–ú–ĺ–ī–Ķ–Ľ–ł—Ä–ĺ–≤–į–Ĺ–ł–Ķ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–į —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –Ņ–ĺ –Ī—É–ī–Ĺ—Ź–ľ –ł –≤—č—Ö–ĺ–ī–Ĺ—č–ľ: –ĺ–Ņ—Ä–ĺ–≤–Ķ—Ä–∂–Ķ–Ĺ–ł–Ķ –ľ–ł—Ą–ĺ–≤, —Ā–≤—Ź–∑–į–Ĺ–Ĺ—č—Ö —Ā¬†—Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–ľ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–ĺ–ľ
- –ź–Ĺ–Ĺ–ĺ—ā–į—Ü–ł—Ź
- –°—ā–į—ā—Ć—Ź
- –°—Ā—č–Ľ–ļ–ł
- English
–ú–į—ā–Ķ—Ä–ł–į–Ľ –ł –ľ–Ķ—ā–ĺ–ī—č. –Ē–Ľ—Ź —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤ –Ī—É–ī–Ĺ–ł –ł –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –Ī—č–Ľ–ł –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ—č 190 –≤—č–Ī–ĺ—Ä–ĺ–ļ –ł–∑ –Ľ–ł—ā–Ķ—Ä–į—ā—É—Ä—č (—Ā—Ä–Ķ–ī–Ĺ–ł–Ļ –≤–ĺ–∑—Ä–į—Ā—ā –ł—Ā–Ņ—č—ā—É–Ķ–ľ—č—Ö –≤–į—Ä—Ć–ł—Ä–ĺ–≤–į–Ľ—Ā—Ź –ĺ—ā –Ņ–ĺ–Ľ—É–≥–ĺ–ī–į –ī–ĺ 60 –Ľ–Ķ—ā). –ü—Ä–ł–ľ–Ķ–Ĺ—Ź–Ľ–į—Ā—Ć –ļ–Ľ–į—Ā—Ā–ł—á–Ķ—Ā–ļ–į—Ź –ī–≤—É—Ö–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–Ĺ–į—Ź –ľ–ĺ–ī–Ķ–Ľ—Ć —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł —Ā–Ĺ–į –ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź, –Ņ—Ä–Ķ–ī–Ľ–ĺ–∂–Ķ–Ĺ–Ĺ–į—Ź S. Daan –ł —Ā–ĺ–į–≤—ā. –≤ 1984 –≥., –≤ –ľ–ĺ–ī–ł—Ą–ł–ļ–į—Ü–ł–ł –į–≤—ā–ĺ—Ä–į 1995 –≥. –ö—Ä–ĺ–ľ–Ķ —ā–ĺ–≥–ĺ, –ī–Ľ—Ź –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–Ķ–Ĺ–ł—Ź –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö –ł–∑ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–ĺ–≤ –Ņ–Ķ—Ä–≤—č—Ö 190 –≤—č–Ī–ĺ—Ä–ĺ–ļ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ľ–ł—Ā—Ć –ī–į–Ĺ–Ĺ—č–Ķ 117 –ī–ĺ–Ņ–ĺ–Ľ–Ĺ–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –≤—č–Ī–ĺ—Ä–ĺ–ļ –ł–∑ –Ņ—É–Ī–Ľ–ł–ļ–į—Ü–ł–Ļ –Ņ–ĺ—Ā–Ľ–Ķ–ī–Ĺ–ł—Ö –ī–≤—É—Ö –Ľ–Ķ—ā.
–†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č. –ź–Ĺ–į–Ľ–ł–∑ —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł—Ö –ī–į–Ĺ–Ĺ—č—Ö –ł –ł—Ö —Ā–ł–ľ—É–Ľ—Ź—Ü–ł—Ź –Ĺ–Ķ –Ņ–ĺ–ī–ī–Ķ—Ä–∂–į–Ľ–ł –Ĺ–ł –ĺ–ī–Ĺ–ĺ –ł–∑ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł–Ļ, –Ī—č—ā—É—é—Č–ł—Ö –≤–ĺ–ļ—Ä—É–≥ —Ź–≤–Ľ–Ķ–Ĺ–ł—Ź —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į, –≤ —á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, —á—ā–ĺ —Ä–į–Ĺ–Ĺ–ł–Ķ –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł—Ź –Ņ–ĺ –Ī—É–ī–Ĺ—Ź–ľ –Ņ—Ä–ł–≤–ĺ–ī—Ź—ā –ļ –į–ļ–ļ—É–ľ—É–Ľ—Ź—Ü–ł–ł –ī–ĺ–Ľ–≥–į —Ā–Ĺ–į –ł —Ā–ĺ–Ĺ, –Ņ–ĺ—ā–Ķ—Ä—Ź–Ĺ–Ĺ—č–Ļ –ł–∑-–∑–į —Ä–į–Ĺ–Ĺ–ł—Ö –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł–Ļ –≤ —ć—ā–ł –ī–Ĺ–ł, –ľ–ĺ–∂–Ĺ–ĺ —Ö–ĺ—ā—Ź –Ī—č —á–į—Ā—ā–ł—á–Ĺ–ĺ –ļ–ĺ–ľ–Ņ–Ķ–Ĺ—Ā–ł—Ä–ĺ–≤–į—ā—Ć, –ĺ—ā–ĺ—Ā–Ņ–į–≤—ą–ł—Ā—Ć –≤ –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ. –ü–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–Ķ–Ĺ–į –ĺ—ą–ł–Ī–ĺ—á–Ĺ–ĺ—Ā—ā—Ć –ľ–Ĺ–Ķ–Ĺ–ł—Ź, —á—ā–ĺ –≤—Ä–Ķ–ľ—Ź –Ĺ–į—á–į–Ľ–į –ł –ĺ–ļ–ĺ–Ĺ—á–į–Ĺ–ł—Ź —Ā–Ĺ–į –≤ –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł –ľ–ĺ–∂–Ĺ–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į—ā—Ć –ī–Ľ—Ź –ł–ī–Ķ–Ĺ—ā–ł—Ą–ł–ļ–į—Ü–ł–ł ¬ę—Ā–ĺ–≤¬Ľ –ł ¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ĺ–≤¬Ľ –ł —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–Ļ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑ –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –≤—č—Ä–į–∂–Ķ–Ĺ —É ¬ę—Ā–ĺ–≤¬Ľ. –ü—Ä–ł–∑–Ĺ–į–Ĺ–ĺ –Ĺ–Ķ–≤–Ķ—Ä–Ĺ—č–ľ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–Ķ, —á—ā–ĺ –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č –Ĺ–į—á–ł–Ĺ–į—é—ā –∑–į–Ņ–į–∑–ī—č–≤–į—ā—Ć –≤ –Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ (—á—ā–ĺ —Ā–Ľ–Ķ–ī—É–Ķ—ā –ł–∑ –∑–į–Ņ–į–∑–ī—č–≤–į–Ĺ–ł—Ź –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤ –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ), –Ĺ–ĺ –∑–į—ā–Ķ–ľ –Ĺ–į—á–ł–Ĺ–į—é—ā —Ā–Ĺ–ĺ–≤–į —Ā–Ņ–Ķ—ą–ł—ā—Ć –Ņ–ĺ –ľ–Ķ—Ä–Ķ –ī–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–≥–ĺ –≤–∑—Ä–ĺ—Ā–Ľ–Ķ–Ĺ–ł—Ź –ł —Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź. –Ě–į–ļ–ĺ–Ĺ–Ķ—Ü, –ĺ–Ņ—Ä–ĺ–≤–Ķ—Ä–≥–Ĺ—É—ā–ĺ –ľ–Ĺ–Ķ–Ĺ–ł–Ķ, —á—ā–ĺ –≤ —ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ī–Ķ–Ľ–ł –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č –ĺ—Ā—ā–į—é—ā—Ā—Ź —Ā–ł–Ĺ—Ą–į–∑–Ĺ—č–ľ–ł 24-—á–į—Ā–ĺ–≤–ĺ–ľ—É —Ä–Ķ–∂–ł–ľ—É –Ķ—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –ĺ—Ā–≤–Ķ—Č–Ķ–Ĺ–ł—Ź –ł —ā–ĺ–Ľ—Ć–ļ–ĺ —Ą–į–∑–į —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ĺ ‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ —Ā–ī–≤–ł–≥–į–Ķ—ā—Ā—Ź –≤–Ņ–Ķ—Ä–Ķ–ī-–Ĺ–į–∑–į–ī.
