Начало исследований однополушарного сна морских млекопитающих
- Аннотация
- Статья
- Ссылки
- English
В 1961 г. опубликована книга профессора Джона Лилли «Человек и дельфин» [1], на русский язык она переведена в 1965 г. [2]. Книга вызвала огромный интерес к дельфинам во всем мире. Конечно, в ней были сформулированы различные недостаточно обоснованные предположения и целый ряд не сбывшихся в дальнейшем прогнозов. Например, Лилли утверждал, что в недалеком будущем дельфины станут первыми животными, с которыми человек начнет разговаривать, используя акустический язык. Может быть, именно некоторые фантазии автора и обусловили особый интерес к этим животным во всем мире. Тем не менее книга Лилли сыграла огромную роль в формировании современного уважительного отношения людей к дельфинам. Имела она и практические последствия. Например, в середине 1960-х гг. был прекращен промысел дельфинов всеми причерноморскими странами, кроме Турции. В 1960-е гг. сначала в США, а затем и в СССР были созданы военные дельфинарии, где делались попытки практического использования дельфинов. В Советском Союзе так же, как и за рубежом, возникло много групп ученых и инженеров, которые занялись исследованиями дельфинов в самых различных аспектах.
Наша группа физиологов сна, обсуждая особенности поведения дельфинов, не могла оставить без внимания явное противоречие: с одной стороны, сон млекопитающих, особенно парадоксальный, обязательно сопровождается их неподвижностью, а с другой – дельфины непрерывно находятся в движении. Они живут в воде, но дышат атмосферным воздухом, поэтому для каждого вдоха всплывают к поверхности воды. Дыхательные паузы у черноморских афалин, как правило, длятся всего 15 секунд. Трудно представить себе неподвижность дельфинов во время сильного волнения на море и шторма. Из данных литературы выяснилось, что речные дельфины вообще плывут непрерывно от рождения до смерти. Естественно, возник вопрос: как сочетаются у дельфинов их постоянная двигательная активность и сон, особенно парадоксальный? Пришлось даже столкнуться с мнением, что сон у дельфинов вообще отсутствует [3]. Ответ можно было получить, только зарегистрировав электрическую активность мозга (ЭЭГ) у свободноподвижных дельфинов.
Такая возможность представилась в Институте эволюционной морфологии и экологии животных им. А.Н. Северцова АН СССР, и я сменил место работы и перешел с кафедры высшей нервной деятельности биофака МГУ в этот институт в лабораторию А.Я. Супина. В 1973 г. была организована экспедиция на Черное море в район мыса Большой Утриш около г. Анапы. Были отловлены черноморские афалины для различных научных экспериментов, в том числе изучения их сна.
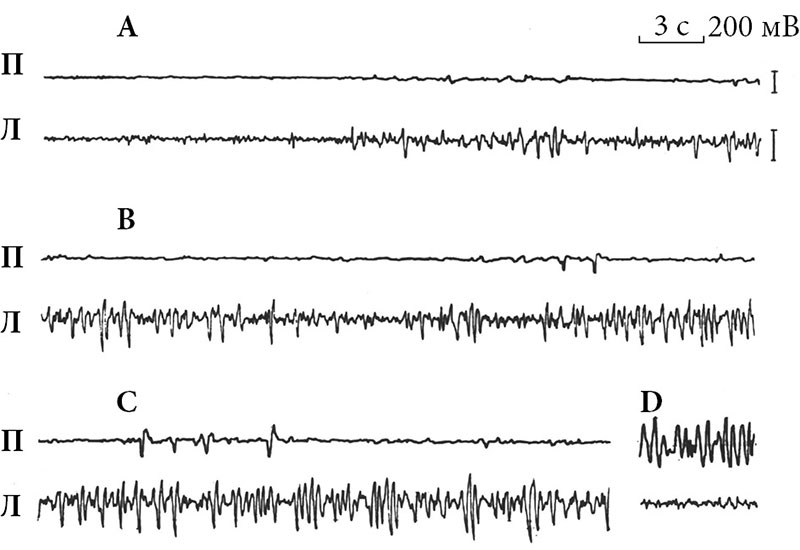
При первых же регистрациях ЭЭГ у афалин было зафиксировано обычное чередование десинхронизации и синхронизации ЭЭГ, характерное для цикла «сон –бодрствование» млекопитающих. Но при этом обнаружено однополушарное возникновение дельта-волн при десинхронизации ЭЭГ в другом полушарии, то есть межполушарная асимметрия ЭЭГ. Это явление сначала нами расценивалось как артефакт в результате некачественной регистрации электроактивности мозга, но тщательный анализ записей показал, что оно действительно существует [4]. В следующем экспедиционном сезоне 1974 г. уже все внимание было посвящено этому феномену, в результате была подтверждена его реальность и описаны его основные характеристики (рис. 1 и 2 [5]).
