–Ь–µ—В–∞–±–Њ–ї–Њ–Љ–љ—Л–є –Ї–Њ–Љ–њ–µ–љ–і–Є—Г–Љ –њ–Њ¬†–Љ–∞–≥–љ–Є—П¬†–Њ—А–Њ—В–∞—В—Г
- –Р–љ–љ–Њ—В–∞—Ж–Є—П
- –°—В–∞—В—М—П
- –°—Б—Л–ї–Ї–Є
- English

–Т–≤–µ–і–µ–љ–Є–µ
–Ъ–Њ–Љ–њ–µ–љ–і–Є—Г–Љ (–Њ—В¬†–ї–∞—В. compendium)¬†вАУ —Б–ґ–∞—В–Њ–µ –Є–Ј–ї–Њ–ґ–µ–љ–Є–µ –њ–Њ–љ—П—В–Є–є, —В–µ–Њ—А–Є–є –Є¬†–Љ–µ—В–Њ–і–Є–Ї –Њ–њ—А–µ–і–µ–ї–µ–љ–љ–Њ–є –Њ–±–ї–∞—Б—В–Є –љ–∞—Г–Ї–Є. –Ъ–Њ–Љ–њ–µ–љ–і–Є—Г–Љ –њ–Њ–Ј–≤–Њ–ї—П–µ—В —Д–Њ—А–Љ–Є—А–Њ–≤–∞—В—М —Ж–µ–ї–Њ—Б—В–љ–Њ–µ, –љ–Њ¬†–љ–µ—Г–њ—А–Њ—Й–µ–љ–љ–Њ–µ –њ–Њ–љ–Є–Љ–∞–љ–Є–µ —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–µ–Љ–Њ–є –њ—А–Њ–±–ї–µ–Љ—Л. –Э–∞–њ—А–Є–Љ–µ—А, —Г–њ—А–Њ—Й–µ–љ–љ–Њ–µ –њ–Њ–љ–Є–Љ–∞–љ–Є–µ —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ –і–µ–є—Б—В–≤–Є—П –Љ–∞–≥–љ–Є–µ–≤—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –њ–Њ—А–Њ–ґ–і–∞–µ—В —Б—В–µ—А–µ–Њ—В–Є–њ–љ–Њ–µ –Њ—В–љ–Њ—И–µ–љ–Є–µ –Ї¬†–љ–Є–Љ: –Љ–∞–≥–љ–Є–є¬†вАУ –≤—Б–њ–Њ–Љ–Њ–≥–∞—В–µ–ї—М–љ—Л–є –њ—А–µ–њ–∞—А–∞—В, –Љ–∞–≥–љ–Є–є —Б–љ–Є–Љ–∞–µ—В —Б—Г–і–Њ—А–Њ–≥–Є, –Љ–∞–≥–љ–Є–є –±–µ—Б–њ–Њ–ї–µ–Ј–µ–љ. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П –њ—А–Є —Н–ї–µ–Ї—В—А–Њ–ї–Є—В–љ–Њ–Љ –і–Є—Б–±–∞–ї–∞–љ—Б–µ, –Ї–Њ–≥–і–∞ –љ–µ¬†–љ–Њ—А–Љ–∞–ї–Є–Ј–Њ–≤–∞–љ—Л –≤¬†–њ–µ—А–≤—Г—О –Њ—З–µ—А–µ–і—М —Г—А–Њ–≤–љ–Є –Љ–∞–≥–љ–Є—П, –Ї–∞–ї–Є—П, –Ї–∞–ї—М—Ж–Є—П –≤¬†–Ї—А–Њ–≤–Є, –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ –і–∞–ґ–µ —Б–∞–Љ—Л—Е —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л—Е –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ—Л—Е —Б—А–µ–і—Б—В–≤ –Љ–Њ–ґ–µ—В –±—Л—В—М –ї–Є–±–Њ –Љ–∞–ї–Њ—Н—Д—Д–µ–Ї—В–Є–≤–љ—Л–Љ, –ї–Є–±–Њ —Б–Њ–≤—Б–µ–Љ –љ–µ¬≠—Н—Д—Д–µ–Ї—В–Є–≤–љ—Л–Љ. –Ф–µ—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–љ—Л–є –Љ–Є–љ–µ—А–∞–ї—М–љ—Л–є –Њ–±–Љ–µ–љ –љ–µ¬†—В–Њ–ї—М–Ї–Њ –Є–≥—А–∞–µ—В —Б—Г—Й–µ—Б—В–≤–µ–љ–љ—Г—О —А–Њ–ї—М –≤¬†–њ–∞—В–Њ–≥–µ–љ–µ–Ј–µ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є, –љ–Њ¬†–Є –Є–Ј–Љ–µ–љ—П–µ—В —Д–∞—А–Љ–∞–Ї–Њ–Ї–Є–љ–µ—В–Є—З–µ—Б–Ї–Є–є –Є¬†—Д–∞—А–Љ–∞–Ї–Њ–і–Є–љ–∞–Љ–Є—З–µ—Б–Ї–Є–є –Њ—В–≤–µ—В –љ–∞¬†–≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ—Л—Е —Б—А–µ–і—Б—В–≤ [1].
–Т¬†—Б–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–є –±–Є–Њ–Љ–µ–і–Є—Ж–Є–љ–µ —Ж–µ–ї–Њ—Б—В–љ–Њ–µ –њ–Њ–љ–Є–Љ–∞–љ–Є–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤ –і–µ–є—Б—В–≤–Є—П –ї—О–±–Њ–≥–Њ —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ –њ—А–µ–њ–∞—А–∞—В–∞ –Њ—Б–љ–Њ–≤–∞–љ–Њ –љ–∞¬†–µ–≥–Њ –≤–ї–Є—П–љ–Є–Є –љ–∞¬†–Љ–µ—В–∞–±–Њ–ї–Њ–Љ –Њ—А–≥–∞–љ–Є–Ј–Љ–∞ [2]. –Я–Њ–і –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–Њ–Љ –њ–Њ–љ–Є–Љ–∞—О—В —Б–Њ–≤–Њ–Ї—Г–њ–љ–Њ—Б—В—М –≤—Б–µ—Е –љ–Є–Ј–Ї–Њ–Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л—Е –≤–µ—Й–µ—Б—В–≤ (–Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ–∞—П –Љ–∞—Б—Б–∞ –Љ–µ–љ–µ–µ 1000 –і–∞–ї—М—В–Њ–љ) –Є¬†–Є–Њ–љ–Њ–≤ (Mg2+, Ca2+, K+, Zn2+ –Є¬†–і—А.), –Њ–±–љ–∞—А—Г–ґ–µ–љ–љ—Л—Е –≤¬†–Ї–ї–µ—В–Ї–∞—Е –Є¬†—В–Ї–∞–љ—П—Е –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–∞ [3]. –Ш–Љ–µ–љ–љ–Њ –∞–љ–∞–ї–Є–Ј –і–µ–є—Б—В–≤–Є—П –њ—А–µ–њ–∞—А–∞—В–∞ –≤¬†–Ї–Њ–љ—В–µ–Ї—Б—В–µ –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–∞ —З–µ–ї–Њ–≤–µ–Ї–∞ –њ–Њ–Ј–≤–Њ–ї—П–µ—В —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞—В—М –Њ–±—К–µ–Ї—В–Є–≤–љ–Њ–µ –њ–Њ–љ–Є–Љ–∞–љ–Є–µ –Ї–∞–Ї —В–µ—А–∞–њ–µ–≤—В–Є—З–µ—Б–Ї–Є—Е, —В–∞–Ї –Є¬†–њ–Њ–±–Њ—З–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤.
–Э–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ—Б—В—М –Љ–∞–≥–љ–Є—П¬†вАУ –њ–∞—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–µ —Б–Њ—Б—В–Њ—П–љ–Є–µ –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–∞ —З–µ–ї–Њ–≤–µ–Ї–∞, –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ–љ–Њ–µ —Б¬†–і–Њ—Б—В–Њ–≤–µ—А–љ—Л–Љ –њ–Њ–≤—Л—И–µ–љ–Є–µ–Љ —А–Є—Б–Ї–∞ —А—П–і–∞ —Е—А–Њ–љ–Є—З–µ—Б–Ї–Є—Е –њ–∞—В–Њ–ї–Њ–≥–Є–є [4]. –І—В–Њ–±—Л –Ї–Њ–Љ–њ–µ–љ—Б–Є—А–Њ–≤–∞—В—М –і–µ—Д–Є—Ж–Є—В –Љ–∞–≥–љ–Є—П, —Б–ї–µ–і—Г–µ—В –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞—В—М –њ—А–µ–њ–∞—А–∞—В—Л –љ–∞¬†–Њ—Б–љ–Њ–≤–µ –Њ—А–≥–∞–љ–Є—З–µ—Б–Ї–Є—Е —Б–Њ–ї–µ–є –Љ–∞–≥–љ–Є—П, –Ї–Њ—В–Њ—А—Л–µ –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–љ–µ–Њ—А–≥–∞–љ–Є—З–µ—Б–Ї–Є–Љ–Є —Б–Њ–ї—П–Љ–Є —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—В—Б—П –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Њ–є –±–Є–Њ—Г—Б–≤–Њ—П–µ–Љ–Њ—Б—В—М—О [1].
–Т—Л—Б–Њ–Ї–Њ–є –±–Є–Њ—Г—Б–≤–Њ—П–µ–Љ–Њ—Б—В—М—О –Њ–±–ї–∞–і–∞—О—В –њ—А–µ–њ–∞—А–∞—В—Л –љ–∞¬†–Њ—Б–љ–Њ–≤–µ —Б–Њ–ї–µ–є –Љ–∞–≥–љ–Є—П —Б¬†–Њ—А–≥–∞–љ–Є—З–µ—Б–Ї–Є–Љ–Є –Ї–Є—Б–ї–Њ—В–∞–Љ–Є, –≤¬†–Ї–Њ—В–Њ—А—Л—Е –∞–љ–Є–Њ–љ –Ї–Є—Б–ї–Њ—В—Л —Б–ї—Г–ґ–Є—В –њ–µ—А–µ–љ–Њ—Б—З–Є–Ї–Њ–Љ –Љ–∞–≥–љ–Є—П –≤–љ—Г—В—А—М –Ї–ї–µ—В–Њ–Ї. –Т–Љ–µ—Б—В–µ —Б¬†—В–µ–Љ –∞–љ–Є–Њ–љ-–њ–µ—А–µ–љ–Њ—Б—З–Є–Ї —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г–µ—В—Б—П —Б–∞–Љ–Њ—Б—В–Њ—П—В–µ–ї—М–љ—Л–Љ–Є —Ж–µ–ї–µ–≤—Л–Љ–Є —Н—Д—Д–µ–Ї—В–∞–Љ–Є, –љ–∞–њ—А–∞–≤–ї–µ–љ–љ—Л–Љ–Є –љ–∞¬†–њ–Њ–і–і–µ—А–ґ–∞–љ–Є–µ —Б–Є—Б—В–µ–Љ –Њ—А–≥–∞–љ–Є–Ј–Љ–∞. –Ґ–∞–Ї–Є–Љ –∞–љ–Є–Њ–љ–Њ–Љ —П–≤–ї—П–µ—В—Б—П –Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ (–∞–љ–Є–Њ–љ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л) —Б¬†—Г–љ–Є–Ї–∞–ї—М–љ—Л–Љ —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ –і–µ–є—Б—В–≤–Є–µ–Љ.
–Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞, –Є–ї–Є –≤–Є—В–∞–Љ–Є–љ –Т13,¬†вАУ –њ—А–µ–і—И–µ—Б—В–≤–µ–љ–љ–Є–Ї –≤¬†–±–Є–Њ—Б–Є–љ—В–µ–Ј–µ –њ–Є—А–Є–Љ–Є–і–Є–љ–Њ–≤—Л—Е –Њ—Б–љ–Њ–≤–∞–љ–Є–є, —Б—В–Є–Љ—Г–ї–Є—А—Г—О—Й–Є–є —А–Њ—Б—В –ґ–Є–≤–Њ—В–љ—Л—Е, —А–∞—Б—В–µ–љ–Є–є –Є¬†–Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤.
–°–Њ–ї–Є –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –њ–Њ–≤—Л—И–∞—О—В –±–Є–Њ–і–Њ—Б—В—Г–њ–љ–Њ—Б—В—М –Є–Њ–љ–Њ–≤ —А–∞–Ј–ї–Є—З–љ—Л—Е –Љ–µ—В–∞–ї–ї–Њ–≤, —Б–Њ–і–µ—А–ґ–∞—Й–Є—Е—Б—П –≤¬†–Љ–Є–љ–µ—А–∞–ї—М–љ—Л—Е –њ–Є—Й–µ–≤—Л—Е –і–Њ–±–∞–≤–Ї–∞—Е –Є¬†–њ—А–µ–њ–∞—А–∞—В–∞—Е –љ–∞¬†–Њ—Б–љ–Њ–≤–µ –Љ–Є–љ–µ—А–∞–ї–Њ–≤. –Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ —Б–Њ–і–µ—А–ґ–Є—В—Б—П –≤¬†–Љ–Њ–ї–Њ–Ї–µ (–Ї–Њ—А–Њ–≤—М–µ–Љ, –Ї–Њ–Ј—М–µ–Љ), –Љ–Њ–ї–Њ—З–љ—Л—Е –њ—А–Њ–і—Г–Ї—В–∞—Е, –∞¬†—В–∞–Ї–ґ–µ –≤¬†–Љ–Њ—А–Ї–Њ–≤–Є –Є¬†—Б–≤–µ–Ї–ї–µ [5, 6]. –Т¬†–Ї–Њ—А–Њ–≤—М–µ–Љ –Љ–Њ–ї–Њ–Ї–µ –µ–µ —Г—А–Њ–≤–µ–љ—М –і–Њ—Б—В–Є–≥–∞–µ—В –≤¬†—Б—А–µ–і–љ–µ–Љ 80 –Љ–Ї–≥/–Љ–ї [7], –≤¬†—Б—Г—Е–Њ–є –Љ–Њ–ї–Њ—З–љ–Њ–є —Б—Л–≤–Њ—А–Њ—В–Ї–µ¬†вАУ –Њ—В¬†64 –і–Њ 146¬†–Љ–≥ –љ–∞¬†100¬†–≥¬†[8]. –І—В–Њ –Ї–∞—Б–∞–µ—В—Б—П –і—А—Г–≥–Є—Е –њ—А–Њ–і—Г–Ї—В–Њ–≤, —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ –≤¬†–љ–Є—Е –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –љ–µ¬†–Є–Ј—Г—З–µ–љ–Њ.
–Я–Њ—В—А–µ–±–љ–Њ—Б—В—М –≤–Ј—А–Њ—Б–ї–Њ–≥–Њ —З–µ–ї–Њ–≤–µ–Ї–∞ –≤¬†–Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В–µ –Њ–Ї–Њ–ї–Њ 500вАУ1500¬†–Љ–≥/—Б—Г—В. –£¬†–Ї–Њ—А–Љ—П—Й–Є—Е –Љ–∞—В–µ—А–µ–є —Н—В–Њ—В –њ–Њ–Ї–∞–Ј–∞—В–µ–ї—М –і–Њ—Б—В–Є–≥–∞–µ—В 3000¬†–Љ–≥/—Б—Г—В, —Г¬†–Є–љ—В–µ–љ—Б–Є–≤–љ–Њ —В—А–µ–љ–Є—А—Г—О—Й–Є—Е—Б—П —Б–њ–Њ—А—В—Б–Љ–µ–љ–Њ–≤¬†вАУ 3000вАУ5000¬†–Љ–≥/—Б—Г—В.
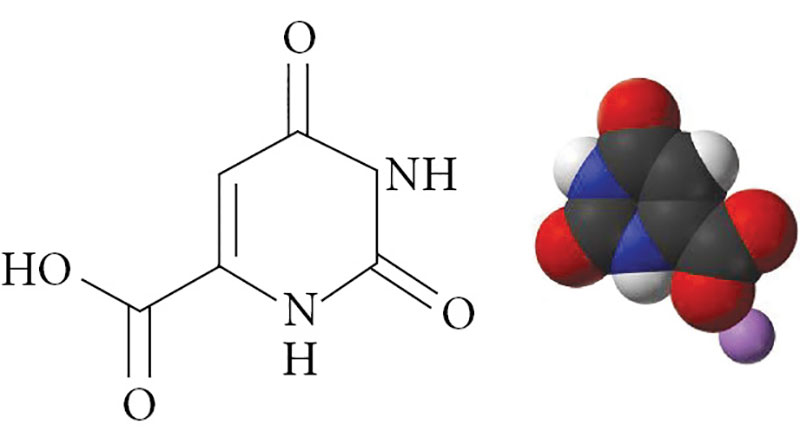
–Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В —Б–Њ–±–Њ–є¬†–≥–µ—В–µ—А–Њ—Ж–Є–Ї–ї–Є—З–µ—Б–Ї–Њ–µ —Б–Њ–µ–і–Є–љ–µ–љ–Є–µ¬†вАУ 2,6-–і–Є–Њ–Ї—Б–Є–њ–Є—А–Є–Љ–Є–і–Є–љ-4-–Ї–∞—А–±–Њ–љ–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ (—А–Є—Б.¬†1), —Б–Є–љ—В–µ–Ј–Є—А—Г–µ—В—Б—П –≤–Њ –≤—Б–µ—Е –Ї–ї–µ—В–Ї–∞—Е –Њ—А–≥–∞–љ–Є–Ј–Љ–∞ –Є¬†—П–≤–ї—П–µ—В—Б—П –њ—А–Њ–Љ–µ–ґ—Г—В–Њ—З–љ—Л–Љ –њ—А–Њ–і—Г–Ї—В–Њ–Љ —Б–Є–љ—В–µ–Ј–∞ –њ–Є—А–Є–Љ–Є–і–Є–љ–Њ–≤—Л—Е –љ—Г–Ї–ї–µ–Њ—В–Є–і–Њ–≤ (–Њ—А–Њ—В–Є–і–Є–љ—Д–Њ—Б—Д–∞—В–∞, —Г—А–Є–і–Є–љ—В—А–Є—Д–Њ—Б—Д–∞—В–∞, —Ж–Є—В–Є–і–Є–љ—В—А–Є—Д–Њ—Б—Д–∞—В–∞) [9]. –§–µ—А–Љ–µ–љ—В—Л –±–Є–Њ—В—А–∞–љ—Б—Д–Њ—А–Љ–∞—Ж–Є–є –Њ—А–Њ—В–∞—В–∞ –Њ–±–љ–∞—А—Г–ґ–µ–љ—Л –≤–Њ –≤—Б–µ—Е –Є–Ј–≤–µ—Б—В–љ—Л—Е —В–Ї–∞–љ—П—Е.
–Т–∞–ґ–љ—Л–є –Љ–Њ–Љ–µ–љ—В: –Њ—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ –Є¬†–µ–µ –њ—А–Њ–Є–Ј–≤–Њ–і–љ—Л–µ —П–≤–ї—П—О—В—Б—П –љ—Г—В—А–Є–µ–љ—В–∞–Љ–Є –і–ї—П –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л [10]. –Э–∞–њ—А–Є–Љ–µ—А,¬†–≥–µ–љ–Њ–Љ—Л –Љ–Њ–ї–Њ—З–љ–Њ–Ї–Є—Б–ї—Л—Е –±–∞–Ї—В–µ—А–Є–є Lactobacillus —Б–Њ–і–µ—А–ґ–∞—В¬†–≥–µ–љ—Л, –Ї–Њ–і–Є—А—Г—О—Й–Є–µ —Б–њ–µ—Ж–Є–∞–ї—М–љ—Л–µ –±–µ–ї–Ї–Є –і–ї—П —В—А–∞–љ—Б–њ–Њ—А—В–∞ –Є¬†–±–Є–Њ—В—А–∞–љ—Б—Д–Њ—А–Љ–∞—Ж–Є–є –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л [11].
–Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ —Г—З–∞—Б—В–≤—Г–µ—В –≤–Њ –Љ–љ–Њ–≥–Є—Е –Њ—Б–љ–Њ–≤–љ—Л—Е –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –њ—А–Њ—Ж–µ—Б—Б–∞—Е, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є:
-
–њ–µ—А–µ—А–∞–±–Њ—В–Ї–µ¬†–≥–ї—О–Ї–Њ–Ј—Л;
-
–њ–Њ–і–і–µ—А–ґ–∞–љ–Є–Є —А–µ–Ј–µ—А–≤–Њ–≤ –∞–і–µ–љ–Њ–Ј–Є–љ—В—А–Є—Д–Њ—Б—Д–∞—В–∞ (–Р–Ґ–§);
-
—Б–Є–љ—В–µ–Ј–µ —А–Є–±–Њ–Ј—Л;
-
—Б–Њ–Ј–і–∞–љ–Є–Є —А–µ–Ј–µ—А–≤–Њ–≤ –Ї–∞—А–љ–Њ–Ј–Є–љ–∞ –≤¬†—Б–Ї–µ–ї–µ—В–љ—Л—Е –Љ—Л—И—Ж–∞—Е –Є¬†–Љ–Є–Њ–Ї–∞—А–і–µ;
-
—А–Њ—Б—В–µ –Є¬†—А–∞–Ј–≤–Є—В–Є–Є –Ї–ї–µ—В–Њ–Ї –Є¬†—В–Ї–∞–љ–µ–є, –љ–∞–њ—А–Є–Љ–µ—А –Љ—Л—И–µ—З–љ–Њ–є (–Ј–∞ —Б—З–µ—В —Б–Є–љ—В–µ–Ј–∞ –†–Э–Ъ);
-
–∞–Ї—В–Є–≤–∞—Ж–Є–Є —Б–Њ–Ї—А–∞—В–Є—В–µ–ї—М–љ—Л—Е –≤–Њ–Ј¬≠–Љ–Њ–ґ–љ–Њ—Б—В–µ–є –Љ—Л—И–µ—З–љ—Л—Е —В–Ї–∞–љ–µ–є¬†[12].
