–Ь–Њ–і—Г–ї—П—Ж–Є—П –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Љ–µ—В—Д–Њ—А–Љ–Є–љ–Њ–Љ
- –Р–љ–љ–Њ—В–∞—Ж–Є—П
- –°—В–∞—В—М—П
- –°—Б—Л–ї–Ї–Є
- English
–Т —Б—В–∞—В—М–µ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ –Њ–±–Ј–Њ—А —Б–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–є –ї–Є—В–µ—А–∞—В—Г—А—Л –Њ –≤–ї–Є—П–љ–Є–Є –њ–µ—А–Њ—А–∞–ї—М–љ–Њ–є —Б–∞—Е–∞—А–Њ—Б–љ–Є–ґ–∞—О—Й–µ–є —В–µ—А–∞–њ–Є–Є, –≤ —З–∞—Б—В–љ–Њ—Б—В–Є –Љ–µ—В—Д–Њ—А–Љ–Є–љ–∞, –љ–∞ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Г –Ї–Є—И–µ—З–љ–Є–Ї–∞. –Ь–µ—В—Д–Њ—А–Љ–Є–љ —П–≤–ї—П–µ—В—Б—П –њ—А–µ–њ–∞—А–∞—В–Њ–Љ –њ–µ—А–≤–Њ–є –ї–Є–љ–Є–Є –Є –љ–∞–Є–±–Њ–ї–µ–µ —И–Є—А–Њ–Ї–Њ –Є—Б–њ–Њ–ї—М–Ј—Г–µ—В—Б—П –і–ї—П –ї–µ—З–µ–љ–Є—П –њ—А–µ–і–Є–∞–±–µ—В–∞ –Є —Б–∞—Е–∞—А–љ–Њ–≥–Њ –і–Є–∞–±–µ—В–∞ 2 —В–Є–њ–∞. –Ю–љ –≤—Л–Ј—Л–≤–∞–µ—В —И–Є—А–Њ–Ї–Є–є —Б–њ–µ–Ї—В—А —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –љ–∞ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ, –Њ–і–љ–∞–Ї–Њ –Љ–µ—Е–∞–љ–Є–Ј–Љ –µ–≥–Њ –і–µ–є—Б—В–≤–Є—П –і–Њ —Б–Є—Е –њ–Њ—А –Є–Ј—Г—З–µ–љ –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ. –Я–Њ—Б–ї–µ–і–љ–Є–µ –і–∞–љ–љ—Л–µ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О—В –Њ –Љ–µ—В—Д–Њ—А–Љ–Є–љ-–Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–љ–љ–Њ–Љ –Є–Ј–Љ–µ–љ–µ–љ–Є–Є –Ї–∞—З–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Ї–Є—И–µ—З–љ–Є–Ї–∞, –≤ —З–∞—Б—В–љ–Њ—Б—В–Є –Њ –њ–Њ–≤—Л—И–µ–љ–Є–Є –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–љ–Њ—Б—В–Є –±–∞–Ї—В–µ—А–Є–є —А–Њ–і–∞ Akkermansia spp. –Є —Г–Љ–µ–љ—М—И–µ–љ–Є–Є —Г—Б–ї–Њ–≤–љ–Њ –њ–∞—В–Њ–≥–µ–љ–љ–Њ–є —Д–ї–Њ—А—Л. –Я—А–µ–њ–∞—А–∞—В —В–∞–Ї–ґ–µ –≤–ї–Є—П–µ—В –љ–∞ —Б–Є–љ—В–µ–Ј –≥–ї—О–Ї–∞–≥–Њ–љ–Њ–њ–Њ–і–Њ–±–љ–Њ–≥–Њ –њ–µ–њ—В–Є–і–∞ 1. –Э–µ –Є—Б–Ї–ї—О—З–µ–љ–Њ, —З—В–Њ —Г–Ї–∞–Ј–∞–љ–љ—Л–є —Н—Д—Д–µ–Ї—В —А–µ–∞–ї–Є–Ј—Г–µ—В—Б—П —З–µ—А–µ–Ј –Љ–Њ–і—Г–ї—П—Ж–Є—О –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Ї–Є—И–µ—З–љ–Є–Ї–∞ –Є —Б–Є–љ—В–µ–Ј –±–∞–Ї—В–µ—А–Є—П–Љ–Є –Ї–Њ—А–Њ—В–Ї–Њ—Ж–µ–њ–Њ—З–µ—З–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В. –Я–Њ—Н—В–Њ–Љ—Г –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В—Б—П –∞–Ї—В—Г–∞–ї—М–љ—Л–Љ –Є–Ј—Г—З–µ–љ–Є–µ –≤–ї–Є—П–љ–Є—П –∞–≥–Њ–љ–Є—Б—В–Њ–≤ —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –≥–ї—О–Ї–∞–≥–Њ–љ–Њ–њ–Њ–і–Њ–±–љ–Њ–≥–Њ –њ–µ–њ—В–Є–і–∞ 1 –љ–∞ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Г –Ї–Є—И–µ—З–љ–Є–Ї–∞ –љ–µ —В–Њ–ї—М–Ї–Њ —Б —Ж–µ–ї—М—О –Ї–Њ—А—А–µ–Ї—Ж–Є–Є –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –љ–∞—А—Г—И–µ–љ–Є–є, –љ–Њ –Є –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ–≥–Њ —Г–ї—Г—З—И–µ–љ–Є—П –µ–µ –Ї–∞—З–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞.
–Т —Б—В–∞—В—М–µ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ –Њ–±–Ј–Њ—А —Б–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–є –ї–Є—В–µ—А–∞—В—Г—А—Л –Њ –≤–ї–Є—П–љ–Є–Є –њ–µ—А–Њ—А–∞–ї—М–љ–Њ–є —Б–∞—Е–∞—А–Њ—Б–љ–Є–ґ–∞—О—Й–µ–є —В–µ—А–∞–њ–Є–Є, –≤ —З–∞—Б—В–љ–Њ—Б—В–Є –Љ–µ—В—Д–Њ—А–Љ–Є–љ–∞, –љ–∞ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Г –Ї–Є—И–µ—З–љ–Є–Ї–∞. –Ь–µ—В—Д–Њ—А–Љ–Є–љ —П–≤–ї—П–µ—В—Б—П –њ—А–µ–њ–∞—А–∞—В–Њ–Љ –њ–µ—А–≤–Њ–є –ї–Є–љ–Є–Є –Є –љ–∞–Є–±–Њ–ї–µ–µ —И–Є—А–Њ–Ї–Њ –Є—Б–њ–Њ–ї—М–Ј—Г–µ—В—Б—П –і–ї—П –ї–µ—З–µ–љ–Є—П –њ—А–µ–і–Є–∞–±–µ—В–∞ –Є —Б–∞—Е–∞—А–љ–Њ–≥–Њ –і–Є–∞–±–µ—В–∞ 2 —В–Є–њ–∞. –Ю–љ –≤—Л–Ј—Л–≤–∞–µ—В —И–Є—А–Њ–Ї–Є–є —Б–њ–µ–Ї—В—А —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Н—Д—Д–µ–Ї—В–Њ–≤ –љ–∞ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ, –Њ–і–љ–∞–Ї–Њ –Љ–µ—Е–∞–љ–Є–Ј–Љ –µ–≥–Њ –і–µ–є—Б—В–≤–Є—П –і–Њ —Б–Є—Е –њ–Њ—А –Є–Ј—Г—З–µ–љ –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ. –Я–Њ—Б–ї–µ–і–љ–Є–µ –і–∞–љ–љ—Л–µ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О—В –Њ –Љ–µ—В—Д–Њ—А–Љ–Є–љ-–Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–љ–љ–Њ–Љ –Є–Ј–Љ–µ–љ–µ–љ–Є–Є –Ї–∞—З–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Ї–Є—И–µ—З–љ–Є–Ї–∞, –≤ —З–∞—Б—В–љ–Њ—Б—В–Є –Њ –њ–Њ–≤—Л—И–µ–љ–Є–Є –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–љ–Њ—Б—В–Є –±–∞–Ї—В–µ—А–Є–є —А–Њ–і–∞ Akkermansia spp. –Є —Г–Љ–µ–љ—М—И–µ–љ–Є–Є —Г—Б–ї–Њ–≤–љ–Њ –њ–∞—В–Њ–≥–µ–љ–љ–Њ–є —Д–ї–Њ—А—Л. –Я—А–µ–њ–∞—А–∞—В —В–∞–Ї–ґ–µ –≤–ї–Є—П–µ—В –љ–∞ —Б–Є–љ—В–µ–Ј –≥–ї—О–Ї–∞–≥–Њ–љ–Њ–њ–Њ–і–Њ–±–љ–Њ–≥–Њ –њ–µ–њ—В–Є–і–∞ 1. –Э–µ –Є—Б–Ї–ї—О—З–µ–љ–Њ, —З—В–Њ —Г–Ї–∞–Ј–∞–љ–љ—Л–є —Н—Д—Д–µ–Ї—В —А–µ–∞–ї–Є–Ј—Г–µ—В—Б—П —З–µ—А–µ–Ј –Љ–Њ–і—Г–ї—П—Ж–Є—О –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Ї–Є—И–µ—З–љ–Є–Ї–∞ –Є —Б–Є–љ—В–µ–Ј –±–∞–Ї—В–µ—А–Є—П–Љ–Є –Ї–Њ—А–Њ—В–Ї–Њ—Ж–µ–њ–Њ—З–µ—З–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В. –Я–Њ—Н—В–Њ–Љ—Г –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В—Б—П –∞–Ї—В—Г–∞–ї—М–љ—Л–Љ –Є–Ј—Г—З–µ–љ–Є–µ –≤–ї–Є—П–љ–Є—П –∞–≥–Њ–љ–Є—Б—В–Њ–≤ —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –≥–ї—О–Ї–∞–≥–Њ–љ–Њ–њ–Њ–і–Њ–±–љ–Њ–≥–Њ –њ–µ–њ—В–Є–і–∞ 1 –љ–∞ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Г –Ї–Є—И–µ—З–љ–Є–Ї–∞ –љ–µ —В–Њ–ї—М–Ї–Њ —Б —Ж–µ–ї—М—О –Ї–Њ—А—А–µ–Ї—Ж–Є–Є –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –љ–∞—А—Г—И–µ–љ–Є–є, –љ–Њ –Є –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ–≥–Њ —Г–ї—Г—З—И–µ–љ–Є—П –µ–µ –Ї–∞—З–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞.

–Т–≤–µ–і–µ–љ–Є–µ
–£—З–µ–љ–Є–µ –Њ¬†–Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–∞—Е, –љ–∞—Б–µ–ї—П—О—Й–Є—Е —З–µ–ї–Њ–≤–µ—З–µ—Б–Ї–Є–є –Њ—А–≥–∞–љ–Є–Ј–Љ, –љ–∞—Б—З–Є—В—Л–≤–∞–µ—В –љ–µ¬†–Њ–і–љ–Њ —Б—В–Њ–ї–µ—В–Є–µ. –Э–∞—З–∞–ї–Њ —Н–≤—А–Є—Б—В–Є—З–µ—Б–Ї–Њ–≥–Њ —Н—В–∞–њ–∞ –∞—Б—Б–Њ—Ж–Є–Є—А—Г–µ—В—Б—П —Б¬†–Р. –≤–∞–љ –Ы–µ–≤–µ–љ–≥—Г–Ї–Њ–Љ (XVIII –≤.), –Ї–Њ—В–Њ—А—Л–є –Њ–±–љ–∞—А—Г–ґ–Є–ї –≤¬†–Њ—А–≥–∞–љ–Є–Ј–Љ–µ —З–µ–ї–Њ–≤–µ–Ї–∞ –Є¬†–ґ–Є–≤–Њ—В–љ—Л—Е –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ—Л. –°–ї–µ–і—Г—О—Й–Є–є —Н—В–∞–њ –Љ–Њ–ґ–љ–Њ –Њ—Е–∞—А–∞–Ї—В–µ—А–Є–Ј–Њ–≤–∞—В—М –Ї–∞–Ї –љ–∞–Ї–Њ–њ–Є—В–µ–ї—М–љ—Л–є. –Ю–≥—А–Њ–Љ–љ—Л–є –≤–Ї–ї–∞–і –≤–љ–µ—Б–ї–Є –Ш.–Ш.¬†–Ь–µ—З–љ–Є–Ї–Њ–≤, –Ы.–У. –Я–µ—А–µ—Ж, –Ы.¬†–Я–∞—Б—В–µ—А, –†.¬†–Ъ–Њ—Е, –Э.–§. –У–∞–Љ–∞–ї–µ–є –Є¬†–і—А. –Т¬†—Н—В–Њ—В –њ–µ—А–Є–Њ–і –±—Л–ї–Є –Њ–±–љ–∞—А—Г–ґ–µ–љ—Л –Є¬†–Є–і–µ–љ—В–Є—Д–Є—Ж–Є—А–Њ–≤–∞–љ—Л –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ—Л –≤¬†—А–∞–Ј–љ—Л—Е –Њ—А–≥–∞–љ–∞—Е —З–µ–ї–Њ–≤–µ–Ї–∞, –Є–Ј—Г—З–µ–љ—Л –Є—Е —Б–≤–Њ–є—Б—В–≤–∞ –Є¬†—А–Њ–ї—М. 30вАУ90-–µ¬†–≥–≥. –њ—А–Њ—И–ї–Њ–≥–Њ —Б—В–Њ–ї–µ—В–Є—П¬†вАУ —Н—В–∞–њ –і–µ—В–∞–ї–Є–Ј–∞—Ж–Є–Є. –С–ї–∞–≥–Њ–і–∞—А—П —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л–Љ –Љ–Є–Ї—А–Њ–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–Љ –Љ–µ—В–Њ–і–∞–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –Є–Ј—Г—З–µ–љ–∞ –љ–µ¬†—В–Њ–ї—М–Ї–Њ —А–Њ–ї—М –Њ—В–і–µ–ї—М–љ—Л—Е –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–µ–є –љ–Њ—А–Љ–∞–ї—М–љ–Њ–є –Є¬†–њ–∞—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є –Љ–Є–Ї—А–Њ—Д–ї–Њ—А—Л, –љ–Њ¬†–Є –Њ–±—Г—Б–ї–Њ–≤–ї–µ–љ–љ—Л–µ –Є–Љ–Є –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –љ–∞—А—Г—И–µ–љ–Є—П¬†–≥–Њ–Љ–µ–Њ—Б—В–∞–Ј–∞.
–Т¬†–Ї–Њ–љ—Ж–µ –•–•¬†вАУ –љ–∞—З–∞–ї–µ XXI –≤. —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–Њ—Б—М –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є–µ –Њ¬†–Љ–Є–Ї—А–Њ—Д–ї–Њ—А–µ –Ї–∞–Ї –µ—Й–µ –Њ–±¬†–Њ–і–љ–Њ–Љ –Њ—А–≥–∞–љ–µ —З–µ–ї–Њ–≤–µ—З–µ—Б–Ї–Њ–≥–Њ –Њ—А–≥–∞–љ–Є–Ј–Љ–∞.
–Ґ–µ—А–Љ–Є–љ ¬Ђ–Љ–Є–Ї—А–Њ–±–Є–Њ—В–∞¬ї –±—Л–ї –≤–≤–µ–і–µ–љ –Ф–ґ. –Ы–µ–і–µ—А–±–µ—А–≥–Њ–Љ [1].
–Т¬†–љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П –њ–Њ–і –Љ–Є–Ї—А–Њ–±–Є–Њ—В–Њ–є –њ–Њ–љ–Є–Љ–∞—О—В —Б–Њ–≤–Њ–Ї—Г–њ–љ–Њ—Б—В—М –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤, –Є—Е¬†–≥–µ–љ–Њ–≤ –Є¬†–≤–Ј–∞–Є–Љ–Њ–Њ—В–љ–Њ—И–µ–љ–Є–є –≤–љ—Г—В—А–Є –Њ–њ—А–µ–і–µ–ї–µ–љ–љ–Њ–є —Б—А–µ–і—Л —Б¬†–∞–Ї—Ж–µ–љ—В–Њ–Љ –љ–∞ –Є—Е¬†—В–∞–Ї—Б–Њ–љ–Њ–Љ–Є—З–µ—Б–Ї–Њ–Љ —Б–Њ—Б—В–∞–≤–µ.
