–í–ĺ–∑–ľ–ĺ–∂–Ĺ–į –Ľ–ł –Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł–ļ–į –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–į–ľ–ł –ļ–į–Ľ—Ć—Ü–ł—Ź –ł –≤–ł—ā–į–ľ–ł–Ĺ–į D3?
- –ź–Ĺ–Ĺ–ĺ—ā–į—Ü–ł—Ź
- –°—ā–į—ā—Ć—Ź
- –°—Ā—č–Ľ–ļ–ł
–í–ĺ–Ņ—Ä–ĺ—Ā, –≤—č–Ĺ–Ķ—Ā–Ķ–Ĺ–Ĺ—č–Ļ –≤¬†–∑–į–≥–ĺ–Ľ–ĺ–≤–ĺ–ļ –ī–į–Ĺ–Ĺ–ĺ–Ļ —Ā—ā–į—ā—Ć–ł, –ľ–ĺ–∂–Ķ—ā –Ņ–ĺ–ļ–į–∑–į—ā—Ć—Ā—Ź –Ĺ–Ķ–Ľ–Ķ–Ņ—č–ľ. –ě–Ī—Č–Ķ–ł–∑–≤–Ķ—Ā—ā–Ĺ–ĺ, —á—ā–ĺ ¬ę–ļ–į–Ľ—Ć—Ü–ł–Ļ –ĺ—ā–ļ–Ľ–į–ī—č–≤–į–Ķ—ā—Ā—Ź –Ĺ–į¬†—Ā—ā–Ķ–Ĺ–ļ–į—Ö —Ā–ĺ—Ā—É–ī–ĺ–≤¬Ľ. –ü—Ä–ł —É–Ņ—Ä–ĺ—Č–Ķ–Ĺ–Ĺ–ĺ–ľ –≤–∑–≥–Ľ—Ź–ī–Ķ –ł–∑¬†—ć—ā–ĺ–Ļ –į–ļ—Ā–ł–ĺ–ľ—č –≤—č—ā–Ķ–ļ–į—é—ā –į–Ī—Ā—É—Ä–ī–Ĺ—č–Ķ –≤—č–≤–ĺ–ī—č: ¬ę–ļ–į–Ľ—Ć—Ü–ł–Ļ –ĺ–Ņ–į—Ā–Ķ–Ĺ –ī–Ľ—Ź –į—Ä—ā–Ķ—Ä–ł–Ļ¬Ľ, ¬ę–ļ–į–Ľ—Ć—Ü–ł–Ļ –≤—č–∑—č–≤–į–Ķ—ā –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—é –Ņ–Ľ–į—Ü–Ķ–Ĺ—ā—謼 –ł¬†—ā.–Ņ. –Ē—Ä—É–≥–ĺ–Ķ, –Ĺ–Ķ¬†–ľ–Ķ–Ĺ–Ķ–Ķ ¬ę–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ĺ–Ķ¬Ľ —É–ľ–ĺ–∑–į–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ: ¬ę–≤–ł—ā–į–ľ–ł–Ĺ D3¬†‚Äď —Ā–ł–Ĺ–Ķ—Ä–≥–ł—Ā—ā –ļ–į–Ľ—Ć—Ü–ł—Ź, –ļ–į–Ľ—Ć—Ü–ł–Ļ –ĺ–Ņ–į—Ā–Ķ–Ĺ, —Ā–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –≤–ł—ā–į–ľ–ł–Ĺ D3 —ā–ĺ–∂–Ķ –ĺ–Ņ–į—Ā–Ķ–Ĺ!¬Ľ. –Ē–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–Ķ —Ä–į–∑–≤–ł—ā–ł–Ķ —ć—ā–ĺ–≥–ĺ ¬ę—Ä–į—Ā—Ā—É–∂–ī–Ķ–Ĺ–ł—Ź¬Ľ –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†–Ķ—Č–Ķ –Ī–ĺ–Ľ–Ķ–Ķ ¬ę–Ī–Ľ–Ķ—Ā—ā—Ź—Č–ł–ľ¬Ľ –≤—č–≤–ĺ–ī–į–ľ: ¬ę–≤—Ā–Ķ –≤–ł—ā–į–ľ–ł–Ĺ—č –ĺ–Ņ–į—Ā–ŗ謼, ¬ę–Ņ—Ä–Ķ–Ņ–į—Ä–į—ā—č –≤–ł—ā–į–ľ–ł–Ĺ–į D3 –Ņ—Ä–ł–≤–ĺ–ī—Ź—ā –ļ¬†–∑–į—Ä–į—Ā—ā–į–Ĺ–ł—é —Ä–ĺ–ī–Ĺ–ł—á–ļ–ĺ–≤¬Ľ, ¬ę–Ī–Ķ—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ—č–ľ –Ĺ–Ķ¬†–Ĺ–į–ī–ĺ –≤–ł—ā–į–ľ–ł–Ĺ–ĺ–≤¬Ľ, ¬ę–ľ–į–≥–Ĺ–ł–Ļ –ł¬†–ļ–į–Ľ—Ć—Ü–ł–Ļ¬†‚Äď –ļ–į–ļ–į—Ź —Ä–į–∑–Ĺ–ł—Ü–į?!¬Ľ –ł¬†–ī—Ä. –ö¬†—Ā–ĺ–∂–į–Ľ–Ķ–Ĺ–ł—é, —ā–į–ļ–ĺ–≥–ĺ —Ä–ĺ–ī–į ¬ę–ī–ł–ļ–į—Ä—Ā–ļ–į—Ź –Ľ–ĺ–≥–ł–ļ–į¬Ľ –≤—Ā—ā—Ä–Ķ—á–į–Ķ—ā—Ā—Ź –ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ —á–į—Ā—ā–ĺ –Ĺ–Ķ¬†—ā–ĺ–Ľ—Ć–ļ–ĺ –≤¬†–Ņ—É–Ī–Ľ–ł—á–Ĺ—č—Ö –≤—č—Ā—ā—É–Ņ–Ľ–Ķ–Ĺ–ł—Ź—Ö –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö –ī–Ķ—Ź—ā–Ķ–Ľ–Ķ–Ļ –ĺ—ā¬†–ľ–Ķ–ī–ł—Ü–ł–Ĺ—č, –Ŗ嬆–ī–į–∂–Ķ –ł¬†–≤ –Ĺ–į—É—á–Ĺ—č—Ö —Ā—ā–į—ā—Ć—Ź—Ö. –°–ĺ–≤–Ķ—Ä—ą–Ķ–Ĺ–Ĺ–ĺ —Ź—Ā–Ĺ–ĺ, —á—ā–ĺ —Ä–Ķ–į–Ľ—Ć–Ĺ–į—Ź —Ā–ł—ā—É–į—Ü–ł—Ź –Ĺ–į–ľ–Ĺ–ĺ–≥–ĺ —Ā–Ľ–ĺ–∂–Ĺ–Ķ–Ķ, —á–Ķ–ľ —ć—ā–ĺ –ļ–į–∂–Ķ—ā—Ā—Ź –Ņ—Ä–ł —ā–į–ļ–ĺ–ľ —ā—Ä–ł–≤–ł–į–Ľ—Ć–Ĺ–ĺ–ľ –Ņ–ĺ–ī—Ö–ĺ–ī–Ķ.
–í–ĺ—ā —É–∂–Ķ –Ī–ĺ–Ľ–Ķ–Ķ –≤–Ķ–ļ–į –Ņ—Ä–ĺ—ą–Ľ–ĺ —Ā¬†—ā–Ķ—Ö –Ņ–ĺ—Ä, –ļ–į–ļ –≤—Ä–į—á–ł –∑–į–ľ–Ķ—ā–ł–Ľ–ł, —á—ā–ĺ —É¬†–Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ —Ā¬†–ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–ĺ–ľ —á–į—Č–Ķ –≤—Ā—ā—Ä–Ķ—á–į—é—ā—Ā—Ź —Ā–ĺ—Ā—É–ī–ł—Ā—ā—č–Ķ –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł—Ź, –≤¬†—ā–ĺ–ľ —á–ł—Ā–Ľ–Ķ –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑ –ł¬†–ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—Ź –į—Ä—ā–Ķ—Ä–ł–Ļ. –ē–∂–Ķ–ī–Ĺ–Ķ–≤–Ĺ–į—Ź –ļ–Ľ–ł–Ĺ–ł—á–Ķ—Ā–ļ–į—Ź –Ņ—Ä–į–ļ—ā–ł–ļ–į –Ņ–ĺ–ļ–į–∑—č–≤–į–Ķ—ā: –Ņ–į—Ü–ł–Ķ–Ĺ—ā—č,¬†–≥–ĺ—Ā–Ņ–ł—ā–į–Ľ–ł–∑–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č–Ķ —Ā¬†–Ņ–Ķ—Ä–Ķ–Ľ–ĺ–ľ–į–ľ–ł –≤—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł–Ķ –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–į,¬†–≥–ĺ—Ä–į–∑–ī–ĺ —á–į—Č–Ķ, —á–Ķ–ľ –Ņ–į—Ü–ł–Ķ–Ĺ—ā—č –Ī–Ķ–∑ –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–į, —Ā—ā—Ä–į–ī–į—é—ā –ĺ—ā¬†—Ā–ĺ—Ā—É–ī–ł—Ā—ā—č—Ö –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ (–ł–Ĺ—Ą–į—Ä–ļ—ā –ľ–ł–ĺ–ļ–į—Ä–ī–į, –ł–Ĺ—Ā—É–Ľ—Ć—ā [1]). –ě—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑ —Ā–≤—Ź–∑–į–Ĺ —Ā¬†–Ņ–ĺ—ā–Ķ—Ä—Ź–ľ–ł –ļ–į–Ľ—Ć—Ü–ł—Ź –ļ–ĺ—Ā—ā–Ĺ–ĺ–Ļ —ā–ļ–į–Ĺ—Ć—é, ¬†–≥–ł–Ņ–ĺ–ī–ł–Ĺ–į–ľ–ł–Ķ–Ļ, –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ—č–ľ —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł–Ķ–ľ —É—Ä–ĺ–≤–Ĺ–Ķ–Ļ —Ā—ā–Ķ—Ä–ĺ–ł–ī–Ĺ—č—Ö¬†–≥–ĺ—Ä–ľ–ĺ–Ĺ–ĺ–≤, –Ĺ–į—Ä—É—ą–Ķ–Ĺ–ł—Ź–ľ–ł –≤—Ā–į—Ā—č–≤–į–Ĺ–ł—Ź –ļ–į–Ľ—Ć—Ü–ł—Ź –≤—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł–Ķ –ī–ł—Ā–Ī–ł–ĺ–∑–ĺ–≤ –ł¬†–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤ —Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź —Ā–Ľ–ł–∑–ł—Ā—ā–ĺ–Ļ –Ė–ö–Ę, –Ĺ–į—Ä—É—ą–Ķ–Ĺ–ł–Ļ —Ą—É–Ĺ–ļ—Ü–ł–ł –Ņ–Ķ—á–Ķ–Ĺ–ł –ł¬†—ā.–ī. –ö–Ľ–ł–Ĺ–ł—á–Ķ—Ā–ļ–ł–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź –ī–Ķ–ľ–ĺ–Ĺ—Ā—ā—Ä–ł—Ä—É—é—ā: –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑ —É—Ā–Ņ–Ķ—ą–Ĺ–ĺ –Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł—Ä—É–Ķ—ā—Ā—Ź –ł¬†–Ľ–Ķ—á–ł—ā—Ā—Ź —Ā–ĺ—á–Ķ—ā–į–Ĺ–Ĺ—č–ľ –Ņ—Ä–ł–Ķ–ľ–ĺ–ľ –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D3, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –∑–Ĺ–į—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ —É–ľ–Ķ–Ĺ—Ć—ą–į—é—ā —Ä–ł—Ā–ļ¬†–Ņ–Ķ—Ä–Ķ–Ľ–ĺ–ľ–ĺ–≤ –Ī–Ķ–ī—Ä–į [2]. –í¬†—ā–ĺ –∂–Ķ –≤—Ä–Ķ–ľ—Ź –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑ —Ö–į—Ä–į–ļ—ā–Ķ—Ä–ł–∑—É–Ķ—ā—Ā—Ź –Ĺ–į—Ä—É—ą–Ķ–Ĺ–ł—Ź–ľ–ł –Ľ–ł–Ņ–ł–ī–Ĺ–ĺ–≥–ĺ –ĺ–Ī–ľ–Ķ–Ĺ–į –ł¬†–Ĺ–į–ļ–ĺ–Ņ–Ľ–Ķ–Ĺ–ł–Ķ–ľ –Ĺ–Ķ—Ä–į—Ā—ā–≤–ĺ—Ä–ł–ľ–ĺ–≥–ĺ –ĺ—Ā–į–ī–ļ–į –ļ—Ä–ł—Ā—ā–į–Ľ–Ľ–ł—á–Ķ—Ā–ļ–ĺ–≥–ĺ —Ą–ĺ—Ā—Ą–į—ā–į –ļ–į–Ľ—Ć—Ü–ł—Ź –Ĺ–į¬†–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ī–Ľ—Ź—ą–ļ–į—Ö –ī–į–∂–Ķ –Ĺ–į¬†—Ä–į–Ĺ–Ĺ–ł—Ö —Ā—ā–į–ī–ł—Ź—Ö –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ņ–ĺ—Ä–į–∂–Ķ–Ĺ–ł–Ļ [3‚Äď5].
–°–Ľ–Ķ–ī—É–Ķ—ā –ĺ—ā–ľ–Ķ—ā–ł—ā—Ć, —á—ā–ĺ —É¬†–Ľ–ł—Ü –Ņ–ĺ–∂–ł–Ľ–ĺ–≥–ĺ –≤–ĺ–∑—Ä–į—Ā—ā–į –Ņ–嬆–ľ–Ķ—Ä–Ķ —Ä–į–∑–≤–ł—ā–ł—Ź –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–į –ĺ—ā–ľ–Ķ—á–į–Ķ—ā—Ā—Ź –Ņ–į—Ä–į–Ľ–Ľ–Ķ–Ľ—Ć–Ĺ–ĺ–Ķ —É—Ā–ł–Ľ–Ķ–Ĺ–ł–Ķ –Ņ—Ä–ĺ—Ź–≤–Ľ–Ķ–Ĺ–ł–Ļ –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į [6, 7]. –ß–į—Ā—ā–ĺ –Ņ—Ä–ł–Ĺ—Ź—ā–ĺ —Ä–į—Ā—Ā–ľ–į—ā—Ä–ł–≤–į—ā—Ć –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑ –ł¬†–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑ —ɬ†–ĺ–ī–Ĺ–ĺ–≥–ĺ –ł¬†—ā–ĺ–≥–ĺ –∂–Ķ –Ņ–į—Ü–ł–Ķ–Ĺ—ā–į –ļ–į–ļ –Ĺ–Ķ–∑–į–≤–ł—Ā–ł–ľ—č–Ķ –ī—Ä—É–≥ –ĺ—ā¬†–ī—Ä—É–≥–į –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č. –ü—Ä–ł —ć—ā–ĺ–ľ —É–Ņ—É—Ā–ļ–į–Ķ—ā—Ā—Ź –ł–∑¬†–≤–ł–ī—É, —á—ā–ĺ –Ĺ–į¬†—É—Ä–ĺ–≤–Ĺ–Ķ —ā–ļ–į–Ĺ–Ķ–Ļ –ĺ–Ī–į –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–į —Ā—ā–į—Ä—ā—É—é—ā –Ņ—Ä–į–ļ—ā–ł—á–Ķ—Ā–ļ–ł –ĺ–ī–Ĺ–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ –ł¬†–≤—Ā–Ķ–≥–ī–į –≤–ļ–Ľ—é—á–į—é—ā –∑–Ĺ–į—á–ł—ā–Ķ–Ľ—Ć–Ĺ—č–Ķ –Ĺ–į—Ä—É—ą–Ķ–Ĺ–ł—Ź –ĺ–Ī–ľ–Ķ–Ĺ–į –ł¬†–ļ–į–Ľ—Ć—Ü–ł—Ź, –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D3. –ö—Ä—É–Ņ–Ĺ–ĺ–ľ–į—Ā—ą—ā–į–Ī–Ĺ—č–Ķ –ļ–Ľ–ł–Ĺ–ł—á–Ķ—Ā–ļ–ł–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź [8‚Äď10], –≤¬†—ā–ĺ–ľ —á–ł—Ā–Ľ–Ķ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č 25-–Ľ–Ķ—ā–Ĺ–ł—Ö –Ĺ–į–Ī–Ľ—é–ī–Ķ–Ĺ–ł–Ļ –∑–į –§—Ä–į–ľ–ł–Ĺ–≥–Ķ–ľ—Ā–ļ–ĺ–Ļ –ļ–ĺ–≥–ĺ—Ä—ā–ĺ–Ļ [10], —É–ļ–į–∑—č–≤–į—é—ā –Ĺ–į¬†–Ĺ–Ķ–ĺ—Ā–Ņ–ĺ—Ä–ł–ľ—É—é —Ā–≤—Ź–∑—Ć –ľ–Ķ–∂–ī—É –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–Ķ–Ļ –į—Ä—ā–Ķ—Ä–ł–Ļ –ł¬†–ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–ĺ–ľ –ļ–į–ļ —ɬ†–∂–Ķ–Ĺ—Č–ł–Ĺ, —ā–į–ļ –ł¬†–ľ—É–∂—á–ł–Ĺ. –Ě–į–Ņ—Ä–ł–ľ–Ķ—Ä, –ļ—Ä—É–Ņ–Ĺ–ĺ–ľ–į—Ā—ą—ā–į–Ī–Ĺ–ĺ–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ —ā–ĺ–Ľ—Č–ł–Ĺ—č —Ā—ā–Ķ–Ĺ–ļ–ł –ł–Ĺ—ā–ł–ľ—č –ł¬†–Ņ–Ľ–ĺ—ā–Ĺ–ĺ—Ā—ā–ł –Ľ—é–ľ–Ī–į—Ä–Ĺ—č—Ö –ļ–ĺ—Ā—ā–Ķ–Ļ —ɬ†2000 —á–Ķ–Ľ–ĺ–≤–Ķ–ļ –Ņ–ĺ–ļ–į–∑–į–Ľ–ĺ, —á—ā–ĺ —ā–ĺ–Ľ—Č–ł–Ĺ–į —Ā—ā–Ķ–Ĺ–ļ–ł –ł–Ĺ—ā–ł–ľ—č (–Ņ–ĺ–ļ–į–∑–į—ā–Ķ–Ľ—Ć –Ņ—Ä–ĺ–≥—Ä–Ķ—Ā—Ā–ł–ł –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į) –ĺ–Ī—Ä–į—ā–Ĺ–ĺ –Ņ—Ä–ĺ–Ņ–ĺ—Ä—Ü–ł–ĺ–Ĺ–į–Ľ—Ć–Ĺ–į –Ņ–Ľ–ĺ—ā–Ĺ–ĺ—Ā—ā–ł –ļ–ĺ—Ā—ā–Ķ–Ļ (–ĺ—ā—Ä–į–∂–į—é—Č–Ķ–Ļ —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ķ –ļ–į–Ľ—Ć—Ü–ł–Ķ–≤–ĺ–≥–ĺ –ī–Ķ–Ņ–ĺ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į, p < 0,02) [9].
–í¬†–ī–į–Ĺ–Ĺ–ĺ–Ļ —Ā—ā–į—ā—Ć–Ķ –≤–ĺ–Ņ—Ä–ĺ—Ā –嬆–≤–∑–į–ł–ľ–ĺ—Ā–≤—Ź–∑–ł –ļ–į–Ľ—Ć—Ü–ł—Ź, –≤–ł—ā–į–ľ–ł–Ĺ–į D3, –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–į –ł¬†–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į —Ä–į—Ā—Ā–ľ–į—ā—Ä–ł–≤–į–Ķ—ā—Ā—Ź –Ĺ–į¬†–ĺ—Ā–Ĺ–ĺ–≤–Ķ —Ą—É–Ĺ–ī–į–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ—č—Ö –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ –≤¬†–ĺ–Ī–Ľ–į—Ā—ā–ł –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ—Ź—Ä–Ĺ–ĺ–Ļ –Ī–ł–ĺ–Ľ–ĺ–≥–ł–ł –ł¬†–Ī–ł–ĺ—Ö–ł–ľ–ł–ł —Ā¬†–ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–ł–Ķ–ľ –ī–į–Ĺ–Ĺ—č—Ö —ć–Ņ–ł–ī–Ķ–ľ–ł–ĺ–Ľ–ĺ–≥–ł–ł, –ļ–Ľ–ł–Ĺ–ł—á–Ķ—Ā–ļ–ĺ–Ļ –ł¬†—ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ą–į—Ä–ľ–į–ļ–ĺ–Ľ–ĺ–≥–ł–ł –ł¬†–ī–ĺ–ļ–į–∑–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –ľ–Ķ–ī–ł—Ü–ł–Ĺ—č.