–≠—В–Є –Є¬†–і—А—Г–≥–Є–µ –±–Є–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Є–µ –∞—Б–њ–µ–Ї—В—Л –і–µ–є—Б—В–≤–Є—П –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Є¬†–Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞ –≤¬†—Б–Њ—Б—В–∞–≤–µ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ –Њ–±—Г—Б–ї–Њ–≤–ї–Є–≤–∞—О—В —Н—Д—Д–µ–Ї—В—Л –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Є¬†–µ–µ –Љ–∞–≥–љ–Є–µ–≤–Њ–є —Б–Њ–ї–Є.
–Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ –Њ–Ї–∞–Ј—Л–≤–∞–µ—В —Б—В–Є–Љ—Г–ї–Є—А—Г—О—Й–µ–µ –і–µ–є—Б—В–≤–Є–µ –љ–∞¬†–±–µ–ї–Ї–Њ–≤—Л–є –Њ–±–Љ–µ–љ, –±–ї–∞–≥–Њ—В–≤–Њ—А–љ–Њ –≤–ї–Є—П–µ—В –љ–∞¬†—Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–µ —Б–Њ—Б—В–Њ—П–љ–Є–µ –њ–µ—З–µ–љ–Є, —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В —Б–љ–Є–ґ–µ–љ–Є—О —Г—А–Њ–≤–љ—П —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ –≤¬†–Ї—А–Њ–≤–Є, —Г–ї—Г—З—И–∞–µ—В —Б–Њ–Ї—А–∞—В–Є—В–µ–ї—М–љ—Г—О —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –Љ–Є–Њ–Ї–∞—А–і–∞ [5, 6, 12].
–°–ї–µ–і—Г–µ—В –Њ—В–Љ–µ—В–Є—В—М, —З—В–Њ –Ї¬†–љ–∞—Б—В–Њ—П—Й–µ–Љ—Г –Љ–Њ–Љ–µ–љ—В—Г –≤—Л–њ–Њ–ї–љ–µ–љ–Њ —Б–≤—Л—И–µ 6500 –±–Є–Њ–Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Є¬†–Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞. –Ъ–∞–ґ–і—Л–є¬†–≥–Њ–і –њ—Г–±–ї–Є–Ї—Г–µ—В—Б—П –Њ—В¬†120 –і–Њ 150 —А–∞–±–Њ—В –њ–Њ¬†—А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ –њ—А–Њ–≤–µ–і–µ–љ–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є. –Ъ–∞–Ј–∞–ї–Њ—Б—М –±—Л, —Б–≤–Њ–є—Б—В–≤–∞ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –±–Є–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Є–µ –Є¬†—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ, —Е–Њ—А–Њ—И–Њ –Є–Ј–≤–µ—Б—В–љ—Л. –Ґ–µ–Љ –љ–µ¬†–Љ–µ–љ–µ–µ —В–Њ—З–љ—Л–µ –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –Ї–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞ –Њ—Б—В–∞—О—В—Б—П –љ–µ¬†–і–Њ –Ї–Њ–љ—Ж–∞ –Є–Ј—Г—З–µ–љ–љ—Л–Љ–Є.
–†–∞—Б—Б–Љ–Њ—В—А–Є–Љ –і–∞–љ–љ—Л–µ —Б–Є—Б—В–µ–Љ–∞—В–Є—З–µ—Б–Ї–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –Є–Љ–µ—О—Й–Є—Е—Б—П –±–Є–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Є—Е, —Н–Ї–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ—Л—Е –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є—Е —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Є¬†–Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞, —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О—Й–Є–µ –Њ–± –Є—Е¬†–њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л—Е, –љ–µ–є—А–Њ- –Є¬†–Ї–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л—Е —Б–≤–Њ–є—Б—В–≤–∞—Е.
–≠–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ—Л–µ –Є –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞ –Є –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л
–Ю—А–Њ—В–Њ–≤–∞—П (–Њ—В¬†–≥—А–µ—З. oros¬†вАУ –Љ–Њ–ї–Њ—З–љ–∞—П —Б—Л–≤–Њ—А–Њ—В–Ї–∞) –Ї–Є—Б–ї–Њ—В–∞ –±—Л–ї–∞ –≤–њ–µ—А–≤—Л–µ –≤—Л–і–µ–ї–µ–љ–∞ –≤¬†1905¬†–≥.¬†–Є–Ј —Б—Л–≤–Њ—А–Њ—В–Ї–Є –Ї–Њ—А–Њ–≤—М–µ–≥–Њ –Љ–Њ–ї–Њ–Ї–∞ [13]. –Ъ¬†1930-–Љ¬†–≥–≥. –±—Л–ї–∞ –Њ–њ–Є—Б–∞–љ–∞ —Б—В—А—Г–Ї—В—Г—А–∞ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Ї–∞–Ї 4-—Г—А–∞—Ж–Є–ї-–Ї–∞—А–±–Њ–љ–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л, –њ—А–Њ–Є–Ј–≤–Њ–і–љ–Њ–≥–Њ —Г—А–∞—Ж–Є–ї–∞¬†[14].
–Т¬†1947вАУ48¬†–≥–≥. H.K. Mitchell –Є¬†—Б–Њ–∞–≤—В. —Г—Б—В–∞–љ–Њ–≤–Є–ї–Є, —З—В–Њ –Њ—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ —П–≤–ї—П–µ—В—Б—П –њ—А–Њ–Љ–µ–ґ—Г—В–Њ—З–љ—Л–Љ –њ—А–Њ–і—Г–Ї—В–Њ–Љ –±–Є–Њ—Б–Є–љ—В–µ–Ј–∞ –њ–Є—А–Є–Љ–Є–і–Є–љ–Њ–≤—Л—Е –љ—Г–Ї–ї–µ–Њ—В–Є–і–Њ–≤ –Є¬†–Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–Њ–Љ –≤—Б–µ—Е –ґ–Є–≤—Л—Е –Ї–ї–µ—В–Њ–Ї [15].
–Т¬†1948¬†–≥.¬†–±—Л–ї –Њ—В–Ї—А—Л—В —Д–∞–Ї—В–Њ—А —А–Њ—Б—В–∞ –Ї—А—Л—Б –Є¬†—Ж—Л–њ–ї—П—В, –љ–∞–Ј–≤–∞–љ–љ—Л–є –≤–Є—В–∞–Љ–Є–љ–Њ–Љ B13. –Ц–Є–≤–Њ—В–љ—Л–µ, –љ–∞—Е–Њ–і–Є–≤—И–Є–µ—Б—П –љ–∞¬†–і–Є–µ—В–µ —Б¬†–і–µ—Д–Є—Ж–Є—В–Њ–Љ –і–∞–љ–љ–Њ–≥–Њ —Д–∞–Ї—В–Њ—А–∞, –њ—А–Њ–Є–Ј–≤–Њ–і–Є–ї–Є –љ–µ–њ–Њ–ї–љ–Њ—Ж–µ–љ–љ–Њ–µ –њ–Њ—В–Њ–Љ—Б—В–≤–Њ, –њ–ї–Њ—Е–Њ —А–Њ—Б–ї–Є, —Е–∞—А–∞–Ї—В–µ—А–Є–Ј–Њ–≤–∞–ї–Є—Б—М –≤—Л—Б–Њ–Ї–Њ–є —Б–Љ–µ—А—В–љ–Њ—Б—В—М—О –≤¬†–њ–µ—А–Є–Њ–і –ї–∞–Ї—В–∞—Ж–Є–Є [16].
–Т¬†1953¬†–≥.¬†L. Manna –Є¬†S.M. Hauge –і–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ –≤–Є—В–∞–Љ–Є–љ B13 –Є¬†–Њ—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞¬†вАУ –Є–і–µ–љ—В–Є—З–љ—Л–µ –≤–µ—Й–µ—Б—В–≤–∞ [17]. –Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ –≤—Л–Ј—Л–≤–∞–µ—В –Ј–∞–Љ–µ—В–љ–Њ–µ —Г—Б–Ї–Њ—А–µ–љ–Є–µ —А–Њ—Б—В–∞ –Љ–Њ–ї–Њ–і—Л—Е –Ї—А—Л—Б, –µ–µ –і–ї–Є—В–µ–ї—М–љ—Л–є –њ—А–Є–µ–Љ –њ–Њ–≤—Л—И–∞–µ—В –њ–ї–Њ–і–Њ–≤–Є—В–Њ—Б—В—М –ґ–Є–≤–Њ—В–љ—Л—Е [18].
–Т–њ–Њ—Б–ї–µ–і—Б—В–≤–Є–Є –±—Л–ї–Є —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ—Л –Є¬†–і—А—Г–≥–Є–µ —Н—Д—Д–µ–Ї—В—Л –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л. –†–µ—З—М –Є–і–µ—В –Њ¬†–≥–µ–њ–∞—В–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–Љ —Б–≤–Њ–є—Б—В–≤–µ [19] –Є¬†–Ј–∞–Љ–µ–і–ї–µ–љ–Є–Є —А–∞–Ј–≤–Є—В–Є—П —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ–Њ–≥–Њ —Ж–Є—А—А–Њ–Ј–∞ –њ–µ—З–µ–љ–Є [20]. –Ю—А–Њ—В–Њ–≤—Г—О –Ї–Є—Б–ї–Њ—В—Г –Є¬†–µ–µ –Љ–∞–≥–љ–Є–µ–≤—Г—О —Б–Њ–ї—М –Љ–Њ–ґ–љ–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞—В—М –≤¬†–Ї–∞—З–µ—Б—В–≤–µ –і–µ–є—Б—В–≤–µ–љ–љ–Њ–≥–Њ –Ј–∞—Й–Є—В–љ–Њ–≥–Њ –≤–µ—Й–µ—Б—В–≤–∞ –њ—А–Њ—В–Є–≤ –Є–Њ–љ–Є–Ј–Є—А—Г—О—Й–µ–є —А–∞–і–Є–∞—Ж–Є–Є [21]. –Ч–∞–Љ–µ—В–Є–Љ, —З—В–Њ –і–Њ—Б—В–∞—В–Њ—З–љ–∞—П –Њ–±–µ—Б–њ–µ—З–µ–љ–љ–Њ—Б—В—М –Љ–∞–≥–љ–Є–µ–Љ —В–∞–Ї–ґ–µ –Ј–∞—Й–Є—Й–∞–µ—В –Њ—В¬†—А–∞–і–Є–∞—Ж–Є–Є –Є¬†–Є–љ—В–Њ–Ї—Б–Є–Ї–∞—Ж–Є–Є —Б–≤–Є–љ—Ж–Њ–Љ [22].
–†–µ–Ј—Г–ї—М—В–∞—В—Л –±–Њ–ї–µ–µ –њ–Њ–Ј–і–љ–Є—Е –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –њ—А–Є–≤–Њ–і–Є—В –Ї¬†–њ–µ—А—В—Г—А–±–∞—Ж–Є—П–Љ —А–∞–Ј–ї–Є—З–љ—Л—Е –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –њ—Г—В–µ–є¬†вАУ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В, —В—А–Є–≥–ї–Є—Ж–µ—А–Є–і–Њ–≤, —Д–Њ—Б—Д–Њ–ї–Є–њ–Є–і–Њ–≤, –±–µ—В–∞-–Њ–Ї–Є—Б–ї–µ–љ–Є—П, —Б–Є–љ—В–µ–Ј–∞ –љ—Г–Ї–ї–µ–Њ—В–Є–і–Њ–≤ –Є¬†–Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ —Г–≥–ї–µ–≤–Њ–і–Њ–≤ [23]. –Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ –њ–Њ–≤—Л—И–∞–µ—В —Г—А–Њ–≤–µ–љ—М –і–Є–∞—Ж–Є–ї–≥–ї–Є—Ж–µ—А–Є–і–Њ–≤ –Є¬†—Б–љ–Є–ґ–∞–µ—В –∞–Ї—В–Є–≤–љ–Њ—Б—В—М —Б—Г–њ–µ—А–Њ–Ї—Б–Є–і–і–Є—Б–Љ—Г—В–∞–Ј—Л –≤¬†–њ–µ—З–µ–љ–Є –Ї—А—Л—Б [24] –Ј–∞ —Б—З–µ—В —Г–≤–µ–ї–Є—З–µ–љ–Є—П –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є —Д–Њ—Б—Д–∞—В–Є–і–∞—В-—Д–Њ—Б—Д–Њ–≥–Є–і—А–Њ–ї–∞–Ј—Л [25]. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П –њ–µ—А–Њ—А–∞–ї—М–љ—Л–є –њ—А–Є–µ–Љ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л —В–Њ—А–Љ–Њ–Ј–Є—В —Б–µ–Ї—А–µ—Ж–Є—О –њ–µ—З–µ–љ—М—О –ї–Є–њ–Њ–њ—А–Њ—В–µ–Є–љ–Њ–≤ –љ–Є–Ј–Ї–Њ–є –Є¬†–Њ—З–µ–љ—М –љ–Є–Ј–Ї–Њ–є –њ–ї–Њ—В–љ–Њ—Б—В–Є, —З—В–Њ —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В —Б–љ–Є–ґ–µ–љ–Є—О —Г—А–Њ–≤–љ—П —В—А–Є–≥–ї–Є—Ж–µ—А–Є–і–Њ–≤ –≤¬†–Ї—А–Њ–≤–Є [26, 27].
–Ш–Ј–Љ–µ—А–µ–љ–Є–µ —Г—А–Њ–≤–љ—П –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –≤¬†–Љ–Њ—З–µ —П–≤–ї—П–µ—В—Б—П —Ж–µ–љ–љ—Л–Љ –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л–Љ —В–µ—Б—В–Њ–Љ –і–ї—П –Є–Ј—Г—З–µ–љ–Є—П –љ–∞—А—Г—И–µ–љ–Є–є –Њ–±–Љ–µ–љ–∞ –≤–µ—Й–µ—Б—В–≤, —Б–≤—П–Ј–∞–љ–љ—Л—Е —Б¬†–±–Є–Њ—Б–Є–љ—В–µ–Ј–Њ–Љ –∞—А–≥–Є–љ–Є–љ–∞¬†вАУ –∞–Љ–Є–љ–Њ–Ї–Є—Б–ї–Њ—В—Л, –Є–Љ–µ—О—Й–µ–є –Њ—Б–Њ–±–Њ–µ –Ј–љ–∞—З–µ–љ–Є–µ –і–ї—П —Д—Г–љ–Ї—Ж–Є–Њ–љ–Є—А–Њ–≤–∞–љ–Є—П –Љ–Є–Њ–Ї–∞—А–і–∞. –Р—А–≥–Є–љ–Є–љ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Є—А—Г–µ—В—Б—П –≤¬†–Њ–Ї—Б–Є–і –∞–Ј–Њ—В–∞ (II)¬†вАУ –≤–∞–ґ–љ–µ–є—И–Є–є –≤–∞–Ј–Њ–і–Є–ї–∞—В–∞—В–Њ—А –Є¬†—Д–∞–Ї—В–Њ—А –∞–љ–≥–Є–Њ–≥–µ–љ–µ–Ј–∞.
–Т¬†–љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П –≤¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Њ–є –њ—А–∞–Ї—В–Є–Ї–µ –Њ–њ—В–Є–Љ–∞–ї—М–љ—Л–Љ –њ—А–µ–њ–∞—А–∞—В–Њ–Љ –і–ї—П –≤–Њ—Б–њ–Њ–ї–љ–µ–љ–Є—П –і–µ—Д–Є—Ж–Є—В–∞ –Љ–∞–≥–љ–Є—П –њ—А–Є–Ј–љ–∞–љ –Ь–∞–≥–љ–µ—А–Њ—В (–Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В). –Я—А–µ–њ–∞—А–∞—В –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Є—Б–њ–Њ–ї—М–Ј—Г–µ—В—Б—П –Ї–∞–Ї –≤¬†—Б–≤–Њ–±–Њ–і–љ–Њ–Љ –≤–Є–і–µ, —В–∞–Ї –Є¬†–≤ –≤–Є–і–µ —Б–Њ–ї–µ–є (–Љ–∞–≥–љ–Є—П –Є–ї–Є –Ї–∞–ї–Є—П). –Я–µ—А–Њ—А–∞–ї—М–љ—Л–є –њ—А–Є–µ–Љ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Њ–±—Л—З–љ–Њ —А–µ–Ї–Њ–Љ–µ–љ–і—Г–µ—В—Б—П –њ–Њ—Б–ї–µ —Е–Є—А—Г—А–≥–Є—З–µ—Б–Ї–Є—Е –Њ–њ–µ—А–∞—Ж–Є–є –љ–∞¬†—Б–µ—А–і—Ж–µ (—В—А–Є –љ–µ–і–µ–ї–Є¬†вАУ –і–≤–∞ –Љ–µ—Б—П—Ж–∞). –Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ –Љ–Њ–ґ–µ—В –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞—В—М—Б—П –њ—А–Є –≤—Б–Ї–∞—А–Љ–ї–Є–≤–∞–љ–Є–Є –љ–µ–і–Њ–љ–Њ—И–µ–љ–љ—Л—Е –і–µ—В–µ–є [28], –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П—Е –њ–µ—З–µ–љ–Є –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Њ–Љ –њ–Њ—Б—В–≥–µ–њ–∞—В–Є—В–љ–Њ–Љ —Б–Є–љ–і—А–Њ–Љ–µ [29], –Љ–µ–≥–∞–ї–Њ–±–ї–∞—Б—В–Є—З–µ—Б–Ї–Є—Е –∞–љ–µ–Љ–Є—П—Е [12], –Њ—Б—В—А–Њ–Љ –Є¬†—Е—А–Њ–љ–Є—З–µ—Б–Ї–Њ–Љ¬†–≥–µ–њ–∞—В–Є—В–µ, —Ж–Є—А—А–Њ–Ј–µ –њ–µ—З–µ–љ–Є [30].