–Э–∞–Є–±–Њ–ї—М—И–µ–є –њ–ї–Њ—В–љ–Њ—Б—В—М—О –Є¬†—Б–Њ–≤–Њ–Ї—Г–њ–љ–Њ–є –±–Є–Њ–Љ–∞—Б—Б–Њ–є, –∞¬†—В–∞–Ї–ґ–µ –Ј–љ–∞—З–µ–љ–Є–µ–Љ –і–ї—П —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є–Є –Њ–±–ї–∞–і–∞–µ—В –Љ–Є–Ї—А–Њ–±–Є–Њ—В–∞ –Ї–Є—И–µ—З–љ–Є–Ї–∞ (–Ь–Ъ). –Х–µ –≤–µ—Б –і–Њ—Б—В–Є–≥–∞–µ—В 3вАУ5 –Ї–≥.
–Т¬†–њ—А–Њ—Б–≤–µ—В–µ –ґ–µ–ї—Г–і–Њ—З–љ–Њ-–Ї–Є—И–µ—З–љ–Њ–≥–Њ —В—А–∞–Ї—В–∞ —Г¬†–Ј–і–Њ—А–Њ–≤—Л—Е –ї–Є—Ж –Њ–±–Є—В–∞–µ—В –±–Њ–ї–µ–µ 1014¬†–±–∞–Ї—В–µ—А–Є–∞–ї—М–љ—Л—Е –Ї–ї–µ—В–Њ–Ї, —З—В–Њ –љ–∞¬†–њ–Њ—А—П–і–Њ–Ї –≤—Л—И–µ –Њ–±—Й–µ–≥–Њ —З–Є—Б–ї–∞ –Ї–ї–µ—В–Њ–Ї –Њ—А–≥–∞–љ–Є–Ј–Љ–∞ [2]. –°–Њ–і–µ—А–ґ–∞–љ–Є–µ —Б–Є–Љ–±–Є–Њ—В–Є—З–µ—Б–Ї–Є—Е –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤ –≤¬†–ґ–µ–ї—Г–і–Њ—З–љ–Њ-–Ї–Є—И–µ—З–љ–Њ–Љ —В—А–∞–Ї—В–µ —И–Є—А–Њ–Ї–Њ –≤–∞—А—М–Є—А—Г–µ—В—Б—П¬†вАУ –Њ—В¬†102вАУ103¬†–Ъ–Ю–Х/–≥¬†–≤ –ґ–µ–ї—Г–і–Ї–µ –і–Њ 1012вАУ1013¬†–Ъ–Ю–Х/–≥ –≤ –і–Є—Б—В–∞–ї—М–љ—Л—Е –Њ—В–і–µ–ї–∞—Е —В–Њ–ї—Б—В–Њ–є –Ї–Є—И–Ї–Є.
–Ф–ї—П –Є–і–µ–љ—В–Є—Д–Є–Ї–∞—Ж–Є–Є –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤ –Ї–∞–Ї –≤¬†–љ–∞—Г—З–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П—Е, —В–∞–Ї –Є¬†–≤ —А—Г—В–Є–љ–љ–Њ–є –Ї–ї–Є–љ–Є–Ї–Њ-–ї–∞–±–Њ—А–∞—В–Њ—А–љ–Њ–є –±–∞–Ї—В–µ—А–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є –њ—А–∞–Ї—В–Є–Ї–µ –Є—Б–њ–Њ–ї—М–Ј—Г–µ—В—Б—П¬†–≥–µ–љ 16S —А–†–Э–Ъ. –Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ—Б–ї–µ–і–љ–µ–≥–Њ –і–µ—Б—П—В–Є–ї–µ—В–Є—П, –њ—А–Њ–≤–µ–і–µ–љ–љ—Л–µ —Б¬†–њ–Њ–Љ–Њ—Й—М—О –Љ–µ—В–∞–≥–µ–љ–Њ–Љ–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞, –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ —Г¬†–Ј–і–Њ—А–Њ–≤—Л—Е –≤–Ј—А–Њ—Б–ї—Л—Е –≤¬†–Ь–Ъ –і–Њ–Љ–Є–љ–Є—А—Г—О—В –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї–Є –њ—П—В–Є —Д–Є–ї—Г–Љ–Њ–≤ (–Њ—В–і–µ–ї–Њ–≤) –±–∞–Ї—В–µ—А–Є–є:¬†–≥—А–∞–Љ–њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ—Л–µ Firmicutes, Actinobacteria –Є¬†–≥—А–∞–Љ–Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ Bacteroidetes, Proteobacteria –Є¬†Verrucomicrobia. –Ф–Њ 90% –њ—А–Є—Е–Њ–і–Є—В—Б—П –љ–∞¬†Firmicutes –Є¬†Bacteroidetes [3]. –Т¬†–Ј–і–Њ—А–Њ–≤–Њ–є –Ь–Ъ Actinobacteria, Proteobacteria –Є¬†Verrucomicrobia, –Ї–∞–Ї –њ—А–∞–≤–Є–ї–Њ, –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л –≤¬†–Љ–µ–љ—М—И–µ–Љ –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ. –Ю–і–љ–∞–Ї–Њ –Є–Љ–µ–љ–љ–Њ —Н—В–Є –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ—Л –Њ–Ї–∞–Ј—Л–≤–∞—О—В –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –љ–∞¬†—Б–Њ—Б—В–Њ—П–љ–Є–µ –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–∞.
–Ь–Є–Ї—А–Њ–±–Є–Њ—В–∞ –Њ—А–≥–∞–љ–Є–Ј–Љ–∞ –≤¬†—Ж–µ–ї–Њ–Љ –Є¬†–Ї–Є—И–µ—З–љ–Є–Ї–∞ –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є —П–≤–ї—П–µ—В—Б—П —Б–≤–Њ–µ–Њ–±—А–∞–Ј–љ—Л–Љ –Є–љ–і–Є–Ї–∞—В–Њ—А–Њ–Љ –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–∞¬†вАУ –≤¬†–Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В¬†—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е, –і–Є–µ—В–Є—З–µ—Б–Ї–Є—Е, –Ї–ї–Є–Љ–∞—В–Є—З–µ—Б–Ї–Є—Е –Є¬†–≥–µ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤ –Є–Ј–Љ–µ–љ—П–µ—В—Б—П –µ–µ –Ї–∞—З–µ—Б—В–≤–µ–љ–љ—Л–є –Є¬†–Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ—Л–є —Б–Њ—Б—В–∞–≤.
–†–µ–Ј—Г–ї—М—В–∞—В—Л –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –њ—А–Њ–і–µ–Љ–Њ–љ—Б—В—А–Є—А–Њ–≤–∞–ї–Є —Б–≤—П–Ј—М –Љ–µ–ґ–і—Г –Ї–∞—З–µ—Б—В–≤–µ–љ–љ—Л–Љ–Є –Є¬†–Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ—Л–Љ–Є –Є–Ј–Љ–µ–љ–µ–љ–Є—П–Љ–Є –Ь–Ъ –Є¬†—Б–Є–љ–і—А–Њ–Љ–∞–Љ–Є, –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ–љ—Л–Љ–Є —Б¬†–љ–µ–њ—А–∞–≤–Є–ї—М–љ—Л–Љ –њ–Є—В–∞–љ–Є–µ–Љ. –†–µ—З—М, –≤¬†—З–∞—Б—В–љ–Њ—Б—В–Є, –Є–і–µ—В –Њ–±¬†–Њ–ґ–Є—А–µ–љ–Є–Є, –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Њ–Љ —Б–Є–љ–і—А–Њ–Љ–µ, —Б–∞—Е–∞—А–љ–Њ–Љ –і–Є–∞–±–µ—В–µ (–°–Ф) 2 —В–Є–њ–∞, –∞—В–µ—А–Њ—Б–Ї–ї–µ—А–Њ–Ј–µ [2, 4вАУ6]. –Я—А–Є –Њ–ґ–Є—А–µ–љ–Є–Є –Є¬†–°–Ф¬†2 —В–Є–њ–∞ –Њ—В–Љ–µ—З–µ–љ–Њ —Г–Љ–µ–љ—М—И–µ–љ–Є–µ —З–Є—Б–ї–µ–љ–љ–Њ—Б—В–Є Bacteroidetes –Є¬†—Г–≤–µ–ї–Є—З–µ–љ–Є–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–∞ Firmicutes [7, 8].
–Я—А–Є—Е–Њ–і–Є—В—Б—П –Ї–Њ–љ—Б—В–∞—В–Є—А–Њ–≤–∞—В—М, —З—В–Њ –љ–∞¬†—Б–µ–≥–Њ–і–љ—П—И–љ–Є–є –і–µ–љ—М –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ —Е–Њ—А–Њ—И–Њ –Є–Ј—Г—З–µ–љ—Л –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –Љ–Њ—А—Д–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Є¬†—Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ—Л—Е –Є–Ј–Љ–µ–љ–µ–љ–Є–є –њ—А–Є –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Њ–Љ —Б–Є–љ–і—А–Њ–Љ–µ, –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ¬†вАУ –≤–Ї–ї–∞–і –≤¬†–µ–≥–Њ —Н—В–Є–Њ–ї–Њ–≥–Є—О –Є¬†–њ–∞—В–Њ–≥–µ–љ–µ–Ј –Ь–Ъ. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –љ–µ—В –µ–і–Є–љ–Њ–≥–Њ –Љ–љ–µ–љ–Є—П –Њ¬†—В–Њ–Љ, —З—В–Њ –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В —Б–Њ–±–Њ–є ¬Ђ—В–Є–њ–Є—З–љ–∞—П¬ї –Љ–Є–Ї—А–Њ–±–Є–Њ—В–∞ –њ—А–Є –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Њ–Љ —Б–Є–љ–і—А–Њ–Љ–µ.
–°–Ї–∞–Ј–∞–љ–љ–Њ–µ –≤—Л—И–µ –Њ–±—Г—Б–ї–Њ–≤–ї–Є–≤–∞–µ—В –∞–Ї—В—Г–∞–ї—М–љ–Њ—Б—В—М –Є–Ј—Г—З–µ–љ–Є—П –њ—А–µ–њ–∞—А–∞—В–Њ–≤, –љ–∞–њ—А–∞–≤–ї–µ–љ–љ—Л—Е –љ–∞¬†–Ї–Њ—А—А–µ–Ї—Ж–Є—О –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –љ–∞—А—Г—И–µ–љ–Є–є, –љ–∞¬†–њ—А–µ–і–Љ–µ—В –Є—Е –≤–Њ–Ј–і–µ–є—Б—В–≤–Є—П —З–µ—А–µ–Ј –Љ–Њ–і—Г–ї—П—Ж–Є—О –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л, —З—В–Њ –Љ–Њ–ґ–µ—В –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ –њ–Њ–≤–ї–Є—П—В—М –Є¬†–љ–∞ –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ.
–Т —Б—В–∞—В—М–µ –±—Г–і—Г—В –њ–Њ–і—А–Њ–±–љ–Њ —А–∞—Б—Б–Љ–Њ—В—А–µ–љ—Л —Н—Д—Д–µ–Ї—В—Л –Љ–µ—В—Д–Њ—А–Љ–Є–љ–∞.
–Т–ї–Є—П–љ–Є–µ –љ–∞¬†—З–Є—Б–ї–µ–љ–љ–Њ—Б—В—М –Є¬†—Б–Њ—Б—В–∞–≤ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Ї–Є—И–µ—З–љ–Є–Ї–∞
–Ь–µ—В—Д–Њ—А–Љ–Є–љ (–Ь–Х–Ґ) –≤—Л–Ј—Л–≤–∞–µ—В —И–Є—А–Њ–Ї–Є–є —Б–њ–µ–Ї—В—А —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Н—Д—Д–µ–Ї—В–Њ–≤, –Њ–і–љ–∞–Ї–Њ –Љ–µ—Е–∞–љ–Є–Ј–Љ –µ–≥–Њ –і–µ–є—Б—В–≤–Є—П –і–Њ —Б–Є—Е –њ–Њ—А –Є–Ј—Г—З–µ–љ –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ.
–£¬†—З–µ–ї–Њ–≤–µ–Ї–∞ –Є¬†–ґ–Є–≤–Њ—В–љ—Л—Е –Ь–Х–Ґ –∞–±—Б–Њ—А–±–Є—А—Г–µ—В—Б—П –Є¬†–љ–∞–Ї–∞–њ–ї–Є–≤–∞–µ—В—Б—П –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –≤¬†–Ї–Є—И–µ—З–љ–Є–Ї–µ. –£—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ –µ–≥–Њ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ –≤¬†—В–Њ–љ–Ї–Њ–є –Ї–Є—И–Ї–µ –≤–Њ –Љ–љ–Њ–≥–Њ —А–∞–Ј –њ—А–µ–≤—Л—И–∞–µ—В —В–∞–Ї–Њ–≤–Њ–µ –≤¬†–њ–ї–∞–Ј–Љ–µ –Є¬†–і—А—Г–≥–Є—Е —В–Ї–∞–љ—П—Е [9]. –Ш–Љ–µ–љ–љ–Њ —Н—В–Є–Љ –Њ–±—К—П—Б–љ—П–µ—В—Б—П —А–∞–Ј–≤–Є—В–Є–µ –њ–Њ–±–Њ—З–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ–≤ —Б–Њ¬†—Б—В–Њ—А–Њ–љ—Л –ґ–µ–ї—Г–і–Њ—З–љ–Њ-–Ї–Є—И–µ—З–љ–Њ–≥–Њ —В—А–∞–Ї—В–∞ –њ—А–Є –њ—А–Є–µ–Љ–µ –њ—А–µ–њ–∞—А–∞—В–∞. –Т–љ—Г—В—А–Є–≤–µ–љ–љ–Њ–µ –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ –Ь–Х–Ґ –≤¬†–Њ—В–ї–Є—З–Є–µ –Њ—В¬†–њ–µ—А–Њ—А–∞–ї—М–љ–Њ–≥–Њ –љ–µ¬†—Г–ї—Г—З—И–∞–µ—В¬†–≥–ї–Є–Ї–µ–Љ–Є—О [10].
–Ь–µ—В—Д–Њ—А–Љ–Є–љ –≤–ї–Є—П–µ—В –љ–∞¬†–∞–Ї—В–Є–≤–љ–Њ—Б—В—М¬†–≥–µ–љ–Њ–≤, —А–µ–≥—Г–ї–Є—А—Г—О—Й–Є—Е –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ –Ї—Б–µ–љ–Њ–±–Є–Њ—В–Є–Ї–Њ–≤, –Ї–ї–µ—В–Њ—З–љ—Л–є —Б—В—А–µ—Б—Б, —Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–Є–є –Њ–±–Љ–µ–љ, –±–Є–Њ—Б–Є–љ—В–µ–Ј, –њ–µ—А–µ–і–∞—З—Г —Б–Є–≥–љ–∞–ї–Њ–≤ –Є¬†—В.–і. –Ш—Е¬†—Н–Ї—Б–њ—А–µ—Б—Б–Є—П –Љ–µ–љ—П–µ—В—Б—П —В–∞–Ї–ґ–µ –њ—А–Є —Б–љ–Є–ґ–µ–љ–Є–Є —Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–Њ–є —Ж–µ–љ–љ–Њ—Б—В–Є –њ–Є—Й–Є [11]. –£¬†–љ–µ–Љ–∞—В–Њ–і Caenorhabditis elegans –Ь–Х–Ґ –≤–Њ–Ј–і–µ–є—Б—В–≤—Г–µ—В –љ–∞¬†–Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ –Љ–µ—В–Є–Њ–љ–Є–љ–∞ –Є¬†—Д–Њ–ї–∞—В–∞ (–Ї–∞–Ї –∞–љ—В–Є—Д–Њ–ї–∞—В–љ—Л–є –њ—А–µ–њ–∞—А–∞—В) –±–∞–Ї—В–µ—А–Є–є-—Б–Є–Љ–±–Є–Њ–љ—В–Њ–≤ –Є¬†–Ј–∞–Љ–µ–і–ї—П–µ—В –њ—А–Њ—Ж–µ—Б—Б —Б—В–∞—А–µ–љ–Є—П –≤¬†–њ—А–Є—Б—Г—В—Б—В–≤–Є–Є Escherichia coli. –§–Њ–ї–∞—В–љ—Л–є —Ж–Є–Ї–ї –њ—А–Є–Ј–љ–∞–љ –Ї–ї—О—З–µ–≤—Л–Љ —А–µ–≥—Г–ї—П—В–Њ—А–Њ–Љ –Ї–ї–µ—В–Њ—З–љ–Њ–≥–Њ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ –Є¬†–Є–љ—В–µ–≥—А–∞—В–Њ—А–Њ–Љ –љ—Г—В—А–Є–µ–љ—В–љ–Њ–≥–Њ —Б—В–∞—В—Г—Б–∞, –њ–Њ—Н—В–Њ–Љ—Г –љ–∞–Ј–љ–∞—З–µ–љ–Є–µ –Ь–Х–Ґ –Є–Љ–Є—В–Є—А—Г–µ—В —Н—Д—Д–µ–Ї—В –Њ–≥—А–∞–љ–Є—З–µ–љ–Є—П –Ї–∞–ї–Њ—А–Є–є–љ–Њ—Б—В–Є –њ–Є—В–∞–љ–Є—П. –£¬†—З–µ–ї–Њ–≤–µ–Ї–∞ –Ь–Х–Ґ —В–∞–Ї–ґ–µ –њ—А–Є–≤–Њ–і–Є—В –Ї¬†–і–µ—Д–Є—Ж–Є—В—Г —Д–Њ–ї–∞—В–∞ –Є¬†–≤–Є—В–∞–Љ–Є–љ–∞ –Т12 –Є¬†–њ–Њ–≤—Л—И–µ–љ–Є—О —Г—А–Њ–≤–љ—П¬†–≥–Њ–Љ–Њ—Ж–Є—Б—В–µ–Є–љ–∞. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –њ—А–µ–њ–∞—А–∞—В –і–µ–є—Б—В–≤—Г–µ—В –Ї–∞–Ї –њ—А—П–Љ–Њ–є –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є–є —Б—В—А–µ—Б—Б–Њ—А [12].
