–°–њ–µ–Ї—В—А–∞–ї—М–љ—Л–µ —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є —Н–ї–µ–Ї—В—А–Њ—Н–љ—Ж–µ—Д–∞–ї–Њ–≥—А–∞–Љ–Љ—Л –њ—А–Є¬†–≤—Л–њ–Њ–ї–љ–µ–љ–Є–Є –Ј–∞–і–∞–љ–Є–є —А–∞–Ј–љ–Њ–≥–Њ —Г—А–Њ–≤–љ—П —Б–ї–Њ–ґ–љ–Њ—Б—В–Є –≤–Њ –≤—А–µ–Љ—П –Ј–∞—Б—Л–њ–∞–љ–Є—П –њ–µ—А–µ–і¬†—Н–њ–Є–Ј–Њ–і–Њ–Љ –і–љ–µ–≤–љ–Њ–≥–Њ —Б–љ–∞
- –Р–љ–љ–Њ—В–∞—Ж–Є—П
- –°—В–∞—В—М—П
- –°—Б—Л–ї–Ї–Є
- English



–Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ —Н–ї–µ–Ї—В—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї –Љ–Њ–Ј–≥–Њ–≤–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –њ–µ—А–µ—Е–Њ–і–∞ –Њ—В¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Ї–Њ¬†—Б–љ—Г, —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–µ–Љ–Њ–≥–Њ –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–є –Є¬†–Ї–Њ–≥–љ–Є—В–Є–≤–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В—М—О, –Є–Љ–µ–µ—В –Њ–њ—А–µ–і–µ–ї–µ–љ–љ—Г—О –і–Є–∞–≥–љ–Њ—Б—В–Є—З–µ—Б–Ї—Г—О —Ж–µ–љ–љ–Њ—Б—В—М –і–ї—П –Њ—Ж–µ–љ–Ї–Є —В—А—Г–і–љ–Њ—Б—В–µ–є –Ј–∞—Б—Л–њ–∞–љ–Є—П —А–∞–Ј–љ–Њ–є —Н—В–Є–Њ–ї–Њ–≥–Є–Є [1, 2]. –Э–∞—А—П–і—Г —Б¬†—Н—В–Є–Љ –≤¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Њ–є –њ—А–∞–Ї—В–Є–Ї–µ –∞–Ї—В—Г–∞–ї—М–љ–Њ –Њ–њ—А–µ–і–µ–ї–µ–љ–Є–µ —Г—А–Њ–≤–љ—П –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є –Є¬†—Б–≤—П–Ј–∞–љ–љ—Л—Е —Б¬†–љ–µ–є —А–µ—Б—Г—А—Б–Њ–≤ –≤–љ–Є–Љ–∞–љ–Є—П –њ—А–Є –±–µ—Б—Б–Њ–љ–љ–Є—Ж–µ –Є¬†–љ–µ–≤—А–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –љ–∞—А—Г—И–µ–љ–Є—П—Е [3].
–Э–µ–є—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Ї–Њ—А—А–µ–ї—П—В—Л –њ—А–Є –Ј–∞—Б—Л–њ–∞–љ–Є–Є —Е–Њ—А–Њ—И–Њ –Є–Ј—Г—З–µ–љ—Л —Г¬†–Ј–і–Њ—А–Њ–≤—Л—Е —Б—Г–±—К–µ–Ї—В–Њ–≤ –Є¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–њ–∞—В–Њ–ї–Њ–≥–Є–µ–є [2, 4]. –Т–Љ–µ—Б—В–µ —Б¬†—В–µ–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Б¬†–Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є –≤¬†–Ї–∞—З–µ—Б—В–≤–µ —В–µ—Б—В–∞, –Њ—Ж–µ–љ–Є–≤–∞—О—Й–µ–≥–Њ —Б–Њ—Б—В–Њ—П–љ–Є–µ —З–µ–ї–Њ–≤–µ–Ї–∞ –њ—А–Є –њ–µ—А–µ—Е–Њ–і–µ –Њ—В¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Ї–Њ¬†—Б–љ—Г, –љ–µ–Љ–љ–Њ–≥–Њ. –≠—В–Њ —А–∞–±–Њ—В—Л –њ–Њ¬†–Є–Ј—Г—З–µ–љ–Є—О —Б–Њ—Б—В–Њ—П–љ–Є—П –Љ–Њ–љ–Њ—В–Њ–љ–Є–Є, –Ї–Њ—В–Њ—А–Њ–µ –≤–Њ–Ј–љ–Є–Ї–∞–µ—В –њ—А–Є –і–ї–Є—В–µ–ї—М–љ–Њ–є –Њ–і–љ–Њ–Њ–±—А–∞–Ј–љ–Њ–є —А–∞–±–Њ—В–µ –Є¬†–Љ–Њ–ґ–µ—В –њ—А–Є–≤–Њ–і–Є—В—М –Ї¬†—Н–њ–Є–Ј–Њ–і–∞–Љ –Ј–∞—Б—Л–њ–∞–љ–Є—П [5]. –Я—А–Є –≤—Л–њ–Њ–ї–љ–µ–љ–Є–Є –Є—Б–њ—Л—В—Г–µ–Љ—Л–Љ–Є –Ј–∞–і–∞—З–Є –њ–Њ¬†–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–Љ—Г –њ—А–Є–љ—П—В–Є—О —А–µ—И–µ–љ–Є–є –Є–Ј¬†–і–≤—Г—Е –≤–∞—А–Є–∞–љ—В–Њ–≤ –≤¬†—Б–Њ—Б—В–Њ—П–љ–Є–Є –і–µ–њ—А–Є–≤–∞—Ж–Є–Є –љ–∞¬†–Њ—Б–љ–Њ–≤–∞–љ–Є–Є —Б–Є–љ—Е—А–Њ–љ–љ—Л—Е –Ј–∞–њ–Є—Б–µ–є —Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–є –Љ–∞–≥–љ–Є—В–љ–Њ-—А–µ–Ј–Њ–љ–∞–љ—Б–љ–Њ–є —В–Њ–Љ–Њ–≥—А–∞—Д–Є–Є (—Д–Ь–†–Ґ) –Є¬†—Н–ї–µ–Ї—В—А–Њ—Н–љ—Ж–µ—Д–∞–ї–Њ–≥—А–∞—Д–Є–Є (–≠–≠–У) –њ–Њ–Ї–∞–Ј–∞–љ—Л —З–∞—Б—В—Л–µ –≤—В–Њ—А–ґ–µ–љ–Є—П —В–µ—В–∞-—А–Є—В–Љ–∞, —Б–≤—П–Ј—Л–≤–∞–µ–Љ—Л–µ —Б¬†–Ј–∞—Б—Л–њ–∞–љ–Є–µ–Љ, –≤¬†–Ї–∞—А—В–Є–љ—Г –±–Є–Њ—Н–ї–µ–Ї—В—А–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є, –Ї–Њ—В–Њ—А–∞—П —Е–∞—А–∞–Ї—В–µ—А–љ–∞ –і–ї—П –≤—Л–њ–Њ–ї–љ—П–µ–Љ–Њ–є –Ї–Њ–≥–љ–Є—В–Є–≤–љ–Њ–є –Ј–∞–і–∞—З–Є. –≠—В–Њ—В —А–Є—В–Љ –±—Л–ї –љ–∞–Є–±–Њ–ї–µ–µ –≤—Л—А–∞–ґ–µ–љ –≤¬†–ї–Њ–±–љ—Л—Е –Є¬†—В–µ–Љ–µ–љ–љ—Л—Е –Њ—В–і–µ–ї–∞—Е –Љ–Њ–Ј–≥–∞ [6]. –Т¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ–Њ–є –Љ–Њ–і–µ–ї–Є, –Є—Б—Б–ї–µ–і—Г—О—Й–µ–є –њ—А–Њ—Ж–µ—Б—Б—Л –Ј–∞—Б—Л–њ–∞–љ–Є—П –±–µ–Ј –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –≤–љ–µ—И–љ–Є–Љ–Є —Б–µ–љ—Б–Њ—А–љ—Л–Љ–Є —Б—В–Є–Љ—Г–ї–∞–Љ–Є, –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ —А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–ї–Є—Б—М –≠–≠–У, —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є –і—Л—Е–∞–љ–Є—П –Є¬†–њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–µ –і–∞–љ–љ—Л–µ (—Б–Є–ї–∞ —Б–і–∞–≤–ї–Є–≤–∞–љ–Є—П –Љ—П—З–Є–Ї–∞ —А—Г–Ї–Њ–є, —Д–Є–Ї—Б–∞—Ж–Є—П –Љ–Њ–Љ–µ–љ—В–∞ –љ–∞—З–∞–ї–∞ —Б–љ–∞, –Ї–Њ–≥–і–∞ –Є—Б–њ—Л—В—Г–µ–Љ—Л–є –≤—Л—А–Њ–љ–Є—В –µ–≥–Њ)¬†[7]. –Ш—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ –њ–Њ–≤–µ–і–µ–љ–Є—П, –∞¬†–Є–Љ–µ–љ–љ–Њ –њ–Њ—Б—В—Г–Ї–Є–≤–∞–љ–Є–є –њ–∞–ї—М—Ж–µ–Љ –Є¬†–љ–∞–ґ–∞—В–Є–є –љ–∞¬†–Ї–љ–Њ–њ–Ї—Г (—В–µ–њ–њ–Є–љ–≥-—В–µ—Б—В) –і–ї—П –Њ–њ—А–µ–і–µ–ї–µ–љ–Є—П –Љ–Њ–Љ–µ–љ—В–∞ –Ј–∞—Б—Л–њ–∞–љ–Є—П, –њ—А–µ–і–ї–∞–≥–∞–ї–Њ—Б—М –Є¬†—А–∞–љ–µ–µ [8]. –Т¬†–љ–∞—И–µ–є –ї–∞–±–Њ—А–∞—В–Њ—А–Є–Є –і–ї—П –Љ–Њ–і–µ–ї–Є—А–Њ–≤–∞–љ–Є—П –Ј–∞—Б—Л–њ–∞–љ–Є—П –≤–Њ –≤—А–µ–Љ—П –Њ–њ–µ—А–∞—В–Њ—А—Б–Ї–Њ–є –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є –њ—А–Њ–≤–Њ–і–Є–ї–Є—Б—М –Њ–њ—Л—В—Л —Б¬†–Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ [9]. –Ю—И–Є–±–Њ—З–љ–∞—П —А–µ–∞–ї–Є–Ј–∞—Ж–Є—П –Ј–∞–і–∞–љ–Є–є —В–µ—Б—В–∞ (–≤¬†—Г–љ–Є–Љ–∞–љ—Г–∞–ї—М–љ–Њ–є, —В–Њ –µ—Б—В—М —Б¬†–љ–∞–ґ–∞—В–Є—П–Љ–Є –љ–∞¬†–Ї–љ–Њ–њ–Ї—Г –Њ–і–љ–Њ–є —А—Г–Ї–Њ–є, –Љ–Њ–і–Є—Д–Є–Ї–∞—Ж–Є–Є), –Њ—Ж–µ–љ–Є–≤–∞–µ–Љ–∞—П –Ї–∞–Ї –і—А–µ–Љ–Њ—В–љ–Њ–µ —Б–Њ—Б—В–Њ—П–љ–Є–µ, —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–ї–∞—Б—М —А–Њ—Б—В–Њ–Љ –Љ–Њ—Й–љ–Њ—Б—В–Є —В–µ—В–∞-—А–Є—В–Љ–∞ –љ–∞¬†–≠–≠–У. –≠–њ–Є–Ј–Њ–і—Л –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є –±–µ–Ј –Њ—И–Є–±–Њ–Ї, —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–µ–Љ—Л–µ –Ї–∞–Ї –±–Њ–і—А—Б—В–≤—Г—О—Й–µ–µ —Б–Њ—Б—В–Њ—П–љ–Є–µ, –њ—А–Є–≤–Њ–і–Є–ї–Є –Ї¬†—А–Њ—Б—В—Г –∞–ї—М—Д–∞-—А–Є—В–Љ–∞¬†[10]. –Ю–і–љ–∞–Ї–Њ –≤¬†–і–∞–љ–љ–Њ–є —А–∞–±–Њ—В–µ –±–Њ–ї—М—И–µ –≤–љ–Є–Љ–∞–љ–Є—П —Г–і–µ–ї—П–ї–Њ—Б—М —Б–Њ–Ј–і–∞–љ–Є—О —Б—В–∞—В–Є—Б—В–Є—З–µ—Б–Ї–Њ–є –Љ–Њ–і–µ–ї–Є –Ј–∞—Б—Л–њ–∞–љ–Є—П –Њ–њ–µ—А–∞—В–Њ—А–∞, –∞¬†–љ–µ –і–µ—В–∞–ї—М–љ–Њ–Љ—Г –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—О –љ–µ–є—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Ї–Њ—А—А–µ–ї—П—В–Њ–≤ –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є –њ—А–Є –њ–µ—А–µ—Е–Њ–і–µ –Њ—В¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Ї–Њ¬†—Б–љ—Г.