–С–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ —Н—Д—Д–µ–Ї—В—Л –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л
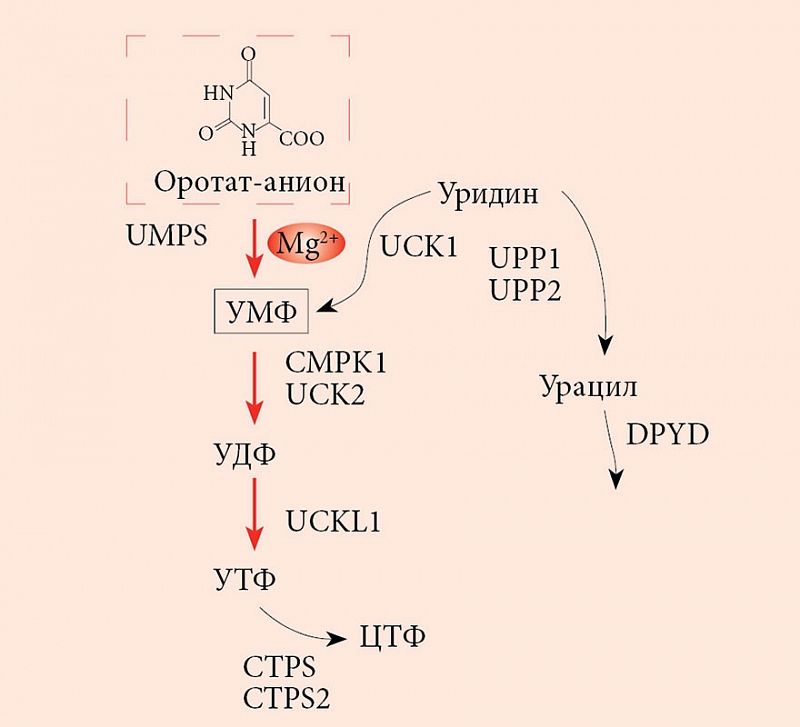
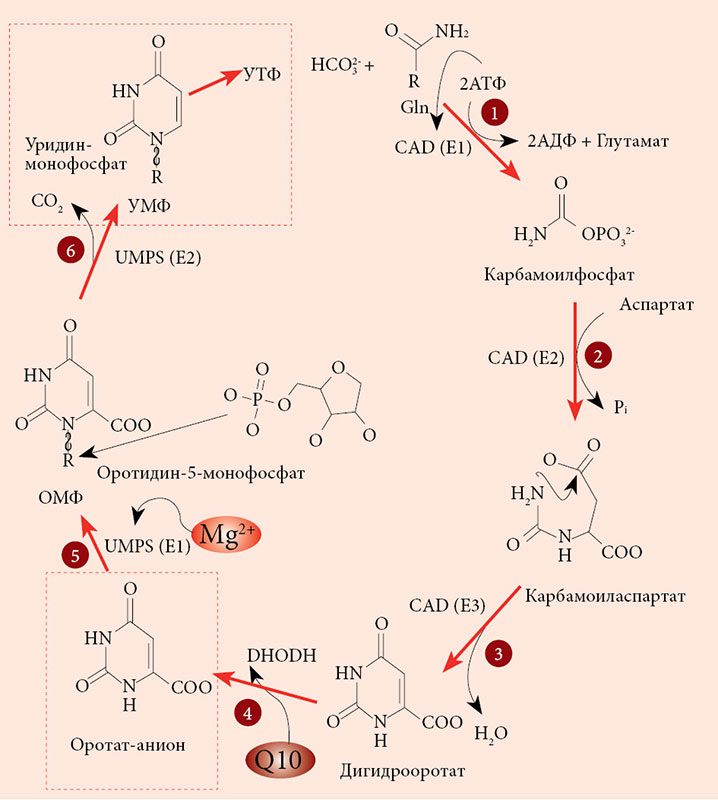
–Т¬†–Њ—А–≥–∞–љ–Є–Ј–Љ–µ —З–µ–ї–Њ–≤–µ–Ї–∞ –Њ—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ —Б–Є–љ—В–µ–Ј–Є—А—Г–µ—В—Б—П –≤¬†–њ–µ—З–µ–љ–Є –Є–Ј¬†–∞—Б–њ–∞—А—В–∞—В–∞ –Є¬†–Ї–∞—А–±–∞–Љ–Њ–Є–ї—Д–Њ—Б—Д–∞—В–∞. –Ь–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є–µ —Н—Д—Д–µ–Ї—В—Л –Њ—А–Њ—В–∞—В–∞ –Њ–≥—А–∞–љ–Є—З–µ–љ—Л —Б–Є–љ—В–µ–Ј–Њ–Љ –њ–Є—А–Є–Љ–Є–і–Є–љ–Њ–≤, —В–Њ—З–љ–µ–µ —Г—А–Є–і–Є–љ–Љ–Њ–љ–Њ—Д–Њ—Б—Д–∞—В–∞ (–£–Ь–§). –°–Є–љ—В–µ–Ј –£–Ь–§ –Њ—Б—Г—Й–µ—Б—В–≤–ї—П–µ—В—Б—П –њ–Њ—Б—А–µ–і—Б—В–≤–Њ–Љ —В—А–µ—Е —Д–µ—А–Љ–µ–љ—В–Њ–≤, –Ї–∞–ґ–і—Л–є –Є–Ј¬†–Ї–Њ—В–Њ—А—Л—Е –Ї–∞—В–∞–ї–Є–Ј–Є—А—Г–µ—В –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ—Л—Е —А–µ–∞–Ї—Ж–Є–є (—А–Є—Б.¬†2). –≠—В–Є–Љ–Є —Д–µ—А–Љ–µ–љ—В–∞–Љ–Є —П–≤–ї—П—О—В—Б—П CAD (–Ї–∞—А–±–∞–Љ–Њ–Є–ї—Д–Њ—Б—Д–∞—В —Б–Є–љ—В–∞–Ј–∞ 2¬†вАУ –∞—Б–њ–∞—А—В–∞—В –Ї–∞—А–±–∞–Љ–Њ–Є–ї—В—А–∞–љ—Б—Д–µ—А–∞–Ј–∞¬†вАУ –і–Є–≥–Є–і—А–Њ–Њ—А–Њ—В–∞–Ј–∞), DHODH (–і–Є–≥–Є–і—А–Њ–Њ—А–Њ—В–∞—В –і–µ–≥–Є–і—А–Њ–≥–µ–љ–∞–Ј–∞) –Є¬†UMPS (—Г—А–Є–і–Є–љ–Љ–Њ–љ–Њ—Д–Њ—Б—Д–∞—В —Б–Є–љ—В–∞–Ј–∞). –Я—А–µ–і–њ–Њ–ї–∞–≥–∞–µ—В—Б—П, —З—В–Њ –≤—Б–µ —В—А–Є —Д–µ—А–Љ–µ–љ—В–∞¬†вАУ CAD, DHOHD –Є¬†UMPS –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ—Л –і—А—Г–≥ —Б¬†–і—А—Г–≥–Њ–Љ –љ–∞¬†–≤–љ–µ—И–љ–µ–є —Б—В–Њ—А–Њ–љ–µ –Љ–Є—В–Њ—Е–Њ–љ–і—А–Є–∞–ї—М–љ–Њ–є –Љ–µ–Љ–±—А–∞–љ—Л, —З—В–Њ –Њ–±–µ—Б–њ–µ—З–Є–≤–∞–µ—В —Н—Д—Д–µ–Ї—В–Є–≤–љ—Г—О –њ–µ—А–µ–і–∞—З—Г —Б—Г–±—Б—В—А–∞—В–Њ–≤/–њ—А–Њ–і—Г–Ї—В–Њ–≤ –Њ—В¬†—Д–µ—А–Љ–µ–љ—В–∞ –Ї¬†—Д–µ—А–Љ–µ–љ—В—Г [23].
–£–≤–µ–ї–Є—З–µ–љ–Є–µ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –Њ—А–Њ—В–∞—В–∞ —З–µ—А–µ–Ј —Б–і–≤–Є–≥ —А–∞–≤–љ–Њ–≤–µ—Б–Є—П —А–µ–∞–Ї—Ж–Є–є, –Ї–∞—В–∞–ї–Є–Ј–Є—А—Г–µ–Љ—Л—Е —Д–µ—А–Љ–µ–љ—В–Њ–Љ –£–Ь–§-—Б–Є–љ—В–∞–Ј–Њ–є, –њ—А–Є–≤–Њ–і–Є—В –Ї¬†—Г–≤–µ–ї–Є—З–µ–љ–Є—О –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–є –£–Ь–§. –Т¬†–Њ—В–ї–Є—З–Є–µ –Њ—В¬†–Њ—А–Њ—В–∞—В–∞ –£–Ь–§ –≤–Њ–≤–ї–µ—З–µ–љ –≤¬†–≥–Њ—А–∞–Ј–і–Њ –±–Њ–ї—М—И–µ–µ —З–Є—Б–ї–Њ –±–Є–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Є—Е –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є–є —Б¬†–Ї–ї–µ—В–Ї–∞–Љ–Є, –Є¬†–і–ї—П —А–∞—Б—Б–Љ–Њ—В—А–µ–љ–Є—П –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –£–Ь–§ –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ –≤–Њ—Б–њ–Њ–ї—М–Ј–Њ–≤–∞—В—М—Б—П –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є–Љ –Ї–Њ–љ—В–µ–Ї—Б—В–Њ–Љ (—А–Є—Б.¬†3). –Ь–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є–є –Ї–Њ–љ—В–µ–Ї—Б—В –Њ–њ—А–µ–і–µ–ї–µ–љ–љ–Њ–≥–Њ –Љ–µ—В–∞–±–Њ–ї–Є—В–∞ –њ—А–µ–і—Г—Б–Љ–∞—В—А–Є–≤–∞–µ—В —Б–њ–µ—Ж–Є—Д–Є—З–µ—Б–Ї–Њ–µ –Њ–њ–Є—Б–∞–љ–Є–µ –±–Є–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Є—Е –њ—А–Њ—Ж–µ—Б—Б–Њ–≤, –≤¬†–Ї–Њ—В–Њ—А—Л–µ —Н—В–Њ—В –Љ–µ—В–∞–±–Њ–ї–Є—В –≤–Њ–≤–ї–µ—З–µ–љ.
–Р–љ–∞–ї–Є–Ј –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Њ–≥–Њ –Ї–Њ–љ—В–µ–Ї—Б—В–∞ –£–Ь–§ –њ–Њ–Ї–∞–Ј—Л–≤–∞–µ—В, —З—В–Њ –∞–Ї—В–Є–≤–љ–Њ—Б—В—М —Г—А–Є–і–Є–љ-—Ж–Є—В–Є–і–Є–љ –Ї–Є–љ–∞–Ј (UCK1, UCK2), —Ж–Є—В–Є–і–Є–ї–∞—В –Ї–Є–љ–∞–Ј—Л (CMPK1), U-C –Ї–Є–љ–∞–Ј—Л-1-–њ–Њ–і–Њ–±–љ–Њ–≥–Њ –±–µ–ї–Ї–∞ (UCKL1) –Є¬†UMPS —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є—О –£–Ь–§ –Є¬†–і–≤—Г—Е –і—А—Г–≥–Є—Е –њ—А–Њ–Є–Ј–≤–Њ–і–љ—Л—Е —Д–Њ—Б—Д–∞—В–Њ–≤ (—Г—А–Є–і–Є–љ–і–Є—Д–Њ—Б—Д–∞—В (–£–Ф–§), —Г—А–Є–і–Є–љ—В—А–Є—Д–Њ—Б—Д–∞—В (–£T–§)). –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М CTP-—Б–Є–љ—В–∞–Ј (CTPS, CTPS2), –і–Є–≥–Є–і—А–Њ–њ–Є—А–Є–Љ–Є–і–Є–љ –і–µ–≥–Є–і—А–Њ–≥–µ–љ–∞–Ј—Л (DPYD) –Є¬†—Г—А–Є–і–Є–љ —Д–Њ—Б—Д–Њ—А–Є–ї–∞–Ј (UPP1, UPP2) —Г–Љ–µ–љ—М—И–∞–µ—В –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є—О –£–Ь–§, –£–Ф–§ –Є¬†–£T–§. –Я–Њ—Б–Ї–Њ–ї—М–Ї—Г —Б–Ї–Њ—А–Њ—Б—В–Є –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –£–Ь–§ –Є–Ј¬†–Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞ –Є¬†—Г—А–Є–і–Є–љ–∞ —Б—А–∞–≤–љ–Є–Љ—Л, –њ–Њ–≤—Л—И–µ–љ–Є–µ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –Њ—А–Њ—В–∞—В–∞ –Ј–∞ —Б—З–µ—В –њ—А–Є–µ–Љ–∞ —Б–Њ–ї–µ–є –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –≤–ї–Є—П–µ—В –љ–∞¬†—Г–≤–µ–ї–Є—З–µ–љ–Є–µ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–є –£–Ь–§, –£–Ф–§ –Є¬†–£T–§. –Т–µ—А–Њ—П—В–љ–Њ, —Н—В–Є —В—А–Є —Г—А–Є–і–Є–љ—Д–Њ—Б—Д–∞—В–∞ –Є¬†—П–≤–ї—П—О—В—Б—П —Б–Є–≥–љ–∞–ї—М–љ—Л–Љ–Є –Љ–Њ–ї–µ–Ї—Г–ї–∞–Љ–Є, –Њ–њ–Њ—Б—А–µ–і—Г—О—Й–Є–Љ–Є –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ–µ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ –Њ—А–Њ—В–∞—В–∞ –љ–∞¬†—Б–µ—А–і–µ—З–љ–Њ-—Б–Њ—Б—Г–і–Є—Б—В—Г—О —Б–Є—Б—В–µ–Љ—Г.
–†–µ–Ј—Г–ї—М—В–∞—В—Л —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –љ–∞¬†–ґ–Є–≤–Њ—В–љ—Л—Е –њ–Њ–і—В–≤–µ—А–ґ–і–∞—О—В, —З—В–Њ –њ–µ—А–Њ—А–∞–ї—М–љ—Л–є –њ—А–Є–µ–Љ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є—О —Г—А–Є–і–Є–љ—Д–Њ—Б—Д–∞—В–Њ–≤ –њ–ї–∞–Ј–Љ—Л –Є¬†–њ—А–Є–≤–Њ–і–Є—В –Ї¬†—Г–Љ–µ–љ—М—И–µ–љ–Є—О¬†–≥–ї–Њ–±–∞–ї—М–љ–Њ–є –Є—И–µ–Љ–Є–Є –њ—А–Є –Є–љ—Д–∞—А–Ї—В–µ –Љ–Є–Њ–Ї–∞—А–і–∞ [31]. –£–Ь–§, —А–∞–≤–љ–Њ –Ї–∞–Ї –£–Ф–§ –Є¬†–£–Ґ–§, –Њ–±–ї–∞–і–∞–µ—В –Ј–љ–∞—З–Є—В–µ–ї—М–љ—Л–Љ –Ї–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–Љ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ–Љ, –Њ—Б–Њ–±–µ–љ–љ–Њ –≤¬†–Њ—Б—В—А—Л–є –њ–µ—А–Є–Њ–і –Є–љ—Д–∞—А–Ї—В–∞ [32, 33]. –Ф–µ–є—Б—В–≤–Є–µ –£T–§ —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г–µ—В—Б—П —Г–Љ–µ–љ—М—И–µ–љ–Є–µ–Љ –њ–µ—А–µ–≥—А—Г–Ј–Ї–Є –Ї–∞–ї—М—Ж–Є–µ–Љ –Љ–Є—В–Њ—Е–Њ–љ–і—А–Є–є –Ї–∞—А–і–Є–Њ–Љ–Є–Њ—Ж–Є—В–Њ–≤ [34], –∞¬†—В–∞–Ї–ґ–µ –≤–∞–Ј–Њ–і–Є–ї–∞—В–∞—Ж–Є–Њ–љ–љ—Л–Љ –Є¬†–ї–µ–≥–Ї–Є–Љ —Б–µ–і–∞—В–Є–≤–љ—Л–Љ —Н—Д—Д–µ–Ї—В–Њ–Љ¬†[35].
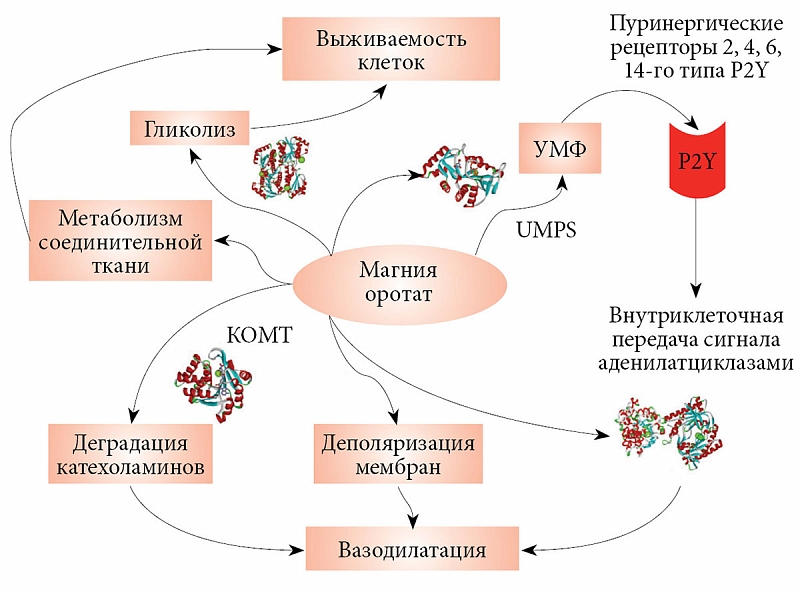
–§–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–µ –і–µ–є—Б—В–≤–Є–µ —Г—А–Є–і–Є–љ—Д–Њ—Б—Д–∞—В–Њ–≤ –Њ—Б—Г—Й–µ—Б—В–≤–ї—П–µ—В—Б—П —З–µ—А–µ–Ј –њ—Г—А–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є–µ —А–µ—Ж–µ–њ—В–Њ—А—Л¬†вАУ G-–±–µ–ї–Њ–Ї-–Ј–∞–≤–Є—Б–Є–Љ—Л–µ —А–µ—Ж–µ–њ—В–Њ—А—Л, –≤–Њ–≤–ї–µ—З–µ–љ–љ—Л–µ –≤¬†–≤–∞–Ј–Њ—А–µ–∞–Ї—В–Є–≤–љ—Л–µ —А–µ–∞–Ї—Ж–Є–Є, –∞–њ–Њ–њ—В–Њ–Ј –Є¬†—Б–µ–Ї—А–µ—Ж–Є—О —Ж–Є—В–Њ–Ї–Є–љ–Њ–≤. –Я–Њ–і–Њ–±–љ–Њ –∞–і—А–µ–љ–µ—А–≥–Є—З–µ—Б–Ї–Є–Љ –Є¬†–∞–і–µ–љ–Њ–Ј–Є–љ-—А–µ—Ж–µ–њ—В–Њ—А–∞–Љ –њ—Г—А–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є–µ —А–µ—Ж–µ–њ—В–Њ—А—Л –∞–Ї—В–Є–≤–Є—А—Г—О—В G-–±–µ–ї–Ї–Є, –∞¬†—З–µ—А–µ–Ј –љ–Є—Е –∞–і–µ–љ–Є–ї–∞—В—Ж–Є–Ї–ї–∞–Ј—Г –і–ї—П –≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ–Њ–є –њ–µ—А–µ–і–∞—З–Є —Б–Є–≥–љ–∞–ї–∞ –≤¬†–Ї–∞—А–і–Є–Њ¬≠–Љ–Є–Њ—Ж–Є—В–∞—Е (—А–Є—Б.¬†4).
–°—Г—Й–µ—Б—В–≤—Г–µ—В —З–µ—В—Л—А–µ –Ї–ї–∞—Б—Б–∞ —Н—В–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤. –Э–∞–њ—А–Є–Љ–µ—А, –Ї–ї–∞—Б—Б P2Y —Б–Њ–і–µ—А–ґ–Є—В 12 –њ—Г—А–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤. –Ю–±—А–∞—В–Є—В–µ –≤–љ–Є–Љ–∞–љ–Є–µ: –Є–Љ–µ–љ–љ–Њ —А–µ—Ж–µ–њ—В–Њ—А—Л –Ї–ї–∞—Б—Б–∞ P2Y —Б–≤—П–Ј—Л–≤–∞—О—В –£–Ф–§ –Є¬†–£T–§ [36].
–Ш–Ј¬†12 –њ—Г—А–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –Ї–ї–∞—Б—Б–∞ P2Y –Њ—Б–Њ–±—Л–є –Є–љ—В–µ—А–µ—Б –њ—А–µ–і—Б—В–∞–≤–ї—П—О—В —А–µ—Ж–µ–њ—В–Њ—А—Л P2RY2, P2RY4, P2RY6 –Є¬†P2RY14. –≠—В–Є —З–µ—В—Л—А–µ —А–µ—Ж–µ–њ—В–Њ—А–∞ –і–µ–є—Б—В–≤—Г—О—В —З–µ—А–µ–Ј –Њ–і–Є–љ –Є¬†—В–Њ—В –ґ–µ G-–±–µ–ї–Њ–Ї Gq/11. –Ъ–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л –£–Ґ–§ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г—О—В –≤–Њ–≤–ї–µ—З–µ–љ–Є—О –Є–Љ–µ–љ–љ–Њ —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ P2Y [37]. P2Y6-—А–µ—Ж–µ–њ—В–Њ—А—Л –≤–Њ–Ј–і–µ–є—Б—В–≤—Г—О—В –љ–∞¬†–≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ—Л–µ –њ—Г—В–Є, –њ—А–µ–і–Њ—В–≤—А–∞—Й–∞—П –∞–њ–Њ–њ—В–Њ–Ј, –≤—Л–Ј–≤–∞–љ–љ—Л–є —Д–∞–Ї—В–Њ—А–Њ–Љ –љ–µ–Ї—А–Њ–Ј–∞ –Њ–њ—Г—Е–Њ–ї–Є –∞–ї—М—Д–∞ [38] –Є¬†–Њ—Б—Г—Й–µ—Б—В–≤–ї—П—П –≤–∞–Ј–Њ–і–Є–ї–∞—В–∞—Ж–Є–Њ–љ–љ—Л–є —Н—Д—Д–µ–Ї—В –£–Ґ–§ [39]. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ, —В—А–∞–љ—Б—Д–Њ—А–Љ–Є—А—Г—П—Б—М –≤¬†–£–Ь–§, –Љ–Њ–ґ–µ—В –Њ—Б—Г—Й–µ—Б—В–≤–ї—П—В—М –Є¬†–Ї–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—Ж–Є—О, –Є¬†–і–Є–ї–∞—В–∞—Ж–Є—О –Ї–Њ—А–Њ–љ–∞—А–љ—Л—Е —Б–Њ—Б—Г–і–Њ–≤ –Ј–∞ —Б—З–µ—В –∞–Ї—В–Є–≤–∞—Ж–Є–Є –њ—Г—А–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ P2Y6.