–Т¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є K. Forslund –Є¬†—Б–Њ–∞–≤—В., –≤¬†–Ї–Њ—В–Њ—А–Њ–Љ —Г—З–∞—Б—В–≤–Њ–≤–∞–ї–Є 784¬†–њ–∞—Ж–Є–µ–љ—В–∞ —Б¬†–°–Ф¬†2 —В–Є–њ–∞, –≤¬†–≥—А—Г–њ–њ–µ –Ь–Х–Ґ –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є¬†–≥—А—Г–њ–њ–Њ–є –Њ—В–Љ–µ—З–∞–ї–Њ—Б—М –Њ–±–Є–ї–Є–µ –±–∞–Ї—В–µ—А–Є–є Subdoligranulum –Є¬†Akkermansia. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, —В–µ—А–∞–њ–Є—П –њ—А–µ–њ–∞—А–∞—В–Њ–Љ –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–ї–∞—Б—М —Б¬†–±–Њ–ї–µ–µ –Ј–і–Њ—А–Њ–≤—Л–Љ —Б–Њ—Б—В–∞–≤–Њ–Љ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Ї–Є—И–µ—З–љ–Є–Ї–∞ [13]. –£¬†–њ–Њ–ї—Г—З–∞–≤—И–Є—Е –µ–≥–Њ –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Њ–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ Adlercreutsia¬†[14].
–§—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ—Л–є –∞–љ–∞–ї–Є–Ј –њ—А–Њ–і–µ–Љ–Њ–љ—Б—В—А–Є—А–Њ–≤–∞–ї –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–µ —Г–≤–µ–ї–Є—З–µ–љ–Є–µ —Б–Є–љ—В–µ–Ј–∞ –Ї–Њ—А–Њ—В–Ї–Њ—Ж–µ–њ–Њ—З–µ—З–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В, —В–∞–Ї–Є—Е –Ї–∞–Ї –±—Г—В–Є—А–∞—В –Є¬†–њ—А–Њ–њ–Є–Њ–љ–∞—В [13]. –£¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–°–Ф¬†2 —В–Є–њ–∞, –љ–µ¬†–њ–Њ–ї—Г—З–∞–≤—И–Є—Е –Ь–Х–Ґ, –њ–Њ–≤—Л—Б–Є–ї–∞—Б—М —З–Є—Б–ї–µ–љ–љ–Њ—Б—В—М Eubacterium –Є¬†Clostridiaceae SMB53 [14] –Є¬†—Б–Њ–Ї—А–∞—В–Є–ї–Њ—Б—М –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –±–∞–Ї—В–µ—А–Є–є, –њ—А–Њ–і—Г—Ж–Є—А—Г—О—Й–Є—Е –Ї–Њ—А–Њ—В–Ї–Њ—Ж–µ–њ–Њ—З–µ—З–љ—Л–µ –ґ–Є—А–љ—Л–µ –Ї–Є—Б–ї–Њ—В—Л (Roseburia, Subdoligranulum), –Є¬†–Ї–ї–∞—Б—В–µ—А–∞ –±—Г—В–Є—А–∞—В-–њ—А–Њ–і—Г—Ж–Є—А—Г—О—Й–Є—Е Clostridiales [13]. –≠—В–Њ –і–Њ–Ї–∞–Ј—Л–≤–∞–µ—В –≤–ї–Є—П–љ–Є–µ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –љ–∞¬†–∞–љ—В–Є–і–Є–∞–±–µ—В–Є—З–µ—Б–Ї–Є–є —Н—Д—Д–µ–Ї—В –Ь–Х–Ґ —З–µ—А–µ–Ј –і–µ–≥—А–∞–і–∞—Ж–Є—О –Љ—Г—Ж–Є–љ–∞ –Є¬†—Б–Є–љ—В–µ–Ј –Ї–Њ—А–Њ—В–Ї–Њ—Ж–µ–њ–Њ—З–µ—З–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В. F.H.¬†Karlsson –Є¬†—Б–Њ–∞–≤—В. —Г—Б—В–∞–љ–Њ–≤–Є–ї–Є —Б–≤—П–Ј—М –Љ–µ–ґ–і—Г –њ—А–Є–µ–Љ–Њ–Љ –Ь–Х–Ґ –Є¬†–њ–Њ–≤—Л—И–µ–љ–Є–µ–Љ –Ї–Њ–ї–Є—З–µ—Б—В–≤–∞ —Н–љ—В–µ—А–Њ–±–∞–Ї—В–µ—А–Є–є (Escherichia, Shigella, Klebsiella –Є¬†Salmonella), –∞¬†—В–∞–Ї–ґ–µ —Б–љ–Є–ґ–µ–љ–Є–µ–Љ —З–Є—Б–ї–∞ Clostridium –Є¬†Eubacterium —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–°–Ф 2¬†—В–Є–њ–∞ [15].
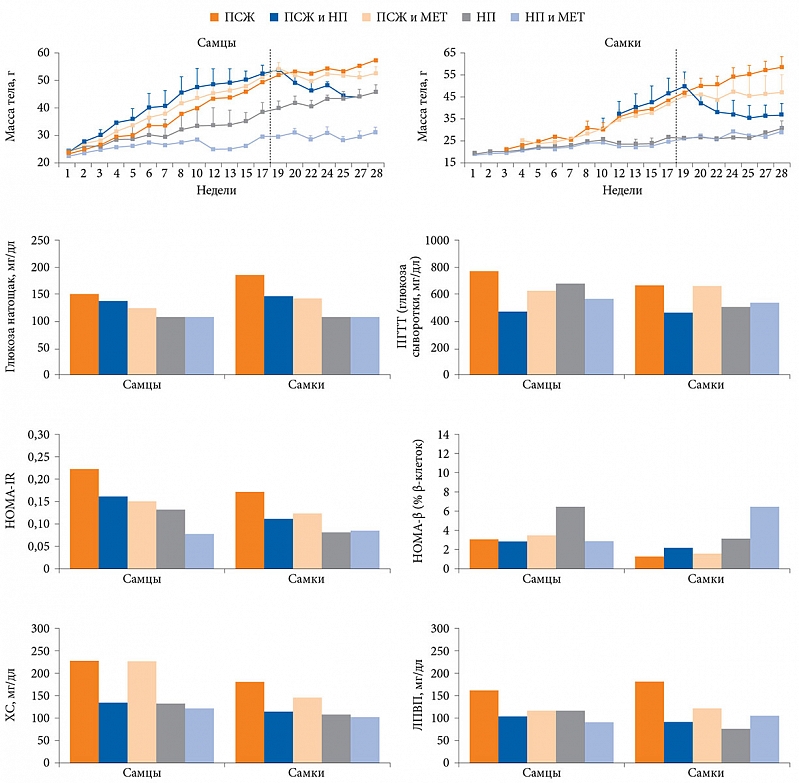
–Т¬†2014¬†–≥.¬†–Є–Ј—Г—З–µ–љ–Њ –≤–ї–Є—П–љ–Є–µ –і–Є–µ—В–Њ—В–µ—А–∞–њ–Є–Є –Є¬†–Ь–Х–Ґ –љ–∞¬†–Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є–µ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є. –®–µ—Б—В–Є–љ–µ–і–µ–ї—М–љ—Л–µ –Љ—Л—И–Є –≤ —В–µ—З–µ–љ–Є–µ 28 –љ–µ–і–µ–ї—М –љ–∞—Е–Њ–і–Є–ї–Є—Б—М –љ–∞¬†–і–Є–µ—В–µ —Б¬†–њ–Њ–≤—Л—И–µ–љ–љ—Л–Љ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ–Љ –ґ–Є—А–Њ–≤ (–Я–°–Ц) [16]. –Ш—Е¬†—А–∞–Ј–і–µ–ї–Є–ї–Є –љ–∞¬†–љ–µ—Б–Ї–Њ–ї—М–Ї–Њ¬†–≥—А—Г–њ–њ: –њ–µ—А–≤–∞—П¬†вАУ –і–Є–µ—В–∞ —Б¬†–Я–°–Ц —Б¬†–њ–µ—А–µ—Е–Њ–і–Њ–Љ –≤¬†–і–∞–ї—М–љ–µ–є—И–µ–Љ –љ–∞¬†–Ь–Х–Ґ (300 –Љ–≥/–Ї–≥) –≤¬†—В–µ—З–µ–љ–Є–µ –і–µ—Б—П—В–Є –љ–µ–і–µ–ї—М, –≤—В–Њ—А–∞—П¬†вАУ –і–Є–µ—В–∞ —Б¬†–Я–°–Ц, —В—А–µ—В—М—П¬†вАУ –і–Є–µ—В–∞ —Б¬†–Я–°–Ц —Б¬†–њ–µ—А–µ—Е–Њ–і–Њ–Љ –љ–∞¬†–љ–Њ—А–Љ–∞–ї—М–љ–Њ–µ –њ–Є—В–∞–љ–Є–µ (–Э–Я) –≤¬†—В–µ—З–µ–љ–Є–µ –і–µ—Б—П—В–Є –љ–µ–і–µ–ї—М, —З–µ—В–≤–µ—А—В–∞—П¬†вАУ –Э–Я, –њ—П—В–∞—П¬†вАУ –Э–Я —Б¬†–њ–µ—А–µ—Е–Њ–і–Њ–Љ –≤¬†–і–∞–ї—М–љ–µ–є—И–µ–Љ –љ–∞¬†–Ь–Х–Ґ –≤¬†—В–µ—З–µ–љ–Є–µ –і–µ—Б—П—В–Є –љ–µ–і–µ–ї—М.
–Т¬†–≥—А—Г–њ–њ–µ –і–Є–µ—В—Л —Б¬†–Я–°–Ц –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –Љ–∞—Б—Б—Л —В–µ–ї–∞, —Г—А–Њ–≤–љ—П¬†–≥–ї—О–Ї–Њ–Ј—Л –љ–∞—В–Њ—Й–∞–Ї –Є¬†–љ–∞—А—Г—И–µ–љ–Є–µ —В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В–Є –Ї¬†–≥–ї—О–Ї–Њ–Ј–µ (–Э–Ґ–У) –њ—А–Є –њ—А–Њ–≤–µ–і–µ–љ–Є–Є –њ–µ—А–Њ—А–∞–ї—М–љ–Њ–≥–Њ¬†–≥–ї—О–Ї–Њ–Ј–Њ—В–Њ–ї–µ—А–∞–љ—В–љ–Њ–≥–Њ —В–µ—Б—В–∞ (–Я–У–Ґ–Ґ), –∞¬†—В–∞–Ї–ґ–µ —Б–љ–Є–ґ–µ–љ–Є–µ –Є–љ—Б—Г–ї–Є–љ–Њ—А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є —В–Ї–∞–љ–µ–є (HOMA-IR), –љ–∞—А—Г—И–µ–љ–Є–µ —Д—Г–љ–Ї—Ж–Є–Є ќ≤-–Ї–ї–µ—В–Њ–Ї (HOMA-ќ≤) –Є¬†–ї–Є–њ–Є–і–љ–Њ–≥–Њ –Њ–±–Љ–µ–љ–∞ (—А–Є—Б.¬†1). –Т¬†–≥—А—Г–њ–њ–µ –Я–°–Ц –Є¬†–Ь–Х–Ґ –Љ–∞—Б—Б–∞ —В–µ–ї–∞ –њ–Њ—Б–ї–µ –і–Њ–±–∞–≤–ї–µ–љ–Є—П –њ—А–µ–њ–∞—А–∞—В–∞ –љ–µ¬†—Г–≤–µ–ї–Є—З–Є–≤–∞–ї–∞—Б—М, –љ–µ—Б–Љ–Њ—В—А—П –љ–∞¬†–њ—А–Њ–і–Њ–ї–ґ–µ–љ–Є–µ –і–Є–µ—В—Л. –Ю—В–Љ–µ—З–∞–ї–Њ—Б—М —Б–љ–Є–ґ–µ–љ–Є–µ¬†–≥–ї–Є–Ї–µ–Љ–Є–Є, –Њ—Б–Њ–±–µ–љ–љ–Њ —Г¬†—Б–∞–Љ–Њ–Ї, –Є¬†HOMA-IR. –Ю–і–љ–∞–Ї–Њ —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ –≤–ї–Є—П–љ–Є—П –Љ–µ—В—Д–Њ—А–Љ–Є–љ–∞ –љ–∞¬†–Э–Ґ–У –Є¬†—Д—Г–љ–Ї—Ж–Є—О ќ≤-–Ї–ї–µ—В–Њ–Ї –љ–µ¬†–Ј–∞—А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–љ–Њ. –≠—Д—Д–µ–Ї—В –Ь–Х–Ґ –љ–∞¬†—Г—А–Њ–≤–µ–љ—М –Њ–±—Й–µ–≥–Њ —Е–Њ–ї–µ—Б—В–µ—А–Є–љ–∞ (–•–°) —А–∞–Ј–ї–Є—З–∞–ї—Б—П –≤¬†–Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В¬†–њ–Њ–ї–∞¬†вАУ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–µ —Б–љ–Є–ґ–µ–љ–Є–µ —Г¬†—Б–∞–Љ–Њ–Ї (—А = 0,023). –Ґ–µ—А–∞–њ–Є—П –Ь–Х–Ґ —В–∞–Ї–ґ–µ —Б–њ–Њ—Б–Њ–±—Б—В–≤–Њ–≤–∞–ї–∞ —Г–Љ–µ–љ—М—И–µ–љ–Є—О —Г—А–Њ–≤–љ—П –ї–Є–њ–Њ–њ—А–Њ—В–µ–Є–љ–Њ–≤ –≤—Л—Б–Њ–Ї–Њ–є –њ–ї–Њ—В–љ–Њ—Б—В–Є (–Ы–Я–Т–Я). –Т¬†–≥—А—Г–њ–њ–µ –Я–°–Ц –Є¬†–Э–Я –≤—Л—П–≤–ї–µ–љ–Њ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–µ —Б–љ–Є–ґ–µ–љ–Є–µ –Љ–∞—Б—Б—Л —В–µ–ї–∞, —Г—А–Њ–≤–љ—П¬†–≥–ї—О–Ї–Њ–Ј—Л –љ–∞—В–Њ—Й–∞–Ї, —Г–ї—Г—З—И–µ–љ–Є–µ –Э–Ґ–У, —Г–Љ–µ–љ—М—И–µ–љ–Є–µ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –•–°, –Ы–Я–Т–Я –Є¬†—А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є –Ї¬†–Є–љ—Б—Г–ї–Є–љ—Г. –Ю–і–љ–∞–Ї–Њ –њ–µ—А–µ—Е–Њ–і –љ–∞¬†–Э–Я, —В–∞–Ї –ґ–µ –Ї–∞–Ї —В–µ—А–∞–њ–Є—П –Љ–µ—В—Д–Њ—А–Љ–Є–љ–Њ–Љ, –љ–µ¬†–Њ–Ї–∞–Ј–∞–ї —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ –≤–ї–Є—П–љ–Є—П –љ–∞¬†–њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є HOMA-ќ≤. –Т¬†–≥—А—Г–њ–њ–µ –Э–Я –љ–µ¬†–±—Л–ї–Њ –≤—Л—П–≤–ї–µ–љ–Њ –љ–Є–Ї–∞–Ї–Є—Е –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –љ–∞—А—Г—И–µ–љ–Є–є, –≤¬†–≥—А—Г–њ–њ–µ –Э–Я –Є¬†–Ь–Х–Ґ –і–Њ–±–∞–≤–ї–µ–љ–Є–µ –њ—А–µ–њ–∞—А–∞—В–∞ –љ–µ¬†–њ–Њ–≤–ї–Є—П–ї–Њ –љ–∞¬†—Г—А–Њ–≤–µ–љ—М –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –±–Є–Њ–Љ–∞—А–Ї–µ—А–Њ–≤ (—Б–Љ. —А–Є—Б.¬†1).