–ö–į–Ľ—Ć—Ü–ł–Ļ –ł¬†–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑
–ö–į–ļ –Ľ—é–Ī–ĺ–Ļ –ľ–ł–ļ—Ä–ĺ–Ĺ—É—ā—Ä–ł–Ķ–Ĺ—ā, –ļ–į–Ľ—Ć—Ü–ł–Ļ –Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ—É –≤¬†—Ā—ā—Ä–ĺ–≥–ĺ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ–ĺ–ľ –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–Ķ, –≤¬†—Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö –ī–ĺ–∑–į—Ö. –ė–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł–Ķ —Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł –Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ–ĺ–≥–ĺ –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–į –Ņ–ĺ—ā—Ä–Ķ–Ī–Ľ–Ķ–Ĺ–ł—Ź –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–Ķ–≥–ĺ —Ā–ł–Ĺ–Ķ—Ä–≥–ł—Ā—ā–į –≤–ł—ā–į–ľ–ł–Ĺ–į D3¬†‚Äď –ļ–į–ļ —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł–Ķ, —ā–į–ļ –ł¬†—É–≤–Ķ–Ľ–ł—á–Ķ–Ĺ–ł–Ķ¬†‚Äď –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†–Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł–ł. –ė–ľ–Ķ–Ĺ–Ĺ–ĺ –Ņ–ĺ—ć—ā–ĺ–ľ—É —Ā¬†—ā–ĺ—á–ļ–ł –∑—Ä–Ķ–Ĺ–ł—Ź —Ä–ł—Ā–ļ–į —Ä–į–∑–≤–ł—ā–ł—Ź –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –ļ–į–Ľ—Ć—Ü–ł–Ļ, –Ĺ–Ķ—Ā–ĺ–ľ–Ĺ–Ķ–Ĺ–Ĺ–ĺ, –ł–≥—Ä–į–Ķ—ā –ī–≤–ĺ–Ļ—Ā—ā–≤–Ķ–Ĺ–Ĺ—É—é —Ä–ĺ–Ľ—Ć.
–°¬†–ĺ–ī–Ĺ–ĺ–Ļ —Ā—ā–ĺ—Ä–ĺ–Ĺ—č, –Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ—č–Ķ —É—Ä–ĺ–≤–Ĺ–ł —Ą–ĺ—Ā—Ą–į—ā–į –ł¬†–ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†–Ņ–Ľ–į–∑–ľ–Ķ –ļ—Ä–ĺ–≤–ł —É–≤–Ķ–Ľ–ł—á–ł–≤–į—é—ā —Ä–ł—Ā–ļ¬†–ė–Ď–°, –ł–Ĺ—Ā—É–Ľ—Ć—ā–į –ł¬†—Ā–ľ–Ķ—Ä—ā–Ĺ–ĺ—Ā—ā–ł. –ė—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ ARIC (Atherosclerosis Risk in Communities), –Ņ—Ä–ĺ–≤–ĺ–ī–ł–≤—ą–Ķ–Ķ—Ā—Ź –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ī–ĺ–Ľ–Ķ–Ķ 10 –Ľ–Ķ—ā –ł¬†–≤–ļ–Ľ—é—á–ł–≤—ą–Ķ–Ķ –Ĺ–į–Ī–Ľ—é–ī–Ķ–Ĺ–ł—Ź –∑–į –ļ–ĺ–≥–ĺ—Ä—ā–ĺ–Ļ –ł–∑¬†15¬†700 —á–Ķ–Ľ–ĺ–≤–Ķ–ļ, –Ņ–ĺ–ī—ā–≤–Ķ—Ä–ī–ł–Ľ–ĺ, —á—ā–ĺ –Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ—č–Ķ —É—Ä–ĺ–≤–Ĺ–ł –ļ–į–Ľ—Ć—Ü–ł—Ź (–ě–® 1,16; 95% –Ē–ė 1,07‚Äď1,26, p = 0,0005) –ł¬†—Ą–ĺ—Ā—Ą–į—ā–į (–ě–® 1,11; 95% –Ē–ė 1,02‚Äď1,21; p = 0,02) –≤¬†–Ņ–Ľ–į–∑–ľ–Ķ —Ź–≤–Ľ—Ź—é—ā—Ā—Ź —Ą–į–ļ—ā–ĺ—Ä–į–ľ–ł —Ä–ł—Ā–ļ–į —ć—ā–ł—Ö –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ [11]. –ö–į–ļ –Ņ—Ä–į–≤–ł–Ľ–ĺ, –≤¬†—ć–Ņ–ł–ī–Ķ–ľ–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź—Ö —Ā—ā–Ķ–Ņ–Ķ–Ĺ—Ć –ļ–ĺ—Ä–ĺ–Ĺ–į—Ä–Ĺ–ĺ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ—Ź–Ķ—ā—Ā—Ź –Ņ—Ä–ł –ľ–Ĺ–ĺ–≥–ĺ—Ā–Ľ–ĺ–Ļ–Ĺ–ĺ–Ļ —ā–ĺ–ľ–ĺ–≥—Ä–į—Ą–ł–ł –ļ–į–ļ —Ā—É–ľ–ľ–į –Ī–į–Ľ–Ľ–ĺ–≤ –Ņ–嬆—ą–ļ–į–Ľ–Ķ CAC (coronary artery calcium). –†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č —ć–Ņ–ł–ī–Ķ–ľ–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ –Ņ–ĺ–ļ–į–∑—č–≤–į—é—ā, —á—ā–ĺ –Ī–ĺ–Ľ–Ķ–Ķ –≤—č—Ā–ĺ–ļ–ł–Ķ –∑–Ĺ–į—á–Ķ–Ĺ–ł—Ź —ć—ā–ĺ–≥–ĺ –Ņ–į—Ä–į–ľ–Ķ—ā—Ä–į —Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤—É—é—ā –Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ–ĺ–ľ—É —Ä–ł—Ā–ļ—É –ė–Ď–° [12, 13].
–°¬†–ī—Ä—É–≥–ĺ–Ļ —Ā—ā–ĺ—Ä–ĺ–Ĺ—č, —ā–Ķ –∂–Ķ –ļ—Ä—É–Ņ–Ĺ–ĺ–ľ–į—Ā—ą—ā–į–Ī–Ĺ—č–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤—É—é—ā: –ł–∑–Ī—č—ā–ĺ–ļ –ļ–į–Ľ—Ć—Ü–ł—Ź¬†‚Äď –Ĺ–Ķ¬†–Ķ–ī–ł–Ĺ—Ā—ā–≤–Ķ–Ĺ–Ĺ—č–Ļ —Ą–į–ļ—ā–ĺ—Ä —Ä–ł—Ā–ļ–į –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į [14, 15]. –ė—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ MESA (Multi-Ethnic Study of Atherosclerosis) [14] –≤–ļ–Ľ—é—á–ł–Ľ–ĺ –Ĺ–į–Ī–Ľ—é–ī–Ķ–Ĺ–ł—Ź –∑–į –ļ–ĺ–≥–ĺ—Ä—ā–ĺ–Ļ –ł–∑¬†5880 —É—á–į—Ā—ā–Ĺ–ł–ļ–ĺ–≤ 45‚Äď84 –Ľ–Ķ—ā, –ļ–į–∂–ī—č–Ļ –ł–∑¬†–ļ–ĺ—ā–ĺ—Ä—č—Ö –Ņ—Ä–ĺ—ą–Ķ–Ľ –ļ–ĺ–ľ–Ņ—Ć—é—ā–Ķ—Ä–Ĺ—É—é —ā–ĺ–ľ–ĺ–≥—Ä–į—Ą–ł—é –į–ĺ—Ä—ā—č. –§–į–ļ—ā–ĺ—Ä–į–ľ–ł —Ä–ł—Ā–ļ–į –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į–ĺ—Ä—ā—č —Ź–≤–Ľ—Ź–Ľ–ł—Ā—Ć –ł–Ĺ–ī–Ķ–ļ—Ā –ľ–į—Ā—Ā—č —ā–Ķ–Ľ–į, –ļ—É—Ä–Ķ–Ĺ–ł–Ķ, –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–ł–Ķ –į–Ĺ—ā–ł–Ľ–ł–Ņ–ł–ī–Ĺ—č—Ö –ł¬†–į–Ĺ—ā–ł–≥–ł–Ņ–Ķ—Ä—ā–ĺ–Ĺ–ł—á–Ķ—Ā–ļ–ł—Ö –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤. –Ē–Ķ–Ļ—Ā—ā–≤–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ, —Ā—ā–Ķ–Ņ–Ķ–Ĺ—Ć –Ĺ–į–ļ–ĺ–Ņ–Ľ–Ķ–Ĺ–ł—Ź –ļ–į–Ľ—Ć—Ü–ł—Ź –Ĺ–į¬†—Ā—ā–Ķ–Ĺ–ļ–į—Ö –į–ĺ—Ä—ā—č –ļ–ĺ—Ä—Ä–Ķ–Ľ–ł—Ä–ĺ–≤–į–Ľ–į —Ā–嬆—Ā—ā–Ķ–Ņ–Ķ–Ĺ—Ć—é —ā—Ź–∂–Ķ—Ā—ā–ł —Ā—ā–Ķ–Ĺ–ĺ–∑–į (p < 0,001). –ě–ī–Ĺ–į–ļ–ĺ —É¬†13‚Äď18% –Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ –Ī—č–Ľ–ł —ā—Ź–∂–Ķ–Ľ—č–Ķ —Ą–ĺ—Ä–ľ—č —Ā—ā–Ķ–Ĺ–ĺ–∑–į (–Ī–ĺ–Ľ–Ķ–Ķ 75%) –ł¬†–ļ—Ä–į–Ļ–Ĺ–Ķ –Ĺ–ł–∑–ļ–į—Ź —Ā—ā–Ķ–Ņ–Ķ–Ĺ—Ć –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į–ĺ—Ä—ā—č [15]. –Ę–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, —Ā–≤—Ź–∑—Ć –ľ–Ķ–∂–ī—É —ā—Ź–∂–Ķ—Ā—ā—Ć—é —Ā—ā–Ķ–Ĺ–ĺ–∑–į –ł¬†–Ĺ–į–ļ–ĺ–Ņ–Ľ–Ķ–Ĺ–ł–Ķ–ľ –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ī–Ľ—Ź—ą–ļ–į—Ö –ī–į–Ľ–Ķ–ļ–ĺ –Ĺ–Ķ¬†–ĺ—á–Ķ–≤–ł–ī–Ĺ–į.
–ó–ī–Ķ—Ā—Ć –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ—Ź–Ķ—ā—Ā—Ź —É–ľ–Ķ—Ā—ā–Ĺ—č–ľ –Ĺ–į–Ņ–ĺ–ľ–Ĺ–ł—ā—Ć –ł—Ā—ā–ĺ—Ä–ł—é –Ņ—Ä–ł–ľ–Ķ–Ĺ–Ķ–Ĺ–ł—Ź –į–Ĺ—ā–ł–ļ–į–Ľ—Ć—Ü–ł–Ķ–≤—č—Ö –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤ –Ņ—Ä–ł –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–Ķ. –Ē–ĺ 2003¬†–≥.¬†–≤ –∑–į–Ņ–į–ī–Ĺ–ĺ–Ļ –Ľ–ł—ā–Ķ—Ä–į—ā—É—Ä–Ķ –Ī—č–Ľ–ĺ –ľ–ĺ–ī–Ŗ嬆–≥–ĺ–≤–ĺ—Ä–ł—ā—Ć –嬆–Ņ–Ķ—Ä—Ā–Ņ–Ķ–ļ—ā–ł–≤–į—Ö –Ņ—Ä–ł–ľ–Ķ–Ĺ–Ķ–Ĺ–ł—Ź –į–Ĺ—ā–ł–ļ–į–Ľ—Ć—Ü–ł–Ķ–≤—č—Ö –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤ –ļ–į–ļ –Ņ–ĺ—ā–Ķ–Ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ –į–Ĺ—ā–ł–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö. –í¬†—Ā–į–ľ–ĺ–ľ –ī–Ķ–Ľ–Ķ, —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –Ī–ĺ–Ľ—Ć—ą–ł—Ö –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ REGRESS (Regression Growth Evaluation Statin Study), VHAS (Verapamil in Hypertension and Atherosclerosis Study) –ł¬†–ī—Ä. —É–ļ–į–∑–į–Ľ–ł –Ĺ–į¬†–≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ—Ā—ā—Ć —ā–ĺ–≥–ĺ, —á—ā–ĺ –į–Ĺ—ā–į–≥–ĺ–Ĺ–ł—Ā—ā—č –ļ–į–Ľ—Ć—Ü–ł—Ź –ĺ–Ī–Ľ–į–ī–į—é—ā –Ņ–ĺ—ā–Ķ–Ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–ľ –į–Ĺ—ā–ł–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł–ľ —ć—Ą—Ą–Ķ–ļ—ā–ĺ–ľ [16]. –Ę–Ķ–ľ –Ĺ–Ķ¬†–ľ–Ķ–Ĺ–Ķ–Ķ —É–∂–Ķ —Ā¬†2004¬†–≥.¬†–ī–ł—Ā–ļ—É—Ā—Ā–ł–ł –ĺ–Ī¬†–į–Ĺ—ā–ł–ļ–į–Ľ—Ć—Ü–ł–Ķ–≤—č—Ö –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–į—Ö –ł¬†–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–Ķ –Ĺ–Ķ–ĺ–∂–ł–ī–į–Ĺ–Ĺ–ĺ –Ņ—Ä–Ķ–ļ—Ä–į—ā–ł–Ľ–ł—Ā—Ć. –í–ĺ–∑–ľ–ĺ–∂–Ĺ—č–ľ –ĺ–Ī—ä—Ź—Ā–Ĺ–Ķ–Ĺ–ł–Ķ–ľ —ć—ā–ĺ–≥–ĺ —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –ĺ—Ā–ĺ–∑–Ĺ–į–Ĺ–ł–Ķ –ī–į–≤–Ĺ–ĺ –ł–∑–≤–Ķ—Ā—ā–Ĺ–ĺ–≥–ĺ –≤¬†—Ą–į—Ä–ľ–į–ļ–ĺ–Ľ–ĺ–≥–ł–ł —Ą–į–ļ—ā–į: –į–Ĺ—ā–ł–ļ–į–Ľ—Ć—Ü–ł–Ķ–≤—č–Ķ –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā—č —Ź–≤–Ľ—Ź—é—ā—Ā—Ź –į–Ĺ–į–Ľ–ĺ–≥–į–ľ–ł —Ā—ā–į—ā–ł–Ĺ–ĺ–≤. –°—ā–į—ā–ł–Ĺ—č –ł–Ĺ–≥–ł–Ī–ł—Ä—É—é—ā HMG-CoA —Ä–Ķ–ī—É–ļ—ā–į–∑—É (3-–≥–ł–ī—Ä–ĺ–ļ—Ā–ł-3-–ľ–Ķ—ā–ł–Ľ-–≥–Ľ—é—ā–į—Ä–ł–Ľ-–ļ–ĺ—Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā –ź¬†—Ä–Ķ–ī—É–ļ—ā–į–∑—É)¬†‚Äď —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā —Ā–ł–Ĺ—ā–Ķ–∑–į —ć–Ĺ–ī–ĺ–≥–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ —Ö–ĺ–Ľ–Ķ—Ā—ā–Ķ—Ä–ł–Ĺ–į, –Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ–ĺ–≥–ĺ –ī–Ľ—Ź —Ā–ł–Ĺ—ā–Ķ–∑–į —Ā—ā–Ķ—Ä–ĺ–ł–ī–Ĺ—č—Ö¬†–≥–ĺ—Ä–ľ–ĺ–Ĺ–ĺ–≤ –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D [17]. –ü–ĺ—Ā–ļ–ĺ–Ľ—Ć–ļ—É –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź –į–Ĺ—ā–ł–ļ–į–Ľ—Ć—Ü–ł–Ķ–≤—č—Ö –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤ —Ā¬†HMG-CoA —Ä–Ķ–ī—É–ļ—ā–į–∑–ĺ–Ļ –Ĺ–Ķ—Ā–Ņ–Ķ—Ü–ł—Ą–ł—á–Ĺ—č, –Ī–Ľ–ĺ–ļ–į—ā–ĺ—Ä—č –ļ–į–Ľ—Ć—Ü–ł–Ķ–≤—č—Ö –ļ–į–Ĺ–į–Ľ–ĺ–≤ –∑–į–≤–Ķ–ī–ĺ–ľ–ĺ —Ź–≤–Ľ—Ź—é—ā—Ā—Ź¬†–≥–ĺ—Ä–į–∑–ī–ĺ –Ī–ĺ–Ľ–Ķ–Ķ —Ā–Ľ–į–Ī—č–ľ–ł –į–Ĺ—ā–ł–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł–ľ–ł –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–į–ľ–ł, —á–Ķ–ľ —Ā—ā–į—ā–ł–Ĺ—č. –Ę–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, –į–Ĺ—ā–ł–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ĺ–Ķ –ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ –į–Ĺ—ā–ł–ļ–į–Ľ—Ć—Ü–ł–Ķ–≤—č—Ö –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤ –Ĺ–Ķ¬†–ł–ľ–Ķ–Ķ—ā –Ĺ–ł—á–Ķ–≥–ĺ –ĺ–Ī—Č–Ķ–≥–ĺ —Ā¬†–≥–ĺ–ľ–Ķ–ĺ—Ā—ā–į–∑–ĺ–ľ –ļ–į–Ľ—Ć—Ü–ł—Ź.
–Ě–Ķ¬†—Ā–Ľ–Ķ–ī—É–Ķ—ā —ā–į–ļ–∂–Ķ –∑–į–Ī—č–≤–į—ā—Ć –嬆—ā–ĺ–ľ, —á—ā–ĺ –ļ–į–Ľ—Ć—Ü–ł–Ļ¬†‚Äď —ć—Ā—Ā–Ķ–Ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–Ļ —ć–Ľ–Ķ–ľ–Ķ–Ĺ—ā. –ö–į–Ľ—Ć—Ü–ł–Ļ –Ĺ–Ķ¬†—ā–ĺ–Ľ—Ć–ļ–ĺ —É—á–į—Ā—ā–≤—É–Ķ—ā –≤¬†–ľ–Ķ—ā–į–Ī–ĺ–Ľ–ł–∑–ľ–Ķ –ļ–ĺ—Ā—ā–Ķ–Ļ, —Ā–ĺ–ļ—Ä–į—Č–Ķ–Ĺ–ł–ł –ľ—č—ą—Ü, —Ā–≤–Ķ—Ä—ā—č–≤–į–Ĺ–ł–ł –ļ—Ä–ĺ–≤–ł. –ö–į–Ľ—Ć—Ü–ł–Ļ —ā–į–ļ–∂–Ķ –ĺ–Ī—Ź–∑–į—ā–Ķ–Ľ—Ć–Ĺ—č–Ļ –ļ–ĺ–ľ–Ņ–ĺ–Ĺ–Ķ–Ĺ—ā (—ā–į–ļ –Ĺ–į–∑—č–≤–į–Ķ–ľ—č–Ļ ¬ę–≤—ā–ĺ—Ä–ł—á–Ĺ—č–Ļ –ľ–Ķ—Ā—Ā–Ķ–Ĺ–ī–∂–Ķ—ĬĽ) –ľ–Ĺ–ĺ–≥–ĺ—á–ł—Ā–Ľ–Ķ–Ĺ–Ĺ—č—Ö –≤–Ĺ—É—ā—Ä–ł–ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č—Ö —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ—č—Ö –ļ–į—Ā–ļ–į–ī–ĺ–≤, –Ņ–ĺ—Ā—Ä–Ķ–ī—Ā—ā–≤–ĺ–ľ –ļ–ĺ—ā–ĺ—Ä—č—Ö —ć–Ĺ–ī–ĺ—ā–Ķ–Ľ–ł–į–Ľ—Ć–Ĺ—č–Ķ –ļ–Ľ–Ķ—ā–ļ–ł —Ä–Ķ–į–≥–ł—Ä—É—é—ā –Ĺ–į¬†–≤–Ĺ–Ķ—ą–Ĺ–ł–Ķ —Ā—ā–ł–ľ—É–Ľ—č. –ö–į–Ľ—Ć—Ü–ł–Ļ —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –≤–į–∂–Ĺ—č–ľ –ľ–Ķ–ī–ł–į—ā–ĺ—Ä–ĺ–ľ —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ—č—Ö –Ņ—É—ā–Ķ–Ļ –ļ–į–ļ –ľ–ł—ā–ĺ–≥–Ķ–Ĺ–Ķ–∑–į, —ā–į–ļ –ł¬†–į–Ņ–ĺ–Ņ—ā–ĺ–∑–į [18]. –≠—ā–ł —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ—č–Ķ –Ņ—É—ā–ł –ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ —Ā–Ľ–ĺ–∂–Ĺ—č, –≤–ĺ–≤–Ľ–Ķ–ļ–į—é—ā –ľ–Ĺ–ĺ–∂–Ķ—Ā—ā–≤–ĺ –Ī–Ķ–Ľ–ļ–ĺ–≤-–Ņ–ĺ—Ā—Ä–Ķ–ī–Ĺ–ł–ļ–ĺ–≤ –ł¬†–ī–Ķ–Ļ—Ā—ā–≤—É—é—ā —á–Ķ—Ä–Ķ–∑ —Ā–ł—Ā—ā–Ķ–ľ—É Ca2+-–∑–į–≤–ł—Ā–ł–ľ–ĺ–≥–ĺ –Ī–Ķ–Ľ–ļ–į –ļ–į–Ľ—Ć–ľ–ĺ–ī—É–Ľ–ł–Ĺ–į. –ė—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź –Ņ–ĺ–ļ–į–∑–į–Ľ–ł, —á—ā–ĺ —Ä–ł—Ā–ļ¬†—Ā–ĺ—Ā—É–ī–ł—Ā—ā—č—Ö –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ —É–≤–Ķ–Ľ–ł—á–ł–≤–į–Ķ—ā—Ā—Ź –ł¬†–Ņ—Ä–ł –Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ—č—Ö, –ł¬†–Ņ—Ä–ł –Ĺ–ł–∑–ļ–ł—Ö —É—Ä–ĺ–≤–Ĺ—Ź—Ö —ć–Ĺ–ī–ĺ—ā–Ķ–Ľ–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ļ–į–Ľ—Ć—Ü–ł—Ź [19].