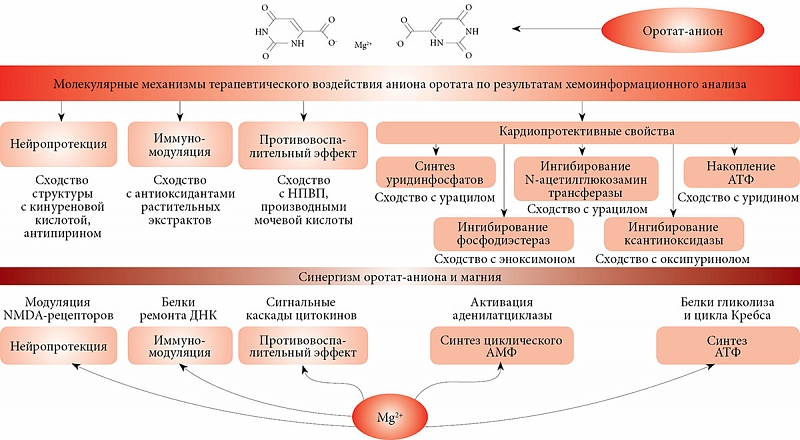
–Ь–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –Ї–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–≥–Њ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –Љ–∞–≥–љ–Є—П –≤ —Б–Њ—Б—В–∞–≤–µ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞
–Ю–±–µ—Б–њ–µ—З–µ–љ–љ–Њ—Б—В—М –Њ—А–≥–∞–љ–Є–Ј–Љ–∞ –Љ–∞–≥–љ–Є–µ–Љ –њ—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ–Њ –≤–∞–ґ–љ–∞ –і–ї—П –Ј–і–Њ—А–Њ–≤—М—П. –Ш–Њ–љ—Л –Љ–∞–≥–љ–Є—П –ґ–Є–Ј–љ–µ–љ–љ–Њ –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л –і–ї—П —Д—Г–љ–Ї—Ж–Є–Њ–љ–Є—А–Њ–≤–∞–љ–Є—П –љ–µ—А–≤–љ–Њ–є, —Б–µ—А–і–µ—З–љ–Њ-—Б–Њ—Б—Г–і–Є—Б—В–Њ–є, –Љ–Њ—З–µ–≤—Л–і–µ–ї–Є—В–µ–ї—М–љ–Њ–є –Є¬†–і—А—Г–≥–Є—Е —Б–Є—Б—В–µ–Љ –Њ—А–≥–∞–љ–Є–Ј–Љ–∞. –Ь–∞–≥–љ–Є–є –Њ–±–ї–∞–і–∞–µ—В –Ј–љ–∞—З–Є—В–µ–ї—М–љ—Л–Љ —Б–Њ–±—Б—В–≤–µ–љ–љ—Л–Љ –Ї–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–Љ —Н—Д—Д–µ–Ї—В–Њ–Љ [40], –∞¬†—В–∞–Ї–ґ–µ —Г—Б–Є–ї–Є–≤–∞–µ—В –њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–є —Н—Д—Д–µ–Ї—В –љ—Г–Ї–ї–µ–Њ–Ј–Є–і–Њ–≤, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є –∞–і–µ–љ–Њ–Ј–Є–љ–∞ [41]. –Я—А–Є–Љ–µ–љ–µ–љ–Є–µ –Љ–∞–≥–љ–Є—П –љ–∞¬†–Љ–Њ–і–µ–ї—П—Е –Є–љ—Д–∞—А–Ї—В–∞ —Г¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ—Л—Е –ґ–Є–≤–Њ—В–љ—Л—Е –њ–Њ–Ј–≤–Њ–ї—П–µ—В —Г–Љ–µ–љ—М—И–Є—В—М —А–∞–Ј–Љ–µ—А –Є–љ—Д–∞—А–Ї—В–∞ –Є¬†—Г–≤–µ–ї–Є—З–Є—В—М –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –Ї–ї–µ—В–Њ–Ї –≤¬†–Є—И–µ–Љ–Є—З–µ—Б–Ї–Њ–є –Њ–±–ї–∞—Б—В–Є [42]. –Ъ–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–є —Н—Д—Д–µ–Ї—В –Њ–±—Г—Б–ї–Њ–≤–ї–µ–љ —Б–љ–Є–ґ–µ–љ–Є–µ–Љ —Г—А–Њ–≤–љ—П –Ї–∞–ї—М—Ж–Є—П –≤¬†–Њ–±–ї–∞—Б—В–Є –Є–љ—Д–∞—А–Ї—В–∞ [43], –Є–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є–µ–Љ —А–Њ—Б—В–∞ —Г—А–Њ–≤–љ—П P-—Б–µ–ї–µ–Ї—В–Є–љ–∞ [44], –∞¬†—В–∞–Ї–ґ–µ —Г–Љ–µ–љ—М—И–µ–љ–Є–µ–Љ –≤–∞–Ј–Њ–Ї–Њ–љ—Б—В—А–Є–Ї—Ж–Є–Є –Є¬†—Г–ї—Г—З—И–µ–љ–Є–µ–Љ –Ї–Њ—А–Њ–љ–∞—А–љ–Њ–≥–Њ –Ї—А–Њ–≤–Њ—В–Њ–Ї–∞ [45]. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –Љ–∞–≥–љ–Є–є –љ–µ–Њ–±—Е–Њ–і–Є–Љ –і–ї—П –љ–Њ—А–Љ–∞–ї—М–љ–Њ–≥–Њ —Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–Њ–≥–Њ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞, –≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ–Њ–є –њ–µ—А–µ–і–∞—З–Є —Б–Є–≥–љ–∞–ї–∞ –Є¬†–њ–Њ–і–і–µ—А–ґ–∞–љ–Є—П —Б—В—А—Г–Ї—В—Г—А—Л —Б–Њ–µ–і–Є–љ–Є—В–µ–ї—М–љ–Њ–є —В–Ї–∞–љ–Є. –Ф–µ—Д–Є—Ж–Є—В –Љ–∞–≥–љ–Є—П –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ–Њ —Б–Ї–∞–Ј—Л–≤–∞–µ—В—Б—П –љ–∞¬†–≤—Б–µ—Е —Н—В–Є—Е –њ—А–Њ—Ж–µ—Б—Б–∞—Е.
–Т–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є–µ –Љ–∞–≥–љ–Є—П –Є¬†–Р–Ґ–§ –Є–≥—А–∞–µ—В –≤–∞–ґ–љ—Г—О —А–Њ–ї—М –≤¬†—Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–Њ–Љ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–µ. –Т¬†–і–Њ–њ–Њ–ї–љ–µ–љ–Є–µ –Ї¬†–≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є—О –Љ–∞–≥–љ–Є—П –Є¬†–Р–Ґ–§ Mg-–Ј–∞–≤–Є—Б–Є–Љ—Л–µ –±–µ–ї–Ї–Є —Г—З–∞—Б—В–≤—Г—О—В –≤¬†—Б–Є–љ—В–µ–Ј–µ –≤–∞–ґ–љ—Л—Е –Ї–Њ—Д–µ—А–Љ–µ–љ—В–Њ–≤, –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–µ —Г–≥–ї–µ–≤–Њ–і–Њ–≤ (–≤¬†–≥–ї–Є–Ї–Њ–ї–Є–Ј–µ). –Т¬†–Љ–Є—В–Њ—Е–Њ–љ–і—А–Є—П—Е Mg-–Ј–∞–≤–Є—Б–Є–Љ—Л–µ –±–µ–ї–Ї–Є —Г—З–∞—Б—В–≤—Г—О—В –≤¬†–Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–µ –њ–Є—А—Г–≤–∞—В–∞ –Є¬†–ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В [46]. –Т¬†—З–∞—Б—В–љ–Њ—Б—В–Є,¬†–≥–ї–Є–Ї–Њ–ї–Є—В–Є—З–µ—Б–Ї–Є–µ —Д–µ—А–Љ–µ–љ—В—Л –µ–љ–Њ–ї–∞–Ј–∞ (ENO1, ENO2), —Д–Њ—Б—Д–Њ–≥–ї—О–Ї–Њ–Љ—Г—В–∞–Ј–∞ (PGM1, PGM2, PGM3) –Є¬†6-—Д–Њ—Б—Д–Њ—Д—А—Г–Ї—В–Њ–Ї–Є–љ–∞–Ј–∞ (PFKP) —В—А–µ–±—Г—О—В –Љ–∞–≥–љ–Є–є –≤¬†–Ї–∞—З–µ—Б—В–≤–µ –Ї–Њ—Д–∞–Ї—В–Њ—А–∞ (—А–Є—Б.¬†5). –Я–Њ–Љ–Є–Љ–Њ –Є–Ј–≤–µ—Б—В–љ—Л—Е —Д—Г–љ–Ї—Ж–Є–є –≤¬†–Ј–∞–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ–Љ —Н—В–∞–њ–µ¬†–≥–ї–Є–Ї–Њ–ї–Є–Ј–∞ –µ–љ–Њ–ї–∞–Ј–∞ —Г—З–∞—Б—В–≤—Г–µ—В –≤¬†—А—П–і–µ –і—А—Г–≥–Є—Е –њ—А–Њ—Ж–µ—Б—Б–Њ–≤¬†вАУ –Ї–Њ–љ—В—А–Њ–ї–µ —А–Њ—Б—В–∞ –Ї–ї–µ—В–Њ–Ї,¬†–≥–Є–њ–Њ–Ї—Б–Є–Є –Є¬†–∞–ї–ї–µ—А–≥–Є—З–µ—Б–Ї–Њ–Љ –Є–Љ–Љ—Г–љ–љ–Њ–Љ –Њ—В–≤–µ—В–µ. –§–Њ—Б—Д–Њ–≥–ї—О–Ї–Њ–Љ—Г—В–∞–Ј–∞-1 —П–≤–ї—П–µ—В—Б—П –±–Є–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–Є–Љ –±–µ–ї–Ї–Њ–Љ, –Ї–Њ—В–Њ—А—Л–є —Г—З–∞—Б—В–≤—Г–µ—В –Ї–∞–Ї –≤¬†–≥–ї–Є–Ї–Њ–ї–Є–Ј–µ, —В–∞–Ї –Є¬†–≤¬†–≥–ї—О–Ї–Њ–љ–µ–Њ–≥–µ–љ–µ–Ј–µ. –§–Њ—Б—Д–Њ—Д—А—Г–Ї—В–Њ–Ї–Є–љ–∞–Ј–∞ –Ї–Њ–љ–≤–µ—А—В–Є—А—Г–µ—В D-—Д—А—Г–Ї—В–Њ–Ј—Г-6-—Д–Њ—Б—Д–∞—В –≤¬†1,6-—Д—А—Г–Ї—В–Њ–Ј—Г-–і–Є—Д–Њ—Б—Д–∞—В –Є¬†–Є–Љ–µ–µ—В –Ј–љ–∞—З–µ–љ–Є–µ –і–ї—П¬†–≥–ї–Є–Ї–Њ–ї–Є—В–Є—З–µ—Б–Ї–Њ–є –і–µ–≥—А–∞–і–∞—Ж–Є–Є —Г–≥–ї–µ–≤–Њ–і–Њ–≤.
–Т–∞–Ј–Њ–Ї–Њ–љ—Б—В—А–Є–Ї—Ж–Є—П/–≤–∞–Ј–Њ–і–Є–ї–∞—В–∞—Ж–Є—П –Њ—Б—Г—Й–µ—Б—В–≤–ї—П–µ—В—Б—П –њ–Њ—Б—А–µ–і—Б—В–≤–Њ–Љ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –љ–µ–є—А–Њ—В—А–∞–љ—Б–Љ–Є—В—В–µ—А–Њ–≤ –љ–∞¬†—А–µ—Ж–µ–њ—В–Њ—А—Л –Љ—Л—И–µ—З–љ—Л—Е –Ї–ї–µ—В–Њ–Ї, –њ–µ—А–µ–і–∞—З–Є —Б–Є–≥–љ–∞–ї–∞ –≤–љ—Г—В—А–Є –Ї–ї–µ—В–Њ–Ї –Є¬†–Є–Ј–Љ–µ–љ–µ–љ–Є—П –Љ–µ–Љ–±—А–∞–љ–љ–Њ–≥–Њ –њ–Њ—В–µ–љ—Ж–Є–∞–ї–∞ —Б¬†–њ–Њ–Љ–Њ—Й—М—О –Є–Њ–љ–љ—Л—Е –Ї–∞–љ–∞–ї–Њ–≤. –†—П–і Mg-–Ј–∞–≤–Є—Б–Є–Љ—Л—Е –њ–ї–∞—Ж–µ–љ—В–∞—А–љ—Л—Е –±–µ–ї–Ї–Њ–≤ —Г—З–∞—Б—В–≤—Г–µ—В –≤¬†—Н—В–Є—Е –њ—А–Њ—Ж–µ—Б—Б–∞—Е. –Ъ–∞—В–µ—Е–Њ–ї-–Ю-–Љ–µ—В–Є–ї—В—А–∞–љ—Б—Д–µ—А–∞–Ј–∞ (–ЪOMT) (—А–Є—Б.¬†6) –Є–љ–∞–Ї—В–Є–≤–Є—А—Г–µ—В –Ї–∞—В–µ—Е–Њ–ї–∞–Љ–Є–љ—Л. –Ф–µ—Д–Є—Ж–Є—В –Љ–∞–≥–љ–Є—П –њ—А–Є–≤–Њ–і–Є—В –Ї¬†—Г–Љ–µ–љ—М—И–µ–љ–Є—О –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –ЪOMT –Є, —Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ, –Ї¬†–њ–Њ–≤—Л—И–µ–љ–љ–Њ–є –Ї–Њ–љ—Б—В—А–Є–Ї—Ж–Є–Є¬†–≥–ї–∞–і–Ї–Њ–є –Љ—Г—Б–Ї—Г–ї–∞—В—Г—А—Л —Б–Њ—Б—Г–і–Њ–≤, –і–∞–ґ–µ –њ—А–Є –љ–Є–Ј–Ї–Є—Е —Г—А–Њ–≤–љ—П—Е —Б—В—А–µ—Б—Б–∞.
–Т–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ—Л–є —Б–Є–≥–љ–∞–ї –Њ—В¬†—А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –Ї–∞—В–µ—Е–Њ–ї–∞–Љ–Є–љ–Њ–≤, –∞¬†—В–∞–Ї–ґ–µ –Њ—В¬†–њ—Г—А–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤, –∞–Ї—В–Є–≤–Є—А—Г–µ–Љ—Л—Е –£–Ф–§ –Є¬†–£–Ґ–§, –њ–µ—А–µ–і–∞–µ—В—Б—П –≤–љ—Г—В—А—М –Ї–ї–µ—В–Ї–Є —З–µ—А–µ–Ј –≤—В–Њ—А–Є—З–љ—Г—О —Б–Є–≥–љ–∞–ї—М–љ—Г—О –Љ–Њ–ї–µ–Ї—Г–ї—Г¬†вАУ —Ж–Є–Ї–ї–Є—З–µ—Б–Ї–Є–є –∞–і–µ–љ–Њ–Ј–Є–љ 3', 5'-–Љ–Њ–љ–Њ—Д–Њ—Б—Д–∞—В (—Ж–Р–Ь–§). –Р–Љ–њ–ї–Є—В—Г–і–∞ —Б–Є–≥–љ–∞–ї–∞ –Њ–≥—А–∞–љ–Є—З–µ–љ–∞ –∞–Ї—В–Є–≤–љ–Њ—Б—В—М—О Mg-–Ј–∞–≤–Є—Б–Є–Љ—Л—Е –∞–і–µ–љ–Є–ї–∞—В—Ж–Є–Ї–ї–∞–Ј (ADCY1, ADCY2 –Є¬†—В.–і., –Њ–Ї–Њ–ї–Њ 10¬†–≥–µ–љ–Њ–≤). –Э–µ–і–Њ—Б—В–∞—В–Њ—З–љ–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –∞–і–µ–љ–Є–ї–∞—В—Ж–Є–Ї–ї–∞–Ј (—А–Є—Б.¬†7), –≤—Л–Ј–≤–∞–љ–љ–∞—П –≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ—Л–Љ –і–µ—Д–Є—Ж–Є—В–Њ–Љ –Љ–∞–≥–љ–Є—П, –њ—А–Є–≤–Њ–і–Є—В –Ї¬†–Ј–∞–Љ–µ–і–ї–µ–љ–Є—О –њ–µ—А–µ–і–∞—З–Є —Б–Є–≥–љ–∞–ї–∞, –∞¬†–Ј–љ–∞—З–Є—В, –Ї¬†—Б–љ–Є–ґ–µ–љ–Є—О —З—Г–≤—Б—В–≤–Є—В–µ–ї—М–љ–Њ—Б—В–Є –Ї–ї–µ—В–Њ–Ї –Ї¬†–≤–љ–µ—И–љ–Є–Љ —Б—В–Є–Љ—Г–ї–∞–Љ (–≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є, –Ї¬†—Н—Д—Д–µ–Ї—В–∞–Љ —Г—А–Є–і–Є–љ—Д–Њ—Б—Д–∞—В–Њ–≤, —Б–≤—П–Ј–∞–љ–љ—Л—Е —Б¬†–њ—Г—А–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є–Љ–Є —А–µ—Ж–µ–њ—В–Њ—А–∞–Љ–Є).
–°–Њ–µ–і–Є–љ–Є—В–µ–ї—М–љ–∞—П —В–Ї–∞–љ—М —Б–Ї—А–µ–њ–ї—П–µ—В —А–∞–Ј–ї–Є—З–љ—Л–µ –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л –њ–ї–∞—Ж–µ–љ—В—Л –Є¬†—Д–Њ—А–Љ–Є—А—Г–µ—В –Њ—Б–љ–Њ–≤—Г —Е–Њ—А–Є–Њ–љ–∞. –Ъ–∞–Ї —Г–ґ–µ –Њ—В–Љ–µ—З–∞–ї–Њ—Б—М, –Љ–∞–≥–љ–Є–є –≤–ї–Є—П–µ—В –љ–∞¬†—Б—В—А—Г–Ї—В—Г—А—Г —Б–Њ–µ–і–Є–љ–Є—В–µ–ї—М–љ–Њ–є —В–Ї–∞–љ–Є [47]. –Т¬†—Ж–µ–ї–Њ–Љ –≤¬†Mg-–Ј–∞–≤–Є—Б–Є–Љ–Њ–µ —А–µ–≥—Г–ї–Є—А–Њ–≤–∞–љ–Є–µ —Б–Њ—Б—В–Њ—П–љ–Є—П —Б–Њ–µ–і–Є–љ–Є—В–µ–ї—М–љ–Њ–є —В–Ї–∞–љ–Є –≤–Њ–≤–ї–µ—З–µ–љ—Л –њ–Њ¬†–Љ–µ–љ—М—И–µ–є –Љ–µ—А–µ 20 –±–µ–ї–Ї–Њ–≤. –Т–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –≤–ї–Є—П–љ–Є—П –і–µ—Д–Є—Ж–Є—В–∞ –Љ–∞–≥–љ–Є—П –љ–∞¬†—Б–Є–љ—В–µ–Ј –Є¬†–і–µ–≥—А–∞–і–∞—Ж–Є—О —Б–Њ–µ–і–Є–љ–Є—В–µ–ї—М–љ–Њ–є —В–Ї–∞–љ–Є –≤–Ї–ї—О—З–∞—О—В –∞–Ї—В–Є–≤–∞—Ж–Є—О –Љ–∞—В—А–Є—З–љ—Л—Е –Љ–µ—В–∞–ї–ї–Њ–њ—А–Њ—В–µ–Є–љ–∞–Ј, –ї–Є–Ј–Є–ї–Њ–Ї—Б–Є–і–∞–Ј—Л,¬†–≥–ї—Г—В–∞–Љ–Є–љ–∞–Ј—Л, –Ј–∞–Љ–µ–і–ї–µ–љ–Є–µ —Б–Є–љ—В–µ–Ј–∞ –Ї–Њ–ї–ї–∞–≥–µ–љ–∞, —Н–ї–∞—Б—В–Є–љ–∞ –Є¬†–≥–Є–∞–ї–Њ—Г—А–Њ–љ–∞–љ–∞. –Я—А–Є –і–µ—Д–Є—Ж–Є—В–µ –Љ–∞–≥–љ–Є—П –±–µ–ї–Ї–Њ–≤—Л–є —Б–Є–љ—В–µ–Ј —Б–Њ–µ–і–Є–љ–Є—В–µ–ї—М–љ–Њ–є —В–Ї–∞–љ–Є –Ј–∞–Љ–µ–і–ї—П–µ—В—Б—П, –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –Љ–∞—В—А–Є—З–љ—Л—Е –Љ–µ—В–∞–ї–ї–Њ–њ—А–Њ—В–µ–Є–љ–∞–Ј —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В—Б—П –Є¬†–≤–љ–µ–Ї–ї–µ—В–Њ—З–љ–∞—П –Љ–∞—В—А–Є—Ж–∞ –њ—А–Њ–≥—А–µ—Б—Б–Є–≤–љ–Њ –і–µ–≥—А–∞–і–Є—А—Г–µ—В, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г —Б—В—А—Г–Ї—В—Г—А–љ–∞—П –њ–Њ–і–і–µ—А–ґ–Ї–∞ —В–Ї–∞–љ–Є (–Ї–Њ–ї–ї–∞–≥–µ–љ–Њ–≤—Л–µ –≤–Њ–ї–Њ–Ї–љ–∞) —А–∞–Ј—А—Г—И–∞–µ—В—Б—П –±—Л—Б—В—А–µ–µ, —З–µ–Љ —Б–Є–љ—В–µ–Ј–Є—А—Г–µ—В—Б—П.
–Я—А–Є–≤–µ–і–µ–љ–љ—Л–є –≤—Л—И–µ –∞–љ–∞–ї–Є–Ј –±–Є–Њ—Е–Є–Љ–Є–Є –Є¬†—Б–Є—Б—В–µ–Љ–љ–Њ–є –±–Є–Њ–ї–Њ–≥–Є–Є –Њ—А–Њ—В–∞—В–∞ –њ–Њ–Ј–≤–Њ–ї—П–µ—В —Б—Д–Њ—А–Љ—Г–ї–Є—А–Њ–≤–∞—В—М –Њ–±–Њ–±—Й–µ–љ–љ—Г—О –Љ–Њ–і–µ–ї—М –µ–≥–Њ —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –љ–∞¬†–Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ–Њ–Љ —Г—А–Њ–≤–љ–µ (—А–Є—Б.¬†8). –Ш–Њ–љ—Л –Љ–∞–≥–љ–Є—П –Є¬†–Њ—А–Њ—В–∞—В–∞ –Њ–Ї–∞–Ј—Л–≤–∞—О—В —В–µ—А–∞–њ–µ–≤—В–Є—З–µ—Б–Ї–Њ–µ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ —З–µ—А–µ–Ј —А–∞–Ј–ї–Є—З–љ—Л–µ –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л. –Ь–∞–≥–љ–Є–є —В–∞–Ї–ґ–µ –≤–Њ–Ј–і–µ–є—Б—В–≤—Г–µ—В –љ–∞¬†—Б–Є–љ—В–µ–Ј –£–Ь–§ –Є¬†–≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ—Г—О –њ–µ—А–µ–і–∞—З—Г —Б–Є–≥–љ–∞–ї–∞ –Њ—В¬†–њ—Г—А–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤. –Т–∞–Ј–Њ–і–Є–ї–∞—В–∞—Ж–Є—П —В–Њ–ґ–µ –≤–ї–Є—П–µ—В –љ–∞¬†–≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М –Ї–ї–µ—В–Њ–Ї –њ—А–Є –Є—И–µ–Љ–Є–Є, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г —Г–≤–µ–ї–Є—З–µ–љ–Є–µ —А–∞–Ј–Љ–µ—А–∞ –ї—О–Љ–µ–љ–∞ —Г–ї—Г—З—И–∞–µ—В –Ї—А–Њ–≤–Њ–Њ–±—А–∞—Й–µ–љ–Є–µ –Є¬†—Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В –±–Њ–ї–µ–µ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ–Љ—Г –Њ–±–Љ–µ–љ—Г –≤–µ—Й–µ—Б—В–≤ –≤¬†–Њ–±–ї–∞—Б—В–Є –Є–љ—Д–∞—А–Ї—В–∞.