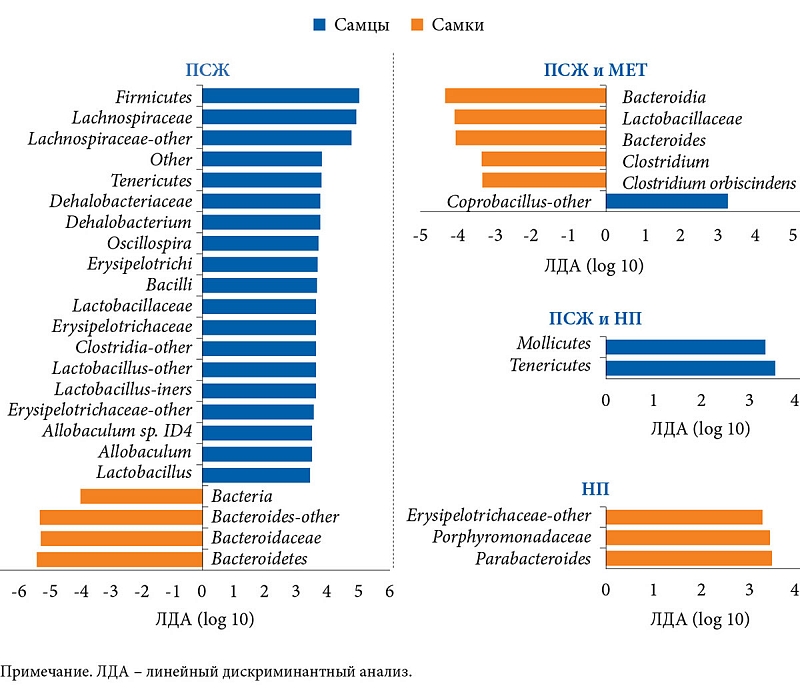
–Я–Њ–Љ–Є–Љ–Њ –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –Љ–∞—А–Ї–µ—А–Њ–≤ –≤¬†—А–∞–±–Њ—В–µ –Є–Ј—Г—З–∞–ї—Б—П —Б–Њ—Б—В–∞–≤ –Ь–Ъ. –Т¬†–≥—А—Г–њ–њ–µ –Я–°–Ц –Њ—В–Љ–µ—З–∞–ї–Њ—Б—М —Г–Љ–µ–љ—М—И–µ–љ–Є–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–∞ Bacteroidetes –і–Њ 43,79 ¬± 22,35% –Є¬†—Г–≤–µ–ї–Є—З–µ–љ–Є–µ —З–Є—Б–ї–µ–љ–љ–Њ—Б—В–Є Firmicutes –і–Њ 50,73 ¬± 19,2%. –Т¬†–≥—А—Г–њ–њ–µ –Э–Я, –љ–∞–Њ–±–Њ—А–Њ—В, –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ Bacteroidetes —Г–≤–µ–ї–Є—З–Є–ї–Њ—Б—М –і–Њ 79,4 ¬± 10,0%. –Т¬†–≥—А—Г–њ–њ–µ –Я–°–Ц –Є¬†–Ь–Х–Ґ –њ–Њ—Б–ї–µ –і–Њ–±–∞–≤–ї–µ–љ–Є—П –њ—А–µ–њ–∞—А–∞—В–∞ —З–Є—Б–ї–µ–љ–љ–Њ—Б—В—М Bacteroidetes –њ–Њ–≤—Л—Б–Є–ї–∞—Б—М –і–Њ 77,45 ¬± 8,73%. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –≤–Њ–Ј—А–Њ—Б–ї–Њ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ Verrucomicrobia –і–Њ 12,4 ¬± 5,26%, Akkermansia muciniphila –і–Њ 12,4 ¬± 5,26% –Є¬†Clostridium cocleatum –і–Њ 0,1 ¬± 0,09% –≤¬†–Њ—В–ї–Є—З–Є–µ –Њ—В¬†–≥—А—Г–њ–њ—Л –Я–°–Ц –Є¬†–≥—А—Г–њ–њ—Л –Я–°–Ц –Є¬†–Э–Я. –Я—А–Є–µ–Љ –Ь–Х–Ґ —В–∞–Ї–ґ–µ –њ–Њ–≤–ї–Є—П–ї –љ–∞¬†—Б–Њ—Б—В–∞–≤ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Љ—Л—И–µ–є, –њ–Њ–ї—Г—З–∞–≤—И–Є—Е –Э–Я. –І–Є—Б–ї–µ–љ–љ–Њ—Б—В—М —Б–µ–Љ–µ–є—Б—В–≤ Rikenellaceae, Ruminococcaceae –Є¬†Verrucomicrobiaceae, –∞¬†—В–∞–Ї–ґ–µ Alistipes spp., Akkermansia spp. –Є¬†Clostridium spp. –≤¬†–≥—А—Г–њ–њ–µ –Э–Я –Є¬†–Ь–Х–Ґ –±—Л–ї–∞ –≤—Л—И–µ, —З–µ–Љ –≤¬†–≥—А—Г–њ–њ–µ¬†–Э–Я.
–Т¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є –≤—Л—П–≤–ї–µ–љ—Л –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В–Є —Б–Њ—Б—В–∞–≤–∞ –Ь–Ъ –≤¬†–Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В¬†–≥–µ–љ–і–µ—А–љ–Њ–є –њ—А–Є–љ–∞–і–ї–µ–ґ–љ–Њ—Б—В–Є. –Ґ–∞–Ї, –≤¬†–≥—А—Г–њ–њ–µ –Я–°–Ц —З–Є—Б–ї–µ–љ–љ–Њ—Б—В—М Bacteroidetes –Њ–Ї–∞–Ј–∞–ї–∞—Б—М –≤—Л—И–µ —Г¬†—Б–∞–Љ–Њ–Ї, –≤¬†–≥—А—Г–њ–њ–µ –Я–°–Ц –Є¬†–Э–Я –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ Tenericutes –±—Л–ї–Њ –±–Њ–ї—М—И–µ —Г¬†—Б–∞–Љ—Ж–Њ–≤, –≤¬†–≥—А—Г–њ–њ–µ –Э–Я¬†вАУ Parabacteroides spp.¬†вАУ —Г¬†—Б–∞–Љ–Њ–Ї, –≤¬†–≥—А—Г–њ–њ–µ –Я–°–Ц –Є¬†–Ь–Х–Ґ¬†вАУ Coprobacillus spp.¬†вАУ —Г¬†—Б–∞–Љ—Ж–Њ–≤, –∞¬†Clostridium spp., Bacteroides spp., —Б–µ–Љ–µ–є—Б—В–≤–∞ Lactobacillaceae, –Ї–ї–∞—Б—Б–∞ Bacteroidia¬†вАУ —Г¬†—Б–∞–Љ–Њ–Ї. –†–∞–Ј–ї–Є—З–Є–є –≤¬†—Б–Њ—Б—В–∞–≤–µ –Ь–Ъ –Љ–µ–ґ–і—Г —Б–∞–Љ—Ж–∞–Љ–Є –Є¬†—Б–∞–Љ–Ї–∞–Љ–Є –≤¬†–≥—А—Г–њ–њ–µ –Э–Я –Є¬†–Ь–Х–Ґ –љ–µ¬†–Њ–±–љ–∞—А—Г–ґ–µ–љ–Њ (—А–Є—Б.¬†2).
–Т¬†–і—А—Г–≥–Њ–Љ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ–Њ–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є –љ–∞¬†–Љ—Л—И–∞—Е –≤—Л—П–≤–ї–µ–љ–Њ, —З—В–Њ –Ь–Х–Ґ –≤–ї–Є—П–µ—В –љ–∞¬†–њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–љ–Њ—Б—В—М –±–∞–Ї—В–µ—А–Є–Є A. muciniphila –Є¬†—Г–ї—Г—З—И–∞–µ—В¬†–≥–ї–Є–Ї–µ–Љ–Є—З–µ—Б–Ї–Є–є –Ї–Њ–љ—В—А–Њ–ї—М. –Э–∞¬†—Д–Њ–љ–µ —В–µ—А–∞–њ–Є–Є —В–∞–Ї–ґ–µ —Г–≤–µ–ї–Є—З–Є–ї–Њ—Б—М –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –±–Њ–Ї–∞–ї–Њ–≤–Є–і–љ—Л—Е –Ї–ї–µ—В–Њ–Ї, –њ—А–Њ–Є–Ј–≤–Њ–і—П—Й–Є—Е –Љ—Г—Ж–Є–љ. –Я–Њ—Б–ї–µ–і–љ–Є–є —П–≤–ї—П–µ—В—Б—П –Є—Б—В–Њ—З–љ–Є–Ї–Њ–Љ –њ–Є—В–∞–љ–Є—П –і–ї—П Akkermansia [17].
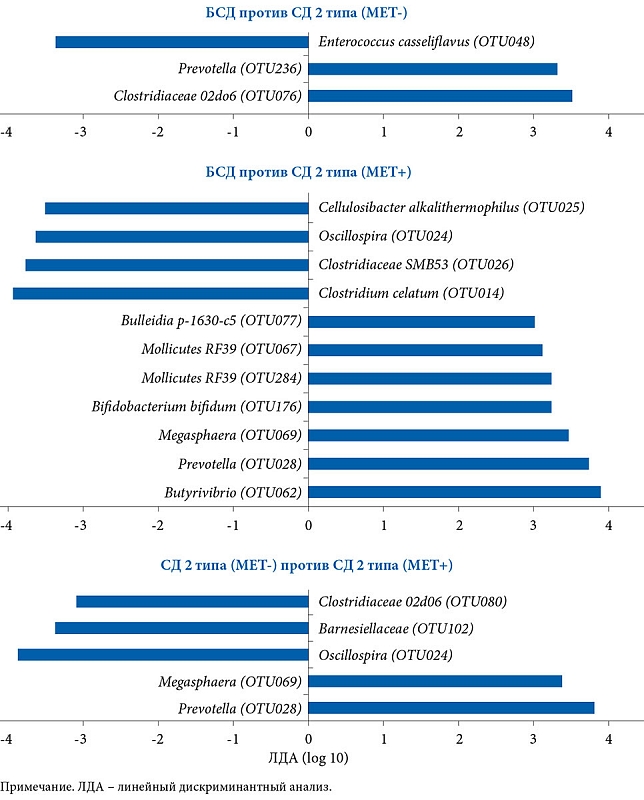
–Т¬†2016¬†–≥.¬†–њ—А–Њ–∞–љ–∞–ї–Є–Ј–Є—А–Њ–≤–∞–љ—Л —З–Є—Б–ї–µ–љ–љ–Њ—Б—В—М –Є¬†—Б–Њ—Б—В–∞–≤ –Ь–Ъ —Г¬†112 –Ї–Њ–ї—Г–Љ–±–Є–є—Б–Ї–Є—Е –ґ–µ–љ—Й–Є–љ [18]. 84 –Є–Ј¬†–љ–Є—Е –љ–µ¬†—Б—В—А–∞–і–∞–ї–Є –°–Ф¬†2 —В–Є–њ–∞ (–С–°–Ф). –°—А–µ–і–Є –њ–∞—Ж–Є–µ–љ—В–Њ–Ї —Б –°–Ф 2 —В–Є–њ–∞ 14¬†–њ—А–Є–љ–Є–Љ–∞–ї–Є –Ь–Х–Ґ (–Ь–Х–Ґ+), 14¬†вАУ –љ–µ—В (M–Х–Ґ-). –Т¬†–≥—А—Г–њ–њ–µ –°–Ф¬†2 —В–Є–њ–∞ –Є¬†–Ь–Х–Ґ- –Њ–і–љ–∞ —Г—З–∞—Б—В–љ–Є—Ж–∞ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –љ–∞—Е–Њ–і–Є–ї–∞—Б—М –љ–∞¬†–Є–љ—Б—Г–ї–Є–љ–µ, –і–≤–µ¬†вАУ –љ–∞¬†–≥–ї–Є–±–µ–љ–Ї–ї–∞–Љ–Є–і–µ, 11 —А–∞–љ–µ–µ –љ–µ¬†–њ–Њ–ї—Г—З–∞–ї–Є –Љ–µ–і–Є–Ї–∞–Љ–µ–љ—В–Њ–Ј–љ–Њ–≥–Њ –ї–µ—З–µ–љ–Є—П.
–°—В–∞—В–Є—Б—В–Є—З–µ—Б–Ї–Є –Ј–љ–∞—З–Є–Љ—Л—Е —А–∞–Ј–ї–Є—З–Є–є –њ–Њ¬†–і–µ–Љ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Њ–Љ—Г, –∞–љ—В—А–Њ–њ–Њ–Љ–µ—В—А–Є—З–µ—Б–Ї–Є–Љ, –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–Љ –њ–∞—А–∞–Љ–µ—В—А–∞–Љ –Љ–µ–ґ–і—Г¬†–≥—А—Г–њ–њ–∞–Љ–Є –љ–µ¬†–≤—Л—П–≤–ї–µ–љ–Њ.
–Т¬†–Њ—В–ї–Є—З–Є–µ –Њ—В¬†–≥—А—Г–њ–њ—Л –С–°–Ф¬†–≥—А—Г–њ–њ–∞ –°–Ф¬†2 —В–Є–њ–∞ –Є¬†–Ь–Х–Ґ+ –Є–Љ–µ–ї–∞ –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Є–µ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є¬†–≥–ї–Є–Ї–µ–Љ–Є–Є –љ–∞—В–Њ—Й–∞–Ї,¬†–≥–ї–Є–Ї–Є—А–Њ–≤–∞–љ–љ–Њ–≥–Њ¬†–≥–µ–Љ–Њ–≥–ї–Њ–±–Є–љ–∞ (HbA1c) –Є¬†HOMA-IR, –∞¬†—В–∞–Ї–ґ–µ –±–Њ–ї–µ–µ –љ–Є–Ј–Ї–Є–є —Г—А–Њ–≤–µ–љ—М –Є–љ—Б—Г–ї–Є–љ-—Б–µ–љ—Б–Є–±–Є–ї–Є–Ј–Є—А—Г—О—Й–µ–≥–Њ¬†–≥–Њ—А–Љ–Њ–љ–∞ –∞–і–Є–њ–Њ–љ–µ–Ї—В–Є–љ–∞ (—А < 0,05).
–Я–Њ—Б–ї–µ –њ—А–Њ–≤–µ–і–µ–љ–Є—П —Б–µ–Ї–≤–µ–љ–Є—А–Њ–≤–∞–љ–Є—П¬†–≥–µ–љ–∞ 16S —А–†–Э–Ъ –Њ–±–љ–∞—А—Г–ґ–µ–љ–∞ —Б–≤—П–Ј—М –Љ–µ–ґ–і—Г –°–Ф, —Б–Њ—Б—В–∞–≤–Њ–Љ –Є¬†—З–Є—Б–ї–µ–љ–љ–Њ—Б—В—М—О –Ь–Ъ, –Ї–Њ—В–Њ—А–∞—П –±—Л–ї–∞ –Љ–Њ–і–Є—Д–Є—Ж–Є—А–Њ–≤–∞–љ–∞ –љ–∞¬†—Д–Њ–љ–µ —В–µ—А–∞–њ–Є–Є –Ь–Х–Ґ (—А–Є—Б.¬†3 –Є¬†4).