–¶–µ–ї—М –љ–∞—И–µ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П¬†вАУ –њ—А–Њ–∞–љ–∞–ї–Є–Ј–Є—А–Њ–≤–∞—В—М, –Ї–∞–Ї —Б–ї–Њ–ґ–љ–∞—П –Є¬†–њ—А–Њ—Б—В–∞—П —Д–∞–Ј—Л –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ (—Б—З–µ—В –≤¬†—Г–Љ–µ, —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–µ–Љ—Л–є —Б–Є–љ—Е—А–Њ–љ–љ—Л–Љ–Є –љ–∞–ґ–∞—В–Є—П–Љ–Є –љ–∞¬†–Ї–љ–Њ–њ–Ї—Г, –Є¬†—В–Њ–ї—М–Ї–Њ —Б—З–µ—В –≤¬†—Г–Љ–µ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ) –Њ—В—А–∞–ґ–∞—О—В—Б—П –≤¬†–±–Є–Њ—Н–ї–µ–Ї—В—А–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є —А–∞–Ј–љ—Л—Е –Њ–±–ї–∞—Б—В–µ–є –µ–µ –Њ—В–≤–µ–і–µ–љ–Є—П –≤¬†–љ–∞—З–∞–ї–µ –Є¬†–Ї–Њ–љ—Ж–µ –Ј–∞—Б—Л–њ–∞–љ–Є—П. –Ч–∞–і–∞—З–Є –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П¬†вАУ —Б—А–∞–≤–љ–Є—В—М –Љ–Њ—Й–љ–Њ—Б—В–љ—Л–µ —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є –≠–≠–У –≤¬†–љ–∞—З–∞–ї–µ –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є —Б¬†–≠–≠–У –њ—А–Є –њ–µ—А–≤–Њ–Љ –њ–∞—В—В–µ—А–љ–µ –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–є –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є —Б¬†–Њ—И–Є–±–Ї–Њ–є —Б¬†–њ–Њ—Б–ї–µ–і–љ–Є–Љ –њ–∞—В—В–µ—А–љ–Њ–Љ –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є, –њ—А–µ–і—И–µ—Б—В–≤–Њ–≤–∞–≤—И–Є–Љ —Н–њ–Є–Ј–Њ–і—Г —Б–љ–∞, –∞¬†—В–∞–Ї–ґ–µ —Б—А–∞–≤–љ–Є—В—М —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є –≠–≠–У —Н–њ–Є–Ј–Њ–і–Њ–≤ –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є —Б¬†–њ–µ—А–≤–Њ–є –Є¬†–њ–Њ—Б–ї–µ–і–љ–µ–є –Њ—И–Є–±–Ї–Њ–є –њ—А–Є –Ј–∞—Б—Л–њ–∞–љ–Є–Є.
–Ь–∞—В–µ—А–Є–∞–ї –Є¬†–Љ–µ—В–Њ–і—Л
–Т¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є —Г—З–∞—Б—В–≤–Њ–≤–∞–ї–Є 34 –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –Ј–і–Њ—А–Њ–≤—Л—Е –ї–Є—Ж–∞¬†вАУ —Б—В—Г–і–µ–љ—В—Л –Љ–Њ—Б–Ї–Њ–≤—Б–Ї–Є—Е –≤—Г–Ј–Њ–≤ (26 –ґ–µ–љ—Й–Є–љ –Є¬†8¬†–Љ—Г–ґ—З–Є–љ, –≤–Њ–Ј—А–∞—Б—В –Њ—В¬†19 –і–Њ 22 –ї–µ—В). –Ю–љ–Є –±—Л–ї–Є –Њ–Ј–љ–∞–Ї–Њ–Љ–ї–µ–љ—Л —Б¬†–њ—А–Њ—Ж–µ–і—Г—А–Њ–є –Њ–њ—Л—В–∞ –Є¬†–і–∞–ї–Є —Б–Њ–≥–ї–∞—Б–Є–µ –љ–∞¬†—Г—З–∞—Б—В–Є–µ –≤¬†–љ–µ–Љ. –Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –±—Л–ї–Њ –Њ–і–Њ–±—А–µ–љ–Њ —Н—В–Є—З–µ—Б–Ї–Њ–є –Ї–Њ–Љ–Є—Б—Б–Є–µ–є –Ш–љ—Б—В–Є—В—Г—В–∞ –≤—Л—Б—И–µ–є –љ–µ—А–≤–љ–Њ–є –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є –Є¬†–љ–µ–є—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є–Є –†–Р–Э (–њ—А–Њ—В–Њ–Ї–Њ–ї вДЦ¬†2 –Њ—В¬†3 –Є—О–љ—П 2019¬†–≥.) –Є¬†—Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Њ–≤–∞–ї–Њ —Н—В–Є—З–µ—Б–Ї–Є–Љ –љ–Њ—А–Љ–∞–Љ –•–µ–ї—М—Б–Є–љ–Ї—Б–Ї–Њ–є –і–µ–Ї–ї–∞—А–∞—Ж–Є–Є –Т—Б–µ–Љ–Є—А–љ–Њ–є –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–є –∞—Б—Б–Њ—Ж–Є–∞—Ж–Є–Є ¬Ђ–≠—В–Є—З–µ—Б–Ї–Є–µ –њ—А–Є–љ—Ж–Є–њ—Л –њ—А–Њ–≤–µ–і–µ–љ–Є—П –љ–∞—Г—З–љ—Л—Е –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Б¬†—Г—З–∞—Б—В–Є–µ–Љ —З–µ–ї–Њ–≤–µ–Ї–∞¬ї —Б¬†–њ–Њ–њ—А–∞–≤–Ї–∞–Љ–Є 2000¬†–≥.¬†–Т—А–µ–Љ—П —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞¬†вАУ —Б¬†13.00 –і–Њ 16.00. –Т—Л–њ–Њ–ї–љ–µ–љ–Є–µ –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ —Б¬†—Н–њ–Є–Ј–Њ–і–∞–Љ–Є —Б–љ–∞¬†вАУ –Њ—В¬†55 –Љ–Є–љ—Г—В –і–Њ 1 —З–∞—Б–∞ 10 –Љ–Є–љ—Г—В. –Э–∞¬†–њ—А–Њ—В—П–ґ–µ–љ–Є–Є –≤—Б–µ–≥–Њ –Њ–њ—Л—В–∞ –Є—Б–њ—Л—В—Г–µ–Љ—Л–µ –љ–∞—Е–Њ–і–Є–ї–Є—Б—М –≤¬†–Ј–∞—В–µ–Љ–љ–µ–љ–љ–Њ–Љ –Ј–≤—Г–Ї–Њ–Є–Ј–Њ–ї–Є—А—Г—О—Й–µ–Љ –њ–Њ–Љ–µ—Й–µ–љ–Є–Є –≤¬†–њ–Њ–ї–Њ–ґ–µ–љ–Є–Є –ї–µ–ґ–∞ –љ–∞¬†–Ї—Г—И–µ—В–Ї–µ —Б¬†–Ј–∞–Ї—А—Л—В—Л–Љ–Є¬†–≥–ї–∞–Ј–∞–Љ–Є. –Т¬†—В–µ—З–µ–љ–Є–µ 5 –Љ–Є–љ—Г—В –њ—А–Њ–≤–Њ–і–Є–ї–Є –Ј–∞–њ–Є—Б—М –≠–≠–У –≤¬†—Б–Њ—Б—В–Њ—П–љ–Є–Є —Б–њ–Њ–Ї–Њ–є–љ–Њ–≥–Њ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П. –°¬†–њ–Њ–Љ–Њ—Й—М—О —Г–љ–Є–Љ–∞–љ—Г–∞–ї—М–љ–Њ–≥–Њ (—Б¬†–љ–∞–ґ–∞—В–Є—П–Љ–Є –љ–∞¬†–Ї–љ–Њ–њ–Ї—Г –Њ–і–љ–Њ–є —А—Г–Ї–Њ–є) –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ [9] –њ–Њ–ї—Г—З–∞–ї–Є —А—П–і –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ—Л—Е –њ–µ—А–Є–Њ–і–Њ–≤ –Ј–∞—Б—Л–њ–∞–љ–Є—П –Є¬†–њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П. –Ш—Б–њ—Л—В—Г–µ–Љ—Л–µ —Б—З–Є—В–∞–ї–Є –≤¬†—Г–Љ–µ –Њ—В¬†–Њ–і–љ–Њ–≥–Њ –і–Њ –і–µ—Б—П—В–Є –Є¬†—Б–Є–љ—Е—А–Њ–љ–љ–Њ –љ–∞–ґ–Є–Љ–∞–ї–Є –Ј–∞–Ї—А–µ–њ–ї–µ–љ–љ—Г—О –љ–∞¬†—Г–Ї–∞–Ј–∞—В–µ–ї—М–љ–Њ–Љ –њ–∞–ї—М—Ж–µ –Ї–љ–Њ–њ–Ї—Г –±–Њ–ї—М—И–Є–Љ –њ–∞–ї—М—Ж–µ–Љ –њ—А–∞–≤–Њ–є —А—Г–Ї–Є (–њ–µ—А–≤–∞—П —Д–∞–Ј–∞ —В–µ—Б—В–∞). –Ф–∞–ї–µ–µ –Њ–љ–Є –њ—А–Њ–і–Њ–ї–ґ–∞–ї–Є —Б—З–Є—В–∞—В—М –≤¬†—Г–Љ–µ –Њ—В¬†–Њ–і–љ–Њ–≥–Њ –і–Њ –і–µ—Б—П—В–Є –±–µ–Ј –љ–∞–ґ–∞—В–Є–є (–≤—В–Њ—А–∞—П —Д–∞–Ј–∞). –І–µ—А–µ–і–Њ–≤–∞–љ–Є–µ —Б—З–µ—В–∞ —Б¬†–љ–∞–ґ–∞—В–Є—П–Љ–Є –Є¬†–±–µ–Ј –љ–∞–ґ–∞—В–Є–є –њ—А–Њ–і–Њ–ї–ґ–∞–ї–Њ—Б—М, –њ–Њ–Ї–∞ –Є—Б–њ—Л—В—Г–µ–Љ—Л–µ –љ–µ¬†–Ј–∞—Б—Л–њ–∞–ї–Є. –Я–Њ—Б–ї–µ —Н–њ–Є–Ј–Њ–і–∞ —Б–љ–∞ –Є¬†–њ–Њ—Б–ї–µ–і—Г—О—Й–µ–≥–Њ —Б–∞–Љ–Њ–њ—А–Њ–Є–Ј–≤–Њ–ї—М–љ–Њ–≥–Њ –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П –Њ–љ–Є –і–Њ–ї–ґ–љ—Л –±—Л–ї–Є –≤–Њ–Ј–Њ–±–љ–Њ–≤–Є—В—М –≤—Л–њ–Њ–ї–љ–µ–љ–Є–µ –Ј–∞–і–∞–љ–Є–є –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ —Б¬†–њ–µ—А–≤–Њ–є —Д–∞–Ј—Л. –Ш—Б–њ—Л—В—Г–µ–Љ—Л—Е, –Ї–Њ—В–Њ—А—Л–µ –љ–µ¬†–Ј–∞—Б—Л–њ–∞–ї–Є –Є–ї–Є –Ј–∞—Б—Л–њ–∞–ї–Є –Є¬†–љ–µ –њ—А–Њ—Б—Л–њ–∞–ї–Є—Б—М –і–Њ –Ї–Њ–љ—Ж–∞ –Њ–њ—Л—В–∞, –Є—Б–Ї–ї—О—З–∞–ї–Є –Є–Ј¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П.