–í—č–≤–ĺ–ī—č. –ě—á–Ķ–≤–ł–ī–Ĺ–ĺ, —á—ā–ĺ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł—Ź –ĺ —Ö—Ä–ĺ–Ĺ–ĺ–Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–į—Ö, –Ľ–Ķ–∂–į—Č–ł—Ö –≤ –ĺ—Ā–Ĺ–ĺ–≤–Ķ —Ź–≤–Ľ–Ķ–Ĺ–ł—Ź —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į, –Ĺ—É–∂–ī–į—é—ā—Ā—Ź –≤ –ĺ—Ā–Ĺ–ĺ–≤–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ–ľ –Ņ–Ķ—Ä–Ķ—Ā–ľ–ĺ—ā—Ä–Ķ.
–ú–į—ā–Ķ—Ä–ł–į–Ľ –ł –ľ–Ķ—ā–ĺ–ī—č. –Ē–Ľ—Ź —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤ –Ī—É–ī–Ĺ–ł –ł –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –Ī—č–Ľ–ł –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ—č 190 –≤—č–Ī–ĺ—Ä–ĺ–ļ –ł–∑ –Ľ–ł—ā–Ķ—Ä–į—ā—É—Ä—č (—Ā—Ä–Ķ–ī–Ĺ–ł–Ļ –≤–ĺ–∑—Ä–į—Ā—ā –ł—Ā–Ņ—č—ā—É–Ķ–ľ—č—Ö –≤–į—Ä—Ć–ł—Ä–ĺ–≤–į–Ľ—Ā—Ź –ĺ—ā –Ņ–ĺ–Ľ—É–≥–ĺ–ī–į –ī–ĺ 60 –Ľ–Ķ—ā). –ü—Ä–ł–ľ–Ķ–Ĺ—Ź–Ľ–į—Ā—Ć –ļ–Ľ–į—Ā—Ā–ł—á–Ķ—Ā–ļ–į—Ź –ī–≤—É—Ö–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–Ĺ–į—Ź –ľ–ĺ–ī–Ķ–Ľ—Ć —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł —Ā–Ĺ–į –ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź, –Ņ—Ä–Ķ–ī–Ľ–ĺ–∂–Ķ–Ĺ–Ĺ–į—Ź S. Daan –ł —Ā–ĺ–į–≤—ā. –≤ 1984 –≥., –≤ –ľ–ĺ–ī–ł—Ą–ł–ļ–į—Ü–ł–ł –į–≤—ā–ĺ—Ä–į 1995 –≥. –ö—Ä–ĺ–ľ–Ķ —ā–ĺ–≥–ĺ, –ī–Ľ—Ź –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–Ķ–Ĺ–ł—Ź –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä—č—Ö –ł–∑ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–ĺ–≤ –Ņ–Ķ—Ä–≤—č—Ö 190 –≤—č–Ī–ĺ—Ä–ĺ–ļ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ľ–ł—Ā—Ć –ī–į–Ĺ–Ĺ—č–Ķ 117 –ī–ĺ–Ņ–ĺ–Ľ–Ĺ–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –≤—č–Ī–ĺ—Ä–ĺ–ļ –ł–∑ –Ņ—É–Ī–Ľ–ł–ļ–į—Ü–ł–Ļ –Ņ–ĺ—Ā–Ľ–Ķ–ī–Ĺ–ł—Ö –ī–≤—É—Ö –Ľ–Ķ—ā.