–°—А–∞–≤–љ–Є—В–µ–ї—М–љ—Л–є –∞–љ–∞–ї–Є–Ј –Љ–µ–ґ–і—Г¬†–≥—А—Г–њ–њ–∞–Љ–Є –°–Ф¬†2 —В–Є–њ–∞ –Є¬†–Ь–Х–Ґ- –Є¬†–С–°–Ф –≤—Л—П–≤–Є–ї, —З—В–Њ –±–∞–Ї—В–µ—А–Є–Є, –Њ—В–љ–Њ—Б—П—Й–Є–µ—Б—П –Ї¬†–≥—А—Г–њ–њ–µ Clostridiaceae (Firmicutes/Clostridiaceae) –Є¬†Prevotella (Bacteroidetes/Prevotellaceae), –±–Њ–ї—М—И–µ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–°–Ф¬†2 —В–Є–њ–∞, –љ–µ¬†–њ–Њ–ї—Г—З–∞–≤—И–Є—Е –Ь–Х–Ґ, —В–Њ–≥–і–∞ –Ї–∞–Ї Enterococcus casseliflavus (Firmicutes/Enterococcaceae)¬†вАУ —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ –±–µ–Ј –°–Ф. –°—А–∞–≤–љ–Є—В–µ–ї—М–љ—Л–є –∞–љ–∞–ї–Є–Ј¬†–≥—А—Г–њ–њ –°–Ф¬†2 —В–Є–њ–∞ –Є¬†–Ь–Х–Ґ+ –Є¬†–С–°–Ф –њ–Њ–Ї–∞–Ј–∞–ї, —З—В–Њ –±–∞–Ї—В–µ—А–Є–Є Butyrivibrio (Firmicutes/Lachnospiraceae), –∞¬†—В–∞–Ї–ґ–µ —А–∞–Ј–љ—Л–µ —В–∞–Ї—Б–Њ–љ–Њ–Љ–Є—З–µ—Б–Ї–Є–µ¬†–≥—А—Г–њ–њ—Л Prevotella (Bacteroidetes/Prevotellaceae), Megasphaera (Firmicutes/Veillonellaceae), Bifidobacterium bifidum (Actinobacteria/Bifidobacteriaceae), –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –і–≤–µ —В–∞–Ї—Б–Њ–љ–Њ–Љ–Є—З–µ—Б–Ї–Є–µ¬†–≥—А—Г–њ–њ—Л Mollicutes (Tenericutes) –Є¬†Bulleidia p-1630-c5 (Firmicutes/Erysipelotrichaceae), –≤ –±–Њ–ї—М—И–µ–є —Б—В–µ–њ–µ–љ–Є –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л –≤¬†–њ–µ—А–≤–Њ–є¬†–≥—А—Г–њ–њ–µ. –Т¬†–≥—А—Г–њ–њ–µ –њ–∞—Ж–Є–µ–љ—В–Њ–≤, –љ–µ¬†—Б—В—А–∞–і–∞–≤—И–Є—Е –°–Ф 2¬†—В–Є–њ–∞, –±–Њ–ї–µ–µ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–љ—Л–Љ–Є –±—Л–ї–Є¬†–≥—А—Г–њ–њ—Л Clostridiales, –≤–Ї–ї—О—З–∞—П Clostridiumcelatum (Firmicutes/Clostridiaceae), Clostridiaceae SMB53 (Firmicutes/Clostridiaceae), Oscillospira (Firmicutes/Ruminococcaceae) –Є¬†Cellulosibacter alkalithermophilus (Firmicutes/Ruminococcaceae). –І–Є—Б–ї–µ–љ–љ–Њ—Б—В—М Prevotella (Bacteroidetes/Prevotellaceae) –Є¬†Megasphaera (Firmicutes/Veillonellaceae) –≤¬†–≥—А—Г–њ–њ–µ –°–Ф¬†2 —В–Є–њ–∞ –Є¬†–њ–Њ–ї—Г—З–∞–≤—И–Є—Е –Ь–Х–Ґ –Њ–Ї–∞–Ј–∞–ї–∞—Б—М –≤—Л—И–µ, —З–µ–Љ –≤¬†–≥—А—Г–њ–њ–µ –°–Ф¬†2 —В–Є–њ–∞ –Є¬†–љ–µ –њ—А–Є–љ–Є–Љ–∞–≤—И–Є—Е –њ—А–µ–њ–∞—А–∞—В, —В–Њ–≥–і–∞ –Ї–∞–Ї Oscillosira (Firmicutes/Ruminococcaceae), Barnesiellaceae (Bacteroidetes), Clostridiaceae 02d06 (Firmicutes/Clostridiaceae)¬†вАУ –≤¬†–њ–Њ—Б–ї–µ–і–љ–µ–є.
–Я–Њ—Б–ї–µ –Њ–±—К–µ–і–Є–љ–µ–љ–Є—П –Љ—Г—Ж–Є–љ-–і–µ–≥—А–∞–і–Є—А—Г—О—Й–Є—Е –Є¬†–±—Г—В–Є—А–∞—В-–њ—А–Њ–і—Г—Ж–Є—А—Г—О—Й–Є—Е –±–∞–Ї—В–µ—А–Є–є –Њ–±–љ–∞—А—Г–ґ–µ–љ–Њ, —З—В–Њ –≤¬†–≥—А—Г–њ–њ–µ –њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–°–Ф 2¬†—В–Є–њ–∞, –њ—А–Є–љ–Є–Љ–∞–≤—И–Є—Е –Ь–Х–Ґ, —З–Є—Б–ї–µ–љ–љ–Њ—Б—В—М –±–∞–Ї—В–µ—А–Є–є –Р. muciniphila –Є¬†Butyrivibrio –±—Л–ї–∞ –≤—Л—И–µ, —З–µ–Љ –≤¬†–≥—А—Г–њ–њ–µ, –љ–µ¬†–њ–Њ–ї—Г—З–∞–≤—И–µ–є —Г–Ї–∞–Ј–∞–љ–љ—Л–є –њ—А–µ–њ–∞—А–∞—В,¬†вАУ –≤¬†3,4¬†–Є¬†4,4 —А–∞–Ј–∞ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ. –†–∞–Ј–ї–Є—З–Є—П –±—Л–ї–Є —Б—В–∞—В–Є—Б—В–Є—З–µ—Б–Ї–Є –Ј–љ–∞—З–Є–Љ—Л–Љ–Є –і–ї—П A.¬†mucini¬≠phila (F1, 109 = 9,46, p = 0,003, q = 0,01), –љ–Њ –љ–µ¬†–і–ї—П Butyrivibrio (F1, 109 = 3,03, p = 0,08, q = 0,21) (—Б–Љ.¬†—А–Є—Б.¬†4–Ц –Є¬†–Ч). –Ф—А—Г–≥–Є—Е —Б—Г—Й–µ—Б—В–≤–µ–љ–љ—Л—Е —А–∞–Ј–ї–Є—З–Є–є –≤¬†–≥—А—Г–њ–њ–∞—Е –±—Г—В–Є—А–∞—В-—Б–Є–љ—В–µ–Ј–Є—А—Г—О—Й–Є—Е –±–∞–Ї—В–µ—А–Є–є –Ь–Ъ –±–Њ–ї—М–љ—Л—Е –°–Ф 2¬†—В–Є–њ–∞, –њ—А–Є–љ–Є–Љ–∞–≤—И–Є—Е –Є¬†–љ–µ –њ—А–Є–љ–Є–Љ–∞–≤—И–Є—Е –Ь–Х–Ґ, –љ–µ¬†–≤—Л—П–≤–ї–µ–љ–Њ. –Ф–ї—П Roseburia¬†вАУ F1, 109 = 1,44 (p = 0,23, q¬†= 0,39), Subdoligranulum¬†вАУ F1, 109 = 0,001 (p = 0,97, q = 0,97), Faecalibacterium¬†вАУ F1, 109 = 0,53 (p = 0,47, q = 0,59). –°—Г—Й–µ—Б—В–≤–µ–љ–љ—Л—Е —А–∞–Ј–ї–Є—З–Є–є –≤¬†–Њ—В–љ–Њ—И–µ–љ–Є–Є —Н—В–Є—Е –±–∞–Ї—В–µ—А–Є–є –њ—А–Є —Б—А–∞–≤–љ–µ–љ–Є–Є –Ь–Ъ –њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–°–Ф 2¬†—В–Є–њ–∞ –Є¬†–±–µ–Ј —В–∞–Ї–Њ–≤–Њ–≥–Њ –љ–µ¬†–Њ—В–Љ–µ—З–µ–љ–Њ (–≤—Б–µ –Ј–љ–∞—З–µ–љ–Є—П p > 0,1 –Є¬†q > 0,2).
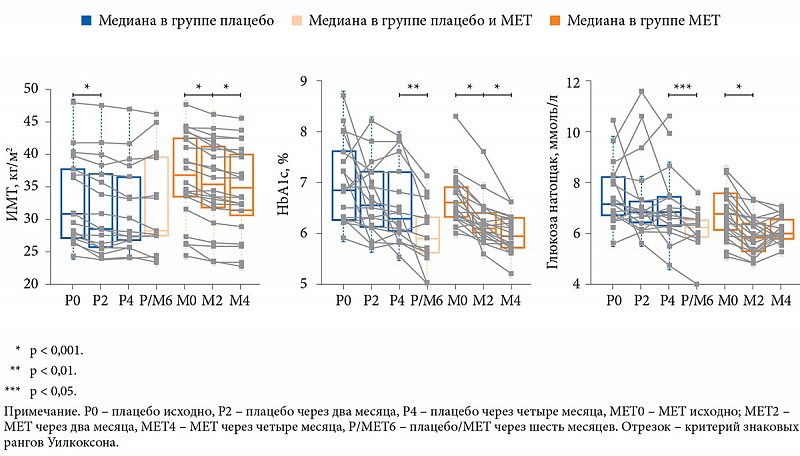
–Т¬†2017¬†–≥.¬†H. Wu –Є¬†—Б–Њ–∞–≤—В. [19] –њ—А–Њ–≤–µ–ї–Є –њ–µ—А–≤–Њ–µ –і–≤–Њ–є–љ–Њ–µ —Б–ї–µ–њ–Њ–µ —А–∞–љ–і–Њ–Љ–Є–Ј–Є—А–Њ–≤–∞–љ–љ–Њ–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –њ–Њ¬†–Њ—Ж–µ–љ–Ї–µ —Н—Д—Д–µ–Ї—В–Њ–≤ –Љ–µ—В—Д–Њ—А–Љ–Є–љ–∞ –љ–∞¬†–Ь–Ъ —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–°–Ф¬†2 —В–Є–њ–∞. –Т¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –≤–Ї–ї—О—З–µ–љ–Њ 40 –њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–≤–њ–µ—А–≤—Л–µ –≤—Л—П–≤–ї–µ–љ–љ–Њ–є –њ–∞—В–Њ–ї–Њ–≥–Є–µ–є. –£—З–∞—Б—В–љ–Є–Ї–Њ–≤ —А–∞–љ–і–Њ–Љ–Є–Ј–Є—А–Њ–≤–∞–ї–Є –љ–∞¬†–і–≤–µ¬†–≥—А—Г–њ–њ—Л: –њ–µ—А–≤–∞—П (n = 22) –њ–Њ–ї—Г—З–∞–ї–∞ –Ь–Х–Ґ 1700 –Љ–≥/—Б—Г—В, –≤—В–Њ—А–∞—П (n = 18)¬†вАУ –њ–ї–∞—Ж–µ–±–Њ. –Ф–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М —В–µ—А–∞–њ–Є–Є¬†вАУ —З–µ—В—Л—А–µ –Љ–µ—Б—П—Ж–∞.
–Я–∞—Ж–Є–µ–љ—В—Л –Њ–±–µ–Є—Е¬†–≥—А—Г–њ–њ –љ–∞—Е–Њ–і–Є–ї–Є—Б—М –љ–∞¬†–і–Є–µ—В–µ —Б¬†–Њ–≥—А–∞–љ–Є—З–µ–љ–Є–µ–Љ –Ї–∞–ї–Њ—А–Є–є–љ–Њ—Б—В–Є.
–І–µ—А–µ–Ј —И–µ—Б—В—М –Љ–µ—Б—П—Ж–µ–≤ –Њ—В¬†–љ–∞—З–∞–ї–∞ –љ–∞–±–ї—О–і–µ–љ–Є—П —З–∞—Б—В—М –њ–∞—Ж–Є–µ–љ—В–Њ–≤¬†–≥—А—Г–њ–њ—Л –њ–ї–∞—Ж–µ–±–Њ –±—Л–ї–∞ –њ–µ—А–µ–≤–µ–і–µ–љ–∞ –љ–∞¬†–Ь–Х–Ґ.
–Ъ–∞–Ї –Є¬†–Њ–ґ–Є–і–∞–ї–Њ—Б—М, –Ј–∞ —З–µ—В—Л—А–µ –Љ–µ—Б—П—Ж–∞ –≤¬†—А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–љ–Є–ґ–µ–љ–Є—П –Ї–∞–ї–Њ—А–Є–є–љ–Њ—Б—В–Є –њ–Є—Й–Є –Љ–∞—Б—Б–∞ —В–µ–ї–∞ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ —Г–Љ–µ–љ—М—И–Є–ї–∞—Б—М –Ї–∞–Ї –≤¬†–≥—А—Г–њ–њ–µ –њ–ї–∞—Ж–µ–±–Њ (—Б¬†85,4 ¬± 5,6 –і–Њ 81,5 ¬± 5,4 –Ї–≥), —В–∞–Ї –Є¬†–≤¬†–≥—А—Г–њ–њ–µ –Ь–Х–Ґ (—Б¬†96,5 ¬± 4,1 –і–Њ 91,4¬†¬± 3,9 –Ї–≥). –Т¬†–≥—А—Г–њ–њ–µ –њ–ї–∞—Ж–µ–±–Њ –љ–µ¬†–Њ—В–Љ–µ—З–∞–ї–Њ—Б—М —Б–љ–Є–ґ–µ–љ–Є—П —Г—А–Њ–≤–љ—П¬†–≥–ї–Є–Ї–µ–Љ–Є–Є –љ–∞—В–Њ—Й–∞–Ї –Є¬†HbA1c, –Њ–і–љ–∞–Ї–Њ —Д–Є–Ї—Б–Є—А–Њ–≤–∞–ї–Њ—Б—М –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–µ —Г–Љ–µ–љ—М—И–µ–љ–Є–µ —Г—А–Њ–≤–љ—П –Њ–±—Й–µ–≥–Њ –•–° (—А < 0,05), –ї–Є–њ–Њ–њ—А–Њ—В–µ–Є–љ–Њ–≤ –љ–Є–Ј–Ї–Њ–є –њ–ї–Њ—В–љ–Њ—Б—В–Є (—А < 0,05) –Є¬†–≥–∞–Љ–Љ–∞-–≥–ї—Г—В–∞–Љ–Є–ї—В—А–∞–љ—Б—Д–µ—А–∞–Ј—Л (—А < 0,01). –Т¬†–≥—А—Г–њ–њ–µ –Ь–Х–Ґ –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–µ —Б–љ–Є–ґ–µ–љ–Є–µ —Г—А–Њ–≤–љ—П HbA1c,¬†–≥–ї–Є–Ї–µ–Љ–Є–Є –љ–∞—В–Њ—Й–∞–Ї, HOMA-IR (—А < 0,05), —Г—А–Њ–≤–љ—П¬†–≥–∞–Љ–Љ–∞-–≥–ї—Г—В–∞–Љ–Є–ї—В—А–∞–љ—Б—Д–µ—А–∞–Ј—Л (—А < 0,05) –Є¬†—Г–≤–µ–ї–Є—З–µ–љ–Є–µ –Ы–Я–Т–Я (—А < 0,05).