–Т¬†—В–µ—З–µ–љ–Є–µ –Њ–њ—Л—В–∞ —Б¬†–њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є¬†–≥–Њ–ї–Њ–≤—Л —А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–ї–Є –≠–≠–У –Њ—В¬†17 –Њ—В–≤–µ–і–µ–љ–Є–є, —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–љ—Л—Е –≤¬†—Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Є–Є —Б–Њ¬†—Б—Е–µ–Љ–Њ–є 10вАУ20% (F3, F4, F7, F8, Fz, C3, C4, Cz, T3, T4, P3, P4, Pz, T5, T6, O1, O2). –Ю—В–≤–µ–і–µ–љ–Є–µ –Љ–Њ–љ–Њ–њ–Њ–ї—П—А–љ–Њ–µ, —А–µ—Д–µ—А–µ–љ—В–љ—Л–є —Н–ї–µ–Ї—В—А–Њ–і¬†вАУ –Њ–±—К–µ–і–Є–љ–µ–љ–љ—Л–є —Г—И–љ–Њ–є. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –Ј–∞–њ–Є—Б—Л–≤–∞–ї–Є —Н–ї–µ–Ї—В—А–Њ–Њ–Ї—Г–ї–Њ–≥—А–∞–Љ–Љ—Г (–≠–Ю–У), –Љ–Є–Њ–≥—А–∞–Љ–Љ—Г (–≠–Ь–У) –Є¬†–Љ–µ—Е–∞–љ–Њ–≥—А–∞–Љ–Љ—Г –љ–∞–ґ–∞—В–Є–є –љ–∞¬†–Ї–љ–Њ–њ–Ї—Г. –†–µ–≥–Є—Б—В—А–∞—Ж–Є—О –≤—Б–µ—Е –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–µ–є –њ—А–Њ–≤–Њ–і–Є–ї–Є —Б¬†–њ–Њ–Љ–Њ—Й—М—О —Б–Є—Б—В–µ–Љ—Л Neocortex-Pro (Neurobotics, –†–Њ—Б—Б–Є—П). –І–∞—Б—В–Њ—В–∞ –і–Є—Б–Ї—А–µ—В–Є–Ј–∞—Ж–Є–Є¬†вАУ 250 –У—Ж. –Я–Њ–ї–Њ—Б–∞ –њ—А–Њ–њ—Г—Б–Ї–∞–љ–Є—П —З–∞—Б—В–Њ—В¬†вАУ 0,5вАУ70 –У—Ж. –≠–≠–У —А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–ї–Є —Б¬†–њ–Њ–Љ–Њ—Й—М—О —И–ї–µ–Љ–∞ —Б¬†—Е–ї–Њ—А—Б–µ—А–µ–±—А—П–љ—Л–Љ–Є —Н–ї–µ–Ї—В—А–Њ–і–∞–Љ–Є —Б¬†—Б–Њ–њ—А–Њ—В–Є–≤–ї–µ–љ–Є–µ–Љ, –љ–µ¬†–њ—А–µ–≤—Л—И–∞—О—Й–Є–Љ 5 –Ъ–Ю–Љ.
–Я–Њ—Б–ї–µ –≤–Є–Ј—Г–∞–ї—М–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –њ–Њ–ї–Є—Б–Њ–Љ–љ–Њ–≥—А–∞–Љ–Љ –Є¬†–Љ–µ—Е–∞–љ–Њ–≥—А–∞–Љ–Љ –≤—Л–і–µ–ї—П–ї–Є –Є—Б–њ—Л—В—Г–µ–Љ—Л—Е, –љ–∞¬†–≠–≠–У –Ї–Њ—В–Њ—А—Л—Е –љ–∞–±–ї—О–і–∞–ї–Є—Б—М –њ–µ—А–Є–Њ–і—Л –Ј–∞—Б—Л–њ–∞–љ–Є—П (n = 24). –Ф–ї—П –∞–љ–∞–ї–Є–Ј–∞ –≤—Л–±–Є—А–∞–ї–Є 30-—Б–µ–Ї—Г–љ–і–љ—Л–µ –Њ—В—А–µ–Ј–Ї–Є –Ј–∞–њ–Є—Б–Є –≠–≠–У –≤¬†—В—А–µ—Е —Б–Є—В—Г–∞—Ж–Є—П—Е:
- —Н–њ–Є–Ј–Њ–і –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ –≤¬†–љ–∞—З–∞–ї–µ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞ (–Ї–Њ–љ—В—А–Њ–ї—М, —Б–Њ—Б—В–Њ—П–љ–Є–µ –њ–Њ–ї–љ–Њ–є –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є);
- –њ–µ—А–≤—Л–є —Н–њ–Є–Ј–Њ–і –Њ—И–Є–±–Њ—З–љ–Њ–≥–Њ –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П –Ј–∞–і–∞–љ–Є–є –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ (—А–∞–љ–љ—П—П —Б—В–∞–і–Є—П –Ј–∞—Б—Л–њ–∞–љ–Є—П);
- –≤—Л–њ–Њ–ї–љ–µ–љ–Є–µ –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –њ–µ—А–µ–і –љ–∞—Б—В—Г–њ–ї–µ–љ–Є–µ–Љ –њ–µ—А–≤–Њ–≥–Њ —Н–њ–Є–Ј–Њ–і–∞ —Б–љ–∞ (–њ–Њ–Ј–і–љ—П—П —Б—В–∞–і–Є—П).
–Ф–ї—П —Б–Є—В—Г–∞—Ж–Є–Є 3 –≤—Л–±–Є—А–∞–ї–Є –Њ—В—А–µ–Ј–Ї–Є, –Ј–∞ –Ї–Њ—В–Њ—А—Л–Љ–Є —Б–ї–µ–і–Њ–≤–∞–ї —Н–њ–Є–Ј–Њ–і —Б–љ–∞ –љ–µ¬†–Љ–µ–љ–µ–µ 1,5 –Љ–Є–љ—Г—В—Л. –Я—А–Є–љ–∞–і–ї–µ–ґ–љ–Њ—Б—В—М —Г—З–∞—Б—В–Ї–Њ–≤ –≠–≠–У –Ї¬†–њ–µ—А–Є–Њ–і—Г –љ–∞—З–∞–ї–∞ —Б–љ–∞ (sleeponset) –Њ—Ж–µ–љ–Є–≤–∞–ї–Є –≤–Є–Ј—Г–∞–ї—М–љ–Њ –њ–Њ¬†—Б—В–∞–љ–і–∞—А—В–љ—Л–Љ –Ї—А–Є—В–µ—А–Є—П–Љ –Р–Љ–µ—А–Є–Ї–∞–љ—Б–Ї–Њ–є –∞–Ї–∞–і–µ–Љ–Є–Є –Љ–µ–і–Є—Ж–Є–љ—Л —Б–љ–∞ (AASM) [11]. –Ф–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ—Л–Љ –Ї—А–Є—В–µ—А–Є–µ–Љ —Б—З–Є—В–∞–ї–Є –Њ—В—Б—Г—В—Б—В–≤–Є–µ –љ–∞¬†–Љ–µ—Е–∞–љ–Њ–≥—А–∞–Љ–Љ–µ –њ—А–Є–Ј–љ–∞–Ї–Њ–≤ –љ–∞–ґ–∞—В–Є–є –љ–∞¬†–Ї–љ–Њ–њ–Ї—Г.
–Ф–ї—П –Њ—Ж–µ–љ–Ї–Є –Љ–Њ—Й–љ–Њ—Б—В–љ—Л—Е —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї –±–Є–Њ—Н–ї–µ–Ї—В—А–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –Ї–Њ—А—Л –њ—А–Њ–≤–Њ–і–Є–ї–Є –љ–µ–њ—А–µ—А—Л–≤–љ–Њ–µ –≤–µ–є–≤–ї–µ—В-–њ—А–µ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є–µ –љ–∞¬†–Њ—Б–љ–Њ–≤–µ ¬Ђ–Љ–∞—В–µ—А–Є–љ—Б–Ї–Њ–≥–Њ¬ї –Ї–Њ–Љ–њ–ї–µ–Ї—Б–љ–Њ–≥–Њ Morlet-–≤–µ–є–≤–ї–µ—В–∞ (Matlab 78.01, –і–ї—П —Б–Ї—А–Є–њ—В–Њ–≤ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–Є—Б—М –њ–∞—А–∞–Љ–µ—В—А—Л, –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–љ—Л–µ –≤¬†—А–∞–±–Њ—В–µ C. Tallon-Baudry –Є¬†—Б–Њ–∞–≤—В. [12]). –Ъ–∞—А—В—Л —А–∞—Б–њ—А–µ–і–µ–ї–µ–љ–Є—П –Ј–љ–∞—З–µ–љ–Є–є –Љ–Њ–і—Г–ї—П –Ї–Њ—Н—Д—Д–Є—Ж–Є–µ–љ—В–∞ –≤–µ–є–≤–ї–µ—В-–њ—А–µ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П (–Ъ–Т–Я) —Б—В—А–Њ–Є–ї–Є –≤¬†–њ–Њ–ї–Њ—Б–µ 0,5вАУ40 –У—Ж —Б¬†—И–∞–≥–Њ–Љ 0,5 –У—Ж –Є¬†—А–∞–Ј—А–µ—И–µ–љ–Є–µ–Љ –њ–Њ¬†–≤—А–µ–Љ–µ–љ–Є 0,01 —Б. –Т¬†—В–µ—В–∞- (4вАУ7 –У—Ж), –∞–ї—М—Д–∞-1- (8вАУ10,5 –У—Ж), –∞–ї—М—Д–∞-2- (11вАУ13,5 –У—Ж) –Є¬†–±–µ—В–∞-–і–Є–∞–њ–∞–Ј–Њ–љ–∞—Е (14вАУ21 –У—Ж) —Г—Б—А–µ–і–љ—П–ї–Є –Ъ–Т–Я –њ–Њ¬†—З–∞—Б—В–Њ—В–µ. –Ф–∞–ї–µ–µ –љ–∞¬†–Њ—Б–љ–Њ–≤–∞–љ–Є–Є –≤–Є–Ј—Г–∞–ї—М–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –Љ–µ—Е–∞–љ–Њ–≥—А–∞–Љ–Љ –≤–љ—Г—В—А–Є –Є—Б—Б–ї–µ–і—Г–µ–Љ—Л—Е 30-—Б–µ–Ї—Г–љ–і–љ—Л—Е –Є–љ—В–µ—А–≤–∞–ї–Њ–≤ —Г—Б—А–µ–і–љ—П–ї–Є —Н–њ–Є–Ј–Њ–і—Л —Б¬†–љ–∞–ґ–∞—В–Є—П–Љ–Є –љ–∞¬†–Ї–љ–Њ–њ–Ї—Г –Є¬†–±–µ–Ј –љ–∞–ґ–∞—В–Є–є –њ–Њ¬†–Њ—В–і–µ–ї—М–љ–Њ—Б—В–Є.
–Я–Њ–ї—Г—З–µ–љ–љ—Л–µ –∞–Љ–њ–ї–Є—В—Г–і–љ–Њ-–Љ–Њ—Й–љ–Њ—Б—В–љ—Л–µ —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є –≠–≠–У –∞–љ–∞–ї–Є–Ј–Є—А–Њ–≤–∞–ї–Є —Б¬†–њ–Њ–Љ–Њ—Й—М—О –і–Є—Б–њ–µ—А—Б–Є–Њ–љ–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ (ANOVARM). –†–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–ї–Є –≤–ї–Є—П–љ–Є–µ –љ–∞¬†–љ–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤:
- ¬Ђ—Б–Є—В—Г–∞—Ж–Є—П¬ї¬†вАУ —В—А–Є —Г—А–Њ–≤–љ—П (–љ–∞—З–∞–ї–Њ –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П —В–µ—Б—В–∞ (—Б–Є—В—Г–∞—Ж–Є—П 1), –њ–µ—А–≤–∞—П –Њ—И–Є–±–Ї–∞ –≤–Њ –≤—А–µ–Љ—П –њ–∞—В—В–µ—А–љ–∞ –љ–∞–ґ–∞—В–Є–є (—Б–Є—В—Г–∞—Ж–Є—П 2), –њ–Њ—Б–ї–µ–і–љ–Є–є –њ–∞—В—В–µ—А–љ –љ–∞–ґ–∞—В–Є–є –њ–µ—А–µ–і –Ј–∞—Б—Л–њ–∞–љ–Є–µ–Љ (—Б–Є—В—Г–∞—Ж–Є—П 3));
- ¬Ђ–і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М¬ї¬†вАУ –і–≤–∞ —Г—А–Њ–≤–љ—П (—Б—З–µ—В –≤¬†—Г–Љ–µ —Б¬†–љ–∞–ґ–∞—В–Є—П–Љ–Є –Є¬†–±–µ–Ј –љ–∞–ґ–∞—В–Є–є);
- ¬Ђ–Њ—В–≤–µ–і–µ–љ–Є–µ¬ї¬†вАУ 17 —Г—А–Њ–≤–љ–µ–є (–њ–Њ¬†—З–Є—Б–ї—Г –Њ—В–≤–µ–і–µ–љ–Є–є).
–Р–љ–∞–ї–Є–Ј –Њ—Б—Г—Й–µ—Б—В–≤–ї—П–ї–Є –і–ї—П –Ї–∞–ґ–і–Њ–≥–Њ –Є–Ј¬†–Є—Б—Б–ї–µ–і—Г–µ–Љ—Л—Е —З–∞—Б—В–Њ—В–љ—Л—Е –і–Є–∞–њ–∞–Ј–Њ–љ–Њ–≤. –Т—Б–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л –њ–Њ–ї—Г—З–∞–ї–Є —Б¬†–Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –њ–Њ–њ—А–∞–≤–Ї–Є –У—А–Є–љ—Е–∞—Г–Ј–∞¬†вАУ –У–µ—Б—Б–µ—А–∞. –Ф–ї—П –≤—Л–±—А–∞–љ–љ—Л—Е –љ–∞¬†–Њ—Б–љ–Њ–≤–∞–љ–Є–Є —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤ –і–Є—Б–њ–µ—А—Б–Є–Њ–љ–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ —З–∞—Б—В–Њ—В–љ—Л—Е –і–Є–∞–њ–∞–Ј–Њ–љ–Њ–≤ –і–ї—П –Ї–∞–ґ–і–Њ–≥–Њ –Њ—В–≤–µ–і–µ–љ–Є—П –≠–≠–У —Б¬†–њ–Њ–Љ–Њ—Й—М—О –њ–∞—А–љ–Њ–≥–Њ –Ї—А–Є—В–µ—А–Є—П –°—В—М—О–і–µ–љ—В–∞ –Њ—Ж–µ–љ–Є–≤–∞–ї–Є –Ј–љ–∞—З–Є–Љ–Њ—Б—В—М –Є–Ј–Љ–µ–љ–µ–љ–Є–є –∞–Љ–њ–ї–Є—В—Г–і–љ–Њ-–Љ–Њ—Й–љ–Њ—Б—В–љ—Л—Е —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї –Њ—В–і–µ–ї—М–љ–Њ –і–ї—П –Є–љ—В–µ—А–≤–∞–ї–Њ–≤ —Б¬†–љ–∞–ґ–∞—В–Є—П–Љ–Є –Є¬†–±–µ–Ј –љ–∞–ґ–∞—В–Є–є –Љ–µ–ґ–і—Г —Б–Є—В—Г–∞—Ж–Є—П–Љ–Є 1 –Є¬†2, 2 –Є¬†3, 1 –Є¬†3. –Т—Б–µ —Б—В–∞—В–Є—Б—В–Є—З–µ—Б–Ї–Є–µ –≤—Л—З–Є—Б–ї–µ–љ–Є—П –њ—А–Њ–≤–Њ–і–Є–ї–Є —Б¬†–Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –њ–∞–Ї–µ—В–∞ –њ—А–Њ–≥—А–∞–Љ–Љ SPSS 13.0.
–†–µ–Ј—Г–ї—М—В–∞—В—Л
–Я–∞—В—В–µ—А–љ –љ–∞–ґ–∞—В–Є–є —Б¬†–њ–µ—А–≤–Њ–є –Њ—И–Є–±–Ї–Њ–є –Є¬†–њ–∞—В—В–µ—А–љ –љ–∞–ґ–∞—В–Є–є, –њ—А–µ–і—И–µ—Б—В–≤–Њ–≤–∞–≤—И–Є–є –і–ї–Є—В–µ–ї—М–љ–Њ–Љ—Г (–њ–Њ–ї—В–Њ—А—Л –Љ–Є–љ—Г—В—Л –Є¬†–±–Њ–ї–µ–µ) —Н–њ–Є–Ј–Њ–і—Г —Б–љ–∞, –љ–∞–±–ї—О–і–∞–ї–Є –њ–Њ—Б–ї–µ –љ–∞—З–∞–ї–∞ –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ —З–µ—А–µ–Ј –Ь = 9,37 ¬± 1,29 –Є¬†–Ь = 15,92 ¬± 2,01 –Љ–Є–љ—Г—В—Л —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ. –Я—А–Њ–Љ–µ–ґ—Г—В–Њ–Ї –Љ–µ–ґ–і—Г –љ–Є–Љ–Є —Б–Њ—Б—В–∞–≤–Є–ї –Ь = 6,61 ¬± 1,36 –Љ–Є–љ—Г—В—Л. –Ю–њ—А–µ–і–µ–ї—П–ї–Є –њ–µ—А–Є–Њ–і –≤—А–µ–Љ–µ–љ–Є –њ–Њ—Б–ї–µ–і–љ–µ–≥–Њ –њ–Њ—П–≤–ї–µ–љ–Є—П –∞–і–µ–Ї–≤–∞—В–љ–Њ–≥–Њ –њ–∞—В—В–µ—А–љ–∞ –љ–∞–ґ–∞—В–Є–є, —З—В–Њ–±—Л –Њ—Ж–µ–љ–Є—В—М –њ–µ—А–Є–Њ–і —Б¬†–Ї–Њ–ї–µ–±–∞–љ–Є—П–Љ–Є –≤¬†–њ—А–∞–≤–Є–ї—М–љ–Њ—Б—В–Є –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П –љ–∞–ґ–∞—В–Є–є. –Ю–љ –њ—А–Њ–і–Њ–ї–ґ–∞–ї—Б—П –Ь = 4,98 ¬± 1,16 –Љ–Є–љ—Г—В—Л, –њ–Њ—Б–ї–µ –љ–µ–≥–Њ –љ–∞–±–ї—О–і–∞–ї—Б—П –њ–µ—А–Є–Њ–і —В–Њ–ї—М–Ї–Њ –Њ—И–Є–±–Њ—З–љ—Л—Е –њ–∞—В—В–µ—А–љ–Њ–≤ –Ь = 1,63 ¬± 0,33 –Љ–Є–љ—Г—В—Л (–њ—А–Є–≤–µ–і–µ–љ—Л —Б—А–µ–і–љ–Є–µ –Ј–љ–∞—З–µ–љ–Є—П —Б¬†–Њ—И–Є–±–Ї–Њ–є —Б—А–µ–і–љ–µ–≥–Њ).
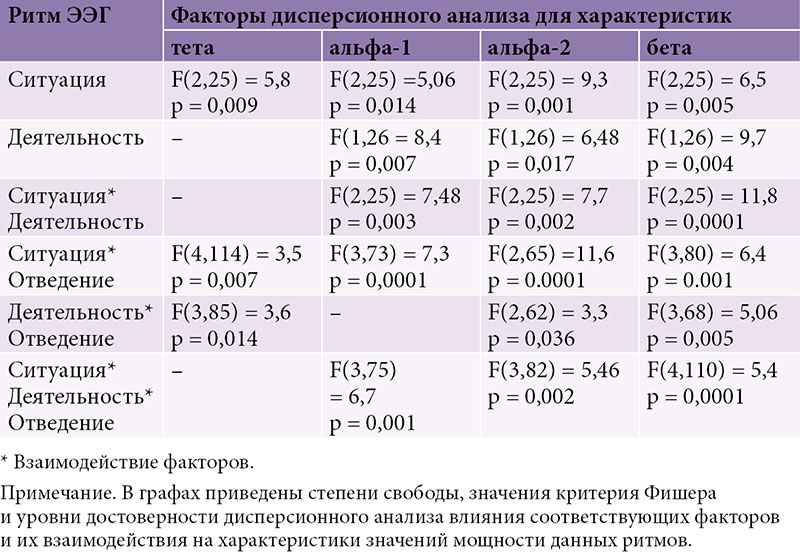
–†–µ–Ј—Г–ї—М—В–∞—В—Л –і–Є—Б–њ–µ—А—Б–Є–Њ–љ–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ (ANOVARM) –∞–Љ–њ–ї–Є—В—Г–і–љ—Л—Е —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї –±–Є–Њ—Н–ї–µ–Ї—В—А–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –≤–Њ –≤—Б–µ—Е –Є—Б—Б–ї–µ–і—Г–µ–Љ—Л—Е –і–Є–∞–њ–∞–Ј–Њ–љ–∞—Е –≠–≠–У –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л –≤¬†—В–∞–±–ї–Є—Ж–µ.