–†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č. –ź–Ĺ–į–Ľ–ł–∑ —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł—Ö –ī–į–Ĺ–Ĺ—č—Ö –ł –ł—Ö —Ā–ł–ľ—É–Ľ—Ź—Ü–ł—Ź –Ĺ–Ķ –Ņ–ĺ–ī–ī–Ķ—Ä–∂–į–Ľ–ł –Ĺ–ł –ĺ–ī–Ĺ–ĺ –ł–∑ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł–Ļ, –Ī—č—ā—É—é—Č–ł—Ö –≤–ĺ–ļ—Ä—É–≥ —Ź–≤–Ľ–Ķ–Ĺ–ł—Ź —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į, –≤ —á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, —á—ā–ĺ —Ä–į–Ĺ–Ĺ–ł–Ķ –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł—Ź –Ņ–ĺ –Ī—É–ī–Ĺ—Ź–ľ –Ņ—Ä–ł–≤–ĺ–ī—Ź—ā –ļ –į–ļ–ļ—É–ľ—É–Ľ—Ź—Ü–ł–ł –ī–ĺ–Ľ–≥–į —Ā–Ĺ–į –ł —Ā–ĺ–Ĺ, –Ņ–ĺ—ā–Ķ—Ä—Ź–Ĺ–Ĺ—č–Ļ –ł–∑-–∑–į —Ä–į–Ĺ–Ĺ–ł—Ö –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł–Ļ –≤ —ć—ā–ł –ī–Ĺ–ł, –ľ–ĺ–∂–Ĺ–ĺ —Ö–ĺ—ā—Ź –Ī—č —á–į—Ā—ā–ł—á–Ĺ–ĺ –ļ–ĺ–ľ–Ņ–Ķ–Ĺ—Ā–ł—Ä–ĺ–≤–į—ā—Ć, –ĺ—ā–ĺ—Ā–Ņ–į–≤—ą–ł—Ā—Ć –≤ –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ. –ü–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–Ķ–Ĺ–į –ĺ—ą–ł–Ī–ĺ—á–Ĺ–ĺ—Ā—ā—Ć –ľ–Ĺ–Ķ–Ĺ–ł—Ź, —á—ā–ĺ –≤—Ä–Ķ–ľ—Ź –Ĺ–į—á–į–Ľ–į –ł –ĺ–ļ–ĺ–Ĺ—á–į–Ĺ–ł—Ź —Ā–Ĺ–į –≤ –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł –ľ–ĺ–∂–Ĺ–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į—ā—Ć –ī–Ľ—Ź –ł–ī–Ķ–Ĺ—ā–ł—Ą–ł–ļ–į—Ü–ł–ł ¬ę—Ā–ĺ–≤¬Ľ –ł ¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ĺ–≤¬Ľ –ł —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–Ļ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑ –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –≤—č—Ä–į–∂–Ķ–Ĺ —É ¬ę—Ā–ĺ–≤¬Ľ. –ü—Ä–ł–∑–Ĺ–į–Ĺ–ĺ –Ĺ–Ķ–≤–Ķ—Ä–Ĺ—č–ľ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–Ķ, —á—ā–ĺ –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č –Ĺ–į—á–ł–Ĺ–į—é—ā –∑–į–Ņ–į–∑–ī—č–≤–į—ā—Ć –≤ –Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ (—á—ā–ĺ —Ā–Ľ–Ķ–ī—É–Ķ—ā –ł–∑ –∑–į–Ņ–į–∑–ī—č–≤–į–Ĺ–ł—Ź –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤ –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ), –Ĺ–ĺ –∑–į—ā–Ķ–ľ –Ĺ–į—á–ł–Ĺ–į—é—ā —Ā–Ĺ–ĺ–≤–į —Ā–Ņ–Ķ—ą–ł—ā—Ć –Ņ–ĺ –ľ–Ķ—Ä–Ķ –ī–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–≥–ĺ –≤–∑—Ä–ĺ—Ā–Ľ–Ķ–Ĺ–ł—Ź –ł —Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź. –Ě–į–ļ–ĺ–Ĺ–Ķ—Ü, –ĺ–Ņ—Ä–ĺ–≤–Ķ—Ä–≥–Ĺ—É—ā–ĺ –ľ–Ĺ–Ķ–Ĺ–ł–Ķ, —á—ā–ĺ –≤ —ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ī–Ķ–Ľ–ł –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č –ĺ—Ā—ā–į—é—ā—Ā—Ź —Ā–ł–Ĺ—Ą–į–∑–Ĺ—č–ľ–ł 24-—á–į—Ā–ĺ–≤–ĺ–ľ—É —Ä–Ķ–∂–ł–ľ—É –Ķ—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –ĺ—Ā–≤–Ķ—Č–Ķ–Ĺ–ł—Ź –ł —ā–ĺ–Ľ—Ć–ļ–ĺ —Ą–į–∑–į —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ĺ ‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ —Ā–ī–≤–ł–≥–į–Ķ—ā—Ā—Ź –≤–Ņ–Ķ—Ä–Ķ–ī-–Ĺ–į–∑–į–ī.
–í—č–≤–ĺ–ī—č. –ě—á–Ķ–≤–ł–ī–Ĺ–ĺ, —á—ā–ĺ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł—Ź –ĺ —Ö—Ä–ĺ–Ĺ–ĺ–Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–į—Ö, –Ľ–Ķ–∂–į—Č–ł—Ö –≤ –ĺ—Ā–Ĺ–ĺ–≤–Ķ —Ź–≤–Ľ–Ķ–Ĺ–ł—Ź —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į, –Ĺ—É–∂–ī–į—é—ā—Ā—Ź –≤ –ĺ—Ā–Ĺ–ĺ–≤–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ–ľ –Ņ–Ķ—Ä–Ķ—Ā–ľ–ĺ—ā—Ä–Ķ.
–í–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ
–ö–ĺ–≥–ī–į –Ĺ–į¬†–Ņ—Ä–ĺ—ā—Ź–∂–Ķ–Ĺ–ł–ł —Ä–į–Ī–ĺ—á–Ķ–Ļ –ł–Ľ–ł —É—á–Ķ–Ī–Ĺ–ĺ–Ļ –Ĺ–Ķ–ī–Ķ–Ľ–ł –Ņ—Ä–ł—Ö–ĺ–ī–ł—ā—Ā—Ź –≤—Ā—ā–į–≤–į—ā—Ć –Ņ–ĺ—Ä–į–Ĺ—Ć—ą–Ķ, —ā–ĺ –Ĺ–ĺ—á–Ĺ–ĺ–Ļ —Ā–ĺ–Ĺ —Ā—ā–į–Ĺ–ĺ–≤–ł—ā—Ā—Ź –ļ–ĺ—Ä–ĺ—á–Ķ –∂–Ķ–Ľ–į–Ķ–ľ–ĺ–≥–ĺ. –í¬†–ļ–ĺ–Ĺ—Ü–Ķ —ā–į–ļ–ĺ–Ļ –Ĺ–Ķ–ī–Ķ–Ľ–ł —Ö–ĺ—á–Ķ—ā—Ā—Ź –ļ–ĺ–ľ–Ņ–Ķ–Ĺ—Ā–ł—Ä–ĺ–≤–į—ā—Ć (—Ö–ĺ—ā—Ź –Ī—č —á–į—Ā—ā–ł—á–Ĺ–ĺ) –Ņ–ĺ—ā–Ķ—Ä—Ź–Ĺ–Ĺ—č–Ļ —Ā–ĺ–Ĺ, –Ņ–ĺ—Ā–Ņ–į—ā—Ć –Ņ–ĺ–ī–ĺ–Ľ—Ć—ą–Ķ –≤ –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ł¬†–Ī–Ľ–į–≥–ĺ–ī–į—Ä—Ź –Ĺ–Ķ–ļ–ĺ—ā–ĺ—Ä–ĺ–ľ—É ¬ę–Ņ–Ķ—Ä–Ķ—Ā—č–Ņ—ɬĽ –≤–Ķ—Ä–Ĺ—É—ā—Ć—Ā—Ź –≤¬†–Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ–Ķ —Ā–ĺ—Ā—ā–ĺ—Ź–Ĺ–ł–Ķ. –Ē–Ķ–Ļ—Ā—ā–≤–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –Ķ—Ā–Ľ–ł —É–ī–į–Ķ—ā—Ā—Ź –Ņ–ĺ—Ā–Ņ–į—ā—Ć –Ņ–ĺ–ī–ĺ–Ľ—Ć—ą–Ķ –≤¬†–Ĺ–ĺ—á—Ć —Ā¬†–Ņ—Ź—ā–Ĺ–ł—Ü—č –Ĺ–į¬†—Ā—É–Ī–Ī–ĺ—ā—É –ł/–ł–Ľ–ł —Ā¬†—Ā—É–Ī–Ī–ĺ—ā—č –Ĺ–į¬†–≤–ĺ—Ā–ļ—Ä–Ķ—Ā–Ķ–Ĺ—Ć–Ķ, —á–Ķ–Ľ–ĺ–≤–Ķ–ļ —á—É–≤—Ā—ā–≤—É–Ķ—ā —Ā–Ķ–Ī—Ź –Ĺ–į–ľ–Ĺ–ĺ–≥–ĺ –Ľ—É—á—ą–Ķ, —á–Ķ–ľ –≤¬†–Ī—É–ī–Ĺ–ł. –í¬†—á–Ķ–ľ –ĺ—ą–ł–Ī–ĺ—á–Ĺ–ĺ—Ā—ā—Ć –Ņ–ĺ–ī–ĺ–Ī–Ĺ–ĺ–≥–ĺ —Ä–į—Ā—Ā—É–∂–ī–Ķ–Ĺ–ł—Ź? –í¬†–ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–ł —ā–ĺ–≥–ĺ, —á—ā–ĺ —É—ā–Ķ—Ä—Ź–Ĺ–Ĺ—č–Ļ —Ā–ĺ–Ĺ –ľ–ĺ–∂–Ĺ–ĺ –≤–Ķ—Ä–Ĺ—É—ā—Ć —Ö–ĺ—ā—Ź –Ī—č —á–į—Ā—ā–ł—á–Ĺ–ĺ.