–Ш–љ–і–µ–Ї—Б –Љ–∞—Б—Б—Л —В–µ–ї–∞ (–Ш–Ь–Ґ) –љ–µ¬†—Г–Љ–µ–љ—М—И–Є–ї—Б—П –≤¬†–њ–Њ–і–≥—А—Г–њ–њ–µ –њ–ї–∞—Ж–µ–±–Њ, –Ї–Њ—В–Њ—А–∞—П —З–µ—А–µ–Ј —И–µ—Б—В—М –Љ–µ—Б—П—Ж–µ–≤ –±—Л–ї–∞ –њ–µ—А–µ–≤–µ–і–µ–љ–∞ –љ–∞¬†–Ь–Х–Ґ, –Њ–і–љ–∞–Ї–Њ —Г—А–Њ–≤–љ–Є HbA1c –Є¬†–≥–ї–Є–Ї–µ–Љ–Є–Є –љ–∞—В–Њ—Й–∞–Ї –≤ –љ–µ–є –і–Њ—Б—В–Њ–≤–µ—А–љ–Њ —Б–љ–Є–Ј–Є–ї–Є—Б—М (—А–Є—Б.¬†5).
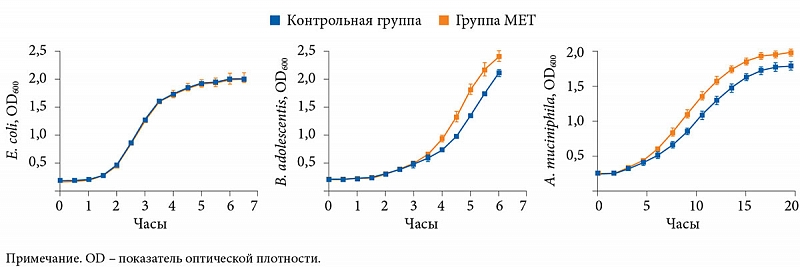
–Ь–µ—В–∞–≥–µ–љ–Њ–Љ–љ—Л–є –∞–љ–∞–ї–Є–Ј —Б¬†–њ–Њ–Љ–Њ—Й—М—О —Б–µ–Ї–≤–µ–љ–Є—А–Њ–≤–∞–љ–Є—П¬†–≥–µ–љ–∞ 16S —А–†–Э–Ъ –њ–Њ–Ї–∞–Ј–∞–ї, —З—В–Њ —В–µ—А–∞–њ–Є—П –Ь–Х–Ґ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї—М—Б—В–≤–Њ –±–∞–Ї—В–µ—А–Є–є Escherichia coli, Bifidobacterium adolescentis (—А = 0,01) –Є¬†A. muciniphila (—А = 0,008), –њ—А–Є —Н—В–Њ–Љ —Б–љ–Є–ґ–∞–µ—В –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ Intestinibacter (—А–Є—Б.¬†6). –Ш—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї–Є —В–∞–Ї–ґ–µ –Њ–±–љ–∞—А—Г–ґ–Є–ї–Є, —З—В–Њ –Љ–µ—В—Д–Њ—А–Љ–Є–љ in¬†vitro —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В —Г–≤–µ–ї–Є—З–µ–љ–Є—О –Ї–Њ–ї–Є—З–µ—Б—В–≤–∞ B. adolescentis –Є¬†A. muciniphila. –Ф–Њ—Б—В–Њ–≤–µ—А–љ—Л—Е –Ї–Њ—А—А–µ–ї—П—Ж–Є–є –Љ–µ–ґ–і—Г —Г—А–Њ–≤–љ–µ–Љ HbA1c –Є¬†A. muciniphila –≤—Л—П–≤–ї–µ–љ–Њ –љ–µ¬†–±—Л–ї–Њ, —Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ, –љ–µ–ї—М–Ј—П —Б–і–µ–ї–∞—В—М –≤—Л–≤–Њ–і –Њ¬†—В–Њ–Љ, —З—В–Њ A. muciniphila —П–≤–ї—П–µ—В—Б—П –Њ—Б–љ–Њ–≤–љ—Л–Љ —Д–∞–Ї—В–Њ—А–Њ–Љ –њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ–≥–Њ –≤–ї–Є—П–љ–Є—П –Ь–Х–Ґ.
H. Wu –Є¬†—Б–Њ–∞–≤—В. —В–∞–Ї–ґ–µ –њ—А–Њ–≤–µ–ї–Є —В—А–∞–љ—Б–њ–ї–∞–љ—В–∞—Ж–Є—О –Ь–Ъ –њ–∞—Ж–Є–µ–љ—В–Њ–≤, –њ–Њ–ї—Г—З–∞–≤—И–Є—Е –Ь–Х–Ґ, —Б—В–µ—А–Є–ї—М–љ—Л–Љ –Љ—Л—И–∞–Љ. –Я–Њ—Б–ї–µ –љ–µ–µ —Г¬†–Љ—Л—И–µ–є —Г–ї—Г—З—И–Є–ї–∞—Б—М —В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В—М –Ї¬†–≥–ї—О–Ї–Њ–Ј–µ. –Ь–∞—Б—Б–∞ —В–µ–ї–∞, –ґ–Є—А–Њ–≤—Л–µ –Њ—В–ї–Њ–ґ–µ–љ–Є—П –Є¬†–Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є—П –Є–љ—Б—Г–ї–Є–љ–∞ –љ–µ¬†–Є–Ј–Љ–µ–љ–Є–ї–Є—Б—М.
–≠—Д—Д–µ–Ї—В –љ–∞¬†–≥–ї—О–Ї–∞–≥–Њ–љ–Њ–њ–Њ–і–Њ–±–љ—Л–є¬†–њ–µ–њ—В–Є–і 1
–У–Њ–Љ–µ–Њ—Б—В–∞–Ј –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–∞ –њ–Њ–і–і–µ—А–ґ–Є–≤–∞–µ—В—Б—П –±–ї–∞–≥–Њ–і–∞—А—П –њ—А–Њ–і—Г–Ї—Ж–Є–Є —А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–љ—Л—Е –Љ–Є–Ї—А–Њ–±–љ—Л—Е —Б–Њ–µ–і–Є–љ–µ–љ–Є–є, –Ї–Њ—В–Њ—А—Л–µ —А–µ–≥—Г–ї–Є—А—Г—О—В —Б–Ї–Њ—А–Њ—Б—В—М –Є¬†–≤—Л—А–∞–ґ–µ–љ–љ–Њ—Б—В—М —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е —Д—Г–љ–Ї—Ж–Є–є, –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –Є¬†–њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є—Е —А–µ–∞–Ї—Ж–Є–є.
–Я–Њ–ї–Є—Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ–∞—П —А–Њ–ї—М –њ—А–Є–љ–∞–і–ї–µ–ґ–Є—В –Ї–Њ—А–Њ—В–Ї–Њ—Ж–µ–њ–Њ—З–µ—З–љ—Л–Љ –ґ–Є—А–љ—Л–Љ –Ї–Є—Б–ї–Њ—В–∞–Љ. –Ю—Б–љ–Њ–≤–љ—Г—О –Є—Е –Љ–∞—Б—Б—Г —Б–Њ—Б—В–∞–≤–ї—П—О—В —Г–Ї—Б—Г—Б–љ–∞—П (–∞—Ж–µ—В–∞—В), –њ—А–Њ–њ–Є–Њ–љ–Њ–≤–∞—П (–њ—А–Њ–њ–Є–Њ–љ–∞—В), –Є–Ј–Њ–Љ–∞—Б–ї—П–љ–∞—П, –Љ–∞—Б–ї—П–љ–∞—П (–±—Г—В–Є—А–∞—В), –Є–Ј–Њ–≤–∞–ї–µ—А–Є–∞–љ–Њ–≤–∞—П, –≤–∞–ї–µ—А–Є–∞–љ–Њ–≤–∞—П (–≤–∞–ї–µ—А–∞—В), –Є–Ј–Њ–Ї–∞–њ—А–Њ–љ–Њ–≤–∞—П –Є¬†–Ї–∞–њ—А–Њ–љ–Њ–≤–∞—П (–≥–µ–Ї—Б–∞–љ–∞—В) –Ї–Є—Б–ї–Њ—В—Л. –Ю–љ–Є –Њ–±—А–∞–Ј—Г—О—В—Б—П –≤¬†—В–Њ–ї—Б—В–Њ–є –Ї–Є—И–Ї–µ –њ—Г—В–µ–Љ —Д–µ—А–Љ–µ–љ—В–∞—Ж–Є–Є —Г–≥–ї–µ–≤–Њ–і–Њ–≤, –ґ–Є—А–Њ–≤ –Є¬†–±–µ–ї–Ї–Њ–≤. –С—Г—В–Є—А–∞—В¬†вАУ –Њ—Б–љ–Њ–≤–љ–Њ–є –Є—Б—В–Њ—З–љ–Є–Ї —Н–љ–µ—А–≥–Є–Є –∞–і–µ–љ–Њ–Ј–Є–љ—В—А–Є—Д–Њ—Б—Д–∞—В–∞. –Р—Ж–µ—В–∞—В –Є¬†–њ—А–Њ–њ–Є–Њ–љ–∞—В –≤¬†–≥–µ–њ–∞—В–Њ—Ж–Є—В–∞—Е —Г—З–∞—Б—В–≤—Г—О—В –≤¬†–≥–ї—О–Ї–Њ–љ–µ–Њ–≥–µ–љ–µ–Ј–µ –Є¬†–ї–Є–њ–Њ–≥–µ–љ–µ–Ј–µ [20].
–Т¬†2014¬†–≥.¬†–њ—А–Њ–≤–µ–і–µ–љ–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –љ–∞¬†–Ї—А—Л—Б–∞—Е, –њ–Њ–і—В–≤–µ—А–і–Є–≤—И–µ–µ, —З—В–Њ –±—Г—В–Є—А–∞—В –љ–∞–њ—А—П–Љ—Г—О –∞–Ї—В–Є–≤–Є—А—Г–µ—В —Н–Ї—Б–њ—А–µ—Б—Б–Є—О¬†–≥–µ–љ–Њ–≤, –Ј–∞–і–µ–є—Б—В–≤–Њ–≤–∞–љ–љ—Л—Е –≤¬†–≥–ї—О–Ї–Њ–љ–µ–Њ–≥–µ–љ–µ–Ј–µ –≤¬†–Ї–ї–µ—В–Ї–∞—Е –Ї–Є—И–µ—З–љ–Є–Ї–∞, –њ–Њ—Б—А–µ–і—Б—В–≤–Њ–Љ —Ж–Є–Ї–ї–Є—З–µ—Б–Ї–Њ–≥–Њ –∞–і–µ–љ–Њ–Ј–Є–љ–Љ–Њ–љ–Њ—Д–Њ—Б—Д–∞—В-–Ј–∞–≤–Є—Б–Є–Љ–Њ–≥–Њ –Љ–µ—Е–∞–љ–Є–Ј–Љ–∞, –≤¬†—В–Њ –≤—А–µ–Љ—П –Ї–∞–Ї –њ—А–Њ–њ–Є–Њ–љ–∞—В¬†вАУ —З–µ—А–µ–Ј –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є–µ —Б¬†—А–µ—Ж–µ–њ—В–Њ—А–∞–Љ–Є —Б–≤–Њ–±–Њ–і–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В 3 (FFAR3), —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–љ—Л–Љ–Є –≤¬†–Ї–ї–µ—В–Ї–∞—Е –љ–µ—А–≤–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л. –°–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ, –Ї–Њ—А–Њ—В–Ї–Њ—Ж–µ–њ–Њ—З–µ—З–љ—Л–µ –ґ–Є—А–љ—Л–µ –Ї–Є—Б–ї–Њ—В—Л –≤–ї–Є—П—О—В –љ–∞¬†—Г–≥–ї–µ–≤–Њ–і–љ—Л–є –Њ–±–Љ–µ–љ —З–µ—А–µ–Ј —А–µ–≥—Г–ї–Є—А–Њ–≤–∞–љ–Є–µ¬†–≥–ї—О–Ї–Њ–љ–µ–Њ–≥–µ–љ–µ–Ј–∞ –≤¬†–Ї–ї–µ—В–Ї–∞—Е –Ї–Є—И–µ—З–љ–Є–Ї–∞ [21].
–Т¬†1987¬†–≥.¬†–±—Л–ї –Њ—В–Ї—А—Л—В —З–µ–ї–Њ–≤–µ—З–µ—Б–Ї–Є–є¬†–≥–ї—О–Ї–∞–≥–Њ–љ–Њ–њ–Њ–і–Њ–±–љ—Л–є –њ–µ–њ—В–Є–і 1 (–У–Я–Я-1), –Ї–Њ—В–Њ—А—Л–є —Б–Њ—Б—В–Њ–Є—В –Є–Ј¬†30¬†–∞–Љ–Є–љ–Њ–Ї–Є—Б–ї–Њ—В–љ—Л—Е –Њ—Б—В–∞—В–Ї–Њ–≤, –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ –і–≤—Г–Љ—П –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є –∞–Ї—В–Є–≤–љ—Л–Љ–Є —Д–Њ—А–Љ–∞–Љ–Є¬†вАУ –У–Я–Я-1-(7-37) –Є¬†–У–Я–Я-1-(7-36)NH2 (80% –≤—Б–µ–≥–Њ –њ—Г–ї–∞ –У–Я–Я-1), —Б–Є–љ—В–µ–Ј–Є—А—Г–µ—В—Б—П L-–Ї–ї–µ—В–Ї–∞–Љ–Є, –ї–Њ–Ї–∞–ї–Є–Ј–Њ–≤–∞–љ–љ—Л–Љ–Є –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –≤¬†—Б–ї–Є–Ј–Є—Б—В–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–µ –і–Є—Б—В–∞–ї—М–љ–Њ–≥–Њ –Њ—В–і–µ–ї–∞ —В–Њ–љ–Ї–Њ–є –Ї–Є—И–Ї–Є, –∞¬†—В–∞–Ї–ґ–µ —В–Њ–ї—Б—В–Њ–є –Ї–Є—И–Ї–Є.
–Т¬†–љ–Њ—А–Љ–µ —Б–Є–љ—В–µ–Ј –Њ—Б–љ–Њ–≤–љ—Л—Е –Є–љ–Ї—А–µ—В–Є–љ–Њ–≤¬†вАУ¬†–≥–∞—Б—В—А–Њ–Є–љ–≥–Є–±–Є—А—Г—О—Й–µ–≥–Њ –њ–µ–њ—В–Є–і–∞ –Є¬†–У–Я–Я-1 –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –≤¬†–Њ—В–≤–µ—В –љ–∞¬†–њ–Њ—Б—В—Г–њ–ї–µ–љ–Є–µ –њ–Є—Й–Є –Є–ї–Є¬†–≥–ї—О–Ї–Њ–Ј—Л –≤¬†–Ї–Є—И–µ—З–љ–Є–Ї. –Ю—Б–љ–Њ–≤–љ—Л–Љ–Є —Н—Д—Д–µ–Ї—В–∞–Љ–Є –У–Я–Я-1 —Б—З–Є—В–∞—О—В—Б—П —Б—В–Є–Љ—Г–ї—П—Ж–Є—П¬†–≥–ї—О–Ї–Њ–Ј–Њ–Ј–∞–≤–Є—Б–Є–Љ–Њ–є —Б–µ–Ї—А–µ—Ж–Є–Є –Є–љ—Б—Г–ї–Є–љ–∞, —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –Љ–∞—Б—Б—Л ќ≤-–Ї–ї–µ—В–Њ–Ї –њ–Њ–і–ґ–µ–ї—Г–і–Њ—З–љ–Њ–є –ґ–µ–ї–µ–Ј—Л, –Є–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є–µ –≤—Л—Б–≤–Њ–±–Њ–ґ–і–µ–љ–Є—П¬†–≥–ї—О–Ї–∞–≥–Њ–љ–∞, –Њ–њ–Њ—А–Њ–ґ–љ–µ–љ–Є–µ –ґ–µ–ї—Г–і–Ї–∞ –Є¬†—Б–љ–Є–ґ–µ–љ–Є–µ –∞–њ–њ–µ—В–Є—В–∞.