–§–∞–Ї—В–Њ—А ¬Ђ—Б–Є—В—Г–∞—Ж–Є—П¬ї –Њ–Ї–∞–Ј—Л–≤–∞–µ—В –Ј–љ–∞—З–Є–Љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –љ–∞¬†–≤—Б–µ –Є—Б—Б–ї–µ–і—Г–µ–Љ—Л–µ —А–Є—В–Љ—Л –≠–≠–У. –Ш–Ј–Њ–ї–Є—А–Њ–≤–∞–љ–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ —Д–∞–Ї—В–Њ—А–∞ ¬Ђ–і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М¬ї –≤—Л—П–≤–ї–µ–љ–Њ –і–ї—П –Њ–±–Њ–Є—Е –њ–Њ–і–і–Є–∞–њ–∞–Ј–Њ–љ–Њ–≤ –∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤. –°–Њ–≤–Љ–µ—Б—В–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ —Д–∞–Ї—В–Њ—А–Њ–≤ ¬Ђ—Б–Є—В—Г–∞—Ж–Є—П –Є¬†–і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М¬ї —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ –і–ї—П –±–Њ–ї—М—И–Є–љ—Б—В–≤–∞ –Є—Б—Б–ї–µ–і—Г–µ–Љ—Л—Е –і–Є–∞–њ–∞–Ј–Њ–љ–Њ–≤, –Ї—А–Њ–Љ–µ —В–µ—В–∞-—А–Є—В–Љ–∞. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –≤¬†–±–Њ–ї—М—И–Є–љ—Б—В–≤–µ –і–Є–∞–њ–∞–Ј–Њ–љ–Њ–≤ –≤—Л—П–≤–ї–µ–љ–Њ —Б–Њ–≤–Љ–µ—Б—В–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ —Д–∞–Ї—В–Њ—А–∞ ¬Ђ–Њ—В–≤–µ–і–µ–љ–Є–µ¬ї —Б¬†—Д–∞–Ї—В–Њ—А–∞–Љ–Є ¬Ђ–і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М¬ї –Є¬†¬Ђ—Б–Є—В—Г–∞—Ж–Є—П¬ї. –≠—В–Њ—В —Д–∞–Ї—В –і–∞–ї –љ–∞–Љ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –Є—Б—Б–ї–µ–і–Њ–≤–∞—В—М —А–µ–≥–Є–Њ–љ–∞–ї—М–љ—Л–µ —А–∞–Ј–ї–Є—З–Є—П –Є–Ј–Љ–µ–љ–µ–љ–Є–є –≤¬†—А–Є—В–Љ–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –≤¬†–Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В¬†—Б–Є—В—Г–∞—Ж–Є–Є –Є¬†—Д–∞–Ј—Л –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞.
–°—В–∞—В–Є—Б—В–Є—З–µ—Б–Ї–∞—П –Њ—Ж–µ–љ–Ї–∞ –Є–Ј–Љ–µ–љ–µ–љ–Є–є –Љ–Њ—Й–љ–Њ—Б—В–љ—Л—Е —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї —А–Є—В–Љ–Њ–≤ –≠–≠–У –њ–Њ¬†–Њ—В–≤–µ–і–µ–љ–Є—П–Љ –Љ–µ–ґ–і—Г —Б–Њ—Б—В–Њ—П–љ–Є–µ–Љ –њ–Њ–ї–љ–Њ–є –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є –Є¬†–њ–µ—А–≤—Л–Љ –Њ—И–Є–±–Њ—З–љ—Л–Љ —Н–њ–Є–Ј–Њ–і–Њ–Љ –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ –њ—А–Є–≤–µ–і–µ–љ–∞ –љ–∞¬†—А–Є—Б. 1. –Т¬†–Ї–∞—Г–і–∞–ї—М–љ—Л—Е –Њ–±–ї–∞—Б—В—П—Е –Њ—В–≤–µ–і–µ–љ–Є—П –≠–≠–У –њ–µ—А–Є–Њ–і –љ–∞–ґ–∞—В–Є–є –љ–∞¬†–Ї–љ–Њ–њ–Ї—Г —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–µ—В—Б—П —А–Њ—Б—В–Њ–Љ –Љ–Њ—Й–љ–Њ—Б—В–Є —В–µ—В–∞-—А–Є—В–Љ–∞ –Є¬†—Г–Љ–µ–љ—М—И–µ–љ–Є–µ–Љ –∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–∞. –Ф–ї—П —Б–Є—В—Г–∞—Ж–Є–Є —Б—З–µ—В–∞ –≤¬†—Г–Љ–µ –±–µ–Ј –љ–∞–ґ–∞—В–Є–є –њ–Њ–Ї–∞–Ј–∞–љ–Њ¬†–≥–µ–љ–µ—А–∞–ї–Є–Ј–Њ–≤–∞–љ–љ–Њ–µ —Б–љ–Є–ґ–µ–љ–Є–µ –Љ–Њ—Й–љ–Њ—Б—В–Є –≤¬†–∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-–і–Є–∞–њ–∞–Ј–Њ–љ–µ –≠–≠–У.
–Э–∞¬†—А–Є—Б. 2 –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л —А–∞–Ј–ї–Є—З–Є—П –Љ–µ–ґ–і—Г —Б–Є—В—Г–∞—Ж–Є—П–Љ–Є –њ–Њ–ї–љ–Њ–є –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є –Є¬†–њ–Њ—Б–ї–µ–і–љ–Є–Љ —Н–њ–Є–Ј–Њ–і–Њ–Љ –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є –њ–µ—А–µ–і –Ј–∞—Б—Л–њ–∞–љ–Є–µ–Љ. –Т¬†–њ–µ—А–Є–Њ–і–µ —Б¬†–љ–∞–ґ–∞—В–Є—П–Љ–Є —В–µ—В–∞-—А–Є—В–Љ –≤–Њ–Ј—А–∞—Б—В–∞–µ—В –≤¬†—Ж–µ–љ—В—А–∞–ї—М–љ—Л—Е, –њ–µ—А–µ–і–љ–µ–≤–Є—Б–Њ—З–љ—Л—Е –Є¬†–Ї–∞—Г–і–∞–ї—М–љ—Л—Е –Њ–±–ї–∞—Б—В—П—Е. –Р–ї—М—Д–∞-1- –Є¬†–±–µ—В–∞-—А–Є—В–Љ—Л –љ–µ–Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ —Г–≤–µ–ї–Є—З–Є–≤–∞—О—В—Б—П –≤¬†–њ–µ—А–µ–і–љ–µ—Ж–µ–љ—В—А–∞–ї—М–љ—Л—Е –Њ–±–ї–∞—Б—В—П—Е –Ї–Њ—А—Л. –Р–ї—М—Д–∞-2 —Б–љ–Є–ґ–∞–µ—В—Б—П –≤¬†–Ј–∞—В—Л–ї–Њ—З–љ—Л—Е –Є¬†–љ–Є–ґ–љ–µ–≤–Є—Б–Њ—З–љ—Л—Е –Ј–Њ–љ–∞—Е. –°—З–µ—В –≤¬†—Г–Љ–µ –±–µ–Ј –љ–∞–ґ–∞—В–Є–є —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–µ—В—Б—П —А–Њ—Б—В–Њ–Љ –∞–ї—М—Д–∞-1-, —Б–љ–Є–ґ–µ–љ–Є–µ–Љ –∞–ї—М—Д–∞-2- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤. –≠—В–Є —А–∞–Ј–ї–Є—З–Є—П –љ–∞–±–ї—О–і–∞—О—В—Б—П –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –њ–Њ¬†–≤—Б–µ–є –Ї–Њ—А–µ.
–†–µ–Ј—Г–ї—М—В–∞—В—Л –∞–љ–∞–ї–Є–Ј–∞ –Є–Ј–Љ–µ–љ–µ–љ–Є–є –Љ–Њ—Й–љ–Њ—Б—В–љ—Л—Е —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї –≠–≠–У –Љ–µ–ґ–і—Г —Б–Є—В—Г–∞—Ж–Є–µ–є –њ–µ—А–≤–Њ–є –Њ—И–Є–±–Ї–Є –Є—Б–њ—Л—В—Г–µ–Љ–Њ–≥–Њ –Є¬†—Б–Є—В—Г–∞—Ж–Є–µ–є –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ –њ–µ—А–µ–і –Ј–∞—Б—Л–њ–∞–љ–Є–µ–Љ –њ—А–Є–≤–µ–і–µ–љ—Л –љ–∞¬†—А–Є—Б. 3. –†–∞–Ј–ї–Є—З–Є—П –Њ–±–љ–∞—А—Г–ґ–µ–љ—Л —В–Њ–ї—М–Ї–Њ –њ—А–Є –љ–∞–ґ–∞—В–Є—П—Е –љ–∞¬†–Ї–љ–Њ–њ–Ї—Г. –†–Њ—Б—В –Љ–Њ—Й–љ–Њ—Б—В–Є –∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤ –њ–Њ–Ї–∞–Ј–∞–љ –≤¬†–њ–µ—А–µ–і–љ–µ—Ж–µ–љ—В—А–∞–ї—М–љ—Л—Е –Њ–±–ї–∞—Б—В—П—Е –Є¬†—В–µ–Љ–µ–љ–љ–Њ–є –Ї–Њ—А–µ.
–Ю–±—Б—Г–ґ–і–µ–љ–Є–µ
–Ь–Њ—Й–љ–Њ—Б—В–љ—Л–µ —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є —В–µ—В–∞-, –∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤ –Є–Ј–Љ–µ–љ—П—О—В—Б—П –≤¬†–Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В¬†–і–≤—Г—Е –Є—Б—Б–ї–µ–і—Г–µ–Љ—Л—Е —Д–∞–Ї—В–Њ—А–Њ–≤: —Б–ї–Њ–ґ–љ–Њ—Б—В–Є –Ј–∞–і–∞–љ–Є–є –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ (—Б—З–µ—В–∞ –≤¬†—Г–Љ–µ —Б¬†—Б–Є–љ—Е—А–Њ–љ–љ—Л–Љ–Є –љ–∞–ґ–∞—В–Є—П–Љ–Є –љ–∞¬†–Ї–љ–Њ–њ–Ї—Г –Є¬†–±–µ–Ј –љ–∞–ґ–∞—В–Є–є) –Є¬†–њ–µ—А–Є–Њ–і–Њ–≤ –Ј–∞—Б—Л–њ–∞–љ–Є—П, –Ї–Њ—В–Њ—А—Л–µ —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—В—Б—П —А–∞–Ј–љ–Њ–є —Б—В–µ–њ–µ–љ—М—О —Б–Њ–љ–ї–Є–≤–Њ—Б—В–Є –Є¬†–±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є.