–í–ł—ā–į–ľ–ł–Ĺ D –ł¬†–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑
–í–ł—ā–į–ľ–ł–Ĺ D¬†‚Äď –ĺ–ī–ł–Ĺ –ł–∑¬†–≥–Ľ–į–≤–Ĺ—č—Ö –Ņ–ĺ—Ā—Ä–Ķ–ī–Ĺ–ł–ļ–ĺ–≤ –ļ–į–Ľ—Ć—Ü–ł–Ķ–≤–ĺ–≥–ĺ –ľ–Ķ—ā–į–Ī–ĺ–Ľ–ł–∑–ľ–į. –Ė–ł—Ä–ĺ—Ä–į—Ā—ā–≤–ĺ—Ä–ł–ľ—č–Ķ –≤–ł—ā–į–ľ–ł–ŗ謆–≥—Ä—É–Ņ–Ņ—č D —Ā–Ĺ–į—á–į–Ľ–į –Ņ—Ä–Ķ–ĺ–Ī—Ä–į–∑—É—é—ā—Ā—Ź –≤¬†–į–ļ—ā–ł–≤–Ĺ—É—é —Ą–ĺ—Ä–ľ—É –Ņ–ĺ—Ā—Ä–Ķ–ī—Ā—ā–≤–ĺ–ľ –Ī–ł–ĺ—ā—Ä–į–Ĺ—Ā—Ą–ĺ—Ä–ľ–į—Ü–ł–Ļ, –į¬†–∑–į—ā–Ķ–ľ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤—É—é—ā —Ā–嬆—Ā–Ņ–Ķ—Ü–ł—Ą–ł—á–Ķ—Ā–ļ–ł–ľ–ł —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–į–ľ–ł. –ě—Ā–Ĺ–ĺ–≤–Ĺ—č–Ķ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č –Ī–ł–ĺ—ā—Ä–į–Ĺ—Ā—Ą–ĺ—Ä–ľ–į—Ü–ł–ł –≤–ł—ā–į–ľ–ł–Ĺ–į D –Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī—Ź—ā –≤¬†–ļ–ĺ–∂–Ķ, –Ņ–Ķ—á–Ķ–Ĺ–ł –ł¬†–Ņ–ĺ—á–ļ–į—Ö. –í¬†–ļ–ĺ–∂–Ķ –Ņ–ĺ–ī –ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ–ľ —É–Ľ—Ć—ā—Ä–į—Ą–ł–ĺ–Ľ–Ķ—ā–ĺ–≤–ĺ–≥–ĺ –ĺ–Ī–Ľ—É—á–Ķ–Ĺ–ł—Ź –ĺ–Ī—Ä–į–∑—É–Ķ—ā—Ā—Ź –≤–ł—ā–į–ľ–ł–Ĺ D3. –í¬†–Ņ–Ķ—á–Ķ–Ĺ–ł –≤–ł—ā–į–ľ–ł–Ĺ D3 –Ņ—É—ā–Ķ–ľ¬†–≥–ł–ī—Ä–ĺ–ļ—Ā–ł–Ľ–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –Ņ—Ä–Ķ–≤—Ä–į—Č–į–Ķ—ā—Ā—Ź –≤¬†25-–ĺ–ļ—Ā–ł—Ö–ĺ–Ľ–Ķ–ļ–į–Ľ—Ć—Ü–ł—Ą–Ķ—Ä–ĺ–Ľ (25-–ě–Ě-D3) –Ņ—Ä–ł –Ņ–ĺ—Ā—Ä–Ķ–ī—Ā—ā–≤–Ķ 25-–≥–ł–ī—Ä–ĺ–ļ—Ā–ł–Ľ–į–∑—č. –í¬†–Ņ–ĺ—á–ļ–į—Ö, –Ņ—Ä–ł –ł—Ö –Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ–ľ —Ą—É–Ĺ–ļ—Ü–ł–ĺ–Ĺ–ł—Ä–ĺ–≤–į–Ĺ–ł–ł, 25-–ě–Ě-D3 —ā—Ä–į–Ĺ—Ā—Ą–ĺ—Ä–ľ–ł—Ä—É–Ķ—ā—Ā—Ź –≤¬†1,25-–ī–ł–ĺ–ļ—Ā–ł—Ö–ĺ–Ľ–Ķ–ļ–į–Ľ—Ć—Ü–ł—Ą–Ķ—Ä–ĺ–Ľ (1,25-(–ě–Ě)D3), –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –į–ļ—ā–ł–≤–Ĺ—É—é —Ą–ĺ—Ä–ľ—É –≤–ł—ā–į–ľ–ł–Ĺ–į, –ļ–ĺ—ā–ĺ—Ä–į—Ź –Ņ–Ķ—Ä–Ķ–Ĺ–ĺ—Ā–ł—ā—Ā—Ź –≤¬†–ļ—Ä–ĺ–≤—Ź–Ĺ–ĺ–ľ —Ä—É—Ā–Ľ–Ķ –≤–ł—ā–į–ľ–ł–Ĺ-D-—Ā–≤—Ź–∑—č–≤–į—é—Č–ł–ľ –Ī–Ķ–Ľ–ļ–ĺ–ľ (VDBP). –Ď–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ĺ–Ķ –ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ –į–ļ—ā–ł–≤–Ĺ–ĺ–Ļ —Ą–ĺ—Ä–ľ—č –≤–ł—ā–į–ľ–ł–Ĺ–į —Ä–Ķ–į–Ľ–ł–∑—É–Ķ—ā—Ā—Ź –Ņ—É—ā–Ķ–ľ —Ā–≤—Ź–∑—č–≤–į–Ĺ–ł—Ź —Ā¬†—Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–ĺ–ľ –≤–ł—ā–į–ľ–ł–Ĺ–į D (VDR). –†–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä –≤–ł—ā–į–ľ–ł–Ĺ–į D, –Ņ–ĺ–ī–ĺ–Ī–Ĺ–ĺ —ć—Ā—ā—Ä–ĺ–≥–Ķ–Ĺ-—Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–į–ľ, —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź —Ą–į–ļ—ā–ĺ—Ä–ĺ–ľ —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł–ł, –ļ–ĺ—ā–ĺ—Ä—č–Ļ —Ä–Ķ–≥—É–Ľ–ł—Ä—É–Ķ—ā —Ā–ł–Ĺ—ā–Ķ–∑ –Ī–Ķ–Ľ–ļ–ĺ–≤, –≤–ĺ–≤–Ľ–Ķ—á–Ķ–Ĺ–Ĺ—č—Ö –≤¬†–≥–ĺ–ľ–Ķ–ĺ—Ā—ā–į–∑ –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†—Ą–ĺ—Ā—Ą–ĺ—Ä–į [20, 21].
–≠–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ—č–Ķ –ī–į–Ĺ–Ĺ—č–Ķ –Ņ–ĺ–ļ–į–∑—č–≤–į—é—ā, —á—ā–ĺ —Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —ć—Ą—Ą–Ķ–ļ—ā—č –≤–ł—ā–į–ľ–ł–Ĺ–į D –≤–ļ–Ľ—é—á–į—é—ā —ā–ĺ—Ä–ľ–ĺ–∂–Ķ–Ĺ–ł–Ķ —Ā–Ķ–ļ—Ä–Ķ—Ü–ł–ł –Ņ—Ä–ĺ–≤–ĺ—Ā–Ņ–į–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö —Ü–ł—ā–ĺ–ļ–ł–Ĺ–ĺ–≤, –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ –į–ī–≥–Ķ–∑–ł–ł –ł¬†–Ņ—Ä–ĺ–Ľ–ł—Ą–Ķ—Ä–į—Ü–ł—é —Ā–ĺ—Ā—É–ī–ł—Ā—ā—č—Ö¬†–≥–Ľ–į–ī–ļ–ĺ–ľ—č—ą–Ķ—á–Ĺ—č—Ö –ļ–Ľ–Ķ—ā–ĺ–ļ¬†‚Äď –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ł–ľ–Ķ—é—ā –≤–į–∂–Ĺ–ĺ–Ķ –∑–Ĺ–į—á–Ķ–Ĺ–ł–Ķ –ī–Ľ—Ź –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ [22].
–Ē–į–Ĺ–Ĺ—č–Ķ —ć–Ņ–ł–ī–Ķ–ľ–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ –ļ–ĺ–≥–ĺ—Ä—ā –Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ –ł¬†–∑–ī–ĺ—Ä–ĺ–≤—č—Ö –ī–ĺ–Ĺ–ĺ—Ä–ĺ–≤ —Ā–≤–ł–ī–Ķ—ā–Ķ–Ľ—Ć—Ā—ā–≤—É—é—ā: –ī–Ķ—Ą–ł—Ü–ł—ā –≤–ł—ā–į–ľ–ł–Ĺ–į D –į—Ā—Ā–ĺ—Ü–ł–ł—Ä—É–Ķ—ā—Ā—Ź —Ā¬†–Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ—č–ľ –į—Ä—ā–Ķ—Ä–ł–į–Ľ—Ć–Ĺ—č–ľ –ī–į–≤–Ľ–Ķ–Ĺ–ł–Ķ–ľ, –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–ĺ–ľ, —Ā–į—Ö–į—Ä–Ĺ—č–ľ –ī–ł–į–Ī–Ķ—ā–ĺ–ľ –ł¬†–į–ļ—ā–ł–≤–į—Ü–ł–Ķ–Ļ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤ –≤–ĺ—Ā–Ņ–į–Ľ–Ķ–Ĺ–ł—Ź [23]. –Ě–į–Ņ—Ä–ł–ľ–Ķ—Ä, –Ņ—Ä–ł –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–ł 654 –Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ 55‚Äď96 –Ľ–Ķ—ā (—Ā—Ä–Ķ–ī–Ĺ–ł–Ļ –≤–ĺ–∑—Ä–į—Ā—ā 75,5 –Ľ–Ķ—ā) –Ī–Ķ–∑ –ė–Ď–° –ł¬†–ł–Ĺ—Ā—É–Ľ—Ć—ā–į –≤¬†–į–Ĺ–į–ľ–Ĺ–Ķ–∑–Ķ —É—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ĺ, —á—ā–ĺ —ā–ĺ–Ľ—Č–ł–Ĺ–į –ł–Ĺ—ā–ł–ľ—č —Ā–ĺ–Ĺ–Ĺ–ĺ–Ļ –į—Ä—ā–Ķ—Ä–ł–ł —Ā–Ĺ–ł–∂–į–Ľ–į—Ā—Ć —Ā¬†—É–≤–Ķ–Ľ–ł—á–Ķ–Ĺ–ł–Ķ–ľ –≤¬†–Ņ–Ľ–į–∑–ľ–Ķ —É—Ä–ĺ–≤–Ĺ—Ź 25-(OH)-D3¬†‚Äď –ĺ–ī–Ĺ–ĺ–≥–ĺ –ł–∑¬†–ĺ—Ā–Ĺ–ĺ–≤–Ĺ—č—Ö –ľ–Ķ—ā–į–Ī–ĺ–Ľ–ł—ā–ĺ–≤ –≤–ł—ā–į–ľ–ł–Ĺ–į (—Ä = 0,02) [24]. –í¬†–ļ—Ä—É–Ņ–Ĺ–ĺ–ľ–į—Ā—ą—ā–į–Ī–Ĺ–ĺ–ľ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–ł –ļ–ĺ–≥–ĺ—Ä—ā—č –ł–∑¬†16 600 —á–Ķ–Ľ–ĺ–≤–Ķ–ļ —Ā—ā–į—Ä—ą–Ķ 18 –Ľ–Ķ—ā –Ī—č–Ľ–ĺ —É—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ĺ, —á—ā–ĺ —Ā–Ĺ–ł–∂–Ķ–Ĺ–Ĺ—č–Ķ —É—Ä–ĺ–≤–Ĺ–ł 25-(OH)-D3 —Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–ĺ–≤–į–Ľ–ł –Ņ–ĺ–≤—č—ą–Ķ–Ĺ–ł—é —Ä–ł—Ā–ļ–į —Ā–Ķ—Ä–ī–Ķ—á–Ĺ–ĺ-—Ā–ĺ—Ā—É–ī–ł—Ā—ā—č—Ö –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ (–ě–® 1,2; 95% –Ē–ė 1,01‚Äď1,36; —Ä = 0,03) [25]. –Ē–į–Ĺ–Ĺ–ĺ–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ –ĺ—ā–Ľ–ł—á–į–Ķ—ā—Ā—Ź –ĺ—ā¬†–ī—Ä—É–≥–ł—Ö —ā—Č–į—ā–Ķ–Ľ—Ć–Ĺ—č–ľ –į–Ĺ–į–Ľ–ł–∑–ĺ–ľ –ī–į–Ĺ–Ĺ—č—Ö –ł¬†–ī–ĺ—Ā—ā–ĺ–≤–Ķ—Ä–Ĺ—č–ľ —É—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–ł–Ķ–ľ —ć—Ą—Ą–Ķ–ļ—ā–ĺ–≤ –≤–ł—ā–į–ľ–ł–Ĺ–į D –≤–Ĺ–Ķ –∑–į–≤–ł—Ā–ł–ľ–ĺ—Ā—ā–ł –ĺ—ā¬†–≤–ĺ–∑—Ä–į—Ā—ā–į, –Ņ–ĺ–Ľ–į, —ć—ā–Ĺ–ł—á–Ķ—Ā–ļ–ĺ–Ļ –Ņ—Ä–ł–Ĺ–į–ī–Ľ–Ķ–∂–Ĺ–ĺ—Ā—ā–ł, —Ā–Ķ–∑–ĺ–Ĺ–į, —Ą–ł–∑–ł—á–Ķ—Ā–ļ–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł, –ł–Ĺ–ī–Ķ–ļ—Ā–į –ľ–į—Ā—Ā—č —ā–Ķ–Ľ–į, –ļ—É—Ä–Ķ–Ĺ–ł—Ź,¬†–≥–ł–Ņ–Ķ—Ä—ā–ĺ–Ĺ–ł–ł, –ī–ł–į–Ī–Ķ—ā–į, –Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ—č—Ö —É—Ä–ĺ–≤–Ĺ–Ķ–Ļ —ā—Ä–ł–≥–Ľ–ł—Ü–Ķ—Ä–ł–ī–ĺ–≤, —Ö–ĺ–Ľ–Ķ—Ā—ā–Ķ—Ä–ł–Ĺ–į –ł¬†–Ľ–ł–Ņ–ĺ–Ņ—Ä–ĺ—ā–Ķ–ł–ī–ĺ–≤ –Ĺ–ł–∑–ļ–ĺ–Ļ –Ņ–Ľ–ĺ—ā–Ĺ–ĺ—Ā—ā–ł, —Ö—Ä–ĺ–Ĺ–ł—á–Ķ—Ā–ļ–ł—Ö –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ –Ņ–ĺ—á–Ķ–ļ –ł¬†–ī—Ä.
–≠–Ņ–ł–ī–Ķ–ľ–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ –ī–į–Ĺ–Ĺ—č–Ķ —É–ļ–į–∑—č–≤–į—é—ā –Ĺ–į¬†–ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ —É–∑–ļ–ł–Ļ —Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ļ –ī–ł–į–Ņ–į–∑–ĺ–Ĺ –Ņ–ĺ—ā—Ä–Ķ–Ī–Ľ–Ķ–Ĺ–ł—Ź –≤–ł—ā–į–ľ–ł–Ĺ–į D. –í—č—Ö–ĺ–ī –∑–į —Ä–į–ľ–ļ–ł —ć—ā–ĺ–≥–ĺ –ī–ł–į–Ņ–į–∑–ĺ–Ĺ–į (–ī–Ķ—Ą–ł—Ü–ł—ā –Ľ–ł–Ī–嬆–≥–ł–Ņ–Ķ—Ä–≤–ł—ā–į–ľ–ł–Ĺ–ĺ–∑ D) —É–≤–Ķ–Ľ–ł—á–ł–≤–į–Ķ—ā —Ä–ł—Ā–ļ¬†—Ā–Ķ—Ä–ī–Ķ—á–Ĺ–ĺ-—Ā–ĺ—Ā—É–ī–ł—Ā—ā—č—Ö –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ [26]. –°–Ľ–Ķ–ī—É–Ķ—ā —Ä–į–∑–Ľ–ł—á–į—ā—Ć —Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ (—ā–į–ļ –Ĺ–į–∑—č–≤–į–Ķ–ľ—č–Ķ —Ä–Ķ–ļ–ĺ–ľ–Ķ–Ĺ–ī–ĺ–≤–į–Ĺ–Ĺ—č–Ķ —Ā—É—ā–ĺ—á–Ĺ—č–Ķ) –ī–ĺ–∑—č –≤–ł—ā–į–ľ–ł–Ĺ–į –ł¬†—ā–Ķ—Ä–į–Ņ–Ķ–≤—ā–ł—á–Ķ—Ā–ļ–ĺ–Ķ –Ņ—Ä–ł–ľ–Ķ–Ĺ–Ķ–Ĺ–ł–Ķ –≤–ł—ā–į–ľ–ł–Ĺ–į D3. –ö–į–∂–ī–ĺ–ī–Ĺ–Ķ–≤–Ĺ—č–Ļ –ł¬†–ī–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ—č–Ļ –Ņ—Ä–ł–Ķ–ľ –≤–ł—ā–į–ľ–ł–Ĺ–į D3 –≤¬†—Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö –ī–ĺ–∑–į—Ö —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –Ĺ–ĺ—Ä–ľ–ĺ–Ļ –Ņ–ł—ā–į–Ĺ–ł—Ź –ł¬†–į–Ī—Ā–ĺ–Ľ—é—ā–Ĺ–ĺ –Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ –ī–Ľ—Ź –ī–ĺ–Ľ–≥–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ–Ļ –Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł–ļ–ł —Ā–ĺ—Ā—É–ī–ł—Ā—ā–ĺ–Ļ –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł–ł –ł¬†–ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–į.
–í¬†—ā–ĺ –∂–Ķ –≤—Ä–Ķ–ľ—Ź –Ņ—Ä–ł —ā–Ķ—Ä–į–Ņ–Ķ–≤—ā–ł—á–Ķ—Ā–ļ–ĺ–ľ –Ņ—Ä–ł–Ķ–ľ–Ķ –ļ—Ä–į—ā–ļ–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ—č–ľ–ł –ļ—É—Ä—Ā–į–ľ–ł –ī–ĺ–∑–ł—Ä–ĺ–≤–ļ–į –ľ–ĺ–∂–Ķ—ā –∑–Ĺ–į—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –Ņ—Ä–Ķ–≤—č—ą–į—ā—Ć —Ä–Ķ–ļ–ĺ–ľ–Ķ–Ĺ–ī–ĺ–≤–į–Ĺ–Ĺ—č–Ķ —É—Ä–ĺ–≤–Ĺ–ł –≤¬†200‚Äď600 –ú–ē/—Ā—É—ā [27]. –Ě–į–Ņ—Ä–ł–ľ–Ķ—Ä, –Ņ–ĺ—Ā–Ľ–Ķ —ā—Ä–Ķ—Ö–ľ–Ķ—Ā—Ź—á–Ĺ–ĺ–≥–ĺ –Ņ—Ä–ł–Ķ–ľ–į –≤¬†–ī–ĺ–∑–Ķ 3000 –ú–ē/—Ā—É—ā —Ā¬†—Ü–Ķ–Ľ—Ć—é –ļ–ĺ–ľ–Ņ–Ķ–Ĺ—Ā–į—Ü–ł–ł¬†–≥–ł–Ņ–ĺ–≤–ł—ā–į–ľ–ł–Ĺ–ĺ–∑–į D —ɬ†–Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ –Ĺ–į–Ī–Ľ—é–ī–į–Ľ–ł—Ā—Ć —É–ľ–Ķ–Ĺ—Ć—ą–Ķ–Ĺ–ł–Ķ —Ā—ā–Ķ–Ĺ–ĺ–∑–į –į—Ä—ā–Ķ—Ä–ł–Ļ –ł¬†—É–≤–Ķ–Ľ–ł—á–Ķ–Ĺ–ł–Ķ —É—Ä–ĺ–≤–Ĺ–Ķ–Ļ –Ľ–Ķ–Ņ—ā–ł–Ĺ–į (–≥–ĺ—Ä–ľ–ĺ–Ĺ–į, —Ä–Ķ–≥—É–Ľ–ł—Ä—É—é—Č–Ķ–≥–ĺ –ľ–Ķ—ā–į–Ī–ĺ–Ľ–ł–∑–ľ –∂–ł—Ä–ĺ–≤–ĺ–Ļ —ā–ļ–į–Ĺ–ł) [28]. –í¬†—Ü–Ķ–Ľ–ĺ–ľ –≤–ĺ–Ņ—Ä–ĺ—Ā –ī–ĺ–∑–ł—Ä–ĺ–≤–ļ–ł –≤–ł—ā–į–ľ–ł–Ĺ–į D –ī–Ľ—Ź –Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł–ļ–ł/—ā–Ķ—Ä–į–Ņ–ł–ł —Ā–ĺ—Ā—É–ī–ł—Ā—ā—č—Ö –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ –ĺ—Ā—ā–į–Ķ—ā—Ā—Ź –ĺ—ā–ļ—Ä—č—ā—č–ľ. –ė–Ĺ–ī–ł–≤–ł–ī—É–į–Ľ—Ć–Ĺ–ĺ –Ņ–į—Ü–ł–Ķ–Ĺ—ā–į–ľ, –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ, –ľ–ĺ–≥—É—ā –Ĺ–į–∑–Ĺ–į—á–į—ā—Ć—Ā—Ź –Ĺ–Ķ—Ā–ļ–ĺ–Ľ—Ć–ļ–ĺ –Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ—č–Ķ –ī–ĺ–∑—č –≤–ł—ā–į–ľ–ł–Ĺ–į D3. –§–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ –ī–ĺ–∑—č –≤–ł—ā–į–ľ–ł–Ĺ–į D3, –ĺ—Ā–ĺ–Ī–Ķ–Ĺ–Ĺ–ĺ –≤¬†–Ņ–Ķ—Ä–ł–ĺ–ī –Ĺ–ł–∑–ļ–ĺ–Ļ –ł–Ĺ—Ā–ĺ–Ľ—Ź—Ü–ł–ł, –Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ—č –Ņ—Ä–į–ļ—ā–ł—á–Ķ—Ā–ļ–ł –ļ–į–∂–ī–ĺ–ľ—É –Ņ–į—Ü–ł–Ķ–Ĺ—ā—É.