–ü–ĺ—Ä–į–∑–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ, —á—ā–ĺ —ć—ā–į –ľ—č—Ā–Ľ—Ć, –≤–Ņ–ĺ–Ľ–Ĺ–Ķ –Ķ—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –≤–ĺ–∑–Ĺ–ł–ļ–į—é—Č–į—Ź –Ĺ–į¬†–Ī—č—ā–ĺ–≤–ĺ–ľ —É—Ä–ĺ–≤–Ĺ–Ķ, –Ĺ–į—ą–Ľ–į –≤–ĺ–Ņ–Ľ–ĺ—Č–Ķ–Ĺ–ł–Ķ –≤¬†–Ĺ–į—É—á–Ĺ–ĺ–Ļ –Ľ–ł—ā–Ķ—Ä–į—ā—É—Ä–Ķ –Ņ–ĺ–ī –Ĺ–į–∑–≤–į–Ĺ–ł–Ķ–ľ —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į (social jet lag). –ü—Ä–į–≤–ī–į, —ć—ā–ĺ –Ņ—Ä–ĺ–ł–∑–ĺ—ą–Ľ–ĺ —Ā—Ä–į–≤–Ĺ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –Ĺ–Ķ–ī–į–≤–Ĺ–ĺ (–≤¬†2006¬†–≥.) –ł, —á—ā–ĺ —É–∂–Ķ —Ā–į–ľ–ĺ –Ņ–嬆—Ā–Ķ–Ī–Ķ –Ņ–ĺ–ī–ĺ–∑—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –Ĺ–į–ľ–Ĺ–ĺ–≥–ĺ –Ņ–ĺ–∑–ī–Ĺ–Ķ–Ķ –ĺ—Ą–ĺ—Ä–ľ–ł–≤—ą–Ķ–Ļ—Ā—Ź –≤¬†–Ĺ–į—á–į–Ľ–Ķ 1960-—Ö¬†–≥–≥. —Ö—Ä–ĺ–Ĺ–ĺ–Ī–ł–ĺ–Ľ–ĺ–≥–ł–ł –≤¬†–ļ–į—á–Ķ—Ā—ā–≤–Ķ –ĺ—ā–ī–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –Ĺ–į—É—á–Ĺ–ĺ–Ļ –ĺ–Ī–Ľ–į—Ā—ā–ł, –ļ–ĺ—ā–ĺ—Ä–į—Ź –ł—Ā—Ā–Ľ–Ķ–ī—É–Ķ—ā –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —Ä–ł—ā–ľ—č, —ā–ĺ –Ķ—Ā—ā—Ć —É–∂–Ķ –Ņ–ĺ—Ā–Ľ–Ķ –Ĺ–į—É—á–Ĺ–ĺ–Ļ —Ą–ł–ļ—Ā–į—Ü–ł–ł –≤—Ā–Ķ—Ö –ĺ—Ā–Ĺ–ĺ–≤–Ĺ—č—Ö –ł–∑—É—á–į–Ķ–ľ—č—Ö –≤¬†–Ķ–Ķ —Ä–į–ľ–ļ–į—Ö —Ą–Ķ–Ĺ–ĺ–ľ–Ķ–Ĺ–ĺ–≤. –ö¬†–Ĺ–ł–ľ, –≤¬†—á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, –ĺ—ā–Ĺ–ĺ—Ā–ł—ā—Ā—Ź —Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ —Ā–ĺ–Ī—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į (jet lag), –Ņ–ĺ–ī –ļ–ĺ—ā–ĺ—Ä—č–ľ –Ņ–ĺ–Ĺ–ł–ľ–į–Ķ—ā—Ā—Ź –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ–ĺ–Ķ —Ä–į—Ā—Ā–ĺ–≥–Ľ–į—Ā–ĺ–≤–į–Ĺ–ł–Ķ —Ą–į–∑ —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ—č—Ö (–ĺ–ļ–ĺ–Ľ–ĺ—Ā—É—ā–ĺ—á–Ĺ—č—Ö) —Ä–ł—ā–ľ–ĺ–≤ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į –≤¬†–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–Ķ –ł—Ö –≤—č–Ĺ—É–∂–ī–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ —Ā–ī–≤–ł–≥–į –≤—Ā–Ľ–Ķ–ī –∑–į —Ā–ī–≤–ł–≥–ĺ–ľ —Ą–į–∑—č –≤–Ĺ–Ķ—ą–Ĺ–Ķ–≥–ĺ 24-—á–į—Ā–ĺ–≤–ĺ–≥–ĺ —Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ł–∑–ł—Ä—É—é—Č–Ķ–≥–ĺ –≤–ĺ–∑–ī–Ķ–Ļ—Ā—ā–≤–ł—Ź. –ö–ĺ–Ĺ—Ü–Ķ–Ņ—Ü–ł—é —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į –Ņ—Ä–Ķ–ī–Ľ–ĺ–∂–ł–Ľ–ł M.¬†Wittmann –ł¬†—Ā–ĺ–į–≤—ā., –ļ–ĺ—ā–ĺ—Ä—č–Ķ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–ł–Ľ–ł –Ķ–≥–ĺ –ļ–į–ļ —Ā–Ņ–Ķ—Ü–ł—Ą–ł—á–Ķ—Ā–ļ—É—é (–ī–ĺ–Ī–į–≤–Ľ—鬆‚Äď –≤–ł–ī–ł–ľ–ĺ, —Ä–į–Ĺ–Ķ–Ķ –Ĺ–ł–ļ–ĺ–ľ—É –Ĺ–Ķ¬†–ł–∑–≤–Ķ—Ā—ā–Ĺ—É—é) —Ą–ĺ—Ä–ľ—É –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į –≤¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ —Ä–į—Ā—Ā–ĺ–≥–Ľ–į—Ā–ĺ–≤–į–Ĺ–ł—Ź –ľ–Ķ–∂–ī—É –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ķ–ľ —Ä–į–Ī–ĺ—ā—č/—É—á–Ķ–Ī—č –ł¬†–Ņ—Ä–Ķ–ī–Ņ–ĺ—á–ł—ā–į–Ķ–ľ—č–ľ –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ķ–ľ —Ā–Ĺ–į¬†–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź. –ü–ĺ—Ā–Ľ–Ķ–ī–Ĺ–Ķ–Ķ –∑–į–≤–ł—Ā–ł—ā –ĺ—ā¬†–≤–Ĺ—É—ā—Ä–Ķ–Ĺ–Ĺ–ł—Ö –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö —á–į—Ā–ĺ–≤, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –≤¬†—Ā–≤–ĺ—é –ĺ—á–Ķ—Ä–Ķ–ī—Ć –ļ–ĺ–Ĺ—ā—Ä–ĺ–Ľ–ł—Ä—É—é—ā—Ā—Ź 24-—á–į—Ā–ĺ–≤—č–ľ —Ä–Ķ–∂–ł–ľ–ĺ–ľ –ĺ—Ā–≤–Ķ—Č–Ķ–Ĺ–ł—Ź, –į¬†–Ĺ–Ķ –ļ–į–ļ–ł–ľ–ł-–Ľ–ł–Ī–ĺ —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–ľ–ł —Ā–ł–≥–Ĺ–į–Ľ–į–ľ–ł –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł, –≤–ļ–Ľ—é—á–į—Ź —á–į—Ā—č —Ä–į–Ī–ĺ—ā—č/—É—á–Ķ–Ī—č [1].
–ė—Ā—Ö–ĺ–ī—Ź –ł–∑¬†—ā–į–ļ–ĺ–≥–ĺ –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–Ķ–Ĺ–ł—Ź, –į–≤—ā–ĺ—Ä—č –Ņ—Ä–ł—ą–Ľ–ł –ļ¬†–≤—č–≤–ĺ–ī—É, —á—ā–ĺ —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–Ļ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑ –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –≤—č—Ä–į–∂–Ķ–Ĺ —ɬ†–Ņ–ĺ–∑–ī–Ĺ–Ķ–≥–ĺ —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į (—ā–ł–Ņ–į —Ā—É—ā–ĺ—á–Ĺ–ĺ–≥–ĺ —Ä–ł—ā–ľ–į), —ā–ĺ –Ķ—Ā—ā—Ć —ɬ†¬ę—Ā–ĺ–≤¬Ľ [1]. –ě–Ĺ–ł –≤—č–Ĺ—É–∂–ī–Ķ–Ĺ—č –Ņ—Ä–ł—Ā–Ņ–ĺ—Ā–į–Ī–Ľ–ł–≤–į—ā—Ć –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ—č–Ķ –Ņ—Ä–ł–≤—č—á–ļ–ł –ļ¬†—Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–ľ —ā—Ä–Ķ–Ī–ĺ–≤–į–Ĺ–ł—Ź–ľ. –ė–Ĺ–į—á–Ķ¬†–≥–ĺ–≤–ĺ—Ä—Ź, ¬ę—Ā–ĺ–≤–į–ľ¬Ľ –Ņ—Ä–ł—Ö–ĺ–ī–ł—ā—Ā—Ź –≤—Ā—ā–į–≤–į—ā—Ć —Ä–į–Ĺ–ĺ –ł¬†–ĺ–Ĺ–ł –Ņ—Ä–ĺ—Ā—ā–ĺ –Ĺ–Ķ —Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ—č —Ā–ī–≤–ł–Ĺ—É—ā—Ć –Ĺ–į¬†–Ī–ĺ–Ľ–Ķ–Ķ —Ä–į–Ĺ–Ĺ–Ķ–Ķ –≤—Ä–Ķ–ľ—Ź –Ĺ–į—á–į–Ľ–ĺ —Ā–≤–ĺ–Ķ–≥–ĺ —Ā–Ĺ–į, –∑–į –ļ–ĺ—ā–ĺ—Ä–ĺ–Ķ –ĺ—ā–≤–Ķ—á–į—é—ā –≤–Ĺ—É—ā—Ä–Ķ–Ĺ–Ĺ–ł–Ķ —á–į—Ā—č, –≤¬†—Ā–≤–ĺ—é –ĺ—á–Ķ—Ä–Ķ–ī—Ć –ļ–ĺ–Ĺ—ā—Ä–ĺ–Ľ–ł—Ä—É–Ķ–ľ—č–Ķ 24-—á–į—Ā–ĺ–≤—č–ľ —Ä–Ķ–∂–ł–ľ–ĺ–ľ –ĺ—Ā–≤–Ķ—Č–Ķ–Ĺ–ł—Ź. –Ę–į–ļ–ł–ľ –ĺ–Ī—Ä–į–∑–ĺ–ľ, –ī—Ä—É–≥–į—Ź –ł–ī–Ķ—Ź –į–≤—ā–ĺ—Ä–ĺ–≤ —Ā–ĺ—Ā—ā–ĺ–ł—ā –≤¬†—ā–ĺ–ľ, —á—ā–ĺ ¬ę—Ā–ĺ–≤—謼 –≤—č–Ĺ—É–∂–ī–Ķ–Ĺ—č –ĺ—ā—Ā—č–Ņ–į—ā—Ć—Ā—Ź –Ĺ–į¬†–≤—č—Ö–ĺ–ī–Ĺ—č—Ö –ī–ĺ–Ľ—Ć—ą–Ķ, —á–Ķ–ľ ¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ł¬Ľ [1]. –°–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –ĺ–Ĺ–ł –≤¬†–Ī–ĺ–Ľ—Ć—ą–Ķ–Ļ —Ā—ā–Ķ–Ņ–Ķ–Ĺ–ł —Ā—ā—Ä–į–ī–į—é—ā –ĺ—ā¬†—Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į, —ā–ĺ –Ķ—Ā—ā—Ć –ĺ—ā¬†—Ą–į–∑–ĺ–≤–ĺ–≥–ĺ —Ä–į—Ā—Ā–ĺ–≥–Ľ–į—Ā–ĺ–≤–į–Ĺ–ł—Ź –ľ–Ķ–∂–ī—É –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ķ–ľ —Ā–Ĺ–į –ł¬†—Ą–į–∑–ĺ–Ļ —Ā–ĺ–Ī—Ā—ā–≤–Ķ–Ĺ–Ĺ—č—Ö –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö —á–į—Ā–ĺ–≤. –ė, –Ķ—Ā–Ľ–ł –≤–Ķ—Ä–ł—ā—Ć –į–≤—ā–ĺ—Ä–į–ľ —Ā—ā–į—ā—Ć–ł, ¬ę—Ā–ĺ–≤—謼 –ļ–ĺ–ľ–Ņ–Ķ–Ĺ—Ā–ł—Ä—É—é—ā –≤ –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –Ĺ–Ķ–ī–Ķ–Ľ—Ć–Ĺ—č–Ļ –ī–ĺ–Ľ–≥ —Ā–Ĺ–į, –ļ–ĺ—ā–ĺ—Ä—č–Ļ —ɬ†–Ĺ–ł—Ö –≤—č—ą–Ķ, —á–Ķ–ľ —ɬ†¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ĺ–≤¬Ľ [1].