–≠—В–Є –Ј–∞–і–∞–љ–Є—П –≤—Л–њ–Њ–ї–љ—П–ї–Є—Б—М –≤¬†–Ј–∞—В–µ–Љ–љ–µ–љ–љ–Њ–Љ –њ–Њ–Љ–µ—Й–µ–љ–Є–Є –≤¬†–њ–Њ–ї–Њ–ґ–µ–љ–Є–Є –ї–µ–ґ–∞ —Б¬†–Ј–∞–Ї—А—Л—В—Л–Љ–Є¬†–≥–ї–∞–Ј–∞–Љ–Є, –њ–Њ—Н—В–Њ–Љ—Г –Љ—Л –њ—А–µ–і–њ–Њ–ї–∞–≥–∞–ї–Є, —З—В–Њ —Г¬†–±–Њ–ї—М—И–Є–љ—Б—В–≤–∞ –Њ–±—Б–ї–µ–і—Г–µ–Љ—Л—Е —Г—А–Њ–≤–µ–љ—М –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є –њ–Њ¬†–Љ–µ—А–µ –Њ—Б—Г—Й–µ—Б—В–≤–ї–µ–љ–Є—П –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є –±—Г–і–µ—В –±—Л—Б—В—А–Њ —Б–љ–Є–ґ–∞—В—М—Б—П, —Г–≤–µ–ї–Є—З–Є—В—Б—П —Б–Њ–љ–ї–Є–≤–Њ—Б—В—М, –≤–њ–ї–Њ—В—М –і–Њ –Ј–∞—Б—Л–њ–∞–љ–Є—П, —В–∞–Ї–Є–µ –Є–Ј–Љ–µ–љ–µ–љ–Є—П –Њ—В—А–∞–Ј—П—В—Б—П –Ї–∞–Ї –≤¬†–њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є—Е, —В–∞–Ї –Є¬†–≤ –±–Є–Њ—Н–ї–µ–Ї—В—А–Є—З–µ—Б–Ї–Є—Е —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–∞—Е. –Э–∞—И–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –њ–Њ–і—В–≤–µ—А–і–Є–ї–Њ —Н—В–Є –њ—А–µ–і–њ–Њ–ї–Њ–ґ–µ–љ–Є—П. –Т¬†—Б–Є—В—Г–∞—Ж–Є–Є –њ–µ—А–≤–Њ–є –Њ—И–Є–±–Ї–Є –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–є –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є (—Г–Љ–µ–љ—М—И–µ–љ–Є–µ —З–Є—Б–ї–∞ –љ–∞–ґ–∞—В–Є–є –≤¬†–њ–∞—В—В–µ—А–љ–µ) –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М—О –≤¬†–љ–∞—З–∞–ї–µ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞ –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –Љ–Њ—Й–љ–Њ—Б—В–Є —В–µ—В–∞-–Ї–Њ–ї–µ–±–∞–љ–Є–є, —Б–љ–Є–ґ–µ–љ–Є–µ –∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤. –Я—А–Є —Б—З–µ—В–µ –Є¬†—Б–Є–љ—Е—А–Њ–љ–љ—Л—Е –љ–∞–ґ–∞—В–Є—П—Е —Г–≤–µ–ї–Є—З–µ–љ–Є–µ —В–µ—В–∞-—А–Є—В–Љ–∞ –Є¬†—Б–љ–Є–ґ–µ–љ–Є–µ –∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤ –љ–Њ—Б–Є–ї–Є –ї–Њ–Ї–∞–ї—М–љ—Л–є —Е–∞—А–∞–Ї—В–µ—А –Є¬†–Њ–±–љ–∞—А—Г–ґ–Є–≤–∞–ї–Є—Б—М –≤¬†–Ї–∞—Г–і–∞–ї—М–љ—Л—Е –Њ–±–ї–∞—Б—В—П—Е –Ї–Њ—А—Л. –Я—А–Є –Љ—Л—Б–ї–µ–љ–љ–Њ–Љ —Б—З–µ—В–µ –±–µ–Ј –љ–∞–ґ–∞—В–Є–є, –Ї–Њ—В–Њ—А—Л–є —Б–ї–µ–і–Њ–≤–∞–ї –Ј–∞ –њ–∞—В—В–µ—А–љ–Њ–Љ –љ–∞–ґ–∞—В–Є–є —Б¬†–Њ—И–Є–±–Ї–Њ–є, –Є–Ј–Љ–µ–љ–µ–љ–Є—П –∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤ –љ–Њ—Б–Є–ї–Є¬†–≥–µ–љ–µ—А–∞–ї–Є–Ј–Њ–≤–∞–љ–љ—Л–є —Е–∞—А–∞–Ї—В–µ—А. –°–љ–Є–ґ–µ–љ–Є–µ –Љ–Њ—Й–љ–Њ—Б—В–Є –≤—Л—Б–Њ–Ї–Њ—З–∞—Б—В–Њ—В–љ—Л—Е —Б–Њ—Б—В–∞–≤–ї—П—О—Й–Є—Е –≠–≠–У —Б¬†–Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ—Л–Љ —А–Њ—Б—В–Њ–Љ —В–µ—В–∞-—А–Є—В–Љ–∞ —Б–≤—П–Ј–∞–љ–Њ —Б¬†—Г—Б–Є–ї–µ–љ–Є–µ–Љ —Б–Њ–љ–ї–Є–≤–Њ—Б—В–Є [13вАУ15]. –Я–Њ—П–≤–ї–µ–љ–Є–µ –Њ—И–Є–±–Ї–Є (—Г–Љ–µ–љ—М—И–µ–љ–Є–µ —З–Є—Б–ї–∞ –љ–∞–ґ–∞—В–Є–є –≤¬†–њ–∞—В—В–µ—А–љ–µ) –Љ–Њ–ґ–љ–Њ —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞—В—М –Ї–∞–Ї –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–є –Љ–∞—А–Ї–µ—А —Б–љ–Є–ґ–µ–љ–Є—П —Г—А–Њ–≤–љ—П –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є, –Є¬†–њ—А–Є –≤—Л–њ–Њ–ї–љ–µ–љ–Є–Є –±–Њ–ї–µ–µ –њ—А–Њ—Б—В–Њ–є –Ј–∞–і–∞—З–Є (–Љ—Л—Б–ї–µ–љ–љ—Л–є —Б—З–µ—В –±–µ–Ј –љ–∞–ґ–∞—В–Є–є) —Н—В–Њ —П–≤–ї–µ–љ–Є–µ –±–Њ–ї–µ–µ –≤—Л—А–∞–ґ–µ–љ–Њ. –Ф–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ–∞—П –Љ–Њ—В–Њ—А–љ–∞—П –і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М —В—А–µ–±—Г–µ—В –±–Њ–ї—М—И–µ–є –∞–Ї—В–Є–≤–∞—Ж–Є–Є. –Я—А–Є —Н—В–Њ–Љ –≠–≠–У-—Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є –љ–∞—З–∞–ї—М–љ–Њ–є —Б—В–∞–і–Є–Є –Ј–∞—Б—Л–њ–∞–љ–Є—П –љ–Њ—Б—П—В –ї–Њ–Ї–∞–ї—М–љ—Л–є —Е–∞—А–∞–Ї—В–µ—А –Є¬†–њ—А–Њ—П–≤–ї—П—О—В—Б—П –≤¬†–Љ–µ–љ—М—И–µ–Љ —З–Є—Б–ї–µ –Њ—В–≤–µ–і–µ–љ–Є–є.
–Я—А–Є —Б—З–µ—В–µ –≤¬†—Г–Љ–µ –њ–µ—А–µ–і –њ–∞—В—В–µ—А–љ–Њ–Љ —Б¬†–љ–∞–ґ–∞—В–Є—П–Љ–Є, –Ї–Њ—В–Њ—А—Л–є –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –њ—А–µ–і—И–µ—Б—В–≤–Њ–≤–∞–ї –њ–µ—А–Є–Њ–і—Г —Б–љ–∞, –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–∞–љ–∞–ї–Њ–≥–Є—З–љ–Њ–є –і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М—О –≤¬†–љ–∞—З–∞–ї–µ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞ –љ–∞–±–ї—О–і–∞–µ—В—Б—П¬†–≥–µ–љ–µ—А–∞–ї–Є–Ј–Њ–≤–∞–љ–љ—Л–є —А–Њ—Б—В –∞–ї—М—Д–∞-1- –Є¬†—Б–љ–Є–ґ–µ–љ–Є–µ –∞–ї—М—Д–∞-2- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤. –Я–Њ—Б–ї–µ–і–љ–Є–є —Н–њ–Є–Ј–Њ–і –Љ–Њ—В–Њ—А–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –њ–µ—А–µ–і –Ј–∞—Б—Л–њ–∞–љ–Є–µ–Љ —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–µ—В—Б—П –ї–Њ–Ї–∞–ї—М–љ—Л–Љ —Г–≤–µ–ї–Є—З–µ–љ–Є–µ–Љ –Љ–Њ—Й–љ–Њ—Б—В–Є —В–µ—В–∞-, –∞–ї—М—Д–∞-1- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤ –Є¬†—Г–Љ–µ–љ—М—И–µ–љ–Є–µ–Љ –∞–ї—М—Д–∞-2. –£–≤–µ–ї–Є—З–µ–љ–Є–µ –Љ–Њ—Й–љ–Њ—Б—В–љ—Л—Е —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї —В–µ—В–∞-, –∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤ –љ–∞¬†–≠–≠–У —П–≤–ї—П–µ—В—Б—П –Љ–∞—А–Ї–µ—А–Њ–Љ —Б–Њ—Б—В–Њ—П–љ–Є—П –Љ–Њ–љ–Њ—В–Њ–љ–Є–Є [5]. –£–≤–µ–ї–Є—З–µ–љ–Є–µ —В–µ—В–∞-, –∞–ї—М—Д–∞-1- —Б¬†–Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ—Л–Љ —Б–љ–Є–ґ–µ–љ–Є–µ–Љ –∞–ї—М—Д–∞-2-—А–Є—В–Љ–Њ–≤ —Б–≤—П–Ј–∞–љ–Њ —Б¬†—Б–Њ—Б—В–Њ—П–љ–Є–µ–Љ –Љ–Є–Ї—А–Њ—Б–љ–∞ —Б¬†–Њ—В–Ї—А—Л—В—Л–Љ–Є¬†–≥–ї–∞–Ј–∞–Љ–Є –њ—А–Є –≤—Л–њ–Њ–ї–љ–µ–љ–Є–Є –Љ–Њ–љ–Њ—В–Њ–љ–љ–Њ–є –Ј—А–Є—В–µ–ї—М–љ–Њ-–њ—А–Њ—Б—В—А–∞–љ—Б—В–≤–µ–љ–љ–Њ–є –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є [16]. –£–≤–µ–ї–Є—З–µ–љ–Є–µ –Љ–Њ—Й–љ–Њ—Б—В–Є —В–µ—В–∞- –Є¬†–∞–ї—М—Д–∞-—А–Є—В–Љ–Њ–≤ –≤–Њ –≤—А–µ–Љ—П –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є –Љ–Њ–ґ–µ—В –±—Л—В—М –Њ–±—Г—Б–ї–Њ–≤–ї–µ–љ–Њ –Є¬†–і—А—Г–≥–Є–Љ–Є —Д–∞–Ї—В–Њ—А–∞–Љ–Є, —Б–≤—П–Ј–∞–љ–љ—Л–Љ–Є —Б¬†–µ–µ —Б–њ–µ—Ж–Є—Д–Є–Ї–Њ–є. –£—Б–Є–ї–µ–љ–Є–µ —В–µ—В–∞-–∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –љ–∞–±–ї—О–і–∞–µ—В—Б—П –њ—А–Є —Г–≤–µ–ї–Є—З–µ–љ–Є–Є —Г–Љ—Б—В–≤–µ–љ–љ–Њ–є –љ–∞–≥—А—Г–Ј–Ї–Є [17]. –†–Њ—Б—В –∞–ї—М—Д–∞-—А–Є—В–Љ–∞ –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞–µ—В—Б—П –њ—А–Є –≤—Л–њ–Њ–ї–љ–µ–љ–Є–Є –Њ–±—А–∞—В–љ–Њ–≥–Њ —Б—З–µ—В–∞ –≤¬†—Г–Љ–µ [18]. –Я–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –±–µ—В–∞-–∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –љ–∞¬†–≠–≠–У –Ї–Њ—А—А–µ–ї–Є—А—Г–µ—В —Б¬†—Г—Б–Є–ї–µ–љ–Є–µ–Љ –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є –Є¬†—Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М—О –ї—Г—З—И–µ –Њ—Б—Г—Й–µ—Б—В–≤–ї—П—В—М –≤—Л—Б—И–Є–µ –Ї–Њ–≥–љ–Є—В–Є–≤–љ—Л–µ —Д—Г–љ–Ї—Ж–Є–Є [19].