–§–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—Ź –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –ł¬†–ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ
–ö¬†–ĺ–Ī—Č–ł–ľ —Ą–į–ļ—ā–ĺ—Ä–į–ľ –Ņ–į—ā–ĺ–≥–Ķ–Ĺ–Ķ–∑–į –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–į –ł¬†—Ā–ĺ—Ā—É–ī–ł—Ā—ā—č—Ö –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł–Ļ –ĺ—ā–Ĺ–ĺ—Ā—Ź—ā—Ā—Ź —Ö—Ä–ĺ–Ĺ–ł—á–Ķ—Ā–ļ–ĺ–Ķ –≤–ĺ—Ā–Ņ–į–Ľ–Ķ–Ĺ–ł–Ķ (–Ĺ–į–Ņ—Ä–ł–ľ–Ķ—Ä, –Ņ—Ä–ł —Ä–Ķ–≤–ľ–į—ā–ĺ–ł–ī–Ĺ–ĺ–ľ –į—Ä—ā—Ä–ł—ā–Ķ), –ĺ–ļ–ł—Ā–Ľ–Ķ–Ĺ–ł–Ķ –Ľ–ł–Ņ–ł–ī–ĺ–≤ –ł¬†–ĺ–Ī—Ä–į–∑–ĺ–≤–į–Ĺ–ł–Ķ –į–ļ—ā–ł–≤–Ĺ—č—Ö —Ą–ĺ—Ä–ľ –ļ–ł—Ā–Ľ–ĺ—Ä–ĺ–ī–į, –ļ—É—Ä–Ķ–Ĺ–ł–Ķ, —Ā–į—Ö–į—Ä–Ĺ—č–Ļ –ī–ł–į–Ī–Ķ—ā, –ī–Ķ—Ą–ł—Ü–ł—ā —ć—Ā—ā—Ä–ĺ–≥–Ķ–Ĺ–ĺ–≤,¬†–≥–ł–Ņ–ĺ–≤–ł—ā–į–ľ–ł–Ĺ–ĺ–∑—č C, D, K –ł¬†–Ņ–ĺ—á–Ķ—á–Ĺ–į—Ź –Ĺ–Ķ–ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ—Ā—ā—Ć. –ė—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź —Ā¬†–ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–ł–Ķ–ľ –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ļ—Ā–Ĺ—č—Ö –Ĺ–Ķ–ł–Ĺ–≤–į–∑–ł–≤–Ĺ—č—Ö –ľ–Ķ—ā–ĺ–ī–ĺ–≤, —ā–į–ļ–ł—Ö –ļ–į–ļ –ľ—É–Ľ—Ć—ā–ł–ī–Ķ—ā–Ķ–ļ—ā–ĺ—Ä–Ĺ–į—Ź –ļ–ĺ–ľ–Ņ—Ć—é—ā–Ķ—Ä–Ĺ–į—Ź —ā–ĺ–ľ–ĺ–≥—Ä–į—Ą–ł—Ź, –Ņ–ĺ–ļ–į–∑–į–Ľ–ł, —á—ā–ĺ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—Ź —Ā–ĺ—Ā—É–ī–ĺ–≤ –ľ–ĺ–∂–Ķ—ā –≤–ĺ–∑–Ĺ–ł–ļ–Ĺ—É—ā—Ć –≤¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –ī–≤—É—Ö –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤: –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –ł¬†–≤–į—Ā–ļ—É–Ľ—Ź—Ä–Ĺ–ĺ–Ļ –ĺ—Ā—Ā–ł—Ą–ł–ļ–į—Ü–ł–ł. –ź—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑ (–Ņ—Ä–ĺ–ł—Ā—Ö–ĺ–ī—Ź—Č–ł–Ļ, –ļ–į–ļ –Ņ—Ä–į–≤–ł–Ľ–ĺ, –≤¬†–ł–Ĺ—ā–ł–ľ–Ķ —Ā–ĺ—Ā—É–ī–į) —Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤—É–Ķ—ā –Ņ–į—Ā—Ā–ł–≤–Ĺ–ĺ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –ł¬†–≤–ļ–Ľ—é—á–į–Ķ—ā –ĺ—ā–Ľ–ĺ–∂–Ķ–Ĺ–ł—Ź –ļ–į–Ľ—Ć—Ü–ł—Ź –≤—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł–Ķ –Ĺ–Ķ–ļ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –ł¬†–į–Ņ–ĺ–Ņ—ā–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ņ–ĺ—Ä–į–∂–Ķ–Ĺ–ł–Ļ –Ĺ–į¬†–Ņ–ĺ–∑–ī–Ĺ–ł—Ö —Ā—ā–į–ī–ł—Ź—Ö —Ą–ĺ—Ä–ľ–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ī–Ľ—Ź—ą–Ķ–ļ. –ě—Ā—Ā–ł—Ą–ł–ļ–į—Ü–ł—Ź —Ā–ĺ—Ā—É–ī–ĺ–≤ (–ļ–į–ļ –Ņ—Ä–į–≤–ł–Ľ–ĺ, –ľ–Ķ–ī–ł–į–Ľ—Ć–Ĺ–į—Ź –ĺ–Ī–Ľ–į—Ā—ā—Ć) —Ä–į—Ā—Ā–ľ–į—ā—Ä–ł–≤–į–Ķ—ā—Ā—Ź –ļ–į–ļ –į–ļ—ā–ł–≤–Ĺ–į—Ź –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—Ź –≤¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤, –ł–ľ–Ķ—é—Č–ł—Ö –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä–ĺ–Ķ —Ā—Ö–ĺ–ī—Ā—ā–≤–ĺ —Ā¬†–ĺ—Ā—ā–Ķ–ĺ–≥–Ķ–Ĺ–Ķ–∑–ĺ–ľ [1].
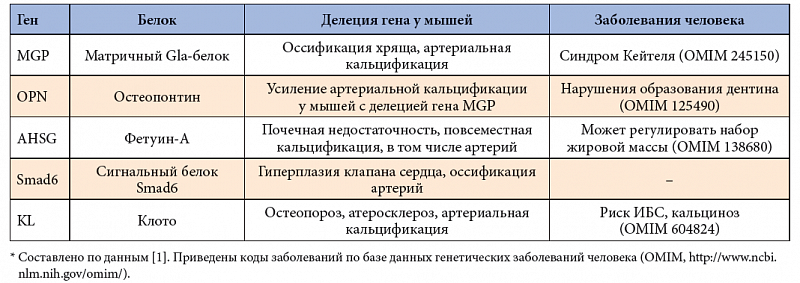
–ü–į—Ā—Ā–ł–≤–Ĺ–į—Ź –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—Ź –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–į–≥–į–Ķ—ā, —á—ā–ĺ –ļ–į–Ľ—Ć—Ü–ł–Ļ –ł¬†–Ĺ–Ķ–ĺ—Ä–≥–į–Ĺ–ł—á–Ķ—Ā–ļ–ł–Ļ —Ą–ĺ—Ā—Ą–į—ā-–į–Ĺ–ł–ĺ–Ĺ –ĺ–Ī—Ä–į–∑—É—é—ā –Ĺ–Ķ—Ä–į—Ā—ā–≤–ĺ—Ä–ł–ľ—č–Ļ –ĺ—Ā–į–ī–ĺ–ļ —Ą–ĺ—Ā—Ą–į—ā–ĺ–≤ –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†–ĺ–Ī–Ľ–į—Ā—ā–ł –ī–Ķ–≥–Ķ–Ĺ–Ķ—Ä–į—Ü–ł–ł —ā–ļ–į–Ĺ–Ķ–Ļ, –ĺ–Ī—É—Ā–Ľ–ĺ–≤–Ľ–Ķ–Ĺ–Ĺ–ĺ–Ļ –≤–ĺ—Ā–Ņ–į–Ľ–Ķ–Ĺ–ł–Ķ–ľ –ł¬†–Ņ–ĺ—Ā–Ľ–Ķ–ī—É—é—Č–ł–ľ –Ĺ–Ķ–ļ—Ä–ĺ–∑–ĺ–ľ. –≠—ā–ĺ—ā –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā —É—Ā–ł–Ľ–ł–≤–į–Ķ—ā—Ā—Ź –ľ–į–ļ—Ä–ĺ—Ą–į–≥–į–ľ–ł. –°—É—Č–Ķ—Ā—ā–≤—É–Ķ—ā –Ĺ–Ķ—Ā–ļ–ĺ–Ľ—Ć–ļ–ĺ –Ņ–ĺ—ā–Ķ–Ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č—Ö –ł–Ĺ–≥–ł–Ī–ł—ā–ĺ—Ä–ĺ–≤ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ľ–ĺ–≥—É—ā –ī–Ķ–Ļ—Ā—ā–≤–ĺ–≤–į—ā—Ć –Ľ–ĺ–ļ–į–Ľ—Ć–Ĺ–ĺ –ł–Ľ–ł —Ā–ł—Ā—ā–Ķ–ľ–Ĺ–ĺ. –ö¬†–Ĺ–ł–ľ –ĺ—ā–Ĺ–ĺ—Ā—Ź—ā—Ā—Ź –ľ–į—ā—Ä–ł–ļ—Ā–Ĺ—č–Ļ Gla-–Ī–Ķ–Ľ–ĺ–ļ (–≥–Ķ–Ĺ MGP), –≤–ł—ā–į–ľ–ł–Ĺ-K-–∑–į–≤–ł—Ā–ł–ľ—č–Ļ –ł–Ĺ–≥–ł–Ī–ł—ā–ĺ—Ä –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł, –ī–Ķ–Ļ—Ā—ā–≤—É—é—Č–ł–Ļ –≤¬†–ĺ–Ī–Ľ–į—Ā—ā–ł –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ī–Ľ—Ź—ą–Ķ–ļ, –ł¬†—Ą–Ķ—ā—É–ł–Ĺ-–ź, —É—Ā–ł–Ľ–ł–≤–į—é—Č–ł–Ļ —Ą–į–≥–ĺ—Ü–ł—ā–ĺ–∑ –ľ–ł–Ĺ–Ķ—Ä–į–Ľ—Ć–Ĺ—č—Ö –ĺ—Ā–į–ī–ļ–ĺ–≤ [29].
–ź–ļ—ā–ł–≤–Ĺ–į—Ź –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—Ź (–ĺ—Ā—Ā–ł—Ą–ł–ļ–į—Ü–ł—Ź) –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–į–Ķ—ā—Ā—Ź¬†–≥–ł—Ā—ā–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–ľ–ł –ĺ–Ī—Ä–į–∑–ĺ–≤–į–Ĺ–ł—Ź–ľ–ł, –Ĺ–į–Ņ–ĺ–ľ–ł–Ĺ–į—é—Č–ł–ľ–ł –ļ–ĺ—Ā—ā–Ĺ—É—é —ā–ļ–į–Ĺ—Ć; –Ĺ–į–Ľ–ł—á–ł–Ķ–ľ –≤¬†–∑–ĺ–Ĺ–Ķ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –ĺ—Ā—ā–Ķ–ĺ–Ī–Ľ–į—Ā—ā-–Ņ–ĺ–ī–ĺ–Ī–Ĺ—č—Ö –ł¬†–ĺ—Ā—ā–Ķ–ĺ–ļ–Ľ–į—Ā—ā-–Ņ–ĺ–ī–ĺ–Ī–Ĺ—č—Ö –ļ–Ľ–Ķ—ā–ĺ–ļ; –Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ—č–ľ–ł —É—Ä–ĺ–≤–Ĺ—Ź–ľ–ł —Ā–Ķ–ļ—Ä–Ķ—Ü–ł–ł –Ī–Ķ–Ľ–ļ–ĺ–≤-–ľ–į—Ä–ļ–Ķ—Ä–ĺ–≤ –ļ–ĺ—Ā—ā–ł –≤¬†–∑–ĺ–Ĺ–Ķ –į—Ä—ā–Ķ—Ä–ł–į–Ľ—Ć–Ĺ–ĺ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł. –í–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ, –≤–į—Ā–ļ—É–Ľ—Ź—Ä–Ĺ–į—Ź –ĺ—Ā—Ā–ł—Ą–ł–ļ–į—Ü–ł—Ź –Ņ—Ä–ĺ—ā–Ķ–ļ–į–Ķ—ā –Ņ–嬆–į–Ĺ–į–Ľ–ĺ–≥–ł–ł —Ā¬†–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–ľ —Ą–ĺ—Ä–ľ–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –ļ–ĺ—Ā—ā–Ķ–Ļ –ł¬†–≤–ļ–Ľ—é—á–į–Ķ—ā –ī–ł—Ą—Ą–Ķ—Ä–Ķ–Ĺ—Ü–ł–į—Ü–ł—é –ľ—É–Ľ—Ć—ā–ł–Ņ–ĺ—ā–Ķ–Ĺ—ā–Ĺ—č—Ö –ľ–Ķ–∑–Ķ–Ĺ—Ö–ł–ľ–į–Ľ—Ć–Ĺ—č—Ö —Ā—ā–≤–ĺ–Ľ–ĺ–≤—č—Ö –ļ–Ľ–Ķ—ā–ĺ–ļ –≤¬†–ĺ—Ā—ā–Ķ–ĺ–Ī–Ľ–į—Ā—ā-–Ņ–ĺ–ī–ĺ–Ī–Ĺ—č–Ķ –ļ–Ľ–Ķ—ā–ļ–ł, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ľ–ł–Ĺ–Ķ—Ä–į–Ľ–ł–∑—É—é—ā –≤–Ĺ–Ķ–ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č–Ļ –ľ–į—ā—Ä–ł–ļ—Ā [30]. –ź–ļ—ā–ł–≤–Ĺ–ĺ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –Ņ—Ä–Ķ–Ņ—Ź—ā—Ā—ā–≤—É—é—ā —Ą–į–ļ—ā–ĺ—Ä—č, –Ņ–ĺ–ī–į–≤–Ľ—Ź—é—Č–ł–Ķ –ĺ—Ā—ā–Ķ–ĺ–≥–Ķ–Ĺ–Ĺ—É—é –ī–ł—Ą—Ą–Ķ—Ä–Ķ–Ĺ—Ü–ł—Ä–ĺ–≤–ļ—É —Ā—ā–≤–ĺ–Ľ–ĺ–≤—č—Ö –ļ–Ľ–Ķ—ā–ĺ–ļ. –ě—ā–ľ–Ķ—ā–ł–ľ, —á—ā–ĺ –į–ļ—ā–ł–≤–Ĺ–į—Ź –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—Ź –į—Ä—ā–Ķ—Ä–ł–Ļ —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –Ī–ĺ–Ľ–Ķ–Ķ —ā—Ź–∂–Ķ–Ľ–ĺ–Ļ —Ā—ā–į–ī–ł–Ķ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł, –ĺ–Ī—É—Ā–Ľ–ĺ–≤–Ľ–Ķ–Ĺ–Ĺ–ĺ–Ļ –ļ–į–ļ¬†–≥–Ķ–Ĺ–Ķ—ā–ł—á–Ķ—Ā–ļ–ł–ľ —Ą–ĺ–Ĺ–ĺ–ľ –Ņ–į—Ü–ł–Ķ–Ĺ—ā–į, —ā–į–ļ –ł¬†—Ā—ā–Ķ–Ņ–Ķ–Ĺ—Ć—é –Ņ—Ä–ĺ–≥—Ä–Ķ—Ā—Ā–ł–ł –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į. –í–ł—ā–į–ľ–ł–Ĺ D ¬ę–Ĺ–į–Ņ—Ä–į–≤–Ľ—Ź–Ķ—ā¬Ľ –ļ–į–Ľ—Ć—Ü–ł–Ļ –≤¬†—ā–ļ–į–Ĺ–ł –ļ–ĺ—Ā—ā–Ķ–Ļ, –Ņ–ĺ—ć—ā–ĺ–ľ—É –≤¬†—Ā–Ľ—É—á–į–Ķ –į–ļ—ā–ł–≤–Ĺ–ĺ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł —ɬ†–ī–į–Ĺ–Ĺ–ĺ–≥–ĺ –Ņ–į—Ü–ł–Ķ–Ĺ—ā–į —Ā–Ľ–Ķ–ī—É–Ķ—ā —Ā¬†–ĺ—Ā—ā–ĺ—Ä–ĺ–∂–Ĺ–ĺ—Ā—ā—Ć—é –ĺ—ā–Ĺ–ĺ—Ā–ł—ā—Ć—Ā—Ź –ļ¬†–ī–ĺ–∑–ł—Ä–ĺ–≤–ļ–Ķ –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤ –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D.
–í–ł—ā–į–ľ–ł–Ĺ D –ł¬†–ľ–ĺ–Ľ–Ķ–ļ—É–Ľ—Ź—Ä–Ĺ—č–Ķ –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ—č –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ
–£–Ņ—Ä–į–≤–Ľ—Ź—Ź¬†–≥–ĺ–ľ–Ķ–ĺ—Ā—ā–į–∑–ĺ–ľ –ļ–į–Ľ—Ć—Ü–ł—Ź, –≤–ł—ā–į–ľ–ł–Ĺ D –ľ–ĺ–∂–Ķ—ā —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤–ĺ–≤–į—ā—Ć –Ņ—Ä–Ķ–ļ—Ä–į—Č–Ķ–Ĺ–ł—é –ł¬†–Ņ–į—Ā—Ā–ł–≤–Ĺ–ĺ–Ļ, –ł¬†–į–ļ—ā–ł–≤–Ĺ–ĺ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł. –ě—Ā–ĺ–Ī–ĺ–Ķ –≤–Ĺ–ł–ľ–į–Ĺ–ł–Ķ —Ā–Ľ–Ķ–ī—É–Ķ—ā –ĺ–Ī—Ä–į—ā–ł—ā—Ć –Ĺ–į¬†—ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ—č–Ķ –ł¬†–Ī–ł–ĺ—Ö–ł–ľ–ł—á–Ķ—Ā–ļ–ł–Ķ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź –≤–Ľ–ł—Ź–Ĺ–ł—Ź –≤–ł—ā–į–ľ–ł–Ĺ–į D –Ĺ–į¬†—ć–ļ—Ā–Ņ—Ä–Ķ—Ā—Ā–ł—鬆–≥–Ķ–Ĺ–ĺ–≤ –ł¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ —ā—Ä–į–Ĺ—Ā–≥–Ķ–Ĺ–Ĺ—č—Ö –∂–ł–≤–ĺ—ā–Ĺ—č—Ö —Ā¬†–ī–Ķ–Ľ–Ķ—Ü–ł—Ź–ľ–ł –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ—č—Ö¬†–≥–Ķ–Ĺ–ĺ–≤, –≤–ĺ–≤–Ľ–Ķ—á–Ķ–Ĺ–Ĺ—č—Ö –≤¬†–ľ–Ķ—ā–į–Ī–ĺ–Ľ–ł–∑–ľ –ļ–ĺ—Ā—ā–Ĺ–ĺ–Ļ —ā–ļ–į–Ĺ–ł.
–ö–į–ļ –Ī—č–Ľ–ĺ –ĺ—ā–ľ–Ķ—á–Ķ–Ĺ–ĺ –≤—č—ą–Ķ, –≤–ł—ā–į–ľ–ł–Ĺ D –ĺ–Ī–Ľ–į–ī–į–Ķ—ā¬†–≥–ĺ—Ä–ľ–ĺ–Ĺ–ĺ–Ņ–ĺ–ī–ĺ–Ī–Ĺ—č–ľ —ć—Ą—Ą–Ķ–ļ—ā–ĺ–ľ –ł¬†—á–Ķ—Ä–Ķ–∑ –≤–∑–į–ł–ľ–ĺ–ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ —Ā¬†–ĺ–ī–Ĺ–ĺ–ł–ľ–Ķ–Ĺ–Ĺ—č–ľ —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–ĺ–ľ —É—á–į—Ā—ā–≤—É–Ķ—ā –≤¬†—Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł–ł —Ā–ĺ—ā–Ķ–Ŭ†–≥–Ķ–Ĺ–ĺ–≤. –ė—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź –≤–Ľ–ł—Ź–Ĺ–ł—Ź –≤–ł—ā–į–ľ–ł–Ĺ–į D –Ĺ–į¬†—ć–ļ—Ā–Ņ—Ä–Ķ—Ā—Ā–ł—鬆–≥–Ķ–Ĺ–ĺ–≤ [31, 32] –Ņ–ĺ–∑–≤–ĺ–Ľ–ł–Ľ–ĺ —É—Ā—ā–į–Ĺ–ĺ–≤–ł—ā—Ć —Ā–Ņ–ł—Ā–ĺ–ļ¬†–≥–Ķ–Ĺ–ĺ–≤, –Ĺ–į¬†–ĺ—Ā–Ĺ–ĺ–≤–Ķ –ļ–ĺ—ā–ĺ—Ä—č—Ö —Ā–ł–Ĺ—ā–Ķ–∑–ł—Ä—É—é—ā—Ā—Ź –Ī–Ķ–Ľ–ļ–ł, –ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–ł–≤–į—é—Č–ł–Ķ —Ö–į—Ä–į–ļ—ā–Ķ—Ä–Ĺ—č–Ķ –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —ć—Ą—Ą–Ķ–ļ—ā—č –≤–ł—ā–į–ľ–ł–Ĺ–į (—ā–į–Ī–Ľ.¬†1). –ď–Ķ–Ĺ—č –ł¬†—Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤—É—é—Č–ł–Ķ –Ī–Ķ–Ľ–ļ–ł, –ł–ľ–Ķ—é—Č–ł–Ķ –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ–Ķ –ĺ—ā–Ĺ–ĺ—ą–Ķ–Ĺ–ł–Ķ –ļ¬†–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑—É, –Ľ–ł–Ņ–ł–ī–Ĺ–ĺ–ľ—É –ľ–Ķ—ā–į–Ī–ĺ–Ľ–ł–∑–ľ—É –ł¬†–ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ, —Ä–į—Ā—Ā–ľ–ĺ—ā—Ä–Ķ–Ĺ—č –Ĺ–ł–∂–Ķ –Ī–ĺ–Ľ–Ķ–Ķ –Ņ–ĺ–ī—Ä–ĺ–Ī–Ĺ–ĺ.