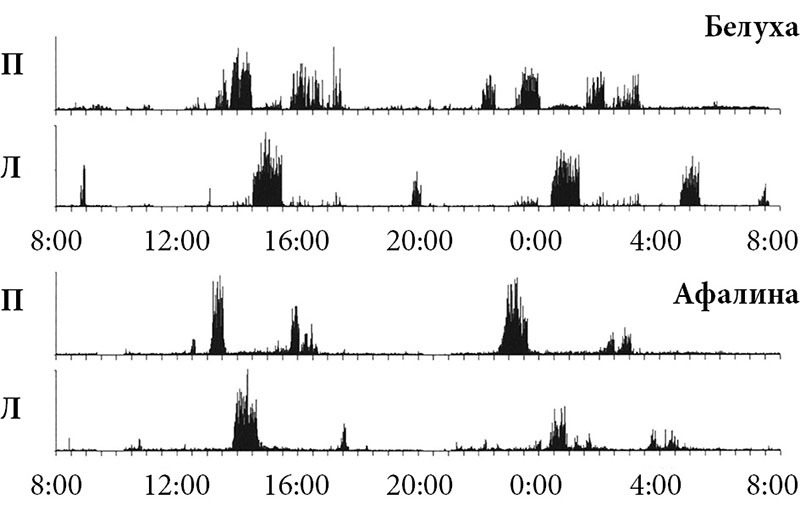
В последующих работах были определены количественные характеристики однополушарного сна дельфинов [6–9]. Подтверждена именно межполушарность этого явления, так как синхронизация возникала в различных полях одного полушария примерно одновременно [10]. Медленноволновая асимметрия наблюдалась и в подкорке – в разных ядрах таламуса синхронизация возникала тоже однополушарно и синхронно с однополушарной синхронизацией в ипсилатеральном полушарии [11]. Вживленные в кору полушарий термисторы показали, что температура коры мозга во время цикла «сон – бодрствование» меняется так же, как и у лабораторных млекопитающих, но однополушарно и синхронно с изменениями электрокортикограммы [12]. У афалин оказалось возможным осуществить депривацию сна не только билатерально, но и однополушарно. В односторонне депривируемом полушарии нарастало давление сна, а после депривации имела место отдача сна [13].
Одним из основных направлений исследований в последующие годы стало сравнительно-физиологическое. Мы пытались понять особенности сна и у других представителей водных млекопитающих: китообразных, ластоногих, сирен. И в первую очередь нас интересовало наличие однополушарного сна и у других млекопитающих. Кроме афалин, на которых впервые был описан однополушарный сон, этот феномен был четко зарегистрирован и у других дельфинов: черноморских морских свиней [14], амазонских дельфинов [15] и белухи [16]. Принципиальных различий в организации медленноволнового сна у этих видов не было.
Мы проводили электрофизиологические исследования сна у представителей всех трех семейств отряда ластоногих: настоящих тюленей (каспийских [17] и гренландских [18, 19]), ушастых тюленей (северных морских котиков [20], сивучей [21], южноафриканских морских котиков [22], южноамериканских морских львов [23]) и тихоокеанских моржей [24]. У настоящих тюленей однополушарный сон не обнаружен. Сон на суше у них такой же, как и у наземных млекопитающих. Но сон в воде у них тоже своеобразен: эпизоды сна, и медленноволнового, и парадоксального, приурочены к дыхательным паузам. Для каждого дыхательного акта, если он сопровождается движениями (всплытие, поднимание ноздрей над поверхностью воды), наступает короткое пробуждение, после которого прерванная стадия сна быстро восстанавливается. Зато у ушастых тюленей однополушарная синхронизация была обнаружена во время сна в воде, который обычно происходит на поверхности воды в позе на боку, когда одна передняя ласта движется, чтобы сохранять позу и держать ноздри над водой. На суше сон выглядит так же, как и у наземных животных, – без асимметрии ЭЭГ и без пробуждений для дыхания. У моржей сон протекает по типу сна настоящих тюленей.