–†–µ–Ј—Г–ї—М—В–∞—В—Л —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–Њ–≤ –љ–∞¬†–Ї—А–Њ–ї–Є–Ї–∞—Е –њ–Њ–Ї–∞–Ј—Л–≤–∞—О—В, —З—В–Њ –Њ—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ –Є¬†–Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г—О—В —Г–ї—Г—З—И–µ–љ–Є—О –ї–Є–њ–Є–і–љ–Њ–≥–Њ –њ—А–Њ—Д–Є–ї—П (—Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є–µ LDL/HDL) –њ–ї–∞–Ј–Љ—Л –Ї—А–Њ–≤–Є [48]. –Э–∞–Ї–Њ–њ–ї–µ–љ–Є–µ –ї–Є–њ–Є–і–Њ–≤ –≤¬†–њ–µ—З–µ–љ–Є, –≤–µ—А–Њ—П—В–љ–Њ, –Њ–±—Г—Б–ї–Њ–≤–ї–µ–љ–Њ –њ–Њ–≤—Л—И–µ–љ–Є–µ–Љ —Н–Ї—Б–њ—А–µ—Б—Б–Є–Є –∞—А–Є–ї–∞—Ж–µ—В–∞–Љ–Є–і-–і–µ–∞—Ж–µ—В–Є–ї–∞–Ј—Л [49] –Є¬†—Д–Њ—Б—Д–∞—В–Є–і–∞—В-—Д–Њ—Б—Д–Њ–≥–Є–і—А–Њ–ї–∞–Ј—Л, –≤¬†—В–Њ –≤—А–µ–Љ—П –Ї–∞–Ї –≤—Л–Ј–≤–∞–љ–љ–Њ–µ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є —Б–љ–Є–ґ–µ–љ–Є–µ –≤¬†—Б—Л–≤–Њ—А–Њ—В–Ї–µ –Ї—А–Њ–≤–Є –ї–Є–њ–Є–і–Њ–≤ —Б–Ї–Њ—А–µ–µ –≤—Б–µ–≥–Њ —А–µ–Ј—Г–ї—М—В–∞—В –њ–Њ–≤—Л—И–µ–љ–Є—П —Г—А–Њ–≤–љ—П –Є¬†–∞–Ї—В–Є–≤–љ–Њ—Б—В–Є —Д–Њ—Б—Д–Њ–ї–Є–њ–∞–Ј—Л C [50]. –°—В–µ–њ–µ–љ—М –≤–ї–Є—П–љ–Є—П –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –љ–∞¬†–ї–Є–њ–Є–і—Л –њ–µ—З–µ–љ–Є –Ј–∞–≤–Є—Б–Є—В –Њ—В¬†–≤–Є–і–∞ –њ–Є—Й–µ–≤–Њ–≥–Њ –±–µ–ї–Ї–∞: –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Њ–µ –≤–Њ–Ј—А–∞—Б—В–∞–љ–Є–µ —Г—А–Њ–≤–љ—П –ї–Є–њ–Є–і–Њ–≤ –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М –њ—А–Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–Є –і–Є–µ—В, –Њ–±–Њ–≥–∞—Й–µ–љ–љ—Л—Е –Ї–∞–Ј–µ–Є–љ–Њ–Љ [51].
–Я–Њ–Ї–∞–Ј–∞–љ–Њ –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ –љ–∞¬†–≤—Л–ґ–Є–≤–∞–љ–Є–µ –њ–∞—Ж–Є–µ–љ—В–Њ–≤, –њ–µ—А–µ–љ–µ—Б—И–Є—Е –Є–љ—Д–∞—А–Ї—В –Љ–Є–Њ–Ї–∞—А–і–∞. –Т¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є 79 –±–Њ–ї—М–љ—Л—Е —Б¬†—В—П–ґ–µ–ї–Њ–є —Б–µ—А–і–µ—З–љ–Њ–є –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ—Б—В—М—О –њ—А–Њ—И–ї–Є –Њ–і–љ–Њ–≥–Њ–і–Є—З–љ—Л–є –Ї—Г—А—Б –ї–µ—З–µ–љ–Є—П —Б¬†–Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞. –°–Њ–≥–ї–∞—Б–љ–Њ –њ–Њ–ї—Г—З–µ–љ–љ—Л–Љ –і–∞–љ–љ—Л–Љ, –≤—Л–ґ–Є–≤–∞–љ–Є–µ —Б–Њ—Б—В–∞–≤–Є–ї–Њ 76% –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†52% –≤¬†–≥—А—Г–њ–њ–µ –њ–ї–∞—Ж–µ–±–Њ [52]. –Ш—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В –њ–µ—А–µ–љ–Њ—Б–Є–Љ–Њ—Б—В—М —Д–Є–Ј–Є—З–µ—Б–Ї–Є—Е –љ–∞–≥—А—Г–Ј–Њ–Ї —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–Є—И–µ–Љ–Є—З–µ—Б–Ї–Њ–є –±–Њ–ї–µ–Ј–љ—М—О —Б–µ—А–і—Ж–∞ (–Ш–С–°) [53]. –Э–∞¬†—Д–Њ–љ–µ –њ—А–Є–Љ–µ–љ–µ–љ–Є—П –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ —Г¬†–њ–Њ–ґ–Є–ї—Л—Е –њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–Ш–С–° —Г–ї—Г—З—И–∞—О—В—Б—П –њ–∞—А–∞–Љ–µ—В—А—Л –Ї–∞—З–µ—Б—В–≤–∞ –ґ–Є–Ј–љ–Є –Є¬†–љ–∞–±–ї—О–і–∞—О—В—Б—П –њ–Њ–Ј–Є—В–Є–≤–љ—Л–µ –Є–Ј–Љ–µ–љ–µ–љ–Є—П –њ—Б–Є—Е–Њ—Н–Љ–Њ—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–≥–Њ —Б—В–∞—В—Г—Б–∞ [54]. –Я–Њ–Ј–Є—В–Є–≤–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –Њ—А–Њ—В–∞—В–∞ –Љ–∞–≥–љ–Є—П –≤¬†—Н—В–Є—Е —Б–ї—Г—З–∞—П—Е –Љ–Њ–ґ–µ—В –±—Л—В—М —Б–≤—П–Ј–∞–љ–Њ –Ї–∞–Ї —Б¬†—Г–≤–µ–ї–Є—З–µ–љ–Є–µ–Љ –≤–∞–Ј–Њ–і–Є–ї–∞—В–∞—Ж–Є–Є –Є¬†—Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–Њ–≥–Њ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ –Ї–∞—А–і–Є–Њ–Љ–Є–Њ¬≠—Ж–Є—В–Њ–≤, —В–∞–Ї –Є¬†—Б –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ—Л–Љ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ–Љ –љ–∞¬†—Б—В—А—Г–Ї—В—Г—А—Г —Б–Њ–µ–і–Є–љ–Є—В–µ–ї—М–љ–Њ–є —В–Ї–∞–љ–Є –Љ–Є–Њ–Ї–∞—А–і–∞, —З—В–Њ –љ–∞–≥–ї—П–і–љ–Њ –њ–Њ–Ї–∞–Ј—Л–≤–∞–µ—В —Г—Б–њ–µ—И–љ–∞—П —В–µ—А–∞–њ–Є—П –њ—А–µ–њ–∞—А–∞—В–Њ–Љ –Ь–∞–≥–љ–µ—А–Њ—В –њ—А–Њ–ї–∞–њ—Б–∞ –Љ–Є—В—А–∞–ї—М–љ–Њ–≥–Њ –Ї–ї–∞–њ–∞–љ–∞ (–Я–Ь–Ъ) [55вАУ57].
–Т¬†–Љ–µ—В–∞–∞–љ–∞–ї–Є–Ј –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –њ—А–Є–Љ–µ–љ–µ–љ–Є—П –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ –≤¬†–Ї–∞—А–і–Є–Њ–ї–Њ–≥–Є–Є –≤–Њ—И–ї–Є –і–∞–љ–љ—Л–µ 19 —А–∞–љ–і–Њ–Љ–Є–Ј–Є—А–Њ–≤–∞–љ–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є, —Б—Г–Љ–Љ–∞—А–љ–Њ –≤–Ї–ї—О—З–∞–≤—И–Є—Е 603 –њ–∞—Ж–Є–µ–љ—В–∞ –Є–Ј¬†–≥—А—Г–њ–њ—Л ¬Ђ—Б–ї—Г—З–∞–є¬ї (–њ—А–Є–µ–Љ –Ь–∞–≥–љ–µ—А–Њ—В–∞) –Є¬†587 –Є–Ј¬†–≥—А—Г–њ–њ—Л ¬Ђ–Ї–Њ–љ—В—А–Њ–ї—М¬ї (–њ–ї–∞—Ж–µ–±–Њ) [58]. –£—Б—В–∞–љ–Њ–≤–ї–µ–љ—Л –і–Њ—Б—В–Њ–≤–µ—А–љ—Л–µ –∞—Б—Б–Њ—Ж–Є–∞—Ж–Є–Є –Љ–µ–ґ–і—Г –њ—А–Є–µ–Љ–Њ–Љ –њ—А–µ–њ–∞—А–∞—В–∞ –Є¬†—Б–љ–Є–ґ–µ–љ–Є–µ–Љ:¬†
-
–≥–Є–њ–Њ–Љ–∞–≥–љ–Є–µ–Љ–Є–Є (–Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ—Л–є —А–Є—Б–Ї (–Ю–†) 0,06; 95%-–љ—Л–є –і–Њ–≤–µ—А–Є—В–µ–ї—М–љ—Л–є –Є–љ—В–µ—А–≤–∞–ї (–Ф–Ш) 0,04вАУ0,09; p = 2,5 √Ч 10-46);
-
–љ–µ–њ–µ—А–µ–љ–Њ—Б–Є–Љ–Њ—Б—В–Є —Д–Є–Ј–Є—З–µ—Б–Ї–Њ–є –љ–∞–≥—А—Г–Ј–Ї–Є (–Ю–† 0,41; 95% –Ф–Ш 0,27вАУ0,62; p = 0,0004);
-
—Б–Є–љ–і—А–Њ–Љ–∞ –≤–µ–≥–µ—В–∞—В–Є–≤–љ–Њ–є –і–Є—Б—В–Њ–љ–Є–Є (–Ю–† 0,08; 95% –Ф–Ш 0,04вАУ0,14; p = 2,4 √Ч 10-21);¬†
-
–≥–Њ–ї–Њ–≤–љ–Њ–є –±–Њ–ї–Є —Г—В—А–Њ–Љ (–Ю–† 0,16; 95% –Ф–Ш 0,09вАУ0,29; p = 1,6 √Ч 10-8);¬†
-
–≥–Њ–ї–Њ–≤–љ–Њ–є –±–Њ–ї–Є –љ–∞–њ—А—П–ґ–µ–љ–Є—П (–Ю–†¬†0,16; 95% –Ф–Ш 0,09вАУ0,27; p¬†= 5,4 √Ч 10-10);¬†
-
–≥–Њ–ї–Њ–≤–Њ–Ї—А—Г–ґ–µ–љ–Є—П (–Ю–† 0,28; 95% –Ф–Ш 0,15вАУ0,50; p = 0,0004);
-
–Я–Ь–Ъ 1-–є¬†—Б—В–µ–њ–µ–љ–Є (–Ю–† 0,05; 95% –Ф–Ш 0,03вАУ0,09; p = 1,2 √Ч 10-25);
-
—А–µ–≥—Г—А–≥–Є—В–∞—Ж–Є–Є 1-–є¬†—Б—В–µ–њ–µ–љ–Є (–Ю–† 0,29; 95% –Ф–Ш 0,14вАУ0,60; p = 0,0075);
-
–љ–∞–і–ґ–µ–ї—Г–і–Њ—З–Ї–Њ–≤–Њ–є —Н–Ї—Б—В—А–∞—Б–Є—Б—В–Њ–ї–Є–Є (–Ю–† 0,30; 95% –Ф–Ш 0,21вАУ0,44; p = 1,0 √Ч 10-8);
-
–ґ–µ–ї—Г–і–Њ—З–Ї–Њ–≤–Њ–є —Н–Ї—Б—В—А–∞—Б–Є—Б—В–Њ–ї–Є–Є (–Ю–† 0,48; 95% –Ф–Ш 0,30вАУ0,76; p = 0,019);
-
–њ–∞—А–Њ–Ї—Б–Є–Ј–Љ–∞–ї—М–љ–Њ–є –љ–∞–і–ґ–µ–ї—Г–і–Њ—З–Ї–Њ–≤–Њ–є —В–∞—Е–Є–Ї–∞—А–і–Є–Є (–Ю–† 0,28; 95% –Ф–Ш 0,15вАУ0,50; p = 0,0002);¬†
-
–≥–Є–њ–µ—А—В–Њ–љ–Є–Є (–Ю–† 0,32; 95% –Ф–Ш 0,17вАУ0,58; p = 0,0027).
–Ь–µ—В–∞–∞–љ–∞–ї–Є–Ј –њ–Њ–Ї–∞–Ј–∞–ї, —З—В–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ –њ–µ—А—Б–њ–µ–Ї—В–Є–≤–љ–Њ –љ–µ¬†—В–Њ–ї—М–Ї–Њ –і–ї—П —В–µ—А–∞–њ–Є–Є –Я–Ь–Ъ –Є¬†–Ї–Њ–Љ–њ–µ–љ—Б–∞—Ж–Є–Є¬†–≥–Є–њ–Њ–Љ–∞–≥–љ–Є–µ–Љ–Є–Є, –љ–Њ¬†–Є –і–ї—П –њ—А–Њ—Д–Є–ї–∞–Ї—В–Є–Ї–Є –Є¬†–ї–µ—З–µ–љ–Є—П –љ–∞—А—Г—И–µ–љ–Є–є —А–Є—В–Љ–∞ —Б–µ—А–і—Ж–∞, —А–µ–≥—Г–ї—П—Ж–Є–Є –∞—А—В–µ—А–Є–∞–ї—М–љ–Њ–≥–Њ –і–∞–≤–ї–µ–љ–Є—П –Є¬†—Г–ї—Г—З—И–µ–љ–Є—П —Д—Г–љ–Ї—Ж–Є–Њ–љ–Є—А–Њ–≤–∞–љ–Є—П –≤–µ–≥–µ—В–∞—В–Є–≤–љ–Њ–є –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л [58].
–•–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Њ–љ–љ—Л–є –∞–љ–∞–ї–Є–Ј –Љ–Њ–ї–µ–Ї—Г–ї—Л –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л
–£¬†–Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л/–Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞ –Ј–∞—Д–Є–Ї—Б–Є—А–Њ–≤–∞–љ—Л —Б–Њ–±—Б—В–≤–µ–љ–љ—Л–µ —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ —Н—Д—Д–µ–Ї—В—Л¬†вАУ —Б–Є–љ—В–µ–Ј –£–Ь–§, –∞–Ї—В–Є–≤–∞—Ж–Є—П –њ—Г—А–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –Є¬†–і—А. –Т–Њ–Ј–љ–Є–Ї–∞—О—В –Ј–∞–Ї–Њ–љ–Њ–Љ–µ—А–љ—Л–µ –≤–Њ–њ—А–Њ—Б—Л: –≤—Б–µ –ї–Є –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ —Н—Д—Д–µ–Ї—В—Л –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ –Є–Ј—Г—З–µ–љ—Л? –Ь–Њ–ґ–љ–Њ –ї–Є –Њ—Ж–µ–љ–Є—В—М –њ–µ—А—Б–њ–µ–Ї—В–Є–≤—Л –і–∞–ї—М–љ–µ–є—И–µ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ–Њ-—Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Б–≤–Њ–є—Б—В–≤ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ –њ–Њ¬†–љ–µ–Њ—З–µ–≤–Є–і–љ—Л–Љ –љ–∞–њ—А–∞–≤–ї–µ–љ–Є—П–Љ?
–Ь–µ—В–∞–±–Њ–ї–Њ–Љ–љ—Л–є –њ–Њ–і—Е–Њ–і –Ї¬†–∞–љ–∞–ї–Є–Ј—Г —Н—Д—Д–µ–Ї—В–Њ–≤ –Њ—А–Њ—В–∞—В–∞ –њ–Њ–Ј–≤–Њ–ї–Є–ї –Њ—Ж–µ–љ–Є—В—М —Н—В–Є –њ–µ—А—Б–њ–µ–Ї—В–Є–≤—Л. –Ъ–∞–Ї –Є–Ј–≤–µ—Б—В–љ–Њ, –Љ–љ–Њ–≥–Є–µ –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ—Л–µ —Б—А–µ–і—Б—В–≤–∞ –Љ–Є–Љ–Є–Ї—А–Є—А—Г—О—В –њ–Њ–і –Њ–њ—А–µ–і–µ–ї–µ–љ–љ—Л–µ –Љ–Њ–ї–µ–Ї—Г–ї—Л –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–∞, –Ј–∞ —Б—З–µ—В —З–µ–≥–Њ –Њ–Ї–∞–Ј—Л–≤–∞—О—В –Ї–∞–Ї –Њ—Б–љ–Њ–≤–љ–Њ–µ —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–µ –і–µ–є—Б—В–≤–Є–µ, —В–∞–Ї –Є¬†–њ–Њ–±–Њ—З–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л. –Я–Њ—Н—В–Њ–Љ—Г —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Є–µ —Б—Е–Њ–ґ–µ—Б—В–Є –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ–Њ–є —Б—В—А—Г–Ї—В—Г—А—Л –і–µ–є—Б—В–≤—Г—О—Й–µ–≥–Њ –љ–∞—З–∞–ї–∞ –њ—А–µ–њ–∞—А–∞—В–∞ (–≤¬†–і–∞–љ–љ–Њ–Љ —Б–ї—Г—З–∞–µ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞) —Б¬†–Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–Љ–Є —Б—В—А—Г–Ї—В—Г—А–∞–Љ–Є –і—А—Г–≥–Є—Е –Љ–Њ–ї–µ–Ї—Г–ї –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–∞ –њ—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ–Њ –≤–∞–ґ–љ–Њ –і–ї—П –≤—Л—П–≤–ї–µ–љ–Є—П –≤—Б–µ—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –Њ—А–Њ—В–∞—В–∞ –љ–∞¬†–Њ—А–≥–∞–љ–Є–Ј–Љ [2]. –Ш–Љ–µ–µ—В—Б—П –≤¬†–≤–Є–і—Г –љ–µ–Ї–Є–є —Б–њ–µ–Ї—В—А —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–µ–є –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞.