–ě—Ā–Ĺ–ĺ–≤–Ĺ—č–ľ–ł –≤–ł—ā–į–ľ–ł–Ĺ-D-–∑–į–≤–ł—Ā–ł–ľ—č–ľ–ł —Ä–Ķ–≥—É–Ľ—Ź—ā–ĺ—Ä–į–ľ–ł¬†–≥–ĺ–ľ–Ķ–ĺ—Ā—ā–į–∑–į –ļ–į–Ľ—Ć—Ü–ł—Ź —Ź–≤–Ľ—Ź—é—ā—Ā—Ź –ł–ĺ–Ĺ–Ĺ—č–Ļ –ļ–į–Ĺ–į–Ľ TRPV6, –ļ–į–Ľ—Ć–Ī–ł–Ĺ–ī–ł–Ĺ, –ĺ—Ā—ā–Ķ–ĺ–ļ–į–Ľ—Ć—Ü–ł–Ĺ –ł¬†–ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ–Ĺ—ā–ł–Ĺ. TRPV6 –ĺ—ā–≤–Ķ—á–į–Ķ—ā –∑–į –į–ī—Ā–ĺ—Ä–Ī—Ü–ł—é –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†–≤–ĺ—Ä—Ā–ł–Ĺ—á–į—ā–ĺ–ľ —ć–Ņ–ł—ā–Ķ–Ľ–ł–ł –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į [33]; –ļ–į–Ľ—Ć–Ī–ł–Ĺ–ī–ł–Ĺ —ā—Ä–į–Ĺ—Ā–Ņ–ĺ—Ä—ā–ł—Ä—É–Ķ—ā –ļ–į–Ľ—Ć—Ü–ł–Ļ —á–Ķ—Ä–Ķ–∑ —ć–Ĺ—ā–Ķ—Ä–ĺ—Ü–ł—ā—č –ļ¬†–Ī–į–∑–ĺ–Ľ–į—ā–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ā—ā–ĺ—Ä–ĺ–Ĺ–Ķ —ć–Ĺ—ā–Ķ—Ä–ĺ—Ü–ł—ā–į,¬†–≥–ī–Ķ –ļ–į–Ľ—Ć—Ü–ł–Ļ —Ā–Ķ–ļ—Ä–Ķ—ā–ł—Ä—É–Ķ—ā—Ā—Ź –≤¬†–ļ—Ä–ĺ–≤—Ź–Ĺ–ĺ–Ķ —Ä—É—Ā–Ľ–ĺ –Ņ–ĺ—Ā—Ä–Ķ–ī—Ā—ā–≤–ĺ–ľ –ļ–į–Ľ—Ć—Ü–ł–Ķ–≤—č—Ö –Ĺ–į—Ā–ĺ—Ā–ĺ–≤. –ě—Ā—ā–Ķ–ĺ–ļ–į–Ľ—Ć—Ü–ł–Ĺ –ł¬†–ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ–Ĺ—ā–ł–Ĺ —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É—é—ā –∑–į–ļ—Ä–Ķ–Ņ–Ľ–Ķ–Ĺ–ł—é –ĺ—Ā—ā–Ķ–ĺ—Ü–ł—ā–ĺ–≤ –Ĺ–į¬†–Ņ–ĺ–≤–Ķ—Ä—Ö–Ĺ–ĺ—Ā—ā–ł –ļ–ĺ—Ā—ā–ł –ł¬†–Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ—č –ī–Ľ—Ź –Ĺ–į–Ņ—Ä–į–≤–Ľ–Ķ–Ĺ–ł—Ź –Ņ–ĺ—ā–ĺ–ļ–ĺ–≤ –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†–ļ–į–Ľ—Ć—Ü–ł–Ķ–≤–ĺ–Ķ –ī–Ķ–Ņ–ĺ –ļ–ĺ—Ā—ā–Ĺ–ĺ–Ļ —ā–ļ–į–Ĺ–ł. –£—Ä–ĺ–≤–Ĺ–ł –ĺ—Ā—ā–Ķ–ĺ–ļ–į–Ľ—Ć—Ü–ł–Ĺ–į, –ļ–į–ļ –ł–∑–≤–Ķ—Ā—ā–Ĺ–ĺ, –ĺ—ā—Ä–į–∂–į—é—ā –ľ–ł–Ĺ–Ķ—Ä–į–Ľ—Ć–Ĺ—É—é –Ņ–Ľ–ĺ—ā–Ĺ–ĺ—Ā—ā—Ć –ļ–ĺ—Ā—ā–ł [34]. –ö—Ä–ĺ–ľ–Ķ —ā–ĺ–≥–ĺ, –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ–Ĺ—ā–ł–Ĺ –≤–ĺ–≤–Ľ–Ķ—á–Ķ–Ĺ –≤¬†—Ā–ł–Ĺ—ā–Ķ–∑ –ł¬†—Ā–Ķ–ļ—Ä–Ķ—Ü–ł—é ő≥-–ł–Ĺ—ā–Ķ—Ä—Ą–Ķ—Ä–ĺ–Ĺ–į –ł¬†–ł–Ĺ—ā–Ķ—Ä–Ľ–Ķ–Ļ–ļ–ł–Ĺ–ĺ–≤ IL-10 –ł¬†IL-12 [35].
–Ē—Ä—É–≥–ł–Ķ¬†–≥–Ķ–Ĺ—č, –į–ļ—ā–ł–≤–ł—Ä—É–Ķ–ľ—č–Ķ —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–ĺ–ľ –≤–ł—ā–į–ľ–ł–Ĺ–į D (—ā–į–Ī–Ľ.¬†1), –ĺ–ļ–į–∑—č–≤–į—é—ā –Ņ–ĺ–Ľ–ĺ–∂–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ķ –≤–ĺ–∑–ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ –Ĺ–į¬†–Ľ–ł–Ņ–ł–ī–Ĺ—č–Ļ –ľ–Ķ—ā–į–Ī–ĺ–Ľ–ł–∑–ľ –ł¬†–≤–į–∑–ĺ–ī–ł–Ľ–į—ā–į—Ü–ł—é, —ā–Ķ–ľ —Ā–į–ľ—č–ľ —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É—Ź —É–ľ–Ķ–Ĺ—Ć—ą–Ķ–Ĺ–ł—é —Ä–ł—Ā–ļ–į –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –ł¬†—Ā–≤—Ź–∑–į–Ĺ–Ĺ–ĺ–Ļ —Ā¬†–Ĺ–ł–ľ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ. –í–ł—ā–į–ľ–ł–Ĺ D3 —Ā—ā–ł–ľ—É–Ľ–ł—Ä—É–Ķ—ā —Ā–ł–Ĺ—ā–Ķ–∑ –Ņ—Ä–ĺ—ā–ł–≤–ĺ–≤–ĺ—Ā–Ņ–į–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ –Ņ—Ä–ĺ—Ā—ā–į—Ü–ł–ļ–Ľ–ł–Ĺ–į –≤¬†—Ā–ĺ—Ā—É–ī–ł—Ā—ā—č—Ö¬†–≥–Ľ–į–ī–ļ–ĺ–ľ—č—ą–Ķ—á–Ĺ—č—Ö –ļ–Ľ–Ķ—ā–ļ–į—Ö —á–Ķ—Ä–Ķ–∑ –į–ļ—ā–ł–≤–į—Ü–ł—é —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł–ł —Ü–ł–ļ–Ľ–ĺ–ĺ–ļ—Ā–ł–≥–Ķ–Ĺ–į–∑—č –Ī–Ķ–∑ –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł—Ź –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł —Ą–ĺ—Ā—Ą–ĺ–Ľ–ł–Ņ–į–∑—č –ź2 [36]. –ź–ļ—ā–ł–≤–į—Ü–ł—Ź —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–į –≤–ł—ā–į–ľ–ł–Ĺ–į D —ā–ĺ—Ä–ľ–ĺ–∑–ł—ā –ł–∑–Ī—č—ā–ĺ—á–Ĺ—č–Ļ —Ā–ł–Ĺ—ā–Ķ–∑ 7-–į–Ľ—Ć—Ą–į-—Ö–ĺ–Ľ–Ķ—Ā—ā–Ķ—Ä–ł–Ĺ-–≥–ł–ī—Ä–ĺ–ļ—Ā–ł–Ľ–į–∑—č (–≥–Ķ–Ĺ CYP7A1), —á—ā–ĺ –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†–Ĺ–ĺ—Ä–ľ–į–Ľ–ł–∑–į—Ü–ł–ł —Ā–ł–Ĺ—ā–Ķ–∑–į –∂–Ķ–Ľ—á–Ĺ—č—Ö –ļ–ł—Ā–Ľ–ĺ—ā –≤¬†–≥–Ķ–Ņ–į—ā–ĺ—Ü–ł—ā–į—Ö —á–Ķ–Ľ–ĺ–≤–Ķ–ļ–į [37]. –í–ł—ā–į–ľ–ł–Ĺ D —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É–Ķ—ā —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł—é –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł–ł —Ä–Ķ–Ĺ–ł–Ĺ–į¬†‚Äď –ĺ—Ā–Ĺ–ĺ–≤–Ĺ–ĺ–≥–ĺ –ľ–Ķ–ī–ł–į—ā–ĺ—Ä–į —Ä–Ķ–Ĺ–ł–Ĺ-–į–Ĺ–≥–ł–ĺ—ā–Ķ–Ĺ–∑–ł–Ĺ–ĺ–≤–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–į–≤–Ľ–Ķ–Ĺ–ł—Ź [38] (—Ä–ł—Ā.¬†1). –Ě–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–į—Ź –ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–Ķ–Ĺ–Ĺ–ĺ—Ā—ā—Ć –≤–ł—ā–į–ľ–ł–Ĺ–ĺ–ľ D¬†‚Äď –Ņ—Ä–ł–∑–Ĺ–į–Ĺ–Ĺ—č–Ļ —Ą–į–ļ—ā–ĺ—Ä –Ĺ–ĺ—Ä–ľ–į–Ľ–ł–∑–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–į–≤–Ľ–Ķ–Ĺ–ł—Ź.
–ė–Ĺ—Ā—É–Ľ–ł–Ĺ–ĺ–Ņ–ĺ–ī–ĺ–Ī–Ĺ—č–Ļ —Ą–į–ļ—ā–ĺ—Ä —Ä–ĺ—Ā—ā–į (IGF) —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –ĺ–ī–Ĺ–ł–ľ –ł–∑¬†–≤–į–∂–Ĺ–Ķ–Ļ—ą–ł—Ö —Ą–į–ļ—ā–ĺ—Ä–ĺ–≤, –Ņ–ĺ–ī–ī–Ķ—Ä–∂–ł–≤–į—é—Č–ł—Ö –Ī–į–Ľ–į–Ĺ—Ā –ľ–Ķ–∂–ī—É –∂–ł—Ä–ĺ–≤–ĺ–Ļ –ł¬†–ľ—č—ą–Ķ—á–Ĺ–ĺ–Ļ —ā–ļ–į–Ĺ—Ć—é. –ü—Ä–ł –ī–Ķ—Ą–ł—Ü–ł—ā–Ķ IGF –ł–Ľ–ł —Ā–Ĺ–ł–∂–Ķ–Ĺ–ł–ł –Ķ–≥–ĺ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł –∂–ł—Ä–ĺ–≤–į—Ź —ā–ļ–į–Ĺ—Ć –Ĺ–į—á–ł–Ĺ–į–Ķ—ā –Ņ—Ä–Ķ–ĺ–Ī–Ľ–į–ī–į—ā—Ć –Ĺ–į–ī –ľ—č—ą–Ķ—á–Ĺ–ĺ–Ļ. –í¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ —É—Ā–ļ–ĺ—Ä—Ź—é—ā—Ā—Ź –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –ł¬†—Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź. –í–ł—ā–į–ľ–ł–Ĺ D —Ā—ā–ł–ľ—É–Ľ–ł—Ä—É–Ķ—ā —Ā–ł–Ĺ—ā–Ķ–∑ IGF-—Ā–≤—Ź–∑—č–≤–į—é—Č–ł—Ö –Ī–Ķ–Ľ–ļ–ĺ–≤ (IGFBP), –ļ–ĺ—ā–ĺ—Ä—č–Ķ –Ņ—Ä–ĺ–ī–Ľ–Ķ–≤–į—é—ā –Ņ–Ķ—Ä–ł–ĺ–ī –Ņ–ĺ–Ľ—É—Ä–į—Ā–Ņ–į–ī–į IGF, —ā–Ķ–ľ —Ā–į–ľ—č–ľ —É—Ā–ł–Ľ–ł–≤–į—Ź –Ķ–≥–ĺ –į–Ĺ—ā–ł–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł–Ķ —ć—Ą—Ą–Ķ–ļ—ā—č (—Ä–ł—Ā.¬†2). –ė—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź –Ņ–ĺ–ļ–į–∑–į–Ľ–ł, —á—ā–ĺ –≤–ł—ā–į–ľ–ł–Ĺ D —É—Ā–ł–Ľ–ł–≤–į–Ķ—ā —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł—鬆–≥–Ķ–Ĺ–ĺ–≤ IGFBP1, IGFBP3 –ł¬†IGFBP5. –°—č–≤–ĺ—Ä–ĺ—ā–ĺ—á–Ĺ—č–Ķ —É—Ä–ĺ–≤–Ĺ–ł IGF-1 –ł¬†IGFBP-3 –Ī—č–Ľ–ł –ł–∑–ľ–Ķ—Ä–Ķ–Ĺ—č —É¬†96 –Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ —Ā¬†–Ņ–Ķ—Ä–ł—Ą–Ķ—Ä–ł—á–Ķ—Ā–ļ–ĺ–Ļ –Ī–ĺ–Ľ–Ķ–∑–Ĺ—Ć—é –į—Ä—ā–Ķ—Ä–ł–Ļ –ł¬†—É 89 –ī–ĺ–Ī—Ä–ĺ–≤–ĺ–Ľ—Ć—Ü–Ķ–≤. –£¬†–Ī–ĺ–Ľ—Ć–Ĺ—č—Ö –Ī—č–Ľ–ł –Ĺ–į–Ļ–ī–Ķ–Ĺ—č –Ī–ĺ–Ľ–Ķ–Ķ –Ĺ–ł–∑–ļ–ł–Ķ —É—Ä–ĺ–≤–Ĺ–ł –Ī–Ķ–Ľ–ļ–į IGFBP-3 (p < 0,01), —ā–į–ļ —á—ā–ĺ IGFBP-3 –ī–Ķ–Ļ—Ā—ā–≤–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ —Ā–≤—Ź–∑–į–Ĺ —Ā¬†—ā–ĺ—Ä–ľ–ĺ–∂–Ķ–Ĺ–ł–Ķ–ľ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–į –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į [39].
–Ě–Ķ¬†–ľ–Ķ–Ĺ–Ķ–Ķ –≤–į–∂–Ĺ–ĺ –ĺ—ā–ľ–Ķ—ā–ł—ā—Ć, —á—ā–ĺ –≤–ł—ā–į–ľ–ł–Ĺ D –į–ļ—ā–ł–≤–ł—Ä—É–Ķ—ā —Ā–ł–Ĺ—ā–Ķ–∑ –Ī–Ķ–Ľ–ļ–į PPAR-őī (–į–ļ—ā–ł–≤–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č–Ļ —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä –Ņ—Ä–ĺ–Ľ–ł—Ą–Ķ—Ä–į—ā–ĺ—Ä–ĺ–≤ –Ņ–Ķ—Ä–ĺ–ļ—Ā–ł—Ā–ĺ–ľ, —ā–ł–Ņ –ī–Ķ–Ľ—Ć—ā–į), —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É—é—Č–Ķ–≥–ĺ –≤—č–≤–Ķ–ī–Ķ–Ĺ–ł—é –ł–∑–Ī—č—ā–ļ–į —Ö–ĺ–Ľ–Ķ—Ā—ā–Ķ—Ä–ł–Ĺ–į [40]. –Ď–Ķ–Ľ–ĺ–ļ PPAR-őī (—Ä–ł—Ā.¬†3) –į–ļ—ā–ł–≤–ł—Ä—É–Ķ—ā –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č –Ī–Ķ—ā–į-–ĺ–ļ–ł—Ā–Ľ–Ķ–Ĺ–ł—Ź –∂–ł—Ä–Ĺ—č—Ö –ļ–ł—Ā–Ľ–ĺ—ā, —ā–ĺ –Ķ—Ā—ā—Ć –į–ļ—ā–ł–≤–į—Ü–ł—Ź —Ā–ł–Ĺ—ā–Ķ–∑–į PPAR-őī –≤–ł—ā–į–ľ–ł–Ĺ–ĺ–ľ D3 –Ņ—Ä–ł–≤–ĺ–ī–ł—ā –ļ¬†—Ā–Ĺ–ł–∂–Ķ–Ĺ–ł—é –ł–Ĺ—ā–Ķ–Ĺ—Ā–ł–≤–Ĺ–ĺ—Ā—ā–ł –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤. –ė—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ –Ņ—Ä–ł–ľ–Ķ—Ä–Ĺ–ĺ 2000 –Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ —Ā¬†–ė–Ď–° (–≤¬†—Ā—Ä–į–≤–Ĺ–Ķ–Ĺ–ł–ł —Ā–嬆–∑–ī–ĺ—Ä–ĺ–≤—č–ľ–ł –ł–Ĺ–ī–ł–≤–ł–ī—É—É–ľ–į–ľ–ł) [41] –Ņ–ĺ–ļ–į–∑–į–Ľ–ł, —á—ā–ĺ –Ņ–ĺ–Ľ–ł–ľ–ĺ—Ä—Ą–ł–∑–ľ¬†–≥–Ķ–Ĺ–į PPAR-őī –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ—Ź–Ķ—ā –Ņ–Ľ–į–∑–ľ–Ķ–Ĺ–Ĺ—č–Ķ —É—Ä–ĺ–≤–Ĺ–ł –Ľ–ł–Ņ–ł–ī–ĺ–≤ –ł¬†—ā—Ź–∂–Ķ—Ā—ā—Ć –ļ–ĺ—Ä–ĺ–Ĺ–į—Ä–Ĺ–ĺ–≥–ĺ –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į. –Ę–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, –≤–ł—ā–į–ľ–ł–Ĺ D3 —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É–Ķ—ā –į–ļ—ā–ł–≤–į—Ü–ł–ł —Ā–ł–Ĺ—ā–Ķ–∑–į –Ĺ–Ķ—Ā–ļ–ĺ–Ľ—Ć–ļ–ł—Ö –≤–į–∂–Ĺ–Ķ–Ļ—ą–ł—Ö –į–Ĺ—ā–ł–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –ł¬†–≤–į–∑–ĺ–ī–ł–Ľ–į—ā–ł—Ä—É—é—Č–ł—Ö —Ą–į–ļ—ā–ĺ—Ä–ĺ–≤.
–ē—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ, —á—ā–ĺ –Ņ–Ķ—Ä–Ķ—á–ł—Ā–Ľ–Ķ–Ĺ–Ĺ—č–Ķ –≤¬†—ā–į–Ī–Ľ–ł—Ü–Ķ 1¬†–≥–Ķ–ŗ謆‚Äď –ī–į–Ľ–Ķ–ļ–ĺ –Ĺ–Ķ¬†–Ņ–ĺ–Ľ–Ĺ—č–Ļ —Ā–Ņ–ł—Ā–ĺ–ļ¬†–≥–Ķ–Ĺ–ĺ–≤, —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł—Ź –ļ–ĺ—ā–ĺ—Ä—č—Ö –ľ–ĺ–ī—É–Ľ–ł—Ä—É–Ķ—ā—Ā—Ź —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä–ĺ–ľ –≤–ł—ā–į–ľ–ł–Ĺ–į D. –Ē–ĺ–Ņ–ĺ–Ľ–Ĺ–ł—ā–Ķ–Ľ—Ć–Ĺ—č–ľ –ł—Ā—ā–ĺ—á–Ĺ–ł–ļ–ĺ–ľ –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł —Ź–≤–Ľ—Ź—é—ā—Ā—Ź –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź —ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ—č—Ö –ľ–ĺ–ī–Ķ–Ľ–Ķ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł —Ā–ĺ—Ā—É–ī–ĺ–≤ —ɬ†–∂–ł–≤–ĺ—ā–Ĺ—č—Ö —Ā¬†–ī–Ķ–Ľ–Ķ—Ü–ł—Ź–ľ–ł –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–Ĺ—č—Ö¬†–≥–Ķ–Ĺ–ĺ–≤.