T. Roenneberg –ł¬†—Ā–ĺ–į–≤—ā. —É—ā–≤–Ķ—Ä–∂–ī–į—é—ā, —á—ā–ĺ —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ¬†‚Äď —Ā–Ņ–Ķ—Ü–ł—Ą–ł—á–Ķ—Ā–ļ–ĺ–Ķ –ī–Ľ—Ź –ł–Ĺ–ī–ł–≤–ł–ī—É—É–ľ–į –ĺ—ā–Ĺ–ĺ—ą–Ķ–Ĺ–ł–Ķ –ľ–Ķ–∂–ī—É —Ą–į–∑–ĺ–Ļ –≤–Ĺ—É—ā—Ä–Ķ–Ĺ–Ĺ–ł—Ö —á–į—Ā–ĺ–≤, –ł–Ĺ–ī–ł–ļ–į—ā–ĺ—Ä–į–ľ–ł –ļ–ĺ—ā–ĺ—Ä–ĺ–Ļ —Ź–≤–Ľ—Ź—é—ā—Ā—Ź —Ą–į–∑—č —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ—č—Ö —Ä–ł—ā–ľ–ĺ–≤ —ā–Ķ–ľ–Ņ–Ķ—Ä–į—ā—É—Ä—č —ā–Ķ–Ľ–į –ł¬†–ľ–Ķ–Ľ–į—ā–ĺ–Ĺ–ł–Ĺ–į, –ł¬†–≤–Ĺ–Ķ—ą–Ĺ–ł–ľ —Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ł–∑–į—ā–ĺ—Ä–ĺ–ľ, –Ĺ–į–Ņ—Ä–ł–ľ–Ķ—Ä –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ķ–ľ –≤–ĺ—Ā—Ö–ĺ–ī–į –ł¬†–∑–į—Ö–ĺ–ī–į —Ā–ĺ–Ľ–Ĺ—Ü–į [2]. –ü–嬆–ł—Ö –ľ–Ĺ–Ķ–Ĺ–ł—é, —Ā¬†—É—á–Ķ—ā–ĺ–ľ —ć—ā–ĺ–≥–ĺ –ĺ—ā–Ĺ–ĺ—ą–Ķ–Ĺ–ł—Ź –ī–Ķ—ā–ł –ĺ–Ī—č—á–Ĺ–ĺ –ł–ľ–Ķ—é—ā —Ä–į–Ĺ–Ĺ–ł–Ļ —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ, –Ĺ–ĺ, –Ņ–ĺ—Ā—ā–Ķ–Ņ–Ķ–Ĺ–Ĺ–ĺ —Ā–ī–≤–ł–≥–į—Ź –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į –ł¬†–ī–ĺ—Ā—ā–ł–≥–į—Ź –ľ–į–ļ—Ā–ł–ľ—É–ľ–į –≤¬†–≤–ĺ–∑—Ä–į—Ā—ā–Ķ –ĺ–ļ–ĺ–Ľ–ĺ 20 –Ľ–Ķ—ā, –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī—Ź—ā –ļ¬†–Ņ–ĺ–∑–ī–Ĺ–Ķ–ľ—É —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ—É, –∑–į—ā–Ķ–ľ –Ņ–嬆–ľ–Ķ—Ä–Ķ –ī–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–≥–ĺ –≤–∑—Ä–ĺ—Ā–Ľ–Ķ–Ĺ–ł—Ź –Ĺ–Ķ—Ä–Ķ–ī–ļ–ĺ –≤–ĺ–∑–≤—Ä–į—Č–į—Ź—Ā—Ć –ļ¬†—Ä–į–Ĺ–Ĺ–Ķ–ľ—É —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ—É [3]. –ü—Ä–ł —ć—ā–ĺ–ľ –∑–į–ľ–Ķ—ā–ł–ľ, —Ä–Ķ—á—Ć –ł–ī–Ķ—ā —É–∂–Ķ –嬆—Ą–į–∑–Ķ —Ā–Ĺ–į, —ā–ĺ–≥–ī–į –ļ–į–ļ –ľ–į—Ä–ļ–Ķ—Ä—č —Ą–į–∑—č –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö —á–į—Ā–ĺ–≤ (—Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ—č–Ķ —Ä–ł—ā–ľ—č —ā–Ķ–ľ–Ņ–Ķ—Ä–į—ā—É—Ä—č —ā–Ķ–Ľ–į –ł¬†–ľ–Ķ–Ľ–į—ā–ĺ–Ĺ–ł–Ĺ–į) –≤¬†—ć—ā–ł—Ö —Ä–į–Ī–ĺ—ā–į—Ö, –ĺ—Ā–Ĺ–ĺ–≤–į–Ĺ–Ĺ—č—Ö –Ĺ–į¬†–ĺ–Ņ—Ä–ĺ—Ā–Ĺ–ł–ļ–Ķ, –Ĺ–Ķ¬†–ł–∑–ľ–Ķ—Ä—Ź–Ľ–ł—Ā—Ć [3].