–Ґ–∞–Ї–Њ–є –∞–љ–∞–ї–Є–Ј —Б—Е–Њ–ґ–µ—Б—В–Є –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л—Е —Б—В—А—Г–Ї—В—Г—А –љ–∞¬†—Г—А–Њ–≤–љ–µ –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–Њ–≤ –≤–Њ–Ј–Љ–Њ–ґ–µ–љ –≤¬†—А–∞–Љ–Ї–∞—Е –Љ–µ—В–Њ–і–Њ–ї–Њ–≥–Є–є —Е–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—В–Є–Ї–Є¬†вАУ –Њ–±–ї–∞—Б—В–Є –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –љ–∞¬†—Б—В—Л–Ї–µ —Б—В—А—Г–Ї—В—Г—А–љ–Њ–є —Е–Є–Љ–Є–Є, —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є–Є –Є¬†–±–Є–Њ–Є–љ—Д–Њ—А–Љ–∞—В–Є–Ї–Є, –≤¬†–Ї–Њ—В–Њ—А–Њ–є –≤–Ј–∞–Є–Љ–Њ—Б–≤—П–Ј—М ¬Ђ—Е–Є–Љ–Є—З–µ—Б–Ї–∞—П —Б—В—А—Г–Ї—В—Г—А–∞¬†вАУ —Б–≤–Њ–є—Б—В–≤–Њ –≤–µ—Й–µ—Б—В–≤–∞¬ї –Є—Б—Б–ї–µ–і—Г–µ—В—Б—П –Љ–µ—В–Њ–і–∞–Љ–Є —Б–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–є –Є–љ—Д–Њ—А–Љ–∞—В–Є–Ї–Є. –Р–љ–∞–ї–Є–Ј —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–µ–є —Е–Є–Љ–Є—З–µ—Б–Ї–Њ–є —Б—В—А—Г–Ї—В—Г—А—Л –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –њ—А–Њ–≤–Њ–і–Є—В—Б—П –њ—Г—В–µ–Љ —Б—А–∞–≤–љ–µ–љ–Є—П —Е–Є–Љ–Є—З–µ—Б–Ї–Њ–є —Б—В—А—Г–Ї—В—Г—А—Л –Љ–Њ–ї–µ–Ї—Г–ї—Л –Њ—А–Њ—В–∞—В–∞ —Б¬†–і–µ—Б—П—В–Ї–∞–Љ–Є —В—Л—Б—П—З –і—А—Г–≥–Є—Е –Љ–Њ–ї–µ–Ї—Г–ї –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–∞ –љ–∞¬†–Њ—Б–љ–Њ–≤–∞–љ–Є–Є —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л—Е –Љ–µ—В–Њ–і–Њ–≤ –њ—А–Є–Ї–ї–∞–і–љ–Њ–є –Љ–∞—В–µ–Љ–∞—В–Є–Ї–Є –Є¬†–Є–љ—В–µ–ї–ї–µ–Ї—В—Г–∞–ї—М–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –і–∞–љ–љ—Л—Е –љ–∞—Г—З–љ–Њ–є —И–Ї–Њ–ї—Л –∞–Ї–∞–і. –†–Р–Э –Ѓ.–Ш. –Ц—Г—А–∞–≤–ї–µ–≤–∞ [59вАУ62]. –•–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Њ–љ–љ—Л–є –њ–Њ–і—Е–Њ–і –њ–Њ–Ј–≤–Њ–ї—П–µ—В –≤—Л—З–Є—Б–ї—П—В—М —Е–Є–Љ–Є—З–µ—Б–Ї–Њ–µ —А–∞—Б—Б—В–Њ—П–љ–Є–µ dѕЗ –Љ–µ–ґ–і—Г –і–≤—Г–Љ—П –њ—А–Њ–Є–Ј–≤–Њ–ї—М–љ—Л–Љ–Є –Љ–Њ–ї–µ–Ї—Г–ї–∞–Љ–Є, –њ–Њ–Ј–≤–Њ–ї—П—О—Й–µ–µ –Њ—Ж–µ–љ–Є—В—М —Б—Е–Њ–ґ–µ—Б—В—М –Љ–Њ–ї–µ–Ї—Г–ї –њ–Њ¬†—Б—В—А—Г–Ї—В—Г—А–µ [63].
–Э–∞¬†–Њ—Б–љ–Њ–≤–∞–љ–Є–Є —Е–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Њ–љ–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –Љ–Њ–ї–µ–Ї—Г–ї—Л –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –±—Л–ї–Њ —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ —Б—Е–Њ–і—Б—В–≤–Њ –Љ–µ–ґ–і—Г –Љ–Њ–ї–µ–Ї—Г–ї–Њ–є –Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞ –Є¬†—А—П–і–Њ–Љ –Љ–Њ–ї–µ–Ї—Г–ї –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–∞ —З–µ–ї–Њ–≤–µ–Ї–∞ –Є¬†—А–∞–Ј–ї–Є—З–љ—Л–Љ–Є –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ—Л–Љ–Є —Б—А–µ–і—Б—В–≤–∞–Љ–Є. –Ъ–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Љ–Њ–≥—Г—В –Њ—Б—Г—Й–µ—Б—В–≤–ї—П—В—М—Б—П –Ј–∞ —Б—З–µ—В —А–µ–≥—Г–ї—П—Ж–Є–Є —Д–µ—А–Љ–µ–љ—В–∞ N-–∞—Ж–µ—В–Є–ї–≥–ї—О–Ї–Њ–Ј–∞–Љ–Є–љ —В—А–∞–љ—Б—Д–µ—А–∞–Ј—Л, –Є–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є—П —Д–Њ—Б—Д–Њ–і–Є—Н—Б—В–µ—А–∞–Ј –Є, –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ, –њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ –і–µ–є—Б—В–≤–Є—П. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, —А–µ–Ј—Г–ї—М—В–∞—В—Л —Е–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Њ–љ–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ —Г–Ї–∞–Ј—Л–≤–∞—О—В –љ–∞¬†–њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л–µ, –∞–љ—В–Є–Њ–Ї—Б–Є–і–∞–љ—В–љ—Л–µ, –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–Є–µ, –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л, –Ї–Њ—В–Њ—А—Л–µ –Њ—Б—В–∞—О—В—Б—П –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –љ–µ–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ—Л–Љ–Є.
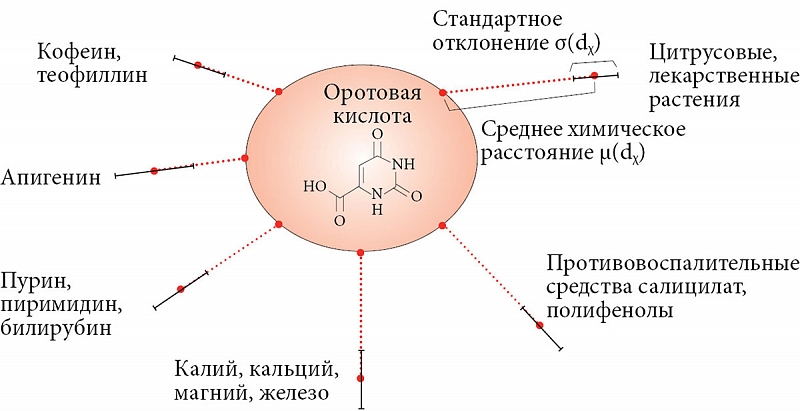
–°¬†–њ–Њ–Љ–Њ—Й—М—О —Е–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Њ–љ–љ–Њ–≥–Њ –Љ–µ—В–Њ–і–∞ –±—Л–ї –њ—А–Њ–≤–µ–і–µ–љ –∞–љ–∞–ї–Є–Ј —Б—Е–Њ–ґ–µ—Б—В–Є —Е–Є–Љ–Є—З–µ—Б–Ї–Њ–є —Б—В—А—Г–Ї—В—Г—А—Л –Љ–Њ–ї–µ–Ї—Г–ї—Л –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (–Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞) —Б¬†–Љ–Њ–ї–µ–Ї—Г–ї–∞–Љ–Є –≤¬†–±–∞–Ј–µ –і–∞–љ–љ—Л—Е –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–∞ —З–µ–ї–Њ–≤–µ–Ї–∞ –Є¬†–±–∞–Ј–∞—Е –і–∞–љ–љ—Л—Е –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ—Л—Е —Б—А–µ–і—Б—В–≤ [64]. –Т¬†—А–µ–Ј—Г–ї—М—В–∞—В–µ –њ—А–Є –∞–љ–∞–ї–Є–Ј–µ —Б–≤—Л—И–µ 40 000 –Љ–Њ–ї–µ–Ї—Г–ї –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–∞ —З–µ–ї–Њ–≤–µ–Ї–∞ –Є¬†2700 –Љ–Њ–ї–µ–Ї—Г–ї –Є–Ј–≤–µ—Б—В–љ—Л—Е –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ—Л—Е —Б—А–µ–і—Б—В–≤ —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ 632 –Љ–Њ–ї–µ–Ї—Г–ї—Л –љ–∞–Є–±–Њ–ї–µ–µ –±–ї–Є–Ј–Ї–Є –Ї¬†–Њ—А–Њ—В–∞—В—Г –њ–Њ¬†—Е–Є–Љ–Є—З–µ—Б–Ї–Њ–є —Б—В—А—Г–Ї—В—Г—А–µ. –Э–∞¬†–Њ—Б–љ–Њ–≤–∞–љ–Є–Є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є (–≤ —З–∞—Б—В–љ–Њ—Б—В–Є, –Ї–ї—О—З–µ–≤—Л—Е —Б–ї–Њ–≤), —Б–Њ–і–µ—А–ґ–∞—Й–µ–є—Б—П –≤¬†HMDB –Є¬†PubChem, –і–ї—П –Ї–∞–ґ–і–Њ–є –Є–Ј¬†—Н—В–Є—Е –Љ–Њ–ї–µ–Ї—Г–ї –±—Л–ї–Є –њ—А–Њ–∞–љ–∞–ї–Є–Ј–Є—А–Њ–≤–∞–љ—Л —Б—А–µ–і–љ–Є–µ —Е–Є–Љ–Є—З–µ—Б–Ї–Є–µ —А–∞—Б—Б—В–Њ—П–љ–Є—П. –†–µ–Ј—Г–ї—М—В–∞—В—Л –Њ–Ї–∞–Ј–∞–ї–Є—Б—М –≤–µ—Б—М–Љ–∞ –Є–љ—В–µ—А–µ—Б–љ—Л–Љ–Є (—А–Є—Б.¬†9).
–Т–Њ-–њ–µ—А–≤—Л—Е, —Б—А–µ–і–Є —Б—Е–Њ–ґ–Є—Е —Б¬†–Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–Њ–Љ –≤–µ—Й–µ—Б—В–≤ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–µ —З–Є—Б–ї–Њ –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–Њ–≤ —А–∞—Б—В–Є—В–µ–ї—М–љ—Л—Е —Н–Ї—Б—В—А–∞–Ї—В–Њ–≤. –Ю—А–Њ—В–∞—В —Б—Е–Њ–ґ —Б–Њ¬†–Љ–љ–Њ–≥–Є–Љ–Є –∞–ї–Ї–∞–ї–Њ–Є–і–∞–Љ–Є –Ї–Њ—А—Л, –Ї–Њ—А–љ–µ–є –Є¬†–њ–ї–Њ–і–Њ–≤ —Ж–Є—В—А—Г—Б–Њ–≤—Л—Е –Є¬†–і—А—Г–≥–Є—Е —А–∞—Б—В–µ–љ–Є–є (–љ–∞–њ—А–Є–Љ–µ—А, –∞–љ—В–Є–Њ–Ї—Б–Є–і–∞–љ—В –Љ–µ—В–Є–ї-2-–∞–Љ–Є–љ–Њ–±–µ–љ–Ј–Њ–є–љ–∞—П –Ї–Є—Б–ї–Њ—В–∞ (dѕЗ = 0,68) —Б–Њ–і–µ—А–ґ–Є—В—Б—П –≤¬†—Н—Д–Є—А–љ—Л—Е –Љ–∞—Б–ї–∞—Е –±–µ—А–≥–∞–Љ–Њ—В–∞, –ґ–∞—Б–Љ–Є–љ–∞, —З–µ—А–љ–Њ–≥–Њ —З–∞—П, —А–Є—Б–Њ–≤—Л—Е –Њ—В—А—Г–±–µ–є, –Ї–Њ–ґ—Г—А—Л –∞–њ–µ–ї—М—Б–Є–љ–∞ –Є¬†–ї–Є–Љ–Њ–љ–∞), –∞¬†—В–∞–Ї–ґ–µ —Б¬†—А—П–і–Њ–Љ –∞–ї–Ї–∞–ї–Њ–Є–і–Њ–≤ —А—Г—В—Л (–≥–Є–і—А–Њ–Ї—Б–Є-3-–Љ–µ—В–Њ–Ї—Б–Є-10-–Љ–Є—В–Є–ї–∞–Ї—А–Є–і–Њ–љ –Є¬†–і—А.). –Т¬†–љ–∞p–Њ–і–љ–Њ–є –Љ–µ–і–Є—Ж–Є–љ–µ —Н–Ї—Б—В—А–∞–Ї—В—Л —А—Г—В—Л (Ruta graveolens) –Є—Б–њ–Њ–ї—М–Ј—Г—О—В—Б—П –Ї–∞–Ї —Б—А–µ–і—Б—В–≤–∞, —Б–љ–Є–ґ–∞—О—Й–Є–µ —А–µ–≤–Љ–∞—В–Є—З–µ—Б–Ї–Є–µ –Є¬†–љ–µ–≤—А–∞–ї–≥–Є—З–µ—Б–Ї–Є–µ –±–Њ–ї–Є. –Э–∞¬†–і–Њ—Б—В–∞—В–Њ—З–љ–Њ –±–ї–Є–Ј–Ї–Њ–Љ —А–∞—Б—Б—В–Њ—П–љ–Є–Є (dѕЗ = 0,58) —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ—Л –∞–љ—В–Є–Њ–Ї—Б–Є–і–∞–љ—В—Л –∞–≤–µ–љ–∞–љ—В—А–∞–Љ–Є–і—Л –Р, –Т, –°,¬†D —Н–Ї—Б—В—А–∞–Ї—В–∞ –Ј–µ—А–µ–љ –Њ–≤—Б–∞ (Avena sativa). –Ь–љ–Њ–≥–Є–µ –Є–Ј¬†—А–∞—Б—В–Є—В–µ–ї—М–љ—Л—Е —Н–Ї—Б—В—А–∞–Ї—В–Њ–≤ —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—В—Б—П –њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л–Љ–Є —Б–≤–Њ–є—Б—В–≤–∞–Љ–Є, –љ–∞–њ—А–Є–Љ–µ—А —Б–∞–ї–Є—Ж–Є–ї–∞—В—Л (dѕЗ = 0,70вАУ0,75), –њ–Њ–ї–Є—Д–µ–љ–Њ–ї—Л —В–Є–њ–∞ —Ж–Є–љ–љ–∞–Љ–Њ–≤–Њ–є, –Ї–Њ—Д–µ–Є–љ–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В (dѕЗ = 0,7вАУ0,8) –Є¬†–∞–њ–Є–≥–µ–љ–Є–љ (dѕЗ = 0,63). –Ч–∞–Љ–µ—В–Є–Љ, —З—В–Њ –њ–Њ–ї–Є—Д–µ–љ–Њ–ї –∞–њ–Є–≥–µ–љ–Є–љ –≤—Е–Њ–і–Є—В –≤¬†—Б–Њ—Б—В–∞–≤ –њ—А–Њ–њ–Њ–ї–Є—Б–∞, —Б–µ–ї—М–і–µ—А–µ—П, –њ–µ—В—А—Г—И–Ї–Є, –∞–њ—В–µ—З–љ–Њ–є —А–Њ–Љ–∞—И–Ї–Є –Є¬†–Њ–±–ї–∞–і–∞–µ—В –Њ—В—З–µ—В–ї–Є–≤—Л–Љ–Є –њ—А–Њ—В–Є–≤–Њ—А–∞–Ї–Њ–≤—Л–Љ–Є —Б–≤–Њ–є—Б—В–≤–∞–Љ–Є.
–Т–Њ-–≤—В–Њ—А—Л—Е, –∞–љ–∞–ї–Є–Ј —Б—Е–Њ–ґ–Є—Е –њ–Њ¬†—Б—В—А—Г–Ї—В—Г—А–µ —Б¬†–Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є –Љ–Њ–ї–µ–Ї—Г–ї –њ–Њ–Ј–≤–Њ–ї–Є–ї –≤—Л–і–µ–ї–Є—В—М –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –і–µ—Б—П—В–Ї–Њ–≤ —Б–Њ–µ–і–Є–љ–µ–љ–Є–є, —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—Й–Є—Е—Б—П –Є–Ј–≤–µ—Б—В–љ–Њ–є —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В—М—О.
–Я—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ–Њ–µ –і–µ–є—Б—В–≤–Є–µ
–Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ —П–≤–ї—П–µ—В—Б—П –Њ–і–љ–Є–Љ –Є–Ј¬†–њ—А–Њ–Љ–µ–ґ—Г—В–Њ—З–љ—Л—Е –њ—А–Њ–і—Г–Ї—В–Њ–≤ –њ–Є—А–Є–Љ–Є–і–Є–љ–Њ–≤–Њ–≥–Њ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞. –Э–µ—Г–і–Є–≤–Є—В–µ–ї—М–љ–Њ, —З—В–Њ —Б—В—А—Г–Ї—В—Г—А–∞ –Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞ —Б—Е–Њ–ґ–∞ —Б–Њ¬†—Б—В—А—Г–Ї—В—Г—А–∞–Љ–Є —А–∞–Ј–ї–Є—З–љ—Л—Е –њ—А–Њ–Є–Ј–≤–Њ–і–љ—Л—Е –Љ–Њ—З–µ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (dѕЗ = 0,40вАУ0,50): 1-–Љ–µ—В–Є–ї–Љ–Њ—З–µ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (–Њ–і–Є–љ –Є–Ј¬†—В—А–µ—Е –Њ—Б–љ–Њ–≤–љ—Л—Е –Љ–µ—В–∞–±–Њ–ї–Є—В–Њ–≤ —В–µ–Њ—Д–Є–ї–ї–Є–љ–∞ [65]), 1,3-–і–Є–Љ–µ—В–Є–ї–Љ–Њ—З–µ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (–њ—А–Њ–і—Г–Ї—В –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ —В–µ–Њ—Д–Є–ї–ї–Є–љ–∞ [66]), 1,7-–і–Є–Љ–µ—В–Є–ї–Љ–Њ—З–µ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (–Њ—Б–љ–Њ–≤–љ–Њ–є –Љ–µ—В–∞–±–Њ–ї–Є—В –Ї–Њ—Д–µ–Є–љ–∞ –≤¬†–Љ–Њ—З–µ, –Њ–±—А–∞–Ј—Г–µ—В—Б—П –њ—А–Є –њ–Њ—Б—А–µ–і—Б—В–≤–µ —Ж–Є—В–Њ—Е—А–Њ–Љ–∞ CYP2A6 [67]). –Ь–Њ—З–µ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ —Б–ї—Г–ґ–Є—В –Љ–∞—А–Ї–µ—А–Њ–Љ —Б—В–∞—А–µ–љ–Є—П. –Х–µ –њ–Њ–≤—Л—И–µ–љ–љ—Л–µ —Г—А–Њ–≤–љ–Є –≤¬†–Њ—А–≥–∞–љ–Є–Ј–Љ–µ –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ—Л —Б¬†–і–Є–∞–±–µ—В–Њ–Љ –Є¬†—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–µ–Љ –Ї–∞–Љ–љ–µ–є –≤¬†–њ–Њ—З–Ї–∞—Е [68].
–°—Е–Њ–і—Б—В–≤–Њ —Б—В—А—Г–Ї—В—Г—А –Љ–Њ—З–µ–≤–Њ–є –Є¬†–Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В –њ–Њ–Ј–≤–Њ–ї—П–µ—В –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ –Њ—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ –Љ–Њ–ґ–µ—В —В–Њ—А–Љ–Њ–Ј–Є—В—М —Н—Д—Д–µ–Ї—В—Л –Љ–Њ—З–µ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л (–≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є, –Є–љ–≥–Є–±–Є—А–Њ–≤–∞—В—М –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є–µ —Г—А–∞—В–љ—Л—Е –Ї–∞–Љ–љ–µ–є –≤¬†–њ–Њ—З–Ї–∞—Е –Є¬†—Б–љ–Є–ґ–∞—В—М –≤–Њ—Б–њ–∞–ї–µ–љ–Є–µ).
–•–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Њ–љ–љ—Л–є –∞–љ–∞–ї–Є–Ј –њ–Њ–Ї–∞–Ј–∞–ї —Б—Е–Њ–і—Б—В–≤–Њ —Б—В—А—Г–Ї—В—Г—А—Л –Љ–Њ–ї–µ–Ї—Г–ї—Л –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л —Б–Њ¬†—Б—В—А—Г–Ї—В—Г—А–∞–Љ–Є —А—П–і–∞ –љ–µ—Б—В–µ—А–Њ–Є–і–љ—Л—Е –њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ (–Э–Я–Т–Я), —В–∞–Ї–Є–Љ–Є –Ї–∞–Ї –Љ–µ—Д–µ–љ–∞–Љ–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ (dѕЗ = 0,64), –Љ–µ–Ї–ї–Њ—Д–µ–љ–∞–Љ–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ (dѕЗ = 0,67), 3-–∞–Љ–Є–љ–Њ—Б–∞–ї–Є—Ж–Є–ї–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞, –∞–љ—В–Є–њ–Є—А–Є–љ, —Б–∞–ї–Є—Ж–Є–ї–∞–Љ–Є–і, –Є–љ–і–Њ–Љ–µ—В–∞—Ж–Є–љ, —В–Њ–ї–Љ–µ—В–Є–љ –Є¬†–љ–Є—Д–ї—Г–Љ–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ (dѕЗ = 0,73вАУ0,75). –Ю—Б–љ–Њ–≤—Л–≤–∞—П—Б—М –љ–∞¬†—Б—Е–Њ–і—Б—В–≤–µ –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ–Њ–є —Б—В—А—Г–Ї—В—Г—А—Л –Њ—А–Њ—В–∞—В–∞ —Б¬†–Э–Я–Т–Я, –Љ–Њ–ґ–љ–Њ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ –Њ—А–Њ—В–∞—В —Б–њ–Њ—Б–Њ–±–µ–љ –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Њ–≤–∞—В—М —Б¬†—Ж–Є–Ї–ї–Њ–Њ–Ї—Б–Є–≥–µ–љ–∞–Ј–Њ–є, –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –Ј–∞–Љ–µ–і–ї—П—П —Б–Є–љ—В–µ–Ј –њ—А–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л—Е –њ—А–Њ—Б—В–∞–≥–ї–∞–љ–і–Є–љ–Њ–≤. –Т¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–µ –њ—А–Є–µ–Љ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л —Б–љ–Є–ґ–∞–ї —Г—А–Њ–≤–љ–Є –Њ–Љ–µ–≥–∞-6-–њ–Њ–ї–Є–љ–µ–љ–∞—Б—Л—Й–µ–љ–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В, –Є–Ј¬†–Ї–Њ—В–Њ—А—Л—Е —Б–Є–љ—В–µ–Ј–Є—А—Г—О—В—Б—П –њ—А–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л–µ —Н–є–Ї–Њ–Ј–∞–љ–Њ–Є–і—Л –Є¬†–Ј–∞ —Б—З–µ—В –Ї–Њ—В–Њ—А—Л—Е –Њ—Б—Г—Й–µ—Б—В–≤–ї—П–µ—В—Б—П —А–µ–≥—Г–ї—П—Ж–Є—П –њ—А–Њ—Б—В–∞–≥–ї–∞–љ–і–Є–љ–Њ–≤–Њ–≥–Њ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞¬†[69].