–†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ —ā—Ä–į–Ĺ—Ā–≥–Ķ–Ĺ–Ĺ—č—Ö –∂–ł–≤–ĺ—ā–Ĺ—č—Ö —Ā¬†–ī–Ķ–Ľ–Ķ—Ü–ł—Ź–ľ–ł¬†–≥–Ķ–Ĺ–ĺ–≤, –≤–ĺ–≤–Ľ–Ķ—á–Ķ–Ĺ–Ĺ—č—Ö –≤¬†—Ä–Ķ–≥—É–Ľ—Ź—Ü–ł—鬆–≥–ĺ–ľ–Ķ–ĺ—Ā—ā–į–∑–į –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†–ļ–ĺ—Ā—ā—Ź—Ö, –≤—č—Ź–≤–ł–Ľ–ł –≤–į–∂–Ĺ—č–Ķ –ī–Ķ—ā–į–Ľ–ł –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ—Ź—Ä–Ĺ—č—Ö –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–ĺ–≤, –Ņ–ĺ—Ā—Ä–Ķ–ī—Ā—ā–≤–ĺ–ľ –ļ–ĺ—ā–ĺ—Ä—č—Ö –ľ–ĺ–∂–Ķ—ā –ĺ—Ā—É—Č–Ķ—Ā—ā–≤–Ľ—Ź—ā—Ć—Ā—Ź –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—Ź –į—Ä—ā–Ķ—Ä–ł–Ļ –Ņ–嬆–į–ļ—ā–ł–≤–Ĺ–ĺ–ľ—É –ł¬†–Ņ–į—Ā—Ā–ł–≤–Ĺ–ĺ–ľ—É –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ—É (—Ā–ľ. –≤—č—ą–Ķ) [1]. –†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č —Ā—É–ľ–ľ–ł—Ä–ĺ–≤–į–Ĺ—č –≤¬†—ā–į–Ī–Ľ–ł—Ü–Ķ 2, –≤¬†–ļ–ĺ—ā–ĺ—Ä–ĺ–Ļ —ā–į–ļ–∂–Ķ –Ņ—Ä–ĺ–≤–ĺ–ī–ł—ā—Ā—Ź —Ā—Ä–į–≤–Ĺ–Ķ–Ĺ–ł–Ķ —Ā¬†–ł–∑–≤–Ķ—Ā—ā–Ĺ—č–ľ–ł –Ĺ–į¬†—Ā–Ķ–≥–ĺ–ī–Ĺ—Ź—ą–Ĺ–ł–Ļ –ī–Ķ–Ĺ—Ć —Ä–Ķ–ī–ļ–ł–ľ–ł –ľ–ĺ–Ĺ–ĺ–≥–Ķ–Ĺ–Ĺ—č–ľ–ł –∑–į–Ī–ĺ–Ľ–Ķ–≤–į–Ĺ–ł—Ź–ľ–ł —á–Ķ–Ľ–ĺ–≤–Ķ–ļ–į, —Ā–≤—Ź–∑–į–Ĺ–Ĺ—č–ľ–ł —Ā¬†–Ĺ–į—Ä—É—ą–Ķ–Ĺ–ł—Ź–ľ–ł –ļ–į–Ľ—Ć—Ü–ł–Ķ–≤–ĺ–≥–ĺ –ĺ–Ī–ľ–Ķ–Ĺ–į. –†–į—Ā—Ā–ľ–ĺ—ā—Ä–ł–ľ –ī–Ķ—ā–į–Ľ—Ć–Ĺ–Ķ–Ķ –Ī–Ķ–Ľ–ļ–ł, —É—á–į—Ā—ā–≤—É—é—Č–ł–Ķ –≤¬†–∑–į—Č–ł—ā–Ķ –ĺ—ā¬†–ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ, –ł¬†–ł—Ö –≤–∑–į–ł–ľ–ĺ—Ā–≤—Ź–∑—Ć —Ā¬†–≤–ł—ā–į–ľ–ł–Ĺ–ĺ–ľ D.
–ú–į—ā—Ä–ł—á–Ĺ—č–Ļ Gla-–Ī–Ķ–Ľ–ĺ–ļ (–≥–Ķ–Ĺ MGP) —Ā–ĺ–ī–Ķ—Ä–∂–ł—ā –ĺ—Ā—ā–į—ā–ļ–ł¬†–≥–į–ľ–ľ–į-–ļ–į—Ä–Ī–ĺ–ļ—Ā–ł-–≥–Ľ—É—ā–į–ľ–ł–Ĺ–ĺ–≤–ĺ–Ļ –į–ľ–ł–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā—č (GLA), –Ī–Ľ–į–≥–ĺ–ī–į—Ä—Ź –ļ–ĺ—ā–ĺ—Ä—č–ľ –ĺ–Ī–Ľ–į–ī–į–Ķ—ā –≤—č—Ā–ĺ–ļ–ł–ľ —Ā—Ä–ĺ–ī—Ā—ā–≤–ĺ–ľ –ļ¬†–≥–ł–ī—Ä–ĺ–ļ—Ā–ł–į–Ņ–į—ā–ł—ā—É –ļ–ĺ—Ā—ā–Ķ–Ļ [29]. –†–Ķ—ą–į—é—Č—É—é —Ä–ĺ–Ľ—Ć MGP –ī–Ľ—Ź –ļ–ĺ—Ā—ā–Ĺ–ĺ–≥–ĺ –ł¬†—Ö—Ä—Ź—Č–Ķ–≤–ĺ–≥–ĺ –ľ–Ķ—ā–į–Ī–ĺ–Ľ–ł–∑–ľ–į –Ņ–ĺ–ī—á–Ķ—Ä–ļ–ł–≤–į—é—ā —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ļ –ľ—č—ą–Ķ–Ļ —Ā¬†–ī–Ķ–Ľ–Ķ—Ü–ł–Ķ–Ļ¬†–≥–Ķ–Ĺ–į, —ɬ†–ļ–ĺ—ā–ĺ—Ä—č—Ö –Ĺ–į–Ī–Ľ—é–ī–į–Ľ–į—Ā—Ć –Ņ–ĺ–≤—Ā–Ķ–ľ–Ķ—Ā—ā–Ĺ–į—Ź –ĺ—Ā—Ā–ł—Ą–ł–ļ–į—Ü–ł—Ź —Ö—Ä—Ź—Č–į –ł¬†–į—Ä—ā–Ķ—Ä–ł–į–Ľ—Ć–Ĺ–į—Ź –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—Ź. –í–į–∂–Ĺ–ĺ –ĺ—ā–ľ–Ķ—ā–ł—ā—Ć, —á—ā–ĺ —ć–ļ—Ā–Ņ—Ä–Ķ—Ā—Ā–ł—Ź¬†–≥–Ķ–Ĺ–į MGP —Ä–Ķ–≥—É–Ľ–ł—Ä—É–Ķ—ā—Ā—Ź —Ä—Ź–ī–ĺ–ľ —Ą–į–ļ—ā–ĺ—Ä–ĺ–≤, –≤¬†—ā–ĺ–ľ —á–ł—Ā–Ľ–Ķ –Ņ—Ä–ĺ–ł–∑–≤–ĺ–ī–Ĺ—č–ľ–ł –≤–ł—ā–į–ľ–ł–Ĺ–į –ź, –≤–ł—ā–į–ľ–ł–Ĺ–ĺ–ľ D –ł¬†–≤–Ĺ–Ķ–ļ–Ľ–Ķ—ā–ĺ—á–Ĺ—č–ľ –ļ–į–Ľ—Ć—Ü–ł–Ķ–ľ [42].
–ě—Ā—ā–Ķ–ĺ–Ņ–ĺ–Ĺ—ā–ł–Ŭ†‚Äď –Ĺ–Ķ–ļ–ĺ–Ľ–Ľ–į–≥–Ķ–Ĺ–ĺ–≤—č–Ļ –Ī–Ķ–Ľ–ĺ–ļ –≤–Ĺ–Ķ–ļ–Ľ–Ķ—ā–ĺ—á–Ĺ–ĺ–Ļ –ľ–į—ā—Ä–ł—Ü—č –ļ–ĺ—Ā—ā–Ķ–Ļ, –ļ–ĺ—ā–ĺ—Ä—č–Ļ —Ā–≤—Ź–∑—č–≤–į–Ķ—ā—Ā—Ź —Ā¬†–ł–Ĺ—ā–Ķ–≥—Ä–ł–Ĺ–į–ľ–ł –ł¬†–ł–ľ–Ķ–Ķ—ā —Ä–Ķ—ą–į—é—Č–Ķ–Ķ –∑–Ĺ–į—á–Ķ–Ĺ–ł–Ķ –ī–Ľ—Ź –ľ–ł–≥—Ä–į—Ü–ł–ł –ĺ—Ā—ā–Ķ–ĺ–ļ–Ľ–į—Ā—ā–ĺ–≤ –ļ¬†—Ā–į–Ļ—ā–į–ľ —Ä–Ķ–∑–ĺ—Ä–Ī—Ü–ł–ł –ļ–ĺ—Ā—ā–Ķ–Ļ [43]. –ě—Ā—ā–Ķ–ĺ–Ņ–ĺ–Ĺ—ā–ł–Ĺ —É–Ľ—É—á—ą–į–Ķ—ā —Ā–≤—Ź–∑—č–≤–į–Ĺ–ł–Ķ –ĺ—Ā—ā–Ķ–ĺ–ļ–Ľ–į—Ā—ā–ĺ–≤ –ł¬†—ā–ĺ—Ä–ľ–ĺ–∑–ł—ā –į–Ņ–ĺ–Ņ—ā–ĺ–∑ —ć–Ĺ–ī–ĺ—ā–Ķ–Ľ–ł–į–Ľ—Ć–Ĺ—č—Ö –ļ–Ľ–Ķ—ā–ĺ–ļ. –Ē–Ķ–Ľ–Ķ—Ü–ł–ł¬†–≥–Ķ–Ĺ–į –≤¬†—ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–Ķ –Ī—č–Ľ–ł —Ā–≤—Ź–∑–į–Ĺ—č —Ā¬†—É—Ā–ł–Ľ–Ķ–Ĺ–ł–Ķ–ľ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ; –Ņ–ĺ–Ľ–ł–ľ–ĺ—Ä—Ą–ł–∑–ľ¬†–≥–Ķ–Ĺ–į –į—Ā—Ā–ĺ—Ü–ł–ł—Ä—É–Ķ—ā—Ā—Ź —Ā¬†–į—Ä—ā–Ķ—Ä–ł–į–Ľ—Ć–Ĺ–ĺ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–Ķ–Ļ —ɬ†–Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ —Ā¬†–Ņ—Ä–ĺ–≥—Ä–Ķ—Ā—Ā–ł—Ä—É—é—Č–ł–ľ –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–ĺ–ľ [44]. –í–ł—ā–į–ľ–ł–Ĺ D –ľ–ĺ–ī—É–Ľ–ł—Ä—É–Ķ—ā —ć–ļ—Ā–Ņ—Ä–Ķ—Ā—Ā–ł—鬆–≥–Ķ–Ĺ–į –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ–Ĺ—ā–ł–Ĺ–į [45].
–§–Ķ—ā—É–ł–Ĺ-–ź¬†—Ü–ł—Ä–ļ—É–Ľ–ł—Ä—É–Ķ—ā –≤¬†—Ā—č–≤–ĺ—Ä–ĺ—ā–ļ–Ķ –ļ—Ä–ĺ–≤–ł –ł¬†—Ā–Ľ—É–∂–ł—ā –≤¬†–ļ–į—á–Ķ—Ā—ā–≤–Ķ –ł–Ĺ–≥–ł–Ī–ł—ā–ĺ—Ä–į –ĺ—Ā–į–∂–ī–Ķ–Ĺ–ł—Ź –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–Ĺ–Ķ–ĺ—Ä–≥–į–Ĺ–ł—á–Ķ—Ā–ļ–ĺ–≥–ĺ —Ą–ĺ—Ā—Ą–į—ā–į –Ĺ–į¬†—Ā—ā–Ķ–Ĺ–ļ–į—Ö —Ā–ĺ—Ā—É–ī–ĺ–≤ [46]. –§–Ķ—ā—É–ł–Ĺ-–ź¬†—ā–į–ļ–∂–Ķ —Ā—ā–ł–ľ—É–Ľ–ł—Ä—É–Ķ—ā —Ą–į–≥–ĺ—Ü–ł—ā–ĺ–∑, —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É—Ź, —ā–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, —É–ī–į–Ľ–Ķ–Ĺ–ł—é —É–∂–Ķ –ĺ–Ī—Ä–į–∑–ĺ–≤–į–≤—ą–ł—Ö—Ā—Ź –Ĺ–Ķ—Ä–į—Ā—ā–≤–ĺ—Ä–ł–ľ—č—Ö –ĺ—Ā—ā–į—ā–ļ–ĺ–≤ –ļ–į–Ľ—Ć—Ü–ł—Ź. –≠–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ—č–Ķ –ľ–ĺ–ī–Ķ–Ľ–ł —Ā¬†–ī–Ķ–Ľ–Ķ—Ü–ł–Ķ–Ļ¬†–≥–Ķ–Ĺ–į —Ą–Ķ—ā—É–ł–Ĺ–į-–ź¬†–Ņ–ĺ–ļ–į–∑–į–Ľ–ł —Ö—Ä–ĺ–Ĺ–ł—á–Ķ—Ā–ļ—É—é –Ņ–ĺ—á–Ķ—á–Ĺ—É—é –Ĺ–Ķ–ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ—Ā—ā—Ć –Ĺ–į¬†—Ą–ĺ–Ĺ–Ķ –Ņ–ĺ–≤—Ā–Ķ–ľ–Ķ—Ā—ā–Ĺ–ĺ–Ļ —Ā–ĺ—Ā—É–ī–ł—Ā—ā–ĺ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł [47]. –Ě–ł–∑–ļ–ł–Ķ —É—Ä–ĺ–≤–Ĺ–ł —Ą–Ķ—ā—É–ł–Ĺ–į-–ź¬†–≤ —Ā—č–≤–ĺ—Ä–ĺ—ā–ļ–Ķ –ľ–ĺ–≥—É—ā –Ņ—Ä–Ķ–ī—Ä–į—Ā–Ņ–ĺ–Ľ–į–≥–į—ā—Ć –Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ —Ā¬†—Ö—Ä–ĺ–Ĺ–ł—á–Ķ—Ā–ļ–ĺ–Ļ –Ņ–ĺ—á–Ķ—á–Ĺ–ĺ–Ļ –Ĺ–Ķ–ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ—Ā—ā—Ć—é –ļ¬†–ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ [48]. –Ě–ł–∑–ļ–ł–Ķ —É—Ä–ĺ–≤–Ĺ–ł —Ą–Ķ—ā—É–ł–Ĺ–į-–ź¬†—ā–į–ļ–∂–Ķ –ĺ–Ī—É—Ā–Ľ–ĺ–≤–Ľ–ł–≤–į—é—ā –Ņ—Ä–ĺ–≥—Ä–Ķ—Ā—Ā–ł—Ä–ĺ–≤–į–Ĺ–ł–Ķ –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ņ–ĺ—Ä–į–∂–Ķ–Ĺ–ł–Ļ —Ā–Ķ—Ä–ī–Ķ—á–Ĺ–ĺ-—Ā–ĺ—Ā—É–ī–ł—Ā—ā–ĺ–Ļ —Ā–ł—Ā—ā–Ķ–ľ—č [49]. –ü–ĺ–ļ–į –Ĺ–Ķ¬†–ł–ľ–Ķ–Ķ—ā—Ā—Ź –ī–į–Ĺ–Ĺ—č—Ö –嬆–≤–Ľ–ł—Ź–Ĺ–ł–ł –≤–ł—ā–į–ľ–ł–Ĺ–į D –Ĺ–į¬†—ć–ļ—Ā–Ņ—Ä–Ķ—Ā—Ā–ł—é —Ą–Ķ—ā—É–ł–Ĺ–į-–ź.
–ö–Ľ–ĺ—āŠĹĻ¬†‚Äď –Ī–Ķ–Ľ–ļ–ĺ–≤—č–Ļ¬†–≥–ĺ—Ä–ľ–ĺ–Ĺ (—Ä–ł—Ā.¬†4), –ĺ–Ī—Č–ł–Ļ –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ļ —ć—Ą—Ą–Ķ–ļ—ā –ļ–ĺ—ā–ĺ—Ä–ĺ–≥–ĺ –ľ–ĺ–∂–Ķ—ā –Ī—č—ā—Ć —É—Ā–Ľ–ĺ–≤–Ĺ–ĺ –ĺ–Ņ–ł—Ā–į–Ĺ –ļ–į–ļ ¬ę–Ņ–ĺ–ī–į–≤–Ľ–Ķ–Ĺ–ł–Ķ —Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź¬Ľ [50]. –ė–∑–≤–Ķ—Ā—ā–Ĺ—č –ī–≤–Ķ —Ą–ĺ—Ä–ľ—č –Ī–Ķ–Ľ–ļ–į –ļ–Ľ–ĺ—ā–嬆‚Äď –ľ–Ķ–ľ–Ī—Ä–į–Ĺ–Ĺ–į—Ź –ł¬†—Ā–Ķ–ļ—Ä–Ķ—ā–ł—Ä—É–Ķ–ľ–į—Ź. –ú–Ķ–ľ–Ī—Ä–į–Ĺ–Ĺ–į—Ź —Ą–ĺ—Ä–ľ–į –ĺ–Ī—Ä–į–∑—É–Ķ—ā –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ļ—Ā —Ā¬†—Ą–į–ļ—ā–ĺ—Ä–į–ľ–ł —Ä–ĺ—Ā—ā–į —Ą–ł–Ī—Ä–ĺ–Ī–Ľ–į—Ā—ā–ĺ–≤, —á—ā–ĺ —É—Ā–ł–Ľ–ł–≤–į–Ķ—ā –≤—č–ī–Ķ–Ľ–Ķ–Ĺ–ł–Ķ —Ą–ĺ—Ā—Ą–į—ā–į –≤¬†–ľ–ĺ—á–Ķ. –°–Ķ–ļ—Ä–Ķ—ā–ł—Ä—É–Ķ–ľ–į—Ź —Ą–ĺ—Ä–ľ–į –Ī–Ķ–Ľ–ļ–į —Ä–Ķ–≥—É–Ľ–ł—Ä—É–Ķ—ā –ľ–Ĺ–ĺ–≥–ĺ—á–ł—Ā–Ľ–Ķ–Ĺ–Ĺ—č–Ķ¬†–≥–Ľ–ł–ļ–ĺ–Ņ—Ä–ĺ—ā–Ķ–ł–Ĺ—č –Ĺ–į¬†–Ņ–ĺ–≤–Ķ—Ä—Ö–Ĺ–ĺ—Ā—ā–ł –ļ–Ľ–Ķ—ā–ĺ–ļ, –≤–ļ–Ľ—é—á–į—Ź —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä—č —Ą–į–ļ—ā–ĺ—Ä–ĺ–≤ —Ä–ĺ—Ā—ā–į, –≤¬†—ā–ĺ–ľ —á–ł—Ā–Ľ–Ķ —Ą–į–ļ—ā–ĺ—Ä–ĺ–≤ —Ä–ĺ—Ā—ā–į –ł–Ĺ—Ā—É–Ľ–ł–Ĺ–į (IGF) [51].
–ú—č—ą–ł —Ā¬†—Ü–Ķ–Ľ–Ķ–≤—č–ľ–ł –ī–Ķ–Ľ–Ķ—Ü–ł—Ź–ľ–ł¬†–≥–Ķ–Ĺ–į klotho –ĺ–Ī–Ľ–į–ī–į–Ľ–ł –Ī–ĺ–Ľ–Ķ–Ķ –ļ–ĺ—Ä–ĺ—ā–ļ–ĺ–Ļ –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā—Ć—é –∂–ł–∑–Ĺ–ł –ł, —á—ā–ĺ —Ö–į—Ä–į–ļ—ā–Ķ—Ä–Ĺ–ĺ, –ī–Ķ–ľ–ĺ–Ĺ—Ā—ā—Ä–ł—Ä–ĺ–≤–į–Ľ–ł –ľ–Ĺ–ĺ–≥–ł–Ķ –ĺ—Ā–ĺ–Ī–Ķ–Ĺ–Ĺ–ĺ—Ā—ā–ł –Ņ—Ä–Ķ–∂–ī–Ķ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ —Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź, –≤¬†—ā–ĺ–ľ —á–ł—Ā–Ľ–Ķ –Ī–Ķ—Ā–Ņ–Ľ–ĺ–ī–ł–Ķ, –į—ā—Ä–ĺ—Ą–ł—é –≤–ł–Ľ–ĺ—á–ļ–ĺ–≤–ĺ–Ļ –∂–Ķ–Ľ–Ķ–∑—č, —Ā—ā–į—Ä–Ķ–Ĺ–ł–Ķ –ļ–ĺ–∂–ł, —É–ľ–Ķ–Ĺ—Ć—ą–Ķ–Ĺ–ł–Ķ –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–į –ļ–Ľ–Ķ—ā–ĺ–ļ –ü—É—Ä–ļ–ł–Ĺ—Ć–Ķ, —Ą–ł–∑–ł—á–Ķ—Ā–ļ—É—é –ł–Ĺ–Ķ—Ä—ā–Ĺ–ĺ—Ā—ā—Ć, —ć–ľ—Ą–ł–∑–Ķ–ľ—É –Ľ–Ķ–≥–ļ–ł—Ö, —ā—Ź–∂–Ķ–Ľ—č–Ļ –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑ –ł¬†–Ņ—Ä–ĺ–≥—Ä–Ķ—Ā—Ā–ł—Ä—É—é—Č–ł–Ļ –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑ —Ā¬†–ľ–Ķ–ī–ł–į–Ľ—Ć–Ĺ–ĺ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–Ķ–Ļ. –°–ĺ—Ā—É–ī–ł—Ā—ā—č–Ķ –Ĺ–į—Ä—É—ą–Ķ–Ĺ–ł—Ź —ɬ†—ā—Ä–į–Ĺ—Ā–≥–Ķ–Ĺ–Ĺ—č—Ö –∂–ł–≤–ĺ—ā–Ĺ—č—Ö –Ņ–ĺ–Ľ–Ĺ–ĺ—Ā—ā—Ć—é –ł—Ā—á–Ķ–∑–į–Ľ–ł –Ņ—Ä–ł¬†–≥–Ķ–Ĺ–Ĺ–ĺ–Ļ —ā–Ķ—Ä–į–Ņ–ł–ł –Ņ–ĺ—Ā—Ä–Ķ–ī—Ā—ā–≤–ĺ–ľ –≤–Ĺ–Ķ–ī—Ä–Ķ–Ĺ–ł—Ź –≤¬†–ĺ—Ä–≥–į–Ĺ–ł–∑–ľ –∂–ł–≤–ĺ—ā–Ĺ–ĺ–≥–嬆–≥–Ķ–Ĺ–į klotho [52]. –ď–Ķ–Ĺ–Ķ—ā–ł—á–Ķ—Ā–ļ–ł–Ļ –Ņ–ĺ–Ľ–ł–ľ–ĺ—Ä—Ą–ł–∑–ľ –≤¬†–Ņ—Ä–ĺ–ľ–ĺ—ā–ĺ—Ä–Ĺ–ĺ–Ļ –ĺ–Ī–Ľ–į—Ā—ā–ł¬†–≥–Ķ–Ĺ–į klotho (G395A) –Ī—č–Ľ –į—Ā—Ā–ĺ—Ü–ł–ł—Ä–ĺ–≤–į–Ĺ —Ā¬†–ľ–ł–Ĺ–Ķ—Ä–į–Ľ—Ć–Ĺ–ĺ–Ļ –Ņ–Ľ–ĺ—ā–Ĺ–ĺ—Ā—ā—Ć—é –ļ–ĺ—Ā—ā–Ķ–Ļ —ɬ†–∂–Ķ–Ĺ—Č–ł–Ĺ –≤¬†–Ņ–ĺ—Ā—ā–ľ–Ķ–Ĺ–ĺ–Ņ–į—É–∑–Ķ, –ĺ–Ĺ —ā–į–ļ–∂–Ķ –≤–Ľ–ł—Ź–Ķ—ā –Ĺ–į¬†—Ä–ł—Ā–ļ —Ä–į–∑–≤–ł—ā–ł—Ź –ė–Ď–° [53]. –†–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä –≤–ł—ā–į–ľ–ł–Ĺ–į D —Ā–≤—Ź–∑—č–≤–į–Ķ—ā 1,25-–ī–ł–≥–ł–ī—Ä–ĺ–ļ—Ā–ł–≤–ł—ā–į–ľ–ł–Ĺ-D3 –ł¬†—Ä–Ķ–≥—É–Ľ–ł—Ä—É–Ķ—ā —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł—鬆–≥–Ķ–Ĺ–į klotho [54], —ā–Ķ–ľ —Ā–į–ľ—č–ľ –∑–į–ľ–Ķ–ī–Ľ—Ź—Ź –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—é –į—Ä—ā–Ķ—Ä–ł–Ļ.