–•—Ä–ĺ–Ĺ–ĺ–Ī–ł–ĺ–Ľ–ĺ–≥–ł—Ź¬†‚Äď —Ą—É–Ĺ–ī–į–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ–į—Ź –ł, –ľ–ĺ–∂–Ĺ–ĺ –ī–į–∂–Ķ —Ā–ļ–į–∑–į—ā—Ć, —ā–ĺ—á–Ĺ–į—Ź –Ĺ–į—É–ļ–į, –Ņ–ĺ—Ā–ļ–ĺ–Ľ—Ć–ļ—É –≤¬†–ĺ—Ā–Ĺ–ĺ–≤–Ķ –≤—Ā–Ķ—Ö –Ķ–Ķ —ā–Ķ–ĺ—Ä–Ķ—ā–ł—á–Ķ—Ā–ļ–ł—Ö –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł–Ļ –Ľ–Ķ–∂–ł—ā –ľ–į—ā–Ķ–ľ–į—ā–ł—á–Ķ—Ā–ļ–į—Ź —ā–Ķ–ĺ—Ä–ł—Ź –ļ–ĺ–Ľ–Ķ–Ī–į–Ĺ–ł–Ļ. –°–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –ĺ–Ī—ä—Ź—Ā–Ĺ–ł—ā—Ć –ļ–į–ļ–ĺ–Ķ-–Ľ–ł–Ī–ĺ –ł–∑—É—á–į–Ķ–ľ–ĺ–Ķ —Ö—Ä–ĺ–Ĺ–ĺ–Ī–ł–ĺ–Ľ–ĺ–≥–į–ľ–ł —Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ –ľ–ĺ–∂–Ĺ–ĺ —ā–ĺ–Ľ—Ć–ļ–ĺ –Ņ—É—ā–Ķ–ľ –Ķ–≥–ĺ –ĺ–Ņ–ł—Ā–į–Ĺ–ł—Ź –ł¬†–ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –Ņ—Ä–Ķ–ī—Ā–ļ–į–∑–į–Ĺ–ł—Ź –≤¬†—ā–Ķ—Ä–ľ–ł–Ĺ–į—Ö —ć—ā–ĺ–Ļ —ā–Ķ–ĺ—Ä–ł–ł. –Ě–į–Ņ—Ä–ł–ľ–Ķ—Ä, –ľ–ĺ–∂–Ĺ–ĺ –Ņ—Ä–Ķ–ī—Ā–ļ–į–∑–į—ā—Ć –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ√≥–Ķ –Ņ–ĺ–≤–Ķ–ī–Ķ–Ĺ–ł–Ķ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į (–≤¬†—á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į –≤¬†–Ī—É–ī–Ĺ–ł –ł¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ) –Ņ—É—ā–Ķ–ľ –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–Ļ —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł —Ā¬†–Ņ–ĺ–ľ–ĺ—Č—Ć—é –ľ–į—ā–Ķ–ľ–į—ā–ł—á–Ķ—Ā–ļ–ĺ–Ļ –ľ–ĺ–ī–Ķ–Ľ–ł —Ä–Ķ–≥—É–Ľ—Ź—ā–ĺ—Ä–į –ļ–ĺ–Ĺ–ļ—Ä–Ķ—ā–Ĺ–ĺ–≥–ĺ —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–≥–ĺ —Ä–ł—ā–ľ–į (–≤¬†–ī–į–Ĺ–Ĺ–ĺ–ľ —Ā–Ľ—É—á–į–Ķ —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ). –£–ī–ł–≤–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –Ŗ嬆–Ĺ–ł–ļ—ā–ĺ –∑–į –Ī–ĺ–Ľ–Ķ–Ķ —á–Ķ–ľ –ī–Ķ—Ā—Ź—ā–ĺ–ļ –Ľ–Ķ—ā –Ĺ–Ķ¬†–Ņ–ĺ–Ņ—č—ā–į–Ľ—Ā—Ź –Ņ—Ä–ł–ľ–Ķ–Ĺ–ł—ā—Ć —ā–į–ļ–ĺ–Ļ —ā—Ä–į–ī–ł—Ü–ł–ĺ–Ĺ–Ĺ–ĺ-—Ö—Ä–ĺ–Ĺ–ĺ–Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ļ –Ņ–ĺ–ī—Ö–ĺ–ī –ļ¬†—ā–Ķ—Ā—ā–ł—Ä–ĺ–≤–į–Ĺ–ł—é —Ź–≤–Ľ–Ķ–Ĺ–ł—Ź —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į. –ü–ĺ—ć—ā–ĺ–ľ—É –≤¬†–ī–≤—É—Ö –Ņ—Ä–Ķ–ī—č–ī—É—Č–ł—Ö —Ā—ā–į—ā—Ć—Ź—Ö [4, 5] –ľ—č —Ā –ļ–ĺ–Ľ–Ľ–Ķ–≥–į–ľ–ł –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ľ–ł –Ņ—Ä–Ķ–ī–Ľ–ĺ–∂–Ķ–Ĺ–Ĺ—É—é –ľ–Ĺ–ĺ–Ļ —Ä–į–Ĺ–Ķ–Ķ –ľ–ĺ–ī–Ķ–Ľ—Ć —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ [6] –ī–Ľ—Ź –ļ–ĺ–Ľ–ł—á–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–Ļ —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł —Ä–į–∑–Ľ–ł—á–ł–Ļ –Ņ–嬆–≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –ľ–Ķ–∂–ī—É —Ä–į–∑–Ĺ—č–ľ–ł —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į–ľ–ł [5] –ł¬†–≤–ĺ–∑—Ä–į—Ā—ā–į–ľ–ł [4]. –í¬†–ļ–į—á–Ķ—Ā—ā–≤–Ķ —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł—Ö –ī–į–Ĺ–Ĺ—č—Ö –Ī—č–Ľ–ł –≤–∑—Ź—ā—č –ĺ–Ņ—É–Ī–Ľ–ł–ļ–ĺ–≤–į–Ĺ–Ĺ—č–Ķ –≤¬†–Ľ–ł—ā–Ķ—Ä–į—ā—É—Ä–Ķ —Ā–≤–Ķ–ī–Ķ–Ĺ–ł—Ź –嬆–≤—Ä–Ķ–ľ–Ķ–Ĺ–ł –ĺ—ā—Ö–ĺ–ī–į –ļ–嬆—Ā–Ĺ—É –ł¬†–Ņ–ĺ–ī—ä–Ķ–ľ–į –Ņ–嬆–Ī—É–ī–Ĺ—Ź–ľ –ł¬†–≤ –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł –ī–Ľ—Ź 190 –≤—č–Ī–ĺ—Ä–ĺ–ļ (–≤—Ā–Ķ–≥–ĺ —á–Ķ—ā—č—Ä–Ķ –Ņ–ĺ–ļ–į–∑–į—ā–Ķ–Ľ—Ź). –ē—Č–Ķ 117¬†–≤—č–Ī–ĺ—Ä–ĺ–ļ –ī–ĺ–Ņ–ĺ–Ľ–Ĺ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ —Ā–ĺ–Ī—Ä–į–Ĺ—č –ł–∑¬†–Ĺ–ĺ–≤—č—Ö —Ä–į–Ī–ĺ—ā 2018 –ł¬†2019¬†–≥–≥. –ł¬†–ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ—č –ī–Ľ—Ź –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–Ķ–Ĺ–ł—Ź —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–ĺ–≤, –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ–Ĺ—č—Ö —Ä–į–Ĺ–Ķ–Ķ [4,¬†5].