–£—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ —Б–µ–Ї—А–µ—Ж–Є—П –У–Я–Я-1 –Њ–±—Г—Б–ї–Њ–≤–ї–µ–љ–∞ –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є–µ–Љ –Ї–Њ—А–Њ—В–Ї–Њ—Ж–µ–њ–Њ—З–µ—З–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В —Б¬†—А–µ—Ж–µ–њ—В–Њ—А–∞–Љ–Є, —Б–≤—П–Ј–∞–љ–љ—Л–Љ–Є —Б¬†G-–±–µ–ї–Ї–∞–Љ–Є —Б–≤–Њ–±–Њ–і–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В 2 –Є¬†3 (GPR41 (FFAR3) –Є¬†GPR43 (FFAR2)) [22]. FFAR2 –Њ–±–µ—Б–њ–µ—З–Є–≤–∞–µ—В —Б–Њ—Е—А–∞–љ–µ–љ–Є–µ —Н–љ–µ—А–≥–Є–Є –Ј–∞ —Б—З–µ—В —Б—В–Є–Љ—Г–ї—П—Ж–Є–Є –ї–Є–њ–Њ–≥–µ–љ–µ–Ј–∞, –Є–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є–µ –ї–Є–њ–Њ–ї–Є–Ј–∞ –Є¬†—Г–Љ–µ–љ—М—И–µ–љ–Є–µ —А–∞—Б—Е–Њ–і–∞ —Н–љ–µ—А–≥–Є–Є. –Т¬†—В–Њ–ї—Б—В–Њ–є –Ї–Є—И–Ї–µ FFAR2 –Є¬†FFAR3 —А–µ–≥—Г–ї–Є—А—Г—О—В –њ–µ—А–Є—Б—В–∞–ї—М—В–Є–Ї—Г –Ї–Є—И–µ—З–љ–Є–Ї–∞ –Є¬†–љ–∞—Б—Л—Й–µ–љ–Є–µ —З–µ—А–µ–Ј –У–Я–Я-1.
–Т¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є H. Lin –Є¬†—Б–Њ–∞–≤—В., –њ—А–Њ–≤–µ–і–µ–љ–љ–Њ–Љ –≤¬†2012¬†–≥.¬†[22], —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ –Ї–Њ—А–Њ—В–Ї–Њ—Ж–µ–њ–Њ—З–µ—З–љ—Л–µ –ґ–Є—А–љ—Л–µ –Ї–Є—Б–ї–Њ—В—Л, –≤–≤–µ–і–µ–љ–љ—Л–µ –Љ—Л—И–∞–Љ, –≤–ї–Є—П—О—В –љ–∞¬†—Б–Є–љ—В–µ–Ј –Ї–Є—И–µ—З–љ—Л—Е¬†–≥–Њ—А–Љ–Њ–љ–Њ–≤ —З–µ—А–µ–Ј —А–µ—Ж–µ–њ—В–Њ—А—Л FFAR2 –Є¬†FFAR3, –Ј–∞—Й–Є—Й–∞—П –Њ—В¬†–Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–љ–љ–Њ–≥–Њ –і–Є–µ—В–Њ–є –Њ–ґ–Є—А–µ–љ–Є—П –Є¬†–Є–љ—Б—Г–ї–Є–љ–Њ—А–µ–Ј–Є—Б—В–µ–љ—В–љ–Њ—Б—В–Є.
–£—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ –±—Г—В–Є—А–∞—В –Є¬†–њ—А–Њ–њ–Є–Њ–љ–∞—В —Б—В–Є–Љ—Г–ї–Є—А—Г—О—В –≤—Л—А–∞–±–Њ—В–Ї—Г –Ї–Є—И–µ—З–љ—Л—Е¬†–≥–Њ—А–Љ–Њ–љ–Њ–≤ –Є¬†—Б–Њ–Ї—А–∞—Й–∞—О—В –Њ–±—Й–µ–µ –љ–∞—Б—Л—Й–µ–љ–Є–µ –љ–µ–Ј–∞–≤–Є—Б–Є–Љ–Њ –Њ—В¬†FFAR3. –≠—В–Є –і–∞–љ–љ—Л–µ —Г–Ї–∞–Ј—Л–≤–∞—О—В –љ–∞¬†–љ–Њ–≤—Л–є –Љ–µ—Е–∞–љ–Є–Ј–Љ –≤–ї–Є—П–љ–Є—П –Ь–Ъ –љ–∞¬†–Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ –Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–∞.
–Т¬†2014¬†–≥.¬†–≤—Л–і–≤–Є–љ—Г—В–∞ —В–µ–Њ—А–Є—П –Њ¬†–њ—А—П–Љ–Њ–є —Б–≤—П–Ј–Є –±–∞–Ї—В–µ—А–Є–є Bifidobacterium —Б¬†—Б–Є–љ—В–µ–Ј–Њ–Љ –У–Я–Я-1 [23]. –°–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ, –µ—Б–ї–Є –≤¬†–Ї–Є—И–µ—З–љ–Є–Ї–µ —Г–Љ–µ–љ—М—И–∞–µ—В—Б—П —З–Є—Б–ї–µ–љ–љ–Њ—Б—В—М Bifidobacterium, —Б–љ–Є–ґ–∞–µ—В—Б—П –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –У–Я–Я-1.
–Р–љ–∞–ї–Њ–≥–Є—З–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ –љ–∞¬†—Б–Є–љ—В–µ–Ј –У–Я–Я-1 –≤–ї–Є—П–µ—В Lactobacillus. –Т¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ–Њ–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є F. Duan –Є¬†—Б–Њ–∞–≤—В. [24] –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ –њ—А–Њ–±–Є–Њ—В–Є–Ї–∞ —В–Є–њ–∞ Lactobacillus, —И—В–∞–Љ–Љ—Л –Ї–Њ—В–Њ—А–Њ–≥–Њ —П–≤–ї—П—О—В—Б—П —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ–є —З–∞—Б—В—М—О –Ј–і–Њ—А–Њ–≤–Њ–є –Ь–Ъ, —Г¬†–Ї—А—Л—Б —Б¬†–°–Ф¬†–≤¬†—В–µ—З–µ–љ–Є–µ 90 –і–љ–µ–є –њ—А–Є–≤–µ–ї–Њ –Ї¬†—Б–љ–Є–ґ–µ–љ–Є—О —Г—А–Њ–≤–љ—П¬†–≥–ї–Є–Ї–µ–Љ–Є–Є –љ–∞¬†30% –±–Њ–ї—М—И–µ, —З–µ–Љ —Г¬†–Ї—А—Л—Б —Б¬†–і–Є–∞–±–µ—В–Њ–Љ, –љ–µ –њ–Њ–ї—Г—З–∞–≤—И–Є—Е –њ—А–Њ–±–Є–Њ—В–Є–Ї–∞.
–Т¬†1998¬†–≥., –µ—Й–µ –і–Њ –Њ—В–Ї—А—Л—В–Є—П –≤–ї–Є—П–љ–Є—П –±–∞–Ї—В–µ—А–Є–є –љ–∞¬†—Б–Є–љ—В–µ–Ј –У–Я–Я-1, R.¬†Lugari –Є¬†—Б–Њ–∞–≤—В. –Њ–±–љ–∞—А—Г–ґ–Є–ї–Є, —З—В–Њ —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–°–Ф¬†2 —В–Є–њ–∞ –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ –Ь–Х–Ґ —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В –њ–Њ–≤—Л—И–µ–љ–Є—О –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –њ–Њ—Б—В–њ—А–∞–љ–і–Є–∞–ї—М–љ–Њ–≥–Њ —Г—А–Њ–≤–љ—П –У–Я–Я-1 [25]. –Т¬†–і–∞–ї—М–љ–µ–є—И–µ–Љ —Г–Ї–∞–Ј–∞–љ–љ—Л–є —Н—Д—Д–µ–Ї—В –±—Л–ї –њ–Њ–і—В–≤–µ—А–ґ–і–µ–љ –≤¬†–Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л—Е –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П—Е [26вАУ28].
–Т¬†2001 –Є¬†2004¬†–≥–≥. E. Mannucci –Є¬†—Б–Њ–∞–≤—В. [26, 27] –Є–Ј—Г—З–∞–ї–Є —Н—Д—Д–µ–Ї—В—Л –Ь–Х–Ґ –љ–∞¬†–Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є—О –У–Я–Я-1 –Є¬†–ї–µ–њ—В–Є–љ–∞ —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–Њ–ґ–Є—А–µ–љ–Є–µ–Љ. –Т¬†–Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ 2001¬†–≥.¬†–±—Л–ї–Є –≤–Ї–ї—О—З–µ–љ—Л 20¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–Њ–ґ–Є—А–µ–љ–Є–µ–Љ (–Ш–Ь–Ґ > 30,0 –Ї–≥/–Љ2) –±–µ–Ј –°–Ф 2¬†—В–Є–њ–∞. –£—З–∞—Б—В–љ–Є–Ї–Њ–≤ —А–∞–љ–і–Њ–Љ–Є–Ј–Є—А–Њ–≤–∞–ї–Є –љ–∞¬†–і–≤–µ¬†–≥—А—Г–њ–њ—Л, –Њ–і–љ–∞ –Є–Ј¬†–Ї–Њ—В–Њ—А—Л—Е –њ—А–Є–љ–Є–Љ–∞–ї–∞ –Ь–Х–Ґ 2550 –Љ–≥/—Б—Г—В (–њ–Њ¬†850 –Љ–≥ —В—А–Є —А–∞–Ј–∞ –≤¬†–і–µ–љ—М) –≤¬†—В–µ—З–µ–љ–Є–µ 14 –і–љ–µ–є.
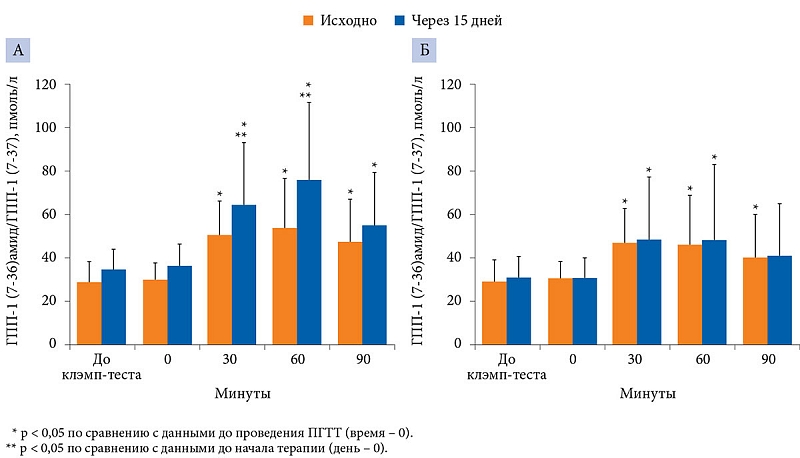
–Ш—Б—Е–Њ–і–љ–Њ –Є¬†–љ–∞ 15-–є¬†–і–µ–љ—М –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –Њ—Ж–µ–љ–Є–≤–∞–ї–Є —Г—А–Њ–≤–µ–љ—М –У–Я–Я-1 –Є¬†–ї–µ–њ—В–Є–љ–∞ –і–Њ –Є¬†–њ–Њ—Б–ї–µ –Я–У–Ґ–Ґ. –Ф–ї—П –Є—Б–Ї–ї—О—З–µ–љ–Є—П –Љ–µ—В—Д–Њ—А–Љ–Є–љ-–Є–љ–і—Г—Ж–Є—А–Њ–≤–∞–љ–љ—Л—Е –Є–Ј–Љ–µ–љ–µ–љ–Є–є —Г—А–Њ–≤–љ—П¬†–≥–ї–Є–Ї–µ–Љ–Є–Є –Є¬†–Є–љ—Б—Г–ї–Є–љ–µ–Љ–Є–Є –Я–У–Ґ–Ґ –њ—А–Њ–≤–Њ–і–Є–ї—Б—П –љ–∞¬†—Д–Њ–љ–µ —Н—Г–≥–ї–Є–Ї–µ–Љ–Є—З–µ—Б–Ї–Њ–≥–Њ¬†–≥–Є–њ–µ—А–Є–љ—Б—Г–ї–Є–љ–µ–Љ–Є—З–µ—Б–Ї–Њ–≥–Њ –Ї–ї—Н–Љ–њ-—В–µ—Б—В–∞.
–Э–∞¬†15-–є –і–µ–љ—М –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ—А–Є –њ—А–Њ–≤–µ–і–µ–љ–Є–Є –Я–У–Ґ–Ґ –≤¬†–≥—А—Г–њ–њ–µ –Ь–Х–Ґ –Ј–∞—Д–Є–Ї—Б–Є—А–Њ–≤–∞–љ–Њ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–µ (—А < 0,05) —Г–≤–µ–ї–Є—З–µ–љ–Є–µ –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є —Ж–Є—А–Ї—Г–ї–Є—А—Г—О—Й–µ–≥–Њ –У–Я–Я-1 (–У–Я–Я-1 (7-36)–∞–Љ–Є–і –Є¬†–У–Я–Я-1 (7-37)) –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–Є—Б—Е–Њ–і–љ—Л–Љ–Є –њ–Њ–Ї–∞–Ј–∞—В–µ–ї—П–Љ–Є. –°—А–∞–≤–љ–Є—В–µ–ї—М–љ—Л–є –∞–љ–∞–ї–Є–Ј¬†–≥—А—Г–њ–њ –≤—Л—П–≤–Є–ї –і–Њ—Б—В–Њ–≤–µ—А–љ–Њ–µ –њ–Њ–≤—Л—И–µ–љ–Є–µ —Г—А–Њ–≤–љ—П –У–Я–Я-1 (—А < 0,05) –≤¬†–≥—А—Г–њ–њ–µ —В–µ—А–∞–њ–Є–Є –Ь–Х–Ґ –њ–Њ¬†—Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†–Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є¬†–≥—А—Г–њ–њ–Њ–є —З–µ—А–µ–Ј 30 (63,8 ¬± 29,0¬†–њ—А–Њ—В–Є–≤ 50,3 ¬± 15,6¬†–њ–Љ–Њ–ї—М/–ї) –Є¬†60 –Љ–Є–љ—Г—В (75,8 ¬± 35,4 –њ—А–Њ—В–Є–≤ 46,9 ¬± 20,0¬†–њ–Љ–Њ–ї—М/–ї). –І–µ—А–µ–Ј¬†90 –Љ–Є–љ—Г—В –і–Њ—Б—В–Њ–≤–µ—А–љ–Њ–≥–Њ –њ–Њ–≤—Л—И–µ–љ–Є—П –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –У–Я–Я-1 –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –Є—Б—Е–Њ–і–љ–Њ–≥–Њ —Г—А–Њ–≤–љ—П –љ–µ¬†–Њ—В–Љ–µ—З–∞–ї–Њ—Б—М. –Т¬†–Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є¬†–≥—А—Г–њ–њ–µ —З–µ—А–µ–Ј 15¬†–і–љ–µ–є —Б—Г—Й–µ—Б—В–≤–µ–љ–љ—Л—Е –Є–Ј–Љ–µ–љ–µ–љ–Є–є —Г—А–Њ–≤–љ—П –У–Я–Я-1 –Њ—В¬†–Є—Б—Е–Њ–і–љ–Њ–≥–Њ –≤¬†—Г–Ї–∞–Ј–∞–љ–љ—Л–µ –≤—А–µ–Љ–µ–љ–љ—Л–µ –њ—А–Њ–Љ–µ–ґ—Г—В–Ї–Є –љ–µ¬†–Ј–∞—Д–Є–Ї—Б–Є—А–Њ–≤–∞–љ–Њ (—А–Є—Б.¬†7).
–£—А–Њ–≤–µ–љ—М –ї–µ–њ—В–Є–љ–∞ –≤¬†–≥—А—Г–њ–њ–µ –Ь–Х–Ґ –Є—Б—Е–Њ–і–љ–Њ —Б–Њ—Б—В–∞–≤–ї—П–ї 14,3 ¬± 6,6 –љ–≥/–Љ–ї, –≤¬†–Ї–Њ–љ—В—А–Њ–ї—М–љ–Њ–є¬†–≥—А—Г–њ–њ–µ¬†вАУ 15,8 ¬± 6,9 –љ–≥/–Љ–ї. –І–µ—А–µ–Ј 15 –і–љ–µ–є –і–Њ –Є¬†–њ–Њ—Б–ї–µ –њ—А–Њ–≤–µ–і–µ–љ–Є—П –Я–У–Ґ–Ґ –Њ–љ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –љ–µ¬†–Њ—В–ї–Є—З–∞–ї—Б—П –Њ—В¬†–Є—Б—Е–Њ–і–љ—Л—Е –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–µ–є (14,8 ¬± 6,4 –Є¬†15,2 ¬± 6,3 –љ–≥/–Љ–ї —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ).