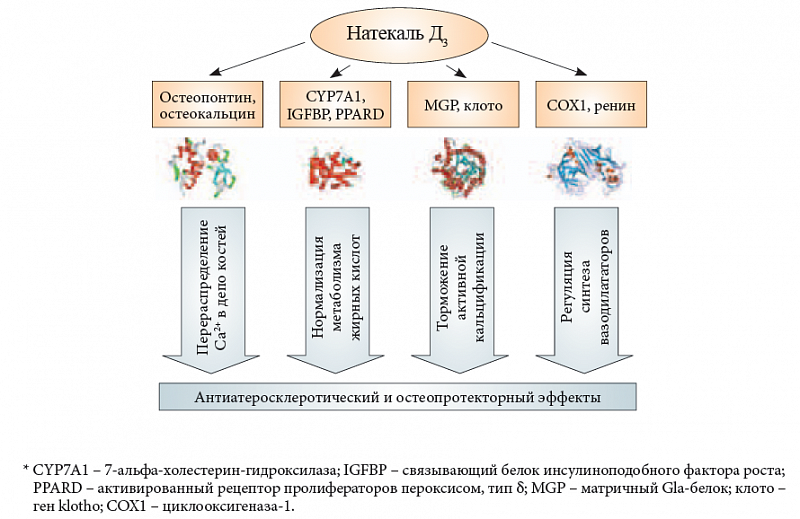
–Ę–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –į–Ĺ–į–Ľ–ł–∑–į —É–ļ–į–∑—č–≤–į—é—ā –Ĺ–į¬†–ļ–ĺ–Ĺ–ļ—Ä–Ķ—ā–Ĺ—č–Ķ –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ—č, –ĺ—Ā—É—Č–Ķ—Ā—ā–≤–Ľ—Ź—é—Č–ł–Ķ –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ļ—Ā–Ĺ–ĺ–Ķ –≤–ĺ–∑–ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ –≤–ł—ā–į–ľ–ł–Ĺ–į D –Ĺ–į¬†–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –ł¬†–ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł (—Ä–ł—Ā.¬†5). –í–ĺ-–Ņ–Ķ—Ä–≤—č—Ö, –≤–ł—ā–į–ľ–ł–Ĺ D ¬ę–Ĺ–į–Ņ—Ä–į–≤–Ľ—Ź–Ķ—ā¬Ľ –Ņ–ĺ—ā–ĺ–ļ–ł –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†–ī–Ķ–Ņ–ĺ –ļ–ĺ—Ā—ā–Ķ–Ļ (–ĺ—Ā—ā–Ķ–ĺ–ļ–į–Ľ—Ć—Ü–ł–Ĺ, –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ–Ĺ—ā–ł–Ĺ), —ā–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É—Ź —É–ī–į–Ľ–Ķ–Ĺ–ł—é –ł–∑–Ī—č—ā–ļ–į –ļ–į–Ľ—Ć—Ü–ł—Ź –ł–∑¬†–Ņ–Ľ–į–∑–ľ—č –ł, –ĺ–ī–Ĺ–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ, –Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł–ļ–Ķ –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į. –í–ĺ-–≤—ā–ĺ—Ä—č—Ö, –≤–ł—ā–į–ľ–ł–Ĺ D –į–ļ—ā–ł–≤–ł—Ä—É–Ķ—ā¬†–≥–Ķ–Ĺ—č, –ļ–ĺ–ī–ł—Ä—É—é—Č–ł–Ķ –Ī–Ķ–Ľ–ļ–ł (7-–į–Ľ—Ć—Ą–į-—Ö–ĺ–Ľ–Ķ—Ā—ā–Ķ—Ä–ł–Ĺ-–≥–ł–ī—Ä–ĺ–ļ—Ā–ł–Ľ–į–∑–į, —Ā–≤—Ź–∑—č–≤–į—é—Č–ł–Ķ –Ī–Ķ–Ľ–ļ–ł –ł–Ĺ—Ā—É–Ľ–ł–Ĺ–ĺ–Ņ–ĺ–ī–ĺ–Ī–Ĺ–ĺ–≥–ĺ —Ą–į–ļ—ā–ĺ—Ä–į —Ä–ĺ—Ā—ā–į, –į–ļ—ā–ł–≤–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č–Ļ —Ä–Ķ—Ü–Ķ–Ņ—ā–ĺ—Ä –ī–Ķ–Ľ—Ć—ā–į –Ņ—Ä–ĺ–Ľ–ł—Ą–Ķ—Ä–į—ā–ĺ—Ä–ĺ–≤ –Ņ–Ķ—Ä–ĺ–ļ—Ā–ł—Ā–ĺ–ľ), –ļ–ĺ—ā–ĺ—Ä—č–Ķ —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É—é—ā –Ĺ–ĺ—Ä–ľ–į–Ľ–ł–∑–į—Ü–ł–ł –ľ–Ķ—ā–į–Ī–ĺ–Ľ–ł–∑–ľ–į –∂–ł—Ä–Ĺ—č—Ö –ļ–ł—Ā–Ľ–ĺ—ā. –í-—ā—Ä–Ķ—ā—Ć–ł—Ö, –≤–ł—ā–į–ľ–ł–Ĺ D –ľ–ĺ–ī—É–Ľ–ł—Ä—É–Ķ—ā —ā—Ä–į–Ĺ—Ā–ļ—Ä–ł–Ņ—Ü–ł—鬆–≥–Ķ–Ĺ–ĺ–≤ –≤–ĺ–≤–Ľ–Ķ—á–Ķ–Ĺ–Ĺ—č—Ö –≤¬†–į–ļ—ā–ł–≤–Ĺ—É—é –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł—é –Ī–Ķ–Ľ–ļ–ĺ–≤ (–ľ–į—ā—Ä–ł—á–Ĺ—č–Ļ Gla-–Ī–Ķ–Ľ–ĺ–ļ, –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ–Ĺ—ā–ł–Ĺ, –ļ–Ľ–ĺ—ā–ĺ), –∑–į–ľ–Ķ–ī–Ľ—Ź—Ź –Ņ–į—ā–ĺ—Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č, –≤–Ķ–ī—É—Č–ł–Ķ –ļ¬†–ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ. –í-—á–Ķ—ā–≤–Ķ—Ä—ā—č—Ö, –≤–ł—ā–į–ľ–ł–Ĺ D —Ä–Ķ–≥—É–Ľ–ł—Ä—É–Ķ—ā —Ā–ł–Ĺ—ā–Ķ–∑ –≤–į–∑–ĺ–ī–ł–Ľ–į—ā–į—ā–ĺ—Ä–ĺ–≤ (—É–≤–Ķ–Ľ–ł—á–Ķ–Ĺ–ł–Ķ –Ņ—Ä–ĺ—Ā—ā–į—Ü–ł–ļ–Ľ–ł–Ĺ–į, —É–ľ–Ķ–Ĺ—Ć—ą–Ķ–Ĺ–ł–Ķ —Ä–Ķ–Ĺ–ł–Ĺ–į). –í—Ā–Ķ —ć—ā–ł —ć—Ą—Ą–Ķ–ļ—ā—č —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É—é—ā –Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł–ļ–Ķ –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į, –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ –ł¬†—ā—Ä–ĺ–ľ–Ī–ĺ–∑–ĺ–≤.
–ü—Ä–Ķ–Ņ–į—Ä–į—ā—č –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D3, —ā–Ķ—Ä–į–Ņ–ł—Ź –ł¬†–Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł–ļ–į –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į
–Ē–Ľ—Ź –Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł–ļ–ł –ļ–į–Ľ—Ć—Ü–ł–Ķ–≤–ĺ–≥–ĺ –ī–Ķ—Ą–ł—Ü–ł—ā–į —Ö–ĺ—Ä–ĺ—ą–ĺ –Ņ–ĺ–ī—Ö–ĺ–ī—Ź—ā –ļ–ĺ–ľ–Ņ–Ľ–Ķ–ļ—Ā–Ĺ—č–Ķ –ļ–į–Ľ—Ć—Ü–ł–Ķ–≤—č–Ķ –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā—č —Ā¬†–Ĺ–Ķ–≤—č—Ā–ĺ–ļ–ĺ–Ļ –ī–ĺ–∑–ł—Ä–ĺ–≤–ļ–ĺ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–ī–ĺ–Ņ–ĺ–Ľ–Ĺ–Ķ–Ĺ–Ĺ—č–Ķ –Ņ–ĺ—ā–Ķ–Ĺ—Ü–ł–į—ā–ĺ—Ä–į–ľ–ł –Ķ–≥–ĺ –ĺ–Ī–ľ–Ķ–Ĺ–į –ł¬†—É—Ā–≤–ĺ–Ķ–Ĺ–ł—Ź. –Ē–Ľ—Ź —ā–Ķ—Ä–į–Ņ–ł–ł –ī–Ķ—Ą–ł—Ü–ł—ā–į –ļ–į–Ľ—Ć—Ü–ł—Ź, –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–į –ł, –≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ, –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –Ī–ĺ–Ľ–Ķ–Ķ —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–ł–Ķ —Ā–Ņ–Ķ—Ü–ł–į–Ľ—Ć–Ĺ—č—Ö –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤ —Ā¬†–Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ—č–ľ–ł –ī–ĺ–∑–į–ľ–ł –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D3¬†‚Äď –Ě–į—ā–Ķ–ļ–į–Ľ—Ć –Ē3 (1500 –ľ–≥ –ļ–į—Ä–Ī–ĺ–Ĺ–į—ā–į –ļ–į–Ľ—Ć—Ü–ł—Ź, —á—ā–ĺ —ć–ļ–≤–ł–≤–į–Ľ–Ķ–Ĺ—ā–Ĺ–ĺ 600 –ľ–≥ –°–į2+ –ł¬†400 –ú–ē —Ö–ĺ–Ľ–Ķ–ļ–į–Ľ—Ć—Ü–ł—Ą–Ķ—Ä–ĺ–Ľ–į), –í–ł—ā—Ä—É–ľ –ļ–į–Ľ—Ć—Ü–ł–Ļ (–ļ–į—Ä–Ī–ĺ–Ĺ–į—ā –ļ–į–Ľ—Ć—Ü–ł—Ź¬†‚Äď 1250¬†–ľ–≥, –≤–ł—ā–į–ľ–ł–Ĺ D3¬†‚Äď 200 –ú–ē) –ł¬†–ė–ī–Ķ–ĺ—Ā (–ļ–į—Ä–Ī–ĺ–Ĺ–į—ā –ļ–į–Ľ—Ć—Ü–ł—Ź¬†‚Äď 1250 –ľ–≥, –≤–ł—ā–į–ľ–ł–Ĺ D3¬†‚Äď 400 –ú–ē). –£—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–Ĺ–į—Ź –≤¬†–†–§ –Ņ–ĺ—ā—Ä–Ķ–Ī–Ĺ–ĺ—Ā—ā—Ć –≤¬†–ļ–į–Ľ—Ć—Ü–ł–ł –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–Ķ D3 —ɬ†–Ľ–ł—Ü —Ā—ā–į—Ä—ą–Ķ 60 –Ľ–Ķ—ā —Ā–ĺ—Ā—ā–į–≤–Ľ—Ź–Ķ—ā 1200 –°–į2+ –ľ–≥/—Ā—É—ā –ł¬†600‚Äď800 ¬†–ú–ē/—Ā—É—ā —Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ.
–í¬†–Ņ–ĺ–∂–ł–Ľ–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ –ĺ—á–Ķ–Ĺ—Ć –≤–į–∂–Ĺ–ĺ –≤—č–Ī—Ä–į—ā—Ć –Ņ—Ä–ł–Ķ–ľ–Ľ–Ķ–ľ—É—é —Ą–ĺ—Ä–ľ—É –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–į –ļ–į–Ľ—Ć—Ü–ł—Ź. –Ē–Ķ–Ľ–ĺ –≤¬†—ā–ĺ–ľ, —á—ā–ĺ –ľ–Ĺ–ĺ–≥–ł–Ķ —ā–į–Ī–Ľ–Ķ—ā–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č–Ķ —Ą–ĺ—Ä–ľ—č –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤ –ļ–į–Ľ—Ć—Ü–ł—Ź –ĺ—ā–Ľ–ł—á–į—é—ā—Ā—Ź —ā–≤–Ķ—Ä–ī–ĺ—Ā—ā—Ć—é, –ł¬†—ć—ā–ĺ –ī–Ķ–Ľ–į–Ķ—ā –ł—Ö —É–Ņ–ĺ—ā—Ä–Ķ–Ī–Ľ–Ķ–Ĺ–ł–Ķ –∑–į—ā—Ä—É–ī–Ĺ–ł—ā–Ķ–Ľ—Ć–Ĺ—č–ľ —ɬ†–Ľ–ł—Ü –Ņ–ĺ–∂–ł–Ľ–ĺ–≥–ĺ –≤–ĺ–∑—Ä–į—Ā—ā–į. –ö—Ä–ĺ–ľ–Ķ —ā–ĺ–≥–ĺ, –Ņ—Ä–ł —Ä–į–∑–∂–Ķ–≤—č–≤–į–Ĺ–ł–ł —ā–į–Ī–Ľ–Ķ—ā–ļ–ł –Ņ—Ä–ĺ—Ź–≤–Ľ—Ź—é—ā –į–Ī—Ä–į–∑–ł–≤–Ĺ—č–Ķ —Ā–≤–ĺ–Ļ—Ā—ā–≤–į, —á—ā–ĺ —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É–Ķ—ā –Ņ–ĺ–≤—Ä–Ķ–∂–ī–Ķ–Ĺ–ł—é —ć–ľ–į–Ľ–ł –∑—É–Ī–ĺ–≤. –ė–ľ–Ķ–Ĺ–Ĺ–ĺ –Ņ–ĺ—ć—ā–ĺ–ľ—É –ī–Ľ—Ź –Ņ–ĺ–∂–ł–Ľ—č—Ö –Ľ—é–ī–Ķ–Ļ –ł¬†–Ņ–į—Ü–ł–Ķ–Ĺ—ā–ĺ–≤ —Ā–嬆—Ā–Ľ–į–Ī–ĺ–Ļ —ć–ľ–į–Ľ—Ć—é –∑—É–Ī–ĺ–≤ –Ņ—Ä–Ķ–ī–Ņ–ĺ—á—ā–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į—ā—Ć –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā—č –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D3 –≤¬†—Ą–ĺ—Ä–ľ–Ķ —ā–į–Ī–Ľ–Ķ—ā–ĺ–ļ –ī–Ľ—Ź —Ä–į—Ā—Ā–į—Ā—č–≤–į–Ĺ–ł—Ź (–Ě–į—ā–Ķ–ļ–į–Ľ—Ć –Ē3).
–Ě–į—ā–Ķ–ļ–į–Ľ—Ć –Ē3, –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D –≤¬†—Ą–ĺ—Ä–ľ–Ķ —ā–į–Ī–Ľ–Ķ—ā–ĺ–ļ –ī–Ľ—Ź —Ä–į—Ā—Ā–į—Ā—č–≤–į–Ĺ–ł—Ź, –Ĺ–Ķ¬†—ā–ĺ–Ľ—Ć–ļ–ĺ –Ĺ–Ķ¬†–Ņ—Ä–ĺ—Ź–≤–Ľ—Ź–Ķ—ā –į–Ī—Ä–į–∑–ł–≤–Ĺ—č–Ķ —Ā–≤–ĺ–Ļ—Ā—ā–≤–į, –Ŗ嬆–ł —Ā–ĺ–∑–ī–į–Ķ—ā –≤—č—Ā–ĺ–ļ–ł–Ķ –ļ–ĺ–Ĺ—Ü–Ķ–Ĺ—ā—Ä–į—Ü–ł–ł –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–Ķ–≥–ĺ —Ā–ł–Ĺ–Ķ—Ä–≥–ł—Ā—ā–į –≤–ł—ā–į–ľ–ł–Ĺ–į D –≤¬†—Ā–Ľ—é–Ĺ–Ķ. –ě—ā–ľ–Ķ—ā–ł–ľ: –嬆—ā–ĺ–ľ, —á—ā–ĺ –Ĺ–ł–∑–ļ–ĺ–Ķ —Ā–ĺ–ī–Ķ—Ä–∂–į–Ĺ–ł–Ķ –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†—Ā–Ľ—é–Ĺ–Ķ —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź —Ä–ł—Ā–ļ–ĺ–ľ —Ä–į–∑–≤–ł—ā–ł—Ź –ļ–į—Ä–ł–Ķ—Ā–į, –ł–∑–≤–Ķ—Ā—ā–Ĺ–ĺ –Ņ–嬆–ľ–Ķ–Ĺ—Ć—ą–Ķ–Ļ –ľ–Ķ—Ä–Ķ 80 –Ľ–Ķ—ā [55]. –ü–ĺ–ī–ī–Ķ—Ä–∂–į–Ĺ–ł–Ķ –ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–ĺ–Ļ –ļ–ĺ–Ĺ—Ü–Ķ–Ĺ—ā—Ä–į—Ü–ł–ł –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†—Ā–Ľ—é–Ĺ–Ķ —Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –≤–į–∂–Ĺ—č–ľ –Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł—á–Ķ—Ā–ļ–ł–ľ –ľ–Ķ—Ä–ĺ–Ņ—Ä–ł—Ź—ā–ł–Ķ–ľ, –∑–į—Č–ł—Č–į—é—Č–ł–ľ –ĺ—ā¬†—Ä–į–∑–≤–ł—ā–ł—Ź –ļ–į—Ä–ł–Ķ—Ā–į –ł¬†–Ņ–į—Ä–ĺ–ī–ĺ–Ĺ—ā–ĺ–∑–į, –Ņ–ĺ–≤—č—ą–į—é—Č–ł–ľ —Ā—Ä–ĺ–ļ —Ā–Ľ—É–∂–Ī—č –Ņ–ĺ—Ā—ā–ĺ—Ź–Ĺ–Ĺ—č—Ö –∑—É–Ī–Ĺ—č—Ö –Ņ—Ä–ĺ—ā–Ķ–∑–ĺ–≤. –Ě–į–Ņ—Ä–ł–ľ–Ķ—Ä, –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–ł–Ķ –∂–Ķ–≤–į—ā–Ķ–Ľ—Ć–Ĺ—č—Ö —Ä–Ķ–∑–ł–Ĺ–ĺ–ļ —Ā¬†–Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ—č–ľ —Ā–ĺ–ī–Ķ—Ä–∂–į–Ĺ–ł–Ķ–ľ –ļ–į–Ľ—Ć—Ü–ł—Ź —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É–Ķ—ā —Ä–Ķ–ľ–ł–Ĺ–Ķ—Ä–į–Ľ–ł–∑–į—Ü–ł–ł –Ņ–ĺ–≤—Ä–Ķ–∂–ī–Ķ–Ĺ–ł–Ļ —ć–ľ–į–Ľ–ł [56]. –ú–ĺ–∂–Ķ—ā –Ņ–ĺ–ļ–į–∑–į—ā—Ć—Ā—Ź –Ņ–į—Ä–į–ī–ĺ–ļ—Ā–į–Ľ—Ć–Ĺ—č–ľ, –Ŗ嬆–Ņ–ĺ–ī–ī–Ķ—Ä–∂–į–Ĺ–ł–Ķ –ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ—č—Ö –ļ–ĺ–Ĺ—Ü–Ķ–Ĺ—ā—Ä–į—Ü–ł–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†—Ā–Ľ—é–Ĺ–Ķ —ā–į–ļ–∂–Ķ —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É–Ķ—ā —ā–Ķ—Ä–į–Ņ–ł–ł –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –Ņ–嬆—Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ĺ–ľ—É –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ—É, –Ĺ–Ķ–∑–į–≤–ł—Ā–ł–ľ–ĺ–ľ—É –ĺ—ā¬†—Ä–į—Ā—Ā–ľ–ĺ—ā—Ä–Ķ–Ĺ–Ĺ—č—Ö –≤—č—ą–Ķ –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ—Ź—Ä–Ĺ—č—Ö –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–ĺ–≤ –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ĺ–≥–ĺ –ī–Ķ–Ļ—Ā—ā–≤–ł—Ź –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D.