–Р–љ–∞–ї–Є–Ј–Є—А—Г–µ–Љ—Л–µ –Љ–Њ—Й–љ–Њ—Б—В–љ—Л–µ —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є –≠–≠–У –њ–∞—В—В–µ—А–љ–∞ —Б—З–µ—В–∞ –≤¬†—Г–Љ–µ –±–µ–Ј –љ–∞–ґ–∞—В–Є–є –њ—А–Є –њ–µ—А–≤–Њ–є –Њ—И–Є–±–Ї–µ –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ –Є¬†–њ–Њ—Б–ї–µ–і–љ–µ–≥–Њ, –Њ—И–Є–±–Њ—З–љ–Њ–≥–Њ, —Н–њ–Є–Ј–Њ–і–∞ –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є –њ–µ—А–µ–і —Б–љ–Њ–Љ¬†вАУ —Б—В–∞—В–Є—Б—В–Є—З–µ—Б–Ї–Є –Ј–љ–∞—З–Є–Љ–Њ –љ–µ¬†—А–∞–Ј–ї–Є—З–∞–ї–Є—Б—М. –Я—А–Є –њ–∞—В—В–µ—А–љ–∞—Е —Б¬†–љ–∞–ґ–∞—В–Є—П–Љ–Є –≤¬†—Н—В–Є—Е –Њ–±—Б—В–Њ—П—В–µ–ї—М—Б—В–≤–∞—Е –њ–Њ–Ї–∞–Ј–∞–љ–Њ —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –∞–ї—М—Д–∞-1-, –∞–ї—М—Д–∞-2- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤ –≤¬†–ї–Њ–±–љ–Њ-—Ж–µ–љ—В—А–∞–ї—М–љ—Л—Е –Є¬†—В–µ–Љ–µ–љ–љ—Л—Е –Њ–±–ї–∞—Б—В—П—Е –Ї–Њ—А—Л. –Я–µ—А–Є–Њ–і –Њ—В¬†–њ–µ—А–≤–Њ–є –Ї¬†–њ–Њ—Б–ї–µ–і–љ–µ–є –Њ—И–Є–±–Ї–µ –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П —В–µ—Б—В–∞ –≤–Ї–ї—О—З–∞–ї –≤¬†—Б–µ–±—П –њ–∞—В—В–µ—А–љ—Л –Ї–∞–Ї —Б¬†–і–µ—Б—П—В—М—О –љ–∞–ґ–∞—В–Є—П–Љ–Є, —В–∞–Ї –Є¬†—Б –Љ–µ–љ—М—И–Є–Љ —З–Є—Б–ї–Њ–Љ –љ–∞–ґ–∞—В–Є–є. –Я—А–Є —Н—В–Њ–Љ –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –≤—А–µ–Љ–µ–љ–љ—Л—Е –Є–љ—В–µ—А–≤–∞–ї–Њ–≤ –Љ–µ–ґ–і—Г –љ–∞–ґ–∞—В–Є—П–Љ–Є. –Ь—Л –љ–µ¬†–Љ–Њ–ґ–µ–Љ –Њ–і–љ–Њ–Ј–љ–∞—З–љ–Њ —Г—В–≤–µ—А–ґ–і–∞—В—М, —З—В–Њ –≤¬†—Н—В–Є –Є–љ—В–µ—А–≤–∞–ї—Л –Є—Б–њ—Л—В—Г–µ–Љ—Л–µ –≤—Л–њ–Њ–ї–љ—П–ї–Є –Ї–Њ–≥–љ–Є—В–Є–≤–љ—Г—О –і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М, –∞¬†–љ–µ –љ–∞—Е–Њ–і–Є–ї–Є—Б—М –≤¬†—Б–Њ—Б—В–Њ—П–љ–Є–Є –Љ–Є–Ї—А–Њ—Б–љ–∞ –Є¬†–њ–Њ—Б–ї–µ–і—Г—О—Й–Є–µ –њ–∞—В—В–µ—А–љ—Л –љ–∞–ґ–∞—В–Є–є –љ–µ¬†–Є—Б–њ—Л—В—Л–≤–∞–ї–Є –≤–ї–Є—П–љ–Є—П –Є–љ–µ—А—Ж–Є–Є —Б–љ–∞. –Т¬†–њ–Њ–ї—М–Ј—Г –≤–ї–Є—П–љ–Є—П —Д–∞–Ї—В–Њ—А–∞ –Ј–∞—Б—Л–њ–∞–љ–Є—П¬†–≥–Њ–≤–Њ—А–Є—В —Д–∞–Ї—В —Б–Љ–µ—Й–µ–љ–Є—П –∞–ї—М—Д–∞-—А–Є—В–Љ–∞ –Є–Ј¬†–Ї–∞—Г–і–∞–ї—М–љ—Л—Е –Њ–±–ї–∞—Б—В–µ–є –≤¬†–њ–µ—А–µ–і–љ–Є–µ [1]. –Э–∞—А—П–і—Г —Б¬†—Н—В–Є–Љ –њ–Њ–Ї–∞–Ј–∞–љ–Њ —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –±–µ—В–∞-–∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –њ–Њ—Б–ї–µ —Н–њ–Є–Ј–Њ–і–∞ –Љ–Є–Ї—А–Њ—Б–љ–∞ [15]. –≠—В–Њ—В —А–µ–Ј—Г–ї—М—В–∞—В –Љ–Њ–ґ–µ—В –Њ—В—А–∞–ґ–∞—В—М –њ–Њ–і—Б–Њ–Ј–љ–∞—В–µ–ї—М–љ–Њ–µ —Б—В—А–µ–Љ–ї–µ–љ–Є–µ –Љ–Њ–Ј–≥–∞ –≤–Њ—Б—Б—В–∞–љ–Њ–≤–Є—В—М —Б–Њ–Ј–љ–∞–љ–Є–µ –Є –њ–µ—А—Ж–µ–њ—В–Є–≤–љ—Г—О —Б–≤—П–Ј—М —Б¬†–≤–љ–µ—И–љ–µ–є —Б—А–µ–і–Њ–є, –∞ —В–∞–Ї–ґ–µ –њ–Њ–≤—В–Њ—А–љ–Њ —Б–Є–љ—Е—А–Њ–љ–Є–Ј–Є—А–Њ–≤–∞—В—М –≤–љ–Є–Љ–∞–љ–Є–µ –Є¬†–њ–∞–Љ—П—В—М –і–ї—П –≤–Њ–Ј–Њ–±–љ–Њ–≤–ї–µ–љ–Є—П –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є. –Т¬†–љ–∞—И–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П—Е –њ–Њ–Ї–∞–Ј–∞–љ —А–Њ—Б—В –∞–ї—М—Д–∞-1- –Є¬†–∞–ї—М—Д–∞-2-—А–Є—В–Љ–Њ–≤ –≤¬†–њ–µ—А–µ–і–љ–µ—Ж–µ–љ—В—А–∞–ї—М–љ—Л—Е –Њ–±–ї–∞—Б—В—П—Е –њ—А–Є –≤—Л–њ–Њ–ї–љ–µ–љ–Є–Є –Ј–∞–і–∞–љ–Є–є –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ –њ–Њ—Б–ї–µ –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П –Є–Ј¬†–±–Њ–ї–µ–µ –і–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ (—Б–≤—Л—И–µ –њ–Њ–ї—Г—В–Њ—А–∞ –Љ–Є–љ—Г—В) –і–љ–µ–≤–љ–Њ–≥–Њ —Б–љ–∞. –Я–Њ–і–Њ–±–љ–Њ–µ —П–≤–ї–µ–љ–Є–µ –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М, —В–Њ–ї—М–Ї–Њ –Ї–Њ–≥–і–∞ –Њ–±—Б–ї–µ–і—Г–µ–Љ—Л–µ –љ–∞—З–Є–љ–∞–ї–Є –≤–Њ—Б—Б—В–∞–љ–∞–≤–ї–Є–≤–∞—В—М —Н—В—Г –і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М —Б¬†–Њ—И–Є–±–Ї–∞–Љ–Є [20]. –Ю–±—Б–ї–µ–і—Г–µ–Љ—Л–є –њ—А–∞–≤–Є–ї—М–љ–Њ –њ—А–Є–њ–Њ–Љ–Є–љ–∞–µ—В –Є–љ—Б—В—А—Г–Ї—Ж–Є—О (¬Ђ–љ–∞–ґ–Є–Љ–∞—В—М¬†вАУ —Б—З–Є—В–∞—В—М¬ї) –≤¬†—Н—В–Є—Е –Њ–±—Б—В–Њ—П—В–µ–ї—М—Б—В–≤–∞—Е, –љ–Њ¬†–љ–µ –≤¬†—Б–Њ—Б—В–Њ—П–љ–Є–Є –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е –Є–љ–µ—А—Ж–Є–Є —Б–љ–∞, —Б–љ–Є–ґ–µ–љ–љ–Њ–≥–Њ —Г—А–Њ–≤–љ—П –Ї–Њ–≥–љ–Є—В–Є–≤–љ–Њ–≥–Њ –Ї–Њ–љ—В—А–Њ–ї—П –Є¬†–Њ—Б–Њ–Ј–љ–∞–љ–Є—П –њ–Њ–ї–љ–Њ—Б—В—М—О –µ–µ —А–µ–∞–ї–Є–Ј–Њ–≤–∞—В—М. –Я—А–Є —З–µ—А–µ–і–Њ–≤–∞–љ–Є–Є —Н–њ–Є–Ј–Њ–і–Њ–≤ –њ—А–∞–≤–Є–ї—М–љ–Њ–≥–Њ –Є¬†–Њ—И–Є–±–Њ—З–љ–Њ–≥–Њ –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П –њ—Б–Є—Е–Њ–Љ–Њ—В–Њ—А–љ–Њ–≥–Њ —В–µ—Б—В–∞ –њ—А–Є –Ј–∞—Б—Л–њ–∞–љ–Є–Є, –Ї–∞–Ї –Љ—Л –њ—А–µ–і–њ–Њ–ї–∞–≥–∞–µ–Љ, —Б—Г–±—К–µ–Ї—В –Є–Ј-–Ј–∞ –Ї–Њ–ї–µ–±–∞–љ–Є–є —Г—А–Њ–≤–љ—П –∞–Ї—В–Є–≤–∞—Ж–Є–Є –љ–µ¬†–≤—Б–µ–≥–і–∞ –Љ–Њ–ґ–µ—В —В–Њ—З–љ–Њ —Б–ї–µ–і–Њ–≤–∞—В—М –Є–љ—Б—В—А—Г–Ї—Ж–Є–Є. –Ъ—А–∞–є–љ–Є–є –њ–∞—В—В–µ—А–љ –љ–∞–ґ–∞—В–Є–є —Б¬†–Њ—И–Є–±–Ї–Њ–є, –Ј–∞ –Ї–Њ—В–Њ—А—Л–Љ —Б–ї–µ–і—Г–µ—В –і–ї–Є—В–µ–ї—М–љ—Л–є —Н–њ–Є–Ј–Њ–і –њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞,¬†вАУ –љ–µ¬†–Є—Б–Ї–ї—О—З–µ–љ–Є–µ.
