżĐüĐéđŞ đĚđ░ đ▓đżđĚđ╝đżđÂđŻĐőđÁ đżĐéĐÇđŞĐćđ░ĐéđÁđ╗ĐîđŻĐőđÁ đ┐đżĐüđ╗đÁđ┤ĐüĐéđ▓đŞĐĆ, đ▓đżđĚđŻđŞđ║ĐłđŞđÁ đ▓ ĐÇđÁđĚĐâđ╗ĐîĐéđ░ĐéđÁ Đüđ░đ╝đżĐüĐéđżĐĆĐéđÁđ╗ĐîđŻđżđ│đż đŞĐüđ┐đżđ╗ĐîđĚđżđ▓đ░đŻđŞĐĆ đĺđ░đ╝đŞ đŞđŻĐäđżĐÇđ╝đ░ĐćđŞđŞ Đü đ┐đżĐÇĐéđ░đ╗đ░ đ▒đÁđĚ đ┐ĐÇđÁđ┤đ▓đ░ĐÇđŞĐéđÁđ╗ĐîđŻđżđ╣ đ║đżđŻĐüĐâđ╗ĐîĐéđ░ĐćđŞđŞ Đü đ▓ĐÇđ░Đçđżđ╝.
đŁđ░đÂđŞđ╝đ░ĐĆ đŻđ░ đ║đŻđżđ┐đ║Đâ ┬źđĺđżđ╣ĐéđŞ┬╗, đĺĐő đ┐đżđ┤Đéđ▓đÁĐÇđÂđ┤đ░đÁĐéđÁ, ĐçĐéđż ĐĆđ▓đ╗ĐĆđÁĐéđÁĐüĐî đ▓ĐÇđ░Đçđżđ╝ đŞđ╗đŞ ĐüĐéĐâđ┤đÁđŻĐéđżđ╝ đ╝đÁđ┤đŞĐćđŞđŻĐüđ║đżđ│đż đ▓ĐâđĚđ░.