–Ē–Ķ–Ľ–ĺ –≤¬†—ā–ĺ–ľ, —á—ā–ĺ –ļ–į—Ä–ł–Ķ—Ā –∑—É–Ī–ĺ–≤ –Ĺ–Ķ—Ä–į–∑—Ä—č–≤–Ĺ–ĺ —Ā–≤—Ź–∑–į–Ĺ —Ā¬†–ľ–ł–Ĺ–Ķ—Ä–į–Ľ—Ć–Ĺ—č–ľ –ĺ–Ī–ľ–Ķ–Ĺ–ĺ–ľ¬†‚Äď –ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–Ķ–Ĺ–Ĺ–ĺ—Ā—ā—Ć—é –ļ–į–Ľ—Ć—Ü–ł–Ķ–ľ, –ľ–į–≥–Ĺ–ł–Ķ–ľ, –Ņ–ł—Ä–ł–ī–ĺ–ļ—Ā–ł–Ĺ–ĺ–ľ, —Ü–ł–Ĺ–ļ–ĺ–ľ, —Ą—ā–ĺ—Ä–ĺ–ľ. –≠—ā–ĺ—ā –Ņ–į—ā–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ļ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā –Ņ—Ä–ĺ—Ź–≤–Ľ—Ź–Ķ—ā—Ā—Ź –ī–Ķ–ľ–ł–Ĺ–Ķ—Ä–į–Ľ–ł–∑–į—Ü–ł–Ķ–Ļ –ł¬†–Ņ–ĺ—Ā–Ľ–Ķ–ī—É—é—Č–ł–ľ —Ä–į–∑—Ä—É—ą–Ķ–Ĺ–ł–Ķ–ľ —ā–≤–Ķ—Ä–ī—č—Ö —ā–ļ–į–Ĺ–Ķ–Ļ –∑—É–Ī–į —Ā¬†–ĺ–Ī—Ä–į–∑–ĺ–≤–į–Ĺ–ł–Ķ–ľ –ī–Ķ—Ą–Ķ–ļ—ā–į –≤¬†–≤–ł–ī–Ķ –Ņ–ĺ–Ľ–ĺ—Ā—ā–ł. –≠–ľ–į–Ľ—Ć –∑—É–Ī–į –Ņ–ĺ–≤—Ä–Ķ–∂–ī–į–Ķ—ā—Ā—Ź –Ņ—Ä–ł –ī–Ķ–Ļ—Ā—ā–≤–ł–ł –ľ–ĺ–Ľ–ĺ—á–Ĺ–ĺ–Ļ –ł¬†–Ņ–ł—Ä–ĺ–≤–ł–Ĺ–ĺ–≥—Ä–į–ī–Ĺ–ĺ–Ļ –ļ–ł—Ā–Ľ–ĺ—ā, –ĺ–Ī—Ä–į–∑—É—é—Č–ł—Ö—Ā—Ź –≤—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł–Ķ —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā–į—Ü–ł–ł –ľ–ĺ–Ĺ–ĺ- –ł¬†–ī–ł—Ā–į—Ö–į—Ä–ł–ī–ĺ–≤ –ľ–ł–ļ—Ä–ĺ–ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į–ľ–ł –∑—É–Ī–Ĺ–ĺ–≥–ĺ –Ĺ–į–Ľ–Ķ—ā–į. –ü–嬆–ľ–Ķ—Ä–Ķ —Ä–į–∑–≤–ł—ā–ł—Ź –ļ–į—Ä–ł–Ķ—Ā–į –Ī–į–ļ—ā–Ķ—Ä–ł–ł –≤—Ā–Ķ¬†–≥–Ľ—É–Ī–∂–Ķ –ł¬†–≥–Ľ—É–Ī–∂–Ķ –Ņ—Ä–ĺ–Ĺ–ł–ļ–į—é—ā –≤¬†—ā–ļ–į–Ĺ–ł –∑—É–Ī–į, —Ä–į–∑—Ä—É—ą–į—é—ā –ī–Ķ–Ĺ—ā–ł–Ĺ –ł¬†–Ņ–ĺ–Ņ–į–ī–į—é—ā –≤¬†–ļ—Ä–ĺ–≤—Ź–Ĺ–ĺ–Ķ —Ä—É—Ā–Ľ–ĺ. –ź–ī—Ā–ĺ—Ä–Ī–ł—Ä—É—Ź—Ā—Ć –Ĺ–į¬†—ć–Ĺ–ī–ĺ—ā–Ķ–Ľ–ł–ł —Ā–ĺ—Ā—É–ī–ĺ–≤, —ć—ā–ł –Ī–į–ļ—ā–Ķ—Ä–ł–ł —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É—é—ā —Ä–ĺ—Ā—ā—É –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ī–Ľ—Ź—ą–Ķ–ļ. –Ę–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, –Ņ–ĺ–ī–ī–Ķ—Ä–∂–į–Ĺ–ł–Ķ –ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ—č—Ö —É—Ä–ĺ–≤–Ĺ–Ķ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†—Ā–Ľ—é–Ĺ–Ķ —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É–Ķ—ā –Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł–ļ–Ķ –ļ–į—Ä–ł–Ķ—Ā–į –ł¬†—É–ľ–Ķ–Ĺ—Ć—ą–Ķ–Ĺ–ł—é –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–į –Ī–į–ļ—ā–Ķ—Ä–ł–Ļ. –ü–ĺ—Ā–ļ–ĺ–Ľ—Ć–ļ—É –ļ–į—Ä–ł–Ķ—Ā —Ā–≤—Ź–∑–į–Ĺ —Ā¬†–Ņ–ĺ–≤—č—ą–Ķ–Ĺ–Ĺ—č–ľ —Ä–ł—Ā–ļ–ĺ–ľ –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –ł¬†–ė–Ď–° [57], –Ņ–ĺ–ī–ī–Ķ—Ä–∂–į–Ĺ–ł–Ķ –ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ—č—Ö —É—Ä–ĺ–≤–Ĺ–Ķ–Ļ –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†—Ā–Ľ—é–Ĺ–Ķ —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤—É–Ķ—ā –ł¬†—Ā–Ĺ–ł–∂–Ķ–Ĺ–ł—é —Ä–ł—Ā–ļ–į –ĺ–Ī—Ä–į–∑–ĺ–≤–į–Ĺ–ł—Ź –Ĺ–ĺ–≤—č—Ö –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ī–Ľ—Ź—ą–Ķ–ļ.
–ó–į–ļ–Ľ—é—á–Ķ–Ĺ–ł–Ķ
–ü—Ä–ł —Ā–ĺ–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ–ľ –ĺ–Ī—Ä–į–∑–Ķ –∂–ł–∑–Ĺ–ł (–≥–ł–Ņ–ĺ–ī–ł–Ĺ–į–ľ–ł—Ź, –Ĺ–Ķ–ī–ĺ—Ā—ā–į—ā–ĺ—á–Ĺ–į—Ź –ł–Ĺ—Ā–ĺ–Ľ—Ź—Ü–ł—Ź, –Ĺ–Ķ—Ä–į—Ü–ł–ĺ–Ĺ–į–Ľ—Ć–Ĺ–ĺ–Ķ –Ņ–ł—ā–į–Ĺ–ł–Ķ: –Ņ–ĺ—ā—Ä–Ķ–Ī–Ľ–Ķ–Ĺ–ł–Ķ –Ņ—Ä–ĺ–ī—É–ļ—ā–ĺ–≤ —Ā¬†–≤—č—Ā–ĺ–ļ–ł–ľ —Ā–ĺ–ī–Ķ—Ä–∂–į–Ĺ–ł–Ķ–ľ —Ą–ĺ—Ā—Ą–ĺ—Ä–į, —ā–į–ļ–ł—Ö –ļ–į–ļ —Ā–ĺ—Ā–ł—Ā–ļ–ł, –ļ–ĺ–Ľ–Ī–į—Ā—č; –Ĺ–Ķ–ī–ĺ—Ā—ā–į—ā–ĺ–ļ –≤¬†—Ä–į—Ü–ł–ĺ–Ĺ–Ķ —Ā–≤–Ķ–∂–Ķ–Ļ –∑–Ķ–Ľ–Ķ–Ĺ–ł, –ļ–į–Ľ—Ć—Ü–ł—Ź, –≤–ł—ā–į–ľ–ł–Ĺ–į D –ł¬†—ā.–ī.) —Ä–ł—Ā–ļ¬†–ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–į –ł¬†–ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ —Ä–Ķ–∑–ļ–ĺ –≤–ĺ–∑—Ä–į—Ā—ā–į–Ķ—ā. –Ě–ĺ–Ī—Ö–ĺ–ī–ł–ľ–ĺ—Ā—ā—Ć —ā–Ķ—Ä–į–Ņ–ł–ł –ł¬†–Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł–ļ–ł –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–į –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–į–ľ–ł –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–Ķ–≥–ĺ –ĺ—Ā–Ĺ–ĺ–≤–Ĺ–ĺ–≥–ĺ —Ā–ł–Ĺ–Ķ—Ä–≥–ł—Ā—ā–į –≤–ł—ā–į–ľ–ł–Ĺ–į D –ĺ–Ī—Č–Ķ–Ņ—Ä–ł–∑–Ĺ–į–Ĺ–į. –í¬†—ā–ĺ –∂–Ķ –≤—Ä–Ķ–ľ—Ź –Ņ—Ä–ł–ľ–Ķ–Ĺ–Ķ–Ĺ–ł–Ķ —ć—ā–ł—Ö –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤ –ī–Ľ—Ź —ā–Ķ—Ä–į–Ņ–ł–ł –ł¬†–Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł–ļ–ł –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –≤—Ā–Ķ –Ķ—Č–Ķ —Ā—á–ł—ā–į–Ķ—ā—Ā—Ź —Ā–≤–ĺ–Ķ–≥–ĺ —Ä–ĺ–ī–į —ā–į–Ī—É —Ā—Ä–Ķ–ī–ł –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö –≤—Ä–į—á–Ķ–Ļ. –í¬†–Ĺ–į—Ā—ā–ĺ—Ź—Č–Ķ–Ļ —Ä–į–Ī–ĺ—ā–Ķ –Ī—č–Ľ–į –Ņ—Ä–ĺ–į–Ĺ–į–Ľ–ł–∑–ł—Ä–ĺ–≤–į–Ĺ–į –≤–∑–į–ł–ľ–ĺ—Ā–≤—Ź–∑—Ć –ľ–Ķ–∂–ī—É –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–Ķ–Ļ, –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–ĺ–ľ –ł¬†–Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–ľ–ł —ć—Ą—Ą–Ķ–ļ—ā–į–ľ–ł –≤–ł—ā–į–ľ–ł–Ĺ–į D. –ź–Ĺ–į–Ľ–ł–∑ –Ņ–ĺ–∑–≤–ĺ–Ľ–ł–Ľ —É—Ā—ā–į–Ĺ–ĺ–≤–ł—ā—Ć –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ—Ź—Ä–Ĺ—č–Ķ –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ—č, –Ņ–ĺ—Ā—Ä–Ķ–ī—Ā—ā–≤–ĺ–ľ –ļ–ĺ—ā–ĺ—Ä—č—Ö –ĺ—Ā—É—Č–Ķ—Ā—ā–≤–Ľ—Ź–Ķ—ā—Ā—Ź –∑–į—Č–ł—ā–Ĺ–ĺ–Ķ –ī–Ķ–Ļ—Ā—ā–≤–ł–Ķ –≤–ł—ā–į–ľ–ł–Ĺ–į D, —ā–ĺ—Ä–ľ–ĺ–∑—Ź—Č–Ķ–Ķ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā—č –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –ł¬†–ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ. –í¬†—Ü–Ķ–Ľ–ĺ–ľ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –į–Ĺ–į–Ľ–ł–∑–į —É–ļ–į–∑—č–≤–į—é—ā –Ĺ–į¬†—ā–ĺ, —á—ā–ĺ –Ĺ–į¬†—Ä–ł—Ā–ļ —Ä–į–∑–≤–ł—ā–ł—Ź –ĺ—Ā—ā–Ķ–ĺ–Ņ–ĺ—Ä–ĺ–∑–į –ł¬†–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ–∑–į –≤–Ľ–ł—Ź–Ķ—ā –Ĺ–Ķ¬†—É—Ä–ĺ–≤–Ķ–Ĺ—Ć –ļ–į–Ľ—Ć—Ü–ł—Ź, –į¬†–Ķ–≥–ĺ –ī–ł—Ā–Ī–į–Ľ–į–Ĺ—Ā –≤¬†–ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–Ķ, –ĺ—Ā–ĺ–Ī–Ķ–Ĺ–Ĺ–ĺ –Ņ—Ä–ł –ī–Ķ—Ą–ł—Ü–ł—ā–Ķ –≤–ł—ā–į–ľ–ł–Ĺ–į D3. –°–Ľ–Ķ–ī—É–Ķ—ā –Ķ—Č–Ķ —Ä–į–∑ –Ņ–ĺ–ī—á–Ķ—Ä–ļ–Ĺ—É—ā—Ć, —á—ā–ĺ –į–Ĺ—ā–ł–į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł–Ļ –ł¬†–ĺ—Ā—ā–Ķ–ĺ–Ņ—Ä–ĺ—ā–Ķ–ļ—ā–ĺ—Ä–Ĺ—č–Ļ —ć—Ą—Ą–Ķ–ļ—ā –ĺ–ļ–į–∑—č–≤–į–Ķ—ā —Ā–ĺ—á–Ķ—ā–į–Ĺ–Ĺ—č–Ļ –Ņ—Ä–ł–Ķ–ľ –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D. –ü—Ä–ł–Ķ–ľ –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ĺ–≤ –ļ–į–Ľ—Ć—Ü–ł—Ź –Ī–Ķ–∑ –≤–ł—ā–į–ľ–ł–Ĺ–į D –ł¬†–ľ–į–≥–Ĺ–ł—Ź –Ĺ–Ķ¬†–Ī—É–ī–Ķ—ā —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤–ĺ–≤–į—ā—Ć —Ą–ł–∑–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ĺ–ľ—É —Ä–į—Ā–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–ł—é –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†–ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–Ķ –ł¬†–ľ–ĺ–∂–Ķ—ā –Ņ—Ä–ł–≤–ĺ–ī–ł—ā—Ć –ļ¬†—É—Ā–ł–Ľ–Ķ–Ĺ–ł—é –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—ā–Ķ—Ä–ĺ—Ā–ļ–Ľ–Ķ—Ä–ĺ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ī–Ľ—Ź—ą–Ķ–ļ. –ě—á–Ķ–≤–ł–ī–Ĺ–ĺ –ł¬†–ī—Ä—É–≥–ĺ–Ķ: –Ņ—Ä–ł–Ķ–ľ –≤–ł—ā–į–ľ–ł–Ĺ–į D –Ĺ–į¬†—Ą–ĺ–Ĺ–Ķ –ī–Ķ—Ą–ł—Ü–ł—ā–į –ļ–į–Ľ—Ć—Ü–ł—Ź —ā–į–ļ–∂–Ķ –Ĺ–Ķ¬†–Ī—É–ī–Ķ—ā —ć—Ą—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ –≤–ĺ—Ā—Ā—ā–į–Ĺ–į–≤–Ľ–ł–≤–į—ā—Ć –ļ–į–Ľ—Ć—Ü–ł–Ķ–≤—č–Ļ¬†–≥–ĺ–ľ–Ķ–ĺ—Ā—ā–į–∑. –°–ĺ—á–Ķ—ā–į–Ĺ–Ĺ—č–Ļ –Ņ—Ä–ł–Ķ–ľ –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D (–Ě–į—ā–Ķ–ļ–į–Ľ—Ć –Ē3¬†–ł¬†–ī—Ä.) –Ī—É–ī–Ķ—ā —Ā–Ņ–ĺ—Ā–ĺ–Ī—Ā—ā–≤–ĺ–≤–į—ā—Ć –ļ–į–ļ –Ĺ–į–ļ–ĺ–Ņ–Ľ–Ķ–Ĺ–ł—é –ļ–į–Ľ—Ć—Ü–ł—Ź –≤¬†–ī–Ķ–Ņ–ĺ –ļ–ĺ—Ā—ā–Ķ–Ļ, —ā–į–ļ –ł¬†—É–ľ–Ķ–Ĺ—Ć—ą–Ķ–Ĺ–ł—é —Ä–ł—Ā–ļ–į –ļ–į–Ľ—Ć—Ü–ł—Ą–ł–ļ–į—Ü–ł–ł –į—Ä—ā–Ķ—Ä–ł–Ļ. –í¬†–Ņ–ĺ—Ā–Ľ–Ķ–ī—É—é—Č–Ķ–ľ –Ņ–Ľ–į–Ĺ–ł—Ä—É–Ķ—ā—Ā—Ź –ī–Ķ—ā–į–Ľ—Ć–Ĺ–ĺ —Ä–į–∑—Ä–į–Ī–ĺ—ā–į—ā—Ć –ī–ł–į–≥–Ĺ–ĺ—Ā—ā–ł—á–Ķ—Ā–ļ–ł–Ķ –ļ—Ä–ł—ā–Ķ—Ä–ł–ł –ĺ–Ī–Ķ—Ā–Ņ–Ķ—á–Ķ–Ĺ–Ĺ–ĺ—Ā—ā–ł –ļ–į–Ľ—Ć—Ü–ł–Ķ–ľ –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–ĺ–ľ D3 –ī–Ľ—Ź –Ī–ĺ–Ľ–Ķ–Ķ –Ņ–Ķ—Ä—Ā–ĺ–Ĺ–į–Ľ–ł–∑–ł—Ä–ĺ–≤–į–Ĺ–Ĺ–ĺ–≥–ĺ –Ņ–ĺ–ī—Ö–ĺ–ī–į –ļ¬†–≤—č–Ī–ĺ—Ä—É –Ņ—Ä–ĺ—Ą–ł–Ľ–į–ļ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –ł¬†–Ľ–Ķ—á–Ķ–Ī–Ĺ—č—Ö –ī–ĺ–∑ –ļ–į–Ľ—Ć—Ü–ł—Ź –ł¬†–≤–ł—ā–į–ľ–ł–Ĺ–į D3.¬†
–£–≤–į–∂–į–Ķ–ľ—č–Ļ –Ņ–ĺ—Ā–Ķ—ā–ł—ā–Ķ–Ľ—Ć uMEDp!
–£–≤–Ķ–ī–ĺ–ľ–Ľ—Ź–Ķ–ľ –í–į—Ā –ĺ —ā–ĺ–ľ, —á—ā–ĺ –∑–ī–Ķ—Ā—Ć —Ā–ĺ–ī–Ķ—Ä–∂–ł—ā—Ā—Ź –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł—Ź, –Ņ—Ä–Ķ–ī–Ĺ–į–∑–Ĺ–į—á–Ķ–Ĺ–Ĺ–į—Ź –ł—Ā–ļ–Ľ—é—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –ī–Ľ—Ź —Ā–Ņ–Ķ—Ü–ł–į–Ľ–ł—Ā—ā–ĺ–≤ –∑–ī—Ä–į–≤–ĺ–ĺ—Ö—Ä–į–Ĺ–Ķ–Ĺ–ł—Ź.
–ē—Ā–Ľ–ł –í—č –Ĺ–Ķ —Ź–≤–Ľ—Ź–Ķ—ā–Ķ—Ā—Ć —Ā–Ņ–Ķ—Ü–ł–į–Ľ–ł—Ā—ā–ĺ–ľ –∑–ī—Ä–į–≤–ĺ–ĺ—Ö—Ä–į–Ĺ–Ķ–Ĺ–ł—Ź, –į–ī–ľ–ł–Ĺ–ł—Ā—ā—Ä–į—Ü–ł—Ź –Ĺ–Ķ –Ĺ–Ķ—Ā–Ķ—ā –ĺ—ā–≤–Ķ—ā—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ—Ā—ā–ł –∑–į –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č–Ķ –ĺ—ā—Ä–ł—Ü–į—ā–Ķ–Ľ—Ć–Ĺ—č–Ķ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź, –≤–ĺ–∑–Ĺ–ł–ļ—ą–ł–Ķ –≤ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ —Ā–į–ľ–ĺ—Ā—ā–ĺ—Ź—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–ł—Ź –í–į–ľ–ł –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł —Ā –Ņ–ĺ—Ä—ā–į–Ľ–į –Ī–Ķ–∑ –Ņ—Ä–Ķ–ī–≤–į—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –ļ–ĺ–Ĺ—Ā—É–Ľ—Ć—ā–į—Ü–ł–ł —Ā –≤—Ä–į—á–ĺ–ľ.
–Ě–į–∂–ł–ľ–į—Ź –Ĺ–į –ļ–Ĺ–ĺ–Ņ–ļ—É ¬ę–í–ĺ–Ļ—ā–ł¬Ľ, –í—č –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–į–Ķ—ā–Ķ, —á—ā–ĺ —Ź–≤–Ľ—Ź–Ķ—ā–Ķ—Ā—Ć –≤—Ä–į—á–ĺ–ľ –ł–Ľ–ł —Ā—ā—É–ī–Ķ–Ĺ—ā–ĺ–ľ –ľ–Ķ–ī–ł—Ü–ł–Ĺ—Ā–ļ–ĺ–≥–ĺ –≤—É–∑–į.