–Ш–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї—П—Ж–Є—П –Є –њ—А–Њ—В–Є–≤–Њ–Њ–њ—Г—Е–Њ–ї–µ–≤–Њ–µ –і–µ–є—Б—В–≤–Є–µ
–†—П–і –Љ–Њ–ї–µ–Ї—Г–ї, —Б—Е–Њ–ґ–Є—Е –њ–Њ¬†—Б—В—А—Г–Ї—В—Г—А–µ —Б¬†–Њ—А–Њ—В–∞—В–Њ–Љ, —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—В—Б—П –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–Є–Љ —Н—Д—Д–µ–Ї—В–Њ–Љ. –Э–∞–њ—А–Є–Љ–µ—А, —В–Є–Љ–Є–љ (dѕЗ = 0,50)¬†вАУ –Њ–і–љ–Њ –Є–Ј¬†—З–µ—В—Л—А–µ—Е –Њ—Б–љ–Њ–≤–∞–љ–Є–є, –Њ–±—А–∞–Ј—Г—О—Й–Є—Е –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ—Б—В—М –Ф–Э–Ъ –Є¬†–Њ–±–ї–∞–і–∞—О—Й–Є—Е –њ—А–Њ—В–Є–≤–Њ–≤–Є—А—Г—Б–љ—Л–Љ –і–µ–є—Б—В–≤–Є–µ–Љ (–њ–Њ¬†–Њ—В–љ–Њ—И–µ–љ–Є—О –Ї¬†–≤–Є—А—Г—Б—Г¬†–≥—А–Є–њ–њ–∞) [70]. –°–Њ–µ–і–Є–љ–µ–љ–Є–µ 5-–≥–Є–і—А–Њ–Ї—Б–Є–Љ–µ—В–Є–ї—Г—А–∞—Ж–Є–ї (dѕЗ = 0,50)¬†вАУ –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї—П—В–Њ—А –Ї–ї–µ—В–Њ—З–љ–Њ–≥–Њ –Є¬†–≥—Г–Љ–Њ—А–∞–ї—М–љ–Њ–≥–Њ –Њ—В–≤–µ—В–∞ –Є—Б–њ–Њ–ї—М–Ј—Г–µ—В—Б—П –і–ї—П –ї–µ—З–µ–љ–Є—П —П–Ј–≤–µ–љ–љ–Њ–є –±–Њ–ї–µ–Ј–љ–Є –ґ–µ–ї—Г–і–Ї–∞ –Є¬†–њ—А–Є —Н—А–Њ–Ј–Є–≤–љ–Њ–Љ –њ–Њ—А–∞–ґ–µ–љ–Є–Є –Ї–Є—И–µ—З–љ–Є–Ї–∞ [71, 72], –∞¬†5-–≥–Є–і—А–Њ–Ї—Б–Є–Љ–µ—В–Є–ї-4-–Љ–µ—В–Є–ї—Г—А–∞—Ж–Є–ї —Б—В–Є–Љ—Г–ї–Є—А—Г–µ—В –ї–µ–є–Ї–Њ—Ж–Є—В–Њ–њ–Њ—Н–Ј (dѕЗ = 0,60).
–Ш–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–µ–µ –і–µ–є—Б—В–≤–Є–µ –њ—А–Є—Б—Г—Й–µ –∞–љ—В–Є–Њ–Ї—Б–Є–і–∞–љ—В–∞–Љ —А–∞—Б—В–Є—В–µ–ї—М–љ—Л—Е —Н–Ї—Б—В—А–∞–Ї—В–Њ–≤¬†вАУ —В–µ–Љ—Г—А–Є–љ—Г, –∞–≤–µ–љ–∞–љ—В—А–∞–Љ–Є–і–∞–Љ, –Љ—Г–Ї–Њ–љ–Є–љ—Г, –њ–Є–њ–µ—А–Њ–ї–∞–Ї—В–∞–Љ—Г. –Ґ–µ–Љ—Г—А–Є–љ, –Є–ї–Є —В–µ–∞–Ї—А–Є–љ (dѕЗ = 0,52),¬†вАУ –њ—Г—А–Є–љ–Њ–≤—Л–є –∞–ї–Ї–∞–ї–Њ–Є–і –њ–ї–Њ–і–Њ–≤–Њ–≥–Њ –і–µ—А–µ–≤–∞ –Ї—Г–њ—Г–∞—Б—Г (Theobroma grandiflorum, —Б–µ–Љ–µ–є—Б—В–≤–Њ ¬Ђ–Ь–∞–ї—М–≤–Њ–≤—Л–µ¬ї) –Є¬†–Ї–∞–Љ–µ–ї–Є–Є (Camellia assamica, —Б–µ–Љ–µ–є—Б—В–≤–Њ ¬Ђ–І–∞–є–љ—Л–µ¬ї) —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г–µ—В—Б—П –њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ—Л–Љ –Є¬†–Њ–±–µ–Ј–±–Њ–ї–Є–≤–∞—О—Й–Є–Љ –і–µ–є—Б—В–≤–Є–µ–Љ [73]. –Я–Њ–ї–Є—Д–µ–љ–Њ–ї –Њ–≤—Б–∞ –∞–≤–µ–љ–∞–љ—В—А–∞–Љ–Є–і L –Є¬†–і—А—Г–≥–Є–µ –∞–≤–µ–љ–∞–љ—В—А–∞–Љ–Є–і—Л (dѕЗ = 0,52) –Њ—В–ї–Є—З–∞—О—В—Б—П –∞–љ—В–Є–Њ–Ї—Б–Є–і–∞–љ—В–љ—Л–Љ –њ–Њ—В–µ–љ—Ж–Є–∞–ї–Њ–Љ –Є¬†–њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В—М—О [74]. –Я–Є–њ–µ—А–Њ–ї–∞–Ї—В–∞–Љ¬†–Р¬†(dѕЗ = 0,61), –∞–ї–Ї–∞–ї–Њ–Є–і –Ї–Њ—А–љ–µ–є –Њ—Б–Њ–±–Њ–≥–Њ –≤–Є–і–∞ –Є–љ–і–Є–є—Б–Ї–Њ–≥–Њ –њ–µ—А—Ж–∞ (Piper longum), –Њ–Ї–∞–Ј—Л–≤–∞–µ—В –њ—А–Њ—В–Є–≤–Њ–Њ–њ—Г—Е–Њ–ї–µ–≤–Њ–µ –і–µ–є—Б—В–≤–Є–µ [75].
–Э–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л
–Ю—В–Љ–µ—З–µ–љ–Њ —Б—Е–Њ–і—Б—В–≤–Њ —Б—В—А—Г–Ї—В—Г—А—Л –Љ–Њ–ї–µ–Ї—Г–ї—Л –Њ—А–Њ—В–∞—В–∞ —Б¬†—А—П–і–Њ–Љ –љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л—Е –Љ–µ—В–∞–±–Њ–ї–Є—В–Њ–≤, –њ—А–µ–ґ–і–µ –≤—Б–µ–≥–Њ —Б¬†–Ї–Є–љ—Г—А–µ–љ–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є (dѕЗ = 0,45). –Ъ–Є–љ—Г—А–µ–љ–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞¬†вАУ —Н–љ–і–Њ–≥–µ–љ–љ—Л–є –∞–љ—В–∞–≥–Њ–љ–Є—Б—В¬†–≥–ї—Г—В–∞–Љ–∞—В–љ—Л—Е –Є–Њ–љ–Њ—В—А–Њ–њ–љ—Л—Е (NMDA) —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ —Б¬†–љ–µ–є—А–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–Љ –Є¬†–њ—А–Њ—В–Є–≤–Њ—Б—Г–і–Њ—А–Њ–ґ–љ—Л–Љ –і–µ–є—Б—В–≤–Є–µ–Љ [76, 77].
–Ъ–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л
–Ґ—А–∞–і–Є—Ж–Є–Њ–љ–љ–Њ –Ї–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ —Б–≤—П–Ј—Л–≤–∞–ї–Є —Б¬†–Ї–Њ–Љ–њ–µ–љ—Б–∞—Ж–Є–µ–є –і–µ—Д–Є—Ж–Є—В–∞ –Љ–∞–≥–љ–Є—П —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–Њ—Б—В—А—Л–Љ –Є–љ—Д–∞—А–Ї—В–Њ–Љ –Љ–Є–Њ–Ї–∞—А–і–∞, –Я–Ь–Ъ, –∞–ї–Ї–Њ–≥–Њ–ї—М–љ–Њ–є –Љ–Є–Њ–Ї–∞—А–і–Є–Њ–і–Є—Б—В—А–Њ—Д–Є–µ–є, —В–∞—Е–Є–∞—А–Є—В–Љ–Є–µ–є –Є¬†–і—А. –†–µ–Ј—Г–ї—М—В–∞—В—Л —Е–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Њ–љ–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ —Г–Ї–∞–Ј—Л–≤–∞—О—В –љ–∞¬†–Є–љ—Л–µ –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л–µ –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л, –њ–Њ—Б—А–µ–і—Б—В–≤–Њ–Љ –Ї–Њ—В–Њ—А—Л—Е –Љ–Њ–ї–µ–Ї—Г–ї–∞ –Њ—А–Њ—В–∞—В–∞ –Љ–Њ–ґ–µ—В —Б—В–Є–Љ—Г–ї–Є—А–Њ–≤–∞—В—М –Ї–∞—А–і–Є–Њ¬≠–њ—А–Њ—В–µ–Ї—Ж–Є—О. –Ъ¬†—В–∞–Ї–Є–Љ –Љ–µ—Е–∞–љ–Є–Ј–Љ–∞–Љ –Њ—В–љ–Њ—Б—П—В—Б—П –њ—А–µ–ґ–і–µ –≤—Б–µ–≥–Њ —А–µ–≥—Г–ї—П—Ж–Є—П —Д–µ—А–Љ–µ–љ—В–∞ N-–∞—Ж–µ—В–Є–ї¬≠–≥–ї—О–Ї–Њ–Ј–∞–Љ–Є–љ —В—А–∞–љ—Б—Д–µ—А–∞–Ј—Л –Є¬†–Є–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є–µ —Д–Њ—Б—Д–Њ–і–Є—Н—Б—В–µ—А–∞–Ј.
–†–µ–≥—Г–ї—П—Ж–Є—П N-–∞—Ж–µ—В–Є–ї–≥–ї—О–Ї–Њ–Ј–∞–Љ–Є–љ —В—А–∞–љ—Б—Д–µ—А–∞–Ј—Л. –Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г–µ—В—Б—П –≤—Л—Б–Њ–Ї–Њ–є —Б—В–µ–њ–µ–љ—М—О —Б—Е–Њ–і—Б—В–≤–∞ —Б¬†—Г—А–∞—Ж–Є–ї–Њ–Љ (dѕЗ = 0,44). –£—А–∞—Ж–Є–ї –Є¬†–µ–≥–Њ —Д–Њ—Б—Д–∞—В-–њ—А–Њ–Є–Ј–≤–Њ–і–љ—Л–µ (–љ–∞–њ—А–Є–Љ–µ—А, –£–Ф–§) —Б–ї—Г–ґ–∞—В –∞–ї–ї–Њ—Б—В–µ—А–Є—З–µ—Б–Ї–Є–Љ–Є —А–µ–≥—Г–ї—П—В–Њ—А–∞–Љ–Є –Є¬†–Ї–Њ—Д–µ—А–Љ–µ–љ—В–∞–Љ–Є –і–ї—П —А—П–і–∞ –±–Є–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Є—Е —А–µ–∞–Ї—Ж–Є–є. –Т¬†—З–∞—Б—В–љ–Њ—Б—В–Є, –£–Ф–§-–≥–ї—О–Ї–Њ–Ј–∞ —А–µ–≥—Г–ї–Є—А—Г–µ—В –њ—А–µ–≤—А–∞—Й–µ–љ–Є–µ¬†–≥–ї—О–Ї–Њ–Ј—Л –Є¬†–≥–∞–ї–∞–Ї—В–Њ–Ј—Л –≤¬†–њ–µ—З–µ–љ–Є –Є¬†–≤ –і—А—Г–≥–Є—Е —В–Ї–∞–љ—П—Е. –£—А–∞—Ж–Є–ї —П–≤–ї—П–µ—В—Б—П —А–µ–≥—Г–ї–Є—А—Г—О—Й–Є–Љ –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–Њ–Љ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ –±–µ—В–∞-–∞–ї–∞–љ–Є–љ–∞, –±–Є–Њ—Б–Є–љ—В–µ–Ј–∞ –њ–∞–љ—В–Њ—В–µ–љ–∞—В–∞ –Є¬†–Ї–Њ—Д–µ—А–Љ–µ–љ—В–∞ –Р, –∞¬†—В–∞–Ї–ґ–µ —Д–µ—А–Љ–µ–љ—В–∞ N-–∞—Ж–µ—В–Є–ї–≥–ї—О–Ї–Њ–Ј–∞–Љ–Є–љ —В—А–∞–љ—Б—Д–µ—А–∞–Ј—Л (–≥–µ–љ OGT) [78], –Ї–Њ—В–Њ—А—Л–є —Г—З–∞—Б—В–≤—Г–µ—В –≤¬†–њ–Њ–і–і–µ—А–ґ–Ї–µ —А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є –Ї¬†–Є–љ—Б—Г–ї–Є–љ—Г –≤¬†–Љ—Л—И—Ж–∞—Е –Є¬†–Ї–ї–µ—В–Ї–∞—Е –∞–і–Є–њ–Њ—Ж–Є—В–Њ–≤. –§–µ—А–Љ–µ–љ—В –Є–љ–≥–Є–±–Є—А—Г–µ—В—Б—П –£–Ф–§ [78] –Є, –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ, –љ–µ–њ–Њ—Б—А–µ–і—Б—В–≤–µ–љ–љ–Њ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є. –Ґ–Њ—А–Љ–Њ–ґ–µ–љ–Є–µ –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є —Д–µ—А–Љ–µ–љ—В–∞ —Б—Е–Њ–ґ–Є–Љ —Б¬†–£–Ф–§ –Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–Њ–Љ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В –∞–Ї—В–Є–≤–∞—Ж–Є–Є —Г–≥–ї–µ–≤–Њ–і–љ–Њ–≥–Њ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ –Є¬†—Б–љ–Є–ґ–µ–љ–Є—О –∞–њ–Њ–њ—В–Њ–Ј–∞ –Ї–∞—А–і–Є–Њ–Љ–Є–Њ¬≠—Ж–Є—В–Њ–≤.
–Ш–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є–µ —Д–Њ—Б—Д–Њ–і–Є—Н—Б—В–µ—А–∞–Ј. –•–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Њ–љ–љ—Л–є –∞–љ–∞–ї–Є–Ј –њ–Њ–Ї–∞–Ј–∞–ї —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ–µ —Б—Е–Њ–і—Б—В–≤–Њ –Љ–µ–ґ–і—Г –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є –Є¬†—А—П–і–Њ–Љ —Б–Њ–µ–і–Є–љ–µ–љ–Є–є¬†вАУ –Є–љ–≥–Є–±–Є—В–Њ—А–Њ–≤ —Д–µ—А–Љ–µ–љ—В–Њ–≤ —Д–Њ—Б—Д–Њ–і–Є—Н—Б—В–µ—А–∞–Ј (—Н–љ–Њ–Ї—Б–Є–Љ–Њ–љ, –∞–Љ—А–Є–љ–Њ–љ, —В–µ–Њ—Д–Є–ї–ї–Є–љ, –Љ–Є–ї—А–Є–љ–Њ–љ). –Ъ–∞–Ї –Є–Ј–≤–µ—Б—В–љ–Њ, —Д–Њ—Б—Д–Њ–і–Є—Н—Б—В–µ—А–∞–Ј—Л —Г—З–∞—Б—В–≤—Г—О—В –≤¬†—А–µ–≥—Г–ї—П—Ж–Є–Є –≤–љ—Г—В—А–Є–Ї–ї–µ—В–Њ—З–љ–Њ–є –њ–µ—А–µ–і–∞—З–Є —Б–Є–≥–љ–∞–ї–∞ –Њ—В¬†—А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ —А–∞–Ј–ї–Є—З–љ—Л—Е —В–Є–њ–Њ–≤ (–∞–і—А–µ–љ–µ—А–≥–Є—З–µ—Б–Ї–Є—Е, –Љ–µ—В–∞–±–Њ—В—А–Њ–њ–љ—Л—Е –∞—Ж–µ—В–Є–ї—Е–Њ–ї–Є–љ–Њ–≤—Л—Е –Є¬†–≥–ї—Г—В–∞–Љ–∞—В–љ—Л—Е, –і–Њ—Д–∞–Љ–Є–љ–Њ–≤—Л—Е, –∞–і–µ–љ–Њ–Ј–Є–љ–Њ–≤—Л—Е,¬†–≥–Є—Б—В–∞–Љ–Є–љ–Њ–≤—Л—Е,¬†–≥–ї—О–Ї–∞–≥–Њ–љ–Њ–≤—Л—Е –Є¬†–і—А.) [79]. –§–µ—А–Љ–µ–љ—В 3',5'-—Д–Њ—Б—Д–Њ–і–Є—Н—Б—В–µ—А–∞–Ј–∞ –і–µ–≥—А–∞–і–Є—А—Г–µ—В —Ж–Є–Ї–ї–Є—З–µ—Б–Ї–Є–є –Р–Ь–§, —В–µ–Љ —Б–∞–Љ—Л–Љ –Њ—Б–ї–∞–±–ї—П—П –њ–µ—А–µ–і–∞—З—Г —Б–Є–≥–љ–∞–ї–Њ–≤ –Њ—В¬†—Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г—О—Й–Є—Е —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤. –Ш¬†–љ–∞–Њ–±–Њ—А–Њ—В, –Є–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є–µ —Д–Њ—Б—Д–Њ–і–Є—Н—Б—В–µ—А–∞–Ј—Л —Г–ї—Г—З—И–∞–µ—В –њ–µ—А–µ–і–∞—З—Г —Б–Є–≥–љ–∞–ї–∞ –њ–Њ¬†—Б–Є–≥–љ–∞–ї—М–љ—Л–Љ –Ї–∞—Б–Ї–∞–і–∞–Љ. –Э–∞–њ—А–Є–Љ–µ—А, —Н–љ–Њ–Ї—Б–Є–Љ–Њ–љ (dѕЗ = 0,56)¬†вАУ —Б–µ–ї–µ–Ї—В–Є–≤–љ—Л–є –Є–љ–≥–Є–±–Є—В–Њ—А —Д–Њ—Б—Д–Њ–і–Є—Н—Б—В–µ—А–∞–Ј. –•–∞—А–∞–Ї—В–µ—А–Є–Ј—Г–µ—В—Б—П —Б–Њ—Б—Г–і–Њ—А–∞—Б—И–Є—А—П—О—Й–Є–Љ –і–µ–є—Б—В–≤–Є–µ–Љ –Є¬†–љ–Њ–Њ—В—А–Њ–њ–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В—М—О, –Ї–Њ—В–Њ—А–∞—П –љ–µ¬†–≤—Л–Ј—Л–≤–∞–µ—В –Є–Ј–Љ–µ–љ–µ–љ–Є–є –≤¬†–њ–Њ—В—А–µ–±–ї–µ–љ–Є–Є –Ї–Є—Б–ї–Њ—А–Њ–і–∞ –Љ–Є–Њ–Ї–∞—А–і–Њ–Љ. –Ш—Б–њ–Њ–ї—М–Ј—Г–µ—В—Б—П —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†—Е—А–Њ–љ–Є—З–µ—Б–Ї–Њ–є —Б–µ—А–і–µ—З–љ–Њ–є –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ—Б—В—М—О [80].
–•–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Њ–љ–љ—Л–є –∞–љ–∞–ї–Є–Ј —Г–Ї–∞–Ј–∞–ї –Є¬†–љ–∞ –і—А—Г–≥–Є–µ –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –Њ—Б—Г—Й–µ—Б—В–≤–ї–µ–љ–Є—П –Ї–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–≥–Њ —Н—Д—Д–µ–Ї—В–∞ –Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞. –Э–∞–њ—А–Є–Љ–µ—А, —Б—Е–Њ–ґ–∞—П —Б¬†–Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є 2-–Є–љ–і–Њ–ї–Ї–∞—А–±–Њ–љ–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ (dѕЗ = 0,50), –Њ–і–Є–љ –Є–Ј¬†–њ–Њ–ї–Є—Д–µ–љ–Њ–ї—М–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–Њ–≤ –ґ–µ–љ—М—И–µ–љ—П –Є¬†–і—А—Г–≥–Є—Е —А–∞—Б—В–Є—В–µ–ї—М–љ—Л—Е —Н–Ї—Б—В—А–∞–Ї—В–Њ–≤, —П–≤–ї—П–µ—В—Б—П —Б–Є–ї—М–љ—Л–Љ –Є–љ–≥–Є–±–Є—В–Њ—А–Њ–Љ –њ–µ—А–µ–Ї–Є—Б–љ–Њ–≥–Њ –Њ–Ї–Є—Б–ї–µ–љ–Є—П –ї–Є–њ–Є–і–Њ–≤ [81]. –°—Е–Њ–ґ–Є–є —Б¬†–Њ—А–Њ—В–∞—В–Њ–Љ –Њ–Ї—Б–Є–њ—Г—А–Є–љ–Њ–ї (dѕЗ = 0,55)¬†вАУ –Є–љ–≥–Є–±–Є—В–Њ—А –Ї—Б–∞–љ—В–Є–љ–Њ–Ї—Б–Є–і–∞–Ј—Л –Є—Б–њ–Њ–ї—М–Ј—Г–µ—В—Б—П –і–ї—П –ї–µ—З–µ–љ–Є—П —Е—А–Њ–љ–Є—З–µ—Б–Ї–Њ–є —Б–µ—А–і–µ—З–љ–Њ–є –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ—Б—В–Є [82]. –Ґ–µ–Њ–±—А–Њ–Љ–Є–љ (dѕЗ = 0,73) –Њ–±–ї–∞–і–∞–µ—В –љ–µ¬†—В–Њ–ї—М–Ї–Њ –±—А–Њ–љ—Е–Њ–ї–Є—В–Є—З–µ—Б–Ї–Є–Љ –Є¬†—Б–ї–∞–±—Л–Љ –Љ–Њ—З–µ–≥–Њ–љ–љ—Л–Љ, –љ–Њ¬†—В–∞–Ї–ґ–µ —Б–Њ—Б—Г–і–Њ—А–∞—Б—И–Є—А—П—О—Й–Є–Љ –Є¬†–Ї–∞—А–і–Є–Њ¬≠—Б—В–Є–Љ—Г–ї–Є—А—Г—О—Й–Є–Љ –і–µ–є—Б—В–≤–Є–µ–Љ [83].