–Т¬†2004¬†–≥.¬†E. Mannucci –Є¬†—Б–Њ–∞–≤—В. [26] –њ—А–Њ–і–Њ–ї–ґ–Є–ї–Є –Є–Ј—Г—З–µ–љ–Є–µ —Н—Д—Д–µ–Ї—В–Њ–≤ –Ь–Х–Ґ –љ–∞¬†–У–Я–Я-1 —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–Њ–ґ–Є—А–µ–љ–Є–µ–Љ –Є¬†–°–Ф¬†2 —В–Є–њ–∞. 34 —Г—З–∞—Б—В–љ–Є–Ї–∞ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –±—Л–ї–Є —А–∞–љ–і–Њ–Љ–Є–Ј–Є—А–Њ–≤–∞–љ—Л –љ–∞¬†–і–≤–µ¬†–≥—А—Г–њ–њ—Л. –Т¬†–њ–µ—А–≤—Г—О¬†–≥—А—Г–њ–њ—Г –≤–Ї–ї—О—З–µ–љ—Л –ї–Є—Ж–∞ —Б¬†–°–Ф 2¬†—В–Є–њ–∞ (n = 22), –≤–Њ –≤—В–Њ—А—Г—О¬†вАУ –±–µ–Ј —Г–Ї–∞–Ј–∞–љ–љ–Њ–є –њ–∞—В–Њ–ї–Њ–≥–Є–Є (n = 12).
–Т—Б–µ–Љ –њ–∞—Ж–Є–µ–љ—В–∞–Љ –љ–∞–Ј–љ–∞—З–µ–љ –Ь–Х–Ґ –≤¬†–і–Њ–Ј–µ 850 –Љ–≥ –≤¬†–њ–µ—А–≤—Л–є –і–µ–љ—М –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –Є¬†2550 –Љ–≥ (–њ–Њ¬†850 –Љ–≥ —В—А–Є —А–∞–Ј–∞ –≤¬†–і–µ–љ—М) –≤¬†—В–µ—З–µ–љ–Є–µ —Б–ї–µ–і—Г—О—Й–Є—Е —З–µ—В—Л—А–µ—Е –љ–µ–і–µ–ї—М.
–£—А–Њ–≤–µ–љ—М –У–У–Я-1 –≤¬†–Њ–±–µ–Є—Е¬†–≥—А—Г–њ–њ–∞—Е –Њ–њ—А–µ–і–µ–ї—П–ї–Є –њ—А–Є –њ—А–Њ–≤–µ–і–µ–љ–Є–Є –Я–У–Ґ–Ґ –≤¬†–њ–µ—А–≤—Л–є –і–µ–љ—М –Є¬†—З–µ—А–µ–Ј —З–µ—В—Л—А–µ –љ–µ–і–µ–ї–Є.
–£—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ –Њ–і–љ–Њ–Ї—А–∞—В–љ—Л–є –њ—А–Є–µ–Љ –Ь–Х–Ґ –љ–µ¬†–≤–ї–Є—П–ї –љ–∞¬†—Г—А–Њ–≤–µ–љ—М –У–Я–Я-1. –Я–Њ¬†–Њ–Ї–Њ–љ—З–∞–љ–Є–Є —В–µ—А–∞–њ–Є–Є –Ј–љ–∞—З–µ–љ–Є—П –У–Я–Я-1 —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–°–Ф¬†2 —В–Є–њ–∞ —Г–≤–µ–ї–Є—З–Є–ї–Є—Б—М —Б¬†3,8 –і–Њ 4,9 –њ–Љ–Њ–ї—М/–ї (—А < 0,05). –Я–Њ—Б–ї–µ –њ—А–Њ–≤–µ–і–µ–љ–Є—П –Я–У–Ґ–Ґ —Г—А–Њ–≤–µ–љ—М –У–Я–Я-1 —Г¬†–љ–Є—Е –±—Л–ї –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –љ–Є–ґ–µ. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П –Є–љ–Ї—А–µ–Љ–µ–љ—В–љ–∞—П –њ–ї–Њ—Й–∞–і—М –њ–Њ–і –Ї—А–Є–≤–Њ–є (AUC) –У–Я–Я-1 –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ —Г–≤–µ–ї–Є—З–Є–≤–∞–ї–∞—Б—М –Ї–∞–Ї —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–°–Ф¬†2 —В–Є–њ–∞ (—Б¬†93,6 (45,6вАУ163,2) –і–Њ 151,2 (36,0вАУ300,5) –њ–Љ–Њ–ї—М √Ч –Љ–Є–љ/–ї (p < 0,05)), —В–∞–Ї –Є¬†—Г –њ–∞—Ж–Є–µ–љ—В–Њ–≤ –±–µ–Ј –°–Ф¬†2 —В–Є–њ–∞ (—Б¬†187,2 (149,4вАУ571,8) –і–Њ 324,0 (238,2вАУ744,0) –њ–Љ–Њ–ї—М √Ч –Љ–Є–љ/–ї (p < 0,05)).
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П, –њ—А–Њ–≤–µ–і–µ–љ–љ—Л–µ E. Mannucci –Є¬†—Б–Њ–∞–≤—В. [26, 27], –њ—А–Њ–і–µ–Љ–Њ–љ—Б—В—А–Є—А–Њ–≤–∞–ї–Є, —З—В–Њ –Ь–Х–Ґ —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В —Г—А–Њ–≤–µ–љ—М –У–Я–Я-1 –Ї–∞–Ї —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–°–Ф¬†2 —В–Є–њ–∞, —В–∞–Ї –Є¬†—Г –њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–љ–Њ—А–Љ–∞–ї—М–љ–Њ–є —В–Њ–ї–µ—А–∞–љ—В–љ–Њ—Б—В—М—О –Ї¬†–≥–ї—О–Ї–Њ–Ј–µ.
–Т¬†–љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П –њ—А–µ–і–ї–Њ–ґ–µ–љ—Л –і–≤–µ¬†–≥–Є–њ–Њ—В–µ–Ј—Л –Њ¬†–Љ–µ—Е–∞–љ–Є–Ј–Љ–∞—Е –≤–ї–Є—П–љ–Є—П –Ь–Х–Ґ –љ–∞¬†—Г—А–Њ–≤–µ–љ—М –У–Я–Я-1. –Я—А–µ–њ–∞—А–∞—В –і–µ–є—Б—В–≤—Г–µ—В –Ї–∞–Ї –њ—А—П–Љ–Њ–є –Є/–Є–ї–Є –Ї–Њ—Б–≤–µ–љ–љ—Л–є —Б–µ–Ї—А–µ—В–∞–≥–Њ–≥ –У–Я–Я-1 –Є–ї–Є –Ї–∞–Ї –Є–љ–≥–Є–±–Є—В–Њ—А –і–Є–њ–µ–њ—В–Є–і–Є–ї–њ–µ–њ—В–Є–і–∞–Ј—Л 4, –њ—А–Њ–і–ї–µ–≤–∞—П –њ–µ—А–Є–Њ–і –њ–Њ–ї—Г–≤—Л–≤–µ–і–µ–љ–Є—П –∞–Ї—В–Є–≤–љ–Њ–≥–Њ –У–Я–Я-1 [28, 29].
–Ч–∞–Ї–ї—О—З–µ–љ–Є–µ
–°¬†—А–∞–Ј–≤–Є—В–Є–µ–Љ –Љ–Њ–ї–µ–Ї—Г–ї—П—А–љ–Њ-–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є—Е —В–µ—Е–љ–Њ–ї–Њ–≥–Є–є –њ–Њ—П–≤–Є–ї–Є—Б—М –љ–Њ–≤—Л–µ –і–∞–љ–љ—Л–µ –Њ¬†—Б–≤—П–Ј–Є –Є–Ј–Љ–µ–љ–µ–љ–љ–Њ–є –Ї–Є—И–µ—З–љ–Њ–є –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –љ–µ¬†—В–Њ–ї—М–Ї–Њ —Б¬†–Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П–Љ–Є –ґ–µ–ї—Г–і–Њ—З–љ–Њ-–Ї–Є—И–µ—З–љ–Њ–≥–Њ —В—А–∞–Ї—В–∞, –љ–Њ¬†–Є —Б¬†–Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є–Љ —Б–Є–љ–і—А–Њ–Љ–Њ–Љ, —Б–∞—Е–∞—А–љ—Л–Љ –і–Є–∞–±–µ—В–Њ–Љ 2 —В–Є–њ–∞. –Я–Њ—Б–ї–µ–і–љ–Є–µ –і–∞–љ–љ—Л–µ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О—В –Њ¬†–њ–Њ—В–µ–љ—Ж–Є–∞–ї—М–љ–Њ–є —А–Њ–ї–Є –Ь–Ъ –Ї–∞–Ї –њ–∞—В–Њ–≥–µ–љ–љ–Њ–≥–Њ —Д–∞–Ї—В–Њ—А–∞, –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–љ–љ–Њ–≥–Њ —Б¬†–Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є–Љ–Є –љ–∞—А—Г—И–µ–љ–Є—П–Љ–Є. –Ю—З–µ–≤–Є–і–љ–Њ, —З—В–Њ –њ–Њ–і–і–µ—А–ґ–∞–љ–Є–µ¬†–≥–Њ–Љ–µ–Њ—Б—В–∞–Ј–∞ –Є¬†–љ–Њ—А–Љ–∞–ї—М–љ–Њ–≥–Њ –Њ–±–Љ–µ–љ–∞ –≤–µ—Й–µ—Б—В–≤ –љ–µ–≤–Њ–Ј–Љ–Њ–ґ–љ–Њ –±–µ–Ј –≤–Њ—Б—Б—В–∞–љ–Њ–≤–ї–µ–љ–Є—П –Ї–∞—З–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ –Љ–Є–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤ –Ї–Є—И–µ—З–љ–Є–Ї–∞.
–Т¬†–љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П –≤¬†–љ–∞—Г—З–љ–Њ–є –ї–Є—В–µ—А–∞—В—Г—А–µ –њ–Њ–Љ–Є–Љ–Њ –і–Є–µ—В–Њ—В–µ—А–∞–њ–Є–Є –Є¬†–њ—А–Є–Љ–µ–љ–µ–љ–Є—П –њ—А–µ- –Є¬†–њ—А–Њ–±–Є–Њ—В–Є–Ї–Њ–≤ –∞–Ї—В–Є–≤–љ–Њ –Њ–±—Б—Г–ґ–і–∞—О—В—Б—П —Н—Д—Д–µ–Ї—В—Л –Ь–Х–Ґ –љ–∞¬†–Љ–Њ–і—Г–ї—П—Ж–Є—О –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л –Є¬†–Љ–∞–Ї—А–Њ–Њ—А–≥–∞–љ–Є–Ј–Љ. –£—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ, —З—В–Њ –Ь–Х–Ґ –≤–ї–Є—П–µ—В –љ–∞¬†—Б–Є–љ—В–µ–Ј –У–Я–Я-1, –Є¬†–љ–µ –Є—Б–Ї–ї—О—З–µ–љ–Њ, —З—В–Њ —Г–Ї–∞–Ј–∞–љ–љ—Л–є —Н—Д—Д–µ–Ї—В —А–µ–∞–ї–Є–Ј—Г–µ—В—Б—П —З–µ—А–µ–Ј –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Г –Є¬†—Б–Є–љ—В–µ–Ј –Ї–Њ—А–Њ—В–Ї–Њ—Ж–µ–њ–Њ—З–µ—З–љ—Л—Е –ґ–Є—А–љ—Л—Е –Ї–Є—Б–ї–Њ—В. –Т —Н—В–Њ–є —Б–≤—П–Ј–Є –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В—Б—П –∞–Ї—В—Г–∞–ї—М–љ—Л–Љ –Є–Ј—Г—З–Є—В—М –≤–ї–Є—П–љ–Є–µ –∞–≥–Њ–љ–Є—Б—В–Њ–≤ —А–µ—Ж–µ–њ—В–Њ—А–Њ–≤ –У–Я–Я-1 –љ–∞¬†–Љ–Є–Ї—А–Њ–±–Є–Њ—В—Г –Ї–Є—И–µ—З–љ–Є–Ї–∞ –љ–µ¬†—В–Њ–ї—М–Ї–Њ —Б¬†—Ж–µ–ї—М—О –Ї–Њ—А—А–µ–Ї—Ж–Є–Є –Љ–µ—В–∞–±–Њ–ї–Є—З–µ—Б–Ї–Є—Е –љ–∞—А—Г—И–µ–љ–Є–є, –љ–Њ¬†–Є –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ–≥–Њ —Г–ї—Г—З—И–µ–љ–Є—П –Ї–∞—З–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ –Є¬†–Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ –Љ–Є–Ї—А–Њ–±–Є–Њ—В—Л.
N.E. Khachaturyan, L.V. Egshatyan
A.I. Yevdokimov Moscow State University of Medicine and Dentistry
A.S. Loginov Moscow Clinical Scientific Practical Center
National Medical Research Center of Endocrinology
Contact person: Lilit Vanikovna Egshatyan, lilit.egshatyan@yandex.ru
Recently data appeared on the role of the gut microbiota in obesity-related metabolic disorders and diabetes mellitus type 2 pathogenesis. Therefore, it is relevant to identify drugs aimed at hyperglycemia and metabolic disorders correction and influencing by the way of gut microbiota modulation. Their use can have positive effect on the macroorganism.
The article presents the literature review on the impact of oral antihyperglycemic therapy, particularly metformin, on gut microbiota. Metformin is commonly used as the first line of medication for the treatment of prediabetes and type 2 diabetes. The effect of metformin on the gut microbiota has been reported; however, the relationship between the gut microbiota and the mechanism of action of metformin in elderly individuals is unclear. Recently, metformin-induced changes in the abundance of Akkermansia muciniphila were shown to be associated with metabolic improvement. Metformin has also been reported to increase levels of glucagon-like peptide 1 in humans and it is possible that this effect is also realized through the modulation of gut microbiota. Metformin increase synthesis of short chain fatty acids by bacteria. It becomes relevant to study the influence of glucagon-like peptide 1 receptor agonists on gut microbiota and improvement of the qualitative composition of the microbiota.
–£–≤–∞–ґ–∞–µ–Љ—Л–є –њ–Њ—Б–µ—В–Є—В–µ–ї—М uMEDp!
–£–≤–µ–і–Њ–Љ–ї—П–µ–Љ –Т–∞—Б –Њ —В–Њ–Љ, —З—В–Њ –Ј–і–µ—Б—М —Б–Њ–і–µ—А–ґ–Є—В—Б—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П, –њ—А–µ–і–љ–∞–Ј–љ–∞—З–µ–љ–љ–∞—П –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –і–ї—П —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П.
–Х—Б–ї–Є –Т—Л –љ–µ —П–≤–ї—П–µ—В–µ—Б—М —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–Љ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П, –∞–і–Љ–Є–љ–Є—Б—В—А–∞—Ж–Є—П –љ–µ –љ–µ—Б–µ—В –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ—Б—В–Є –Ј–∞ –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ –њ–Њ—Б–ї–µ–і—Б—В–≤–Є—П, –≤–Њ–Ј–љ–Є–Ї—И–Є–µ –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–∞–Љ–Њ—Б—В–Њ—П—В–µ–ї—М–љ–Њ–≥–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Т–∞–Љ–Є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є —Б –њ–Њ—А—В–∞–ї–∞ –±–µ–Ј –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є –Ї–Њ–љ—Б—Г–ї—М—В–∞—Ж–Є–Є —Б –≤—А–∞—З–Њ–Љ.
–Э–∞–ґ–Є–Љ–∞—П –љ–∞ –Ї–љ–Њ–њ–Ї—Г ¬Ђ–Т–Њ–є—В–Є¬ї, –Т—Л –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В–µ, —З—В–Њ —П–≤–ї—П–µ—В–µ—Б—М –≤—А–∞—З–Њ–Љ –Є–ї–Є —Б—В—Г–і–µ–љ—В–Њ–Љ –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–≥–Њ –≤—Г–Ј–∞.