В ходе зарубежных экспедиционных работ удалось также зарегистрировать электрополиграмму у одного карибского [25] и одного амазонского ламантина [26]. Сон обоих представителей отряда сирен оказался в основном схож со сном настоящих тюленей. То есть сон, и медленноволновой, и парадоксальный, развивается во время дыхательных пауз, для дыхательных актов происходит короткое пробуждение, и затем сон быстро возвращается к прерванной стадии. У амазонского ламантина мы наблюдали также четкую межполярную асимметрию медленных волн электрокортикограммы при отсутствии двигательной активности, а также отсутствии фазических компонентов парадоксального сна.
При регистрации медленноволнового сна у морских млекопитающих мы получили четкие данные: либо это однополушарный сон дельфинов и ушастых тюленей, либо сон урывками во время дыхательных пауз с обязательными короткими побудками во время дыхательных актов, если они сопровождались движениями. Но идентифицировать парадоксальный сон у дельфинов до сих пор не удалось. У тюленей настоящих и ушастых, моржей и представителей сирен парадоксальный сон четко выражен. У дельфинов поведенчески удавалось хорошо регистрировать типичные для парадоксального сна других млекопитающих вздрагивания головы, грудных плавников, мышц тела на фоне медленного стереотипного плавания. Но при параллельной регистрации ЭЭГ эти вздрагивания могли возникать и на фоне десинхронизации, и на фоне медленных волн. Важно, что вздрагивания, типичные для парадоксального сна других млекопитающих, у дельфинов возникают во время плавания, то есть двигательной активности. Можно предполагать, что парадоксальный сон у дельфинов либо отсутствует [27], либо существенным образом видоизменен, что вероятнее.
Поскольку известно, что жвачные млекопитающие продолжают движения челюстей во время сна, мы проверили электрополиграмму сна у кабарги [28] и косули [29]. Оказалось, что они действительно продолжают жевать во время медленноволнового сна, но прекращают во время парадоксального сна. Межполушарная асимметрия ЭЭГ у этих оленей не была обнаружена.
Если нейрофизиологические механизмы однополушарного сна остаются неизвестными, то в отношении функционального значения этого явления можно предполагать, что оно обеспечивает постоянную двигательную и дыхательную активность дельфинов. У обитателей морей и рек полная или почти полная неподвижность практически невозможна из-за постоянного движения водной среды (волны, течения, шторма). Относительная неподвижность наблюдается только у крупных животных и в штилевом море. Всплывать для дыхательных актов тоже надо непрерывно и с весьма короткими интервалами. Медленноволновой сон в одном полушарии при активном состоянии другого – один из возможных выходов из ситуации, который и был реализован у водных млекопитающих. В естественном сне у дельфинов никогда не наблюдались дыхательные акты на фоне билатеральных дельта-волн. Билатеральные дельта-волны можно было спровоцировать инъекцией транквилизатора диазепама, но перед дыхательным актом обязательно возникает билатеральная или однополушарная десинхронизация, и только тогда происходит дыхательный акт [14]. Есть основания предполагать, что увеличением доз диазепама можно сохранить билатеральные дельта-волны на сравнительно долгий срок, и тогда дыхание прекратится.
Таким образом, в ходе эволюции при адаптации млекопитающих к водной среде обитания природа пошла на существенное изменение физиологических механизмов сна.