–Ч–∞–Ї–ї—О—З–µ–љ–Є–µ
–Т¬†—А–∞–Ј–љ—Л–µ –њ–µ—А–Є–Њ–і—Л –Ј–∞—Б—Л–њ–∞–љ–Є—П –≤–Њ –≤—А–µ–Љ—П –і–љ–µ–≤–љ–Њ–≥–Њ —Б–љ–∞ —Г—А–Њ–≤–µ–љ—М –∞–Ї—В–Є–≤–∞—Ж–Є–Є –Љ–Њ–Ј–≥–Њ–≤—Л—Е –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ –њ—А–Є –Њ—Б—Г—Й–µ—Б—В–≤–ї–µ–љ–Є–Є –і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є —А–∞–Ј–љ–Њ–є —Б–ї–Њ–ґ–љ–Њ—Б—В–Є –Є–Ј–Љ–µ–љ—П–ї—Б—П –њ–Њ-—А–∞–Ј–љ–Њ–Љ—Г. –Э–∞¬†—А–∞–љ–љ–µ–є —Б—В–∞–і–Є–Є –≤—Л–њ–Њ–ї–љ–µ–љ–Є–µ –њ—А–Њ—Б—В–Њ–є –Ј–∞–і–∞—З–Є (—Б—З–µ—В –≤¬†—Г–Љ–µ –Њ—В¬†–Њ–і–љ–Њ–≥–Њ –і–Њ –і–µ—Б—П—В–Є) —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–ї–Њ—Б—М¬†–≥–µ–љ–µ—А–∞–ї–Є–Ј–Њ–≤–∞–љ–љ—Л–Љ —Б–љ–Є–ґ–µ–љ–Є–µ–Љ –∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤. –Я—А–Є —Н—В–Њ–Љ –±–Њ–ї–µ–µ —Б–ї–Њ–ґ–љ–∞—П –Ј–∞–і–∞—З–∞ (–љ–∞–ґ–Є–Љ–∞—В—М –Є¬†—Б—З–Є—В–∞—В—М), –≤—Л–њ–Њ–ї–љ–µ–љ–љ–∞—П —Б¬†–Њ—И–Є–±–Ї–Њ–є, –≤—Л–Ј—Л–≤–∞–ї–∞ –Є—Е —Б–љ–Є–ґ–µ–љ–Є–µ —В–Њ–ї—М–Ї–Њ –≤¬†–Ї–∞—Г–і–∞–ї—М–љ—Л—Е –Њ–±–ї–∞—Б—В—П—Е –Њ—В–≤–µ–і–µ–љ–Є—П –≠–≠–У. –Э–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –њ–µ—А–µ–і —Н–њ–Є–Ј–Њ–і–Њ–Љ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –і–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ —Б–љ–∞ (–Њ—В¬†–њ–Њ–ї—Г—В–Њ—А–∞ –Љ–Є–љ—Г—В –Є¬†–±–Њ–ї–µ–µ) –Љ–Њ—Й–љ–Њ—Б—В–љ—Л–µ —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є –≠–≠–У –њ—А–Њ—Б—В–Њ–є –Ј–∞–і–∞—З–Є –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†—Н–њ–Є–Ј–Њ–і–Њ–Љ –њ–µ—А–≤–Њ–є –Љ–Њ—В–Њ—А–љ–Њ–є –Њ—И–Є–±–Ї–Є –љ–µ¬†–Є–Ј–Љ–µ–љ–Є–ї–Є—Б—М. –Т—Л–њ–Њ–ї–љ–µ–љ–Є–µ –±–Њ–ї–µ–µ —Б–ї–Њ–ґ–љ–Њ–є –Ј–∞–і–∞—З–Є –њ—А–Є —Н—В–Њ–Љ —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–ї–Њ—Б—М —А–Њ—Б—В–Њ–Љ –∞–ї—М—Д–∞- –Є¬†–±–µ—В–∞-—А–Є—В–Љ–Њ–≤ –≤¬†–њ–µ—А–µ–і–љ–µ—Ж–µ–љ—В—А–∞–ї—М–љ—Л—Е –Њ–±–ї–∞—Б—В—П—Е –Є¬†—В–µ–Љ–µ–љ–љ–Њ–є –Ї–Њ—А–µ. –Ь—Л –њ—А–µ–і–њ–Њ–ї–∞–≥–∞–µ–Љ, —З—В–Њ –њ—А–Є —Г–≥–ї—Г–±–ї–µ–љ–Є–Є —Б–Њ–љ–ї–Є–≤–Њ—Б—В–Є –Є¬†—Б–љ–Є–ґ–µ–љ–Є–Є –±–і–Є—В–µ–ї—М–љ–Њ—Б—В–Є –њ–µ—А–Є–Њ–і–Є—З–µ—Б–Ї–∞—П –і–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ–∞—П –Љ–Њ—В–Њ—А–љ–∞—П –і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М, —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞—О—Й–∞—П —Б—З–µ—В –≤¬†—Г–Љ–µ, –њ–Њ–і–і–µ—А–ґ–Є–≤–∞–µ—В –Љ–Њ–Ј–≥ –≤¬†–Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –±–Њ–і—А—Б—В–≤—Г—О—Й–µ–Љ —Б–Њ—Б—В–Њ—П–љ–Є–Є.¬†
–†–∞–±–Њ—В–∞ –≤—Л–њ–Њ–ї–љ–µ–љ–∞ –≤¬†—А–∞–Љ–Ї–∞—Е¬†–≥–Њ—Б—Г–і–∞—А—Б—В–≤–µ–љ–љ–Њ–≥–Њ –Ј–∞–і–∞–љ–Є—П –Ь–Є–љ–Є—Б—В–µ—А—Б—В–≤–∞ –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –Є¬†–љ–∞—Г–Ї–Є –†–Њ—Б—Б–Є–є—Б–Ї–Њ–є –§–µ–і–µ—А–∞—Ж–Є–Є –љ–∞¬†2021вАУ2023¬†–≥–≥.
Ye.A. Cheremushkin, PhD, N.Ye. Petrenko, PhD, V.B. Dorokhov, PhD
Institute of Higher Nervous Activity and Neurophysiology RAS
Contact person: Yevgeny A. Cheremushkin, khton@mail.ru
Investigating the neurophysiologic correlates of the transition from wakefulness to sleep that accompanies activity has diagnostic value for determining the alertness level and associated attention resources in insomnia and neurologic disorders. Cognitive performance of varying difficulty in falling asleep during daytime sleep stage 1 was investigated in 24 adult healthy subjects. A psychomotor test of two consecutive tasks was used: a counting pattern in mind from 1 to 10 with synchronous button presses with one hand and a counting pattern without presses. EEG spectral characteristics were analyzed at the test performance (control) beginning, at the first error in the pressing pattern (less than 10 presses) and before a prolonged (1.5 minutes or more) sleep episode. EEG segments were compared: 1) during mental counting (simple task); 2) during mental counting with synchronized presses (complex task). At the falling asleep early stage (in the first error situation), a generalized decrease in alpha and beta rhythms is shown when performing a simple task and localized (in caudal cortical areas) when performing a complex task. There were no changes in EEG during the simple task in the pre-sleep stage compared to the early sleep stage. The complex task was accompanied by an increase in the alpha and beta rhythms power in anterior, central and parietal cortical areas. We hypothesize that as sleepiness deepens and vigilance decreases, the periodic additional motor activity that accompanies mind counting keeps the brain in a relatively awake state.
–£–≤–∞–ґ–∞–µ–Љ—Л–є –њ–Њ—Б–µ—В–Є—В–µ–ї—М uMEDp!
–£–≤–µ–і–Њ–Љ–ї—П–µ–Љ –Т–∞—Б –Њ —В–Њ–Љ, —З—В–Њ –Ј–і–µ—Б—М —Б–Њ–і–µ—А–ґ–Є—В—Б—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П, –њ—А–µ–і–љ–∞–Ј–љ–∞—З–µ–љ–љ–∞—П –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –і–ї—П —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П.
–Х—Б–ї–Є –Т—Л –љ–µ —П–≤–ї—П–µ—В–µ—Б—М —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–Љ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П, –∞–і–Љ–Є–љ–Є—Б—В—А–∞—Ж–Є—П –љ–µ –љ–µ—Б–µ—В –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ—Б—В–Є –Ј–∞ –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ –њ–Њ—Б–ї–µ–і—Б—В–≤–Є—П, –≤–Њ–Ј–љ–Є–Ї—И–Є–µ –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–∞–Љ–Њ—Б—В–Њ—П—В–µ–ї—М–љ–Њ–≥–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Т–∞–Љ–Є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є —Б –њ–Њ—А—В–∞–ї–∞ –±–µ–Ј –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є –Ї–Њ–љ—Б—Г–ї—М—В–∞—Ж–Є–Є —Б –≤—А–∞—З–Њ–Љ.
–Э–∞–ґ–Є–Љ–∞—П –љ–∞ –Ї–љ–Њ–њ–Ї—Г ¬Ђ–Т–Њ–є—В–Є¬ї, –Т—Л –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В–µ, —З—В–Њ —П–≤–ї—П–µ—В–µ—Б—М –≤—А–∞—З–Њ–Љ –Є–ї–Є —Б—В—Г–і–µ–љ—В–Њ–Љ –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–≥–Њ –≤—Г–Ј–∞.