–Т–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є–µ –Њ—А–Њ—В–∞—В–∞ –Љ–∞–≥–љ–Є—П —Б –і—А—Г–≥–Є–Љ–Є –≤–µ—Й–µ—Б—В–≤–∞–Љ–Є –Є –ї–µ–Ї–∞—А—Б—В–≤–µ–љ–љ—Л–Љ–Є —Б—А–µ–і—Б—В–≤–∞–Љ–Є
–Ю—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ –Є¬†–Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В –њ–ї–Њ—Е–Њ —А–∞—Б—В–≤–Њ—А—П—О—В—Б—П –≤¬†–≤–Њ–і–µ –Є¬†–Њ—А–≥–∞–љ–Є—З–µ—Б–Ї–Є—Е —А–∞—Б—В–≤–Њ—А–Є—В–µ–ї—П—Е. –Ь–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В —А–∞–Ј—А—Г—И–∞–µ—В—Б—П –њ–Њ–і –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ–Љ –њ—А—П–Љ—Л—Е —Б–Њ–ї–љ–µ—З–љ—Л—Е –ї—Г—З–µ–є. –Я—А–Є —Б–Њ–≤–Љ–µ—Б—В–љ–Њ–Љ –њ—А–Є–µ–Љ–µ —Б¬†—Д–Њ–ї–Є–µ–≤–Њ–є –Ї–Є—Б–ї–Њ—В–Њ–є –Є¬†—Д–Њ–ї–∞—В–∞–Љ–Є –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ —Г–ї—Г—З—И–∞–µ—В –Є—Е –±–Є–Њ–і–Њ—Б—В—Г–њ–љ–Њ—Б—В—М. –Ю—А–Њ—В–Њ–≤–∞—П –Ї–Є—Б–ї–Њ—В–∞ –њ—А–Є¬†–≥–Є–њ–Њ–≤–Є—В–∞–Љ–Є–љ–Њ–Ј–µ –Т12 —Б–њ–Њ—Б–Њ–±–љ–∞ –љ–Є–≤–µ–ї–Є—А–Њ–≤–∞—В—М –і–µ—Д–Є—Ж–Є—В —Ж–Є–∞–љ–Њ–Ї–Њ–±–∞–ї–∞–Љ–Є–љ–∞. –Я—А–Є–µ–Љ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ –≤–Њ –≤—А–µ–Љ—П –Ї—Г—А—Б–∞ –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Њ–≤ –њ–Њ—В–µ–љ—Ж–Є—А—Г–µ—В —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М –∞–љ—В–Є–±–∞–Ї—В–µ—А–Є–∞–ї—М–љ–Њ–≥–Њ –і–µ–є—Б—В–≤–Є—П –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Њ–≤ —А–∞–Ј–љ—Л—Е –Ї–ї–∞—Б—Б–Њ–≤ (–њ–µ–љ–Є—Ж–Є–ї–ї–Є–љ—Л, —Ж–µ—Д–∞–ї–Њ—Б–њ–Њ—А–Є–љ—Л –Є¬†–і—А.), —Б–љ–Є–ґ–∞–µ—В —А–Є—Б–Ї¬†–њ–Њ–±–Њ—З–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –Њ—В¬†–њ—А–Є–Љ–µ–љ–µ–љ–Є—П –∞–љ—В–Є–±–Є–Њ—В–Є–Ї–Њ–≤ (—Б–љ–Є–ґ–µ–љ–Є–µ —Б–ї—Г—Е–∞, –њ–Њ—А–∞–ґ–µ–љ–Є–µ –њ–Њ—З–µ–Ї, –љ–µ–є—А–Њ—В–Њ–Ї—Б–Є—З–µ—Б–Ї–Є–µ –њ—А–Њ—П–≤–ї–µ–љ–Є—П, –і–Є—Б–±–Є–Њ–Ј –Ї–Є—И–µ—З–љ–Є–Ї–∞ –Є¬†—В.–і.).
–Ч–∞–Ї–ї—О—З–µ–љ–Є–µ
–Ф–Њ–ї–≥–Њ–µ –≤—А–µ–Љ—П –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–µ —Н—Д—Д–µ–Ї—В—Л, –љ–∞–±–ї—О–і–∞–µ–Љ—Л–µ –њ—А–Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–Є –Њ—А–≥–∞–љ–Є—З–µ—Б–Ї–Є—Е —Б–Њ–ї–µ–є –Љ–∞–≥–љ–Є—П, –њ—А–Є–њ–Є—Б—Л–≤–∞–ї–Є—Б—М –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –Љ–∞–≥–љ–Є—О, –∞¬†—А–Њ–ї—М –Њ—А–≥–∞–љ–Є—З–µ—Б–Ї–Є—Е –∞–љ–Є–Њ–љ–Њ–≤ –Њ–≥—А–∞–љ–Є—З–Є–≤–∞–ї–∞—Б—М –њ–Њ–≤—Л—И–µ–љ–Є–µ–Љ –±–Є–Њ–і–Њ—Б—В—Г–њ–љ–Њ—Б—В–Є –Љ–∞–≥–љ–Є—П –њ—А–Є –њ–µ—А–Њ—А–∞–ї—М–љ–Њ–Љ –њ—А–Є–µ–Љ–µ –њ—А–µ–њ–∞—А–∞—В–Њ–≤. –Э–µ–Ј–∞–≤–Є—Б–Є–Љ—Л–µ –њ–Њ–Ј–Є—В–Є–≤–љ—Л–µ —Н—Д—Д–µ–Ї—В—Л –∞–љ–Є–Њ–љ–Њ–≤ –љ–∞¬†—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—О —З–µ–ї–Њ–≤–µ–Ї–∞, –Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ, –љ–µ¬†—А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–ї–Є—Б—М, –Є¬†–Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ –Њ—Б—В–∞–≤–∞–ї—Б—П –Њ–і–љ–Є–Љ –Є–Ј¬†–љ–∞–Є–Љ–µ–љ–µ–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ—Л—Е –∞–љ–Є–Њ–љ–Њ–≤, –Ї–Њ—В–Њ—А—Л–є, –Ї–∞–Ї —Б—З–Є—В–∞–ї–Њ—Б—М, –њ—А–Њ—Б—В–Њ —Г—З–∞—Б—В–≤—Г–µ—В –≤¬†—В—А–∞–љ—Б–њ–Њ—А—В–µ –Љ–∞–≥–љ–Є—П –≤–љ—Г—В—А—М –Ї–∞—А–і–Є–Њ–Љ–Є–Њ—Ж–Є—В–Њ–≤ –Є¬†–і—А—Г–≥–Є—Е –Ї–ї–µ—В–Њ–Ї.
–Р–љ–∞–ї–Є–Ј –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ—Л—Е –Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤ —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞, –≤—Е–Њ–і—П—Й–µ–≥–Њ –≤¬†—Б–Њ—Б—В–∞–≤ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞, –њ–Њ–Ї–∞–Ј–∞–ї, —З—В–Њ –љ–∞–Є–±–Њ–ї–µ–µ –≤–µ—А–Њ—П—В–љ—Л–є –Љ–µ—Е–∞–љ–Є–Ј–Љ –Њ—Б—Г—Й–µ—Б—В–≤–ї—П–µ—В—Б—П —З–µ—А–µ–Ј —Г–≤–µ–ї–Є—З–µ–љ–Є–µ —Б–Є–љ—В–µ–Ј–∞ –£–Ь–§ –Є¬†–і—А—Г–≥–Є—Е —Г—А–Є–і–Є–љ—Д–Њ—Б—Д–∞—В–Њ–≤. –£—А–Є–і–Є–љ—Д–Њ—Б—Д–∞—В—Л —П–≤–ї—П—О—В—Б—П —Б–Є–≥–љ–∞–ї—М–љ—Л–Љ–Є –Љ–Њ–ї–µ–Ї—Г–ї–∞–Љ–Є, –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤—Г—О—Й–Є–Љ–Є —Б¬†–њ—Г—А–Є–љ–µ—А–≥–Є—З–µ—Б–Ї–Є–Љ–Є —А–µ—Ж–µ–њ—В–Њ—А–∞–Љ–Є, –∞–Ї—В–Є–≤–∞—Ж–Є—П –Ї–Њ—В–Њ—А—Л—Е –њ—А–Є–≤–Њ–і–Є—В –Ї¬†–≤–∞–Ј–Њ–і–Є–ї–∞—В–∞—Ж–Є–Є –Є¬†–њ–Њ–Ј–Є—В–Є–≤–љ—Л–Љ –Є–Ј–Љ–µ–љ–µ–љ–Є—П–Љ –ї–Є–њ–Є–і–љ–Њ–≥–Њ –њ—А–Њ—Д–Є–ї—П.
–Р–љ–∞–ї–Є–Ј —Б—Е–Њ–ґ–µ—Б—В–Є —Е–Є–Љ–Є—З–µ—Б–Ї–Њ–є —Б—В—А—Г–Ї—В—Г—А—Л –Њ—А–Њ—В–∞—В-–∞–љ–Є–Њ–љ–∞ —Б¬†–Љ–Њ–ї–µ–Ї—Г–ї–∞–Љ–Є –Љ–µ—В–∞–±–Њ–ї–Њ–Љ–∞ —З–µ–ї–Њ–≤–µ–Ї–∞ (—Е–µ–Љ–Њ–Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Њ–љ–љ—Л–є –∞–љ–∞–ї–Є–Ј) –њ–Њ–Ј–≤–Њ–ї–Є–ї –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М —Д–∞—А–Љ–∞–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї—Г—О –њ—А–µ–і–Њ–њ—А–µ–і–µ–ї–µ–љ–љ–Њ—Б—В—М –Є–Љ–Љ—Г–љ–Њ–Љ–Њ–і—Г–ї–Є—А—Г—О—Й–µ–≥–Њ, –њ—А–Њ—В–Є–≤–Њ–≤–Њ—Б–њ–∞–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ, –љ–µ–є—А–Њ- –Є¬†–Ї–∞—А–і–Є–Њ¬≠–њ—А–Њ—В–µ–Ї—В–Є–≤–љ–Њ–≥–Њ –≤–µ–Ї—В–Њ—А–∞ –і–µ–є—Б—В–≤–Є—П –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ (—А–Є—Б.¬†10). –Ъ–∞—А–і–Є–Њ¬≠–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л–µ —Б–≤–Њ–є—Б—В–≤–∞ –Љ–Њ–≥—Г—В –Њ—Б—Г—Й–µ—Б—В–≤–ї—П—В—М—Б—П –Ј–∞ —Б—З–µ—В –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –Ї–∞—А–і–Є–Њ–њ—А–Њ—В–µ–Ї—В–Є–≤–љ—Л—Е –£–Ф–§, –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П –љ–∞¬†–∞–і–µ–љ–Њ–Ј–Є–љ–Њ–≤—Л–µ —А–µ—Ж–µ–њ—В–Њ—А—Л, —А–µ–≥—Г–ї—П—Ж–Є–Є N-–∞—Ж–µ—В–Є–ї–≥–ї—О–Ї–Њ–Ј–∞–Љ–Є–љ —В—А–∞–љ—Б—Д–µ—А–∞–Ј—Л, –Є–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є—П —Д–Њ—Б—Д–Њ–і–Є—Н—Б—В–µ—А–∞–Ј –Є¬†–Ї—Б–∞–љ—В–Є–љ –Њ–Ї—Б–Є–і–∞–Ј—Л, –∞–љ—В–Є–Њ–Ї—Б–Є–і–∞–љ—В–љ–Њ–≥–Њ —Н—Д—Д–µ–Ї—В–∞. –Я–Њ–ї—Г—З–µ–љ–љ—Л–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л —Г–Ї–∞–Ј—Л–≤–∞—О—В –љ–∞¬†–њ–µ—А—Б–њ–µ–Ї—В–Є–≤–љ—Л–µ –љ–∞–њ—А–∞–≤–ї–µ–љ–Є—П –і–∞–ї—М–љ–µ–є—И–Є—Е —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ—Л—Е –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л.
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, —Б¬†—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є —В–Њ—З–Ї–Є –Ј—А–µ–љ–Є—П —Б–Њ–≤–Љ–µ—Б—В–љ–Њ–µ –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ –Љ–∞–≥–љ–Є—П –Є¬†–Њ—А–Њ—В–∞—В–∞ –Є–Љ–µ–µ—В —П—А–Ї–Є–є —Б–Є–љ–µ—А–≥–Є–Ј–Љ –і–ї—П –Њ–њ—В–Є–Љ–Є–Ј–∞—Ж–Є–Є –Ї—А–Њ–≤–Њ—Б–љ–∞–±–ґ–µ–љ–Є—П –Ј–Њ–љ—Л –Є–љ—Д–∞—А–Ї—В–∞. –Ъ–∞–Ї –њ–Њ–Ї–∞–Ј—Л–≤–∞—О—В –і–∞–љ–љ—Л–µ —А—П–і–∞ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –њ–Њ¬†–і–Њ–Ї–∞–Ј–∞—В–µ–ї—М–љ–Њ–є –Љ–µ–і–Є—Ж–Є–љ–µ –Є¬†–љ–∞—И–∞ —Б–Њ–±—Б—В–≤–µ–љ–љ–∞—П –Ї–ї–Є–љ–Є—З–µ—Б–Ї–∞—П –њ—А–∞–Ї—В–Є–Ї–∞, –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ –≤¬†–Ї–Њ–Љ–њ–ї–µ–Ї—Б–љ–Њ–є –њ—А–Њ–≥—А–∞–Љ–Љ–µ (–љ–∞—А—П–і—Г —Б¬†–і—А—Г–≥–Є–Љ–Є –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є–Љ–Є, —Д–Є–Ј–Є—З–µ—Б–Ї–Є–Љ–Є –Є¬†–њ—Б–Є—Е–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ–Є –Љ–µ—А–∞–Љ–Є —В–µ—А–∞–њ–µ–≤—В–Є—З–µ—Б–Ї–Њ–≥–Њ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П) –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ –≤–ї–Є—П–µ—В –љ–∞¬†–≤—Л–ґ–Є–≤–∞–љ–Є–µ –Є¬†—Б–∞–Љ–Њ—З—Г–≤—Б—В–≤–Є–µ –њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–Ш–С–°.
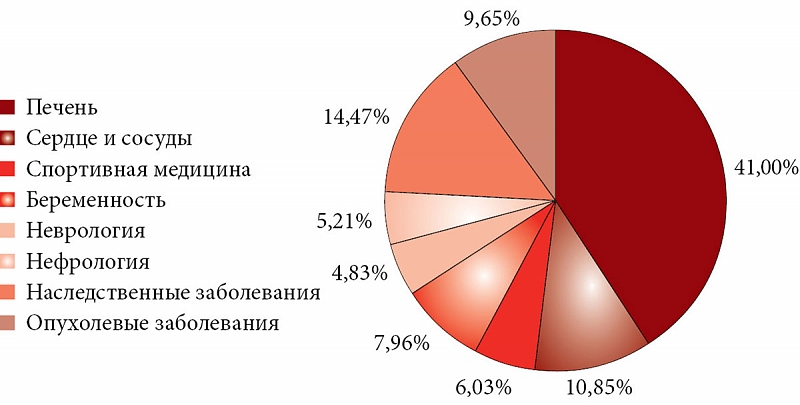
–°–Њ–≥–ї–∞—Б–љ–Њ –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–Љ—Г –∞–љ–∞–ї–Є–Ј—Г 6500 –њ—Г–±–ї–Є–Ї–∞—Ж–Є–є –њ–Њ¬†–Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В–µ –Є¬†–Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В—Г, –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–µ —З–Є—Б–ї–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Б–Ї–Њ–љ—Ж–µ–љ—В—А–Є—А–Њ–≤–∞–љ—Л –≤¬†—В–∞–Ї–Є—Е –Њ–±–ї–∞—Б—В—П—Е, –Ї–∞–Ї¬†–≥–µ–њ–∞—В–Њ–ї–Њ–≥–Є—П, —Б–µ—А–і–µ—З–љ–Њ-—Б–Њ—Б—Г–і–Є—Б—В–∞—П —Б–Є—Б—В–µ–Љ–∞, —Б–њ–Њ—А—В–Є–≤–љ–∞—П –Љ–µ–і–Є—Ж–Є–љ–∞, –љ–∞—Б–ї–µ–і—Б—В–≤–µ–љ–љ—Л–µ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П (—Б–≤—П–Ј–∞–љ–љ—Л–µ —Б¬†–љ–∞—А—Г—И–µ–љ–Є–µ–Љ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ –Њ—А–Њ—В–∞—В–∞ –Є¬†–і—А—Г–≥–Є—Е –њ–Є—А–Є–Љ–Є–і–Є–љ–Њ–≤). –Я–µ—А—Б–њ–µ–Ї—В–Є–≤–љ—Л–Љ–Є –љ–∞–њ—А–∞–≤–ї–µ–љ–Є—П–Љ–Є –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П —П–≤–ї—П—О—В—Б—П –∞–љ—В–Є–Њ–љ–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ —Н—Д—Д–µ–Ї—В—Л, –љ—Г—В—А–Є—Ж–Є–Њ–љ–љ–∞—П –њ–Њ–і–і–µ—А–ґ–Ї–∞ –Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–Њ–Љ –њ—А–Є –±–µ—А–µ–Љ–µ–љ–љ–Њ—Б—В–Є, –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ –Њ—А–Њ—В–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л –Є¬†–Љ–∞–≥–љ–Є—П –Њ—А–Њ—В–∞—В–∞ –≤¬†–ї–µ—З–µ–љ–Є–Є –љ–µ–≤—А–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є (—А–Є—Б.¬†11).
O.A. Gromova, I.Yu. Torshin, A.G. Kalacheva
Ivanovo State Medical University
Russian Satellite Centre of Trace Element вАУ Institute for Unesco
Moscow Institute of Physics and Technology (State University)
Contact person: Olga Alekseyevna Gromova, unesco.gromova@gmail.com
Orotic acid and its magnesium salt are used in a fomr of pharmacological agents belonging to group of anabolics. Magnesium orotate is widely applied in cardiology practice. Here we present the results from the latest studies pointing at a broad range of molecular mechanisms, whereby therapeutic effects of magnesium orotate are fulfilled.
–£–≤–∞–ґ–∞–µ–Љ—Л–є –њ–Њ—Б–µ—В–Є—В–µ–ї—М uMEDp!
–£–≤–µ–і–Њ–Љ–ї—П–µ–Љ –Т–∞—Б –Њ —В–Њ–Љ, —З—В–Њ –Ј–і–µ—Б—М —Б–Њ–і–µ—А–ґ–Є—В—Б—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П, –њ—А–µ–і–љ–∞–Ј–љ–∞—З–µ–љ–љ–∞—П –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –і–ї—П —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П.
–Х—Б–ї–Є –Т—Л –љ–µ —П–≤–ї—П–µ—В–µ—Б—М —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–Љ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П, –∞–і–Љ–Є–љ–Є—Б—В—А–∞—Ж–Є—П –љ–µ –љ–µ—Б–µ—В –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ—Б—В–Є –Ј–∞ –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ –њ–Њ—Б–ї–µ–і—Б—В–≤–Є—П, –≤–Њ–Ј–љ–Є–Ї—И–Є–µ –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–∞–Љ–Њ—Б—В–Њ—П—В–µ–ї—М–љ–Њ–≥–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Т–∞–Љ–Є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є —Б –њ–Њ—А—В–∞–ї–∞ –±–µ–Ј –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є –Ї–Њ–љ—Б—Г–ї—М—В–∞—Ж–Є–Є —Б –≤—А–∞—З–Њ–Љ.
–Э–∞–ґ–Є–Љ–∞—П –љ–∞ –Ї–љ–Њ–њ–Ї—Г ¬Ђ–Т–Њ–є—В–Є¬ї, –Т—Л –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В–µ, —З—В–Њ —П–≤–ї—П–µ—В–µ—Б—М –≤—А–∞—З–Њ–Љ –Є–ї–Є —Б—В—Г–і–µ–љ—В–Њ–Љ –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–≥–Њ –≤—Г–Ј–∞.