Послесловие
В.М. Ковальзон, руководитель секции сомнологии Физиологического общества им. И.П. Павлова
Крупнейшим вкладом послевоенной отечественной сомнологии в мировую науку стало открытие в начале 1970-х гг. группой ученых из Института эволюционной морфологии и экологии животных им. А.Н. Северцова АН СССР (ныне – ИПЭЭ РАН) под руководством Льва Мухарамовича Мухаметова однополушарного сна у дельфинов. Дело в том, что вторичноводные млекопитающие, такие как дельфины, вынуждены постоянно всплывать на поверхность, чтобы вдохнуть, поскольку они, в отличие от рыб, дышат легкими. Регистрация ЭЭГ дельфинов показала, что мозг этих животных никогда не находится целиком в состоянии глубокого медленного сна, а полушария головного мозга спят по очереди. Если в левом полушарии наблюдается картина глубокого сна (высоковольтные медленные волны), то в правом – бодрствования (низковольтные быстрые колебания), и наоборот. Именно поэтому сон китообразных так заинтересовал Л.М. Мухаметова и его коллег. До того момента считалось, что головной мозг засыпает и спит «целиком».
Первые же результаты стадирования записей на черноморских дельфинах-афалинах показали, что асимметрия в ЭЭГ дельфина – не эпифеномен и не случайность. Нормальный сон дельфина – именно чередующийся однополушарный медленный сон [30, 31]. Л.М. Мухаметов и В.М. Ковальзон показали, что асимметрия при этом возникает в динамике не только электрической активности, но и температуры мозга [12, 32]. Никаких признаков быстрого сна у этих животных ни по ЭЭГ, ни по колебаниям температуры мозга обнаружено не было. Инструментальная депривация сна в одном полушарии вызывала «отдачу» сна только в том же самом полушарии, тем самым подтверждая, что каждое полушарие обладает собственной потребностью во сне и недостаток сна в одном полушарии не может быть скомпенсирован удлинением и/или учащением периодов глубокого сна в другом [13]. Дальнейшая 40-летняя история изучения сна дельфинов двумя поколениями исследователей с применением новейших методик показала, что такая организация естественного сна характерна, видимо, для всех видов зубатых китов, и выявила множество интересных деталей, однако ничего принципиально нового в первоначальную картину, обрисованную с помощью самой примитивной экспериментальной техники, не внесла [5].
Таким образом, в 1970–80-х гг. работы ученых из группы Л.М. Мухаметова привели к очередному пересмотру представлений о механизмах сна. В дальнейшем группа Л.М. Мухаметова и О.И. Лямина показала, что различные виды глубокой межполушарной функциональной асимметрии медленного сна довольно широко распространены среди многих групп водных млекопитающих, и не только зубатых китов, но и ушастых тюленей, моржей, дюгоней.
В 1997 г. Иван Николаевич Пигарев, сотрудник Института проблем передачи информации им. А.А. Харкевича РАН, работавший тогда в Германии, опубликовал в одном международном журнале небольшую статью (совместно с немецким и американским коллегами) о том, что в опытах на обезьянах были обнаружены периоды асинхронного развития медленного сна в разных отделах коры одного и того же полушария [33]. Длительное выполнение монотонной двигательной задачи приводило к «засыпанию» зрительной коры, что отмечалось по активности нейронов, в то время как рука обезьяны продолжала выполнять движения, демонстрируя активность моторной коры. В дальнейшем сходные результаты были получены и другими группами в опытах на крысах и привели к формированию понятия так называемого локального сна у наземных млекопитающих, что значительно обогатило представления о механизмах сна. По мнению некоторых авторов, однополушарный медленный сон дельфинов – некий «предельный вариант» локального сна, распространившегося на целое полушарие, характерного также и для наземных млекопитающих и некоторых видов птиц [34, 35]. Однако аналогичны (схожи по внешним проявлениям) эти феномены или гомологичны (схожи по механизмам) – остается неизвестным и является предметом текущих исследований.
L.M. Mukhametov
A.N. Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences, Moscow
Contact person: Lev Mukharamovich Mukhametov, lev.mukhametov@gmail.com
Narration on the discovery of hemisphere sleep in dolphins by the Institute of Evolutionary Morphology and Ecology of Animals research group in the early 1970s, which led to the revision of the knowledge on mechanisms of sleep.
Уважаемый посетитель uMEDp!
Уведомляем Вас о том, что здесь содержится информация, предназначенная исключительно для специалистов здравоохранения.
Если Вы не являетесь специалистом здравоохранения, администрация не несет ответственности за возможные отрицательные последствия, возникшие в результате самостоятельного использования Вами информации с портала без предварительной консультации с врачом.
Нажимая на кнопку «Войти», Вы подтверждаете, что являетесь врачом или студентом медицинского вуза.