–¶–Ķ–Ľ—Ć –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź
–ü—Ä–ĺ–≤–Ķ—Ä–ł—ā—Ć –Ņ—Ä–į–≤–ī–ĺ–Ņ–ĺ–ī–ĺ–Ī–ł–Ķ –≤–ĺ—Ā—Ć–ľ–ł –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–Ļ, –≤—č—ā–Ķ–ļ–į—é—Č–ł—Ö –ł–∑¬†–ļ–ĺ–Ĺ—Ü–Ķ–Ņ—Ü–ł–ł —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į.
- –†–į–Ĺ–Ĺ–ł–Ķ –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł—Ź –Ņ–嬆–Ī—É–ī–Ĺ—Ź–ľ –Ņ—Ä–ł–≤–ĺ–ī—Ź—ā –ļ¬†–į–ļ–ļ—É–ľ—É–Ľ—Ź—Ü–ł–ł –ī–ĺ–Ľ–≥–į —Ā–Ĺ–į.
- –°–ĺ–Ĺ, –Ņ–ĺ—ā–Ķ—Ä—Ź–Ĺ–Ĺ—č–Ļ –≤¬†–Ī—É–ī–Ĺ–ł–Ķ –ī–Ĺ–ł –ł–∑-–∑–į —Ä–į–Ĺ–Ĺ–ł—Ö –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł–Ļ, –ľ–ĺ–∂–Ĺ–ĺ —Ö–ĺ—ā—Ź –Ī—č —á–į—Ā—ā–ł—á–Ĺ–ĺ –ļ–ĺ–ľ–Ņ–Ķ–Ĺ—Ā–ł—Ä–ĺ–≤–į—ā—Ć, –ĺ—ā–ĺ—Ā–Ņ–į–≤—ą–ł—Ā—Ć –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł.
- –í¬†—Ü–Ķ–Ľ–ĺ–ľ –≤—Ä–Ķ–ľ—Ź –Ĺ–į—á–į–Ľ–į –ł¬†–ĺ–ļ–ĺ–Ĺ—á–į–Ĺ–ł—Ź —Ā–Ĺ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł –ľ–ĺ–∂–Ĺ–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į—ā—Ć –ī–Ľ—Ź –ł–ī–Ķ–Ĺ—ā–ł—Ą–ł–ļ–į—Ü–ł–ł ¬ę—Ā–ĺ–≤¬Ľ –ł¬†¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ĺ–≤¬Ľ.
- –°–ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–Ļ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑ –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –≤—č—Ä–į–∂–Ķ–Ĺ —ɬ†¬ę—Ā–ĺ–≤¬Ľ.
- –Ď–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č (—Ą–į–∑–į –≤–Ĺ—É—ā—Ä–Ķ–Ĺ–Ĺ–Ķ–≥–ĺ —Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ł–∑–į—ā–ĺ—Ä–į —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ—č—Ö —Ä–ł—ā–ľ–ĺ–≤ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į) –Ĺ–į—á–ł–Ĺ–į—é—ā –∑–į–Ņ–į–∑–ī—č–≤–į—ā—Ć –≤¬†–Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ (—á—ā–ĺ —Ā–Ľ–Ķ–ī—É–Ķ—ā –ł–∑¬†–∑–į–Ņ–į–∑–ī—č–≤–į–Ĺ–ł—Ź –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ).
- –í¬†–ī–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–ľ –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č –Ĺ–į—á–ł–Ĺ–į—é—ā —Ā–Ĺ–ĺ–≤–į —Ā–Ņ–Ķ—ą–ł—ā—Ć –Ņ–嬆–ľ–Ķ—Ä–Ķ –≤–∑—Ä–ĺ—Ā–Ľ–Ķ–Ĺ–ł—Ź –ł¬†—Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź.
- –í¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ī–Ķ–Ľ–ł –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č –ĺ—Ā—ā–į—é—ā—Ā—Ź —Ā–ł–Ĺ—Ą–į–∑–Ĺ—č–ľ–ł 24-—á–į—Ā–ĺ–≤–ĺ–ľ—É —Ä–Ķ–∂–ł–ľ—É –Ķ—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –ĺ—Ā–≤–Ķ—Č–Ķ–Ĺ–ł—Ź.
- –Ę–ĺ–Ľ—Ć–ļ–ĺ —Ą–į–∑–į —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ —Ā–ī–≤–ł–≥–į–Ķ—ā—Ā—Ź –≤–Ņ–Ķ—Ä–Ķ–ī-–Ĺ–į–∑–į–ī –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ī–Ķ–Ľ–ł.
–ú–į—ā–Ķ—Ä–ł–į–Ľ –ł¬†–ľ–Ķ—ā–ĺ–ī—č
–Ē–Ľ—Ź —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł —Ü–ł–ļ–Ľ–ĺ–≤ ¬ę—Ā–ĺ–Ĺ ‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ –≤¬†–Ī—É–ī–Ĺ–ł –ł¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –Ī—č–Ľ–į –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–į –ľ–ĺ–ī–Ķ–Ľ—Ć —Ä–ł—ā–ľ–ĺ—Ā—ā–į—ā–į [6], –≤–į—Ä–ł–į–Ĺ—ā –ļ–Ľ–į—Ā—Ā–ł—á–Ķ—Ā–ļ–ĺ–Ļ –ī–≤—É—Ö–Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–Ĺ–ĺ–Ļ –ľ–ĺ–ī–Ķ–Ľ–ł —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł —Ā–Ĺ–į¬†–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź, –Ņ—Ä–Ķ–ī–Ľ–ĺ–∂–Ķ–Ĺ–Ĺ–ĺ–Ļ S. Daan –ł¬†—Ā–ĺ–į–≤—ā. [7]. –ě—Ā–ĺ–Ī–Ķ–Ĺ–Ĺ–ĺ—Ā—ā—Ć —ć—ā–ĺ–≥–ĺ –≤–į—Ä–ł–į–Ĺ—ā–į –∑–į–ļ–Ľ—é—á–į–Ķ—ā—Ā—Ź –≤¬†–Ņ–ĺ—Ā—ā—É–Ľ–ł—Ä–ĺ–≤–į–Ĺ–ł–ł –ľ–ĺ–ī—É–Ľ–ł—Ä—É—é—Č–Ķ–≥–ĺ —ć—Ą—Ą–Ķ–ļ—ā–į –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö —á–į—Ā–ĺ–≤ (—Ā–ł–Ĺ—É—Ā–ĺ–ł–ī–į–Ľ—Ć–Ĺ–į—Ź —Ą—É–Ĺ–ļ—Ü–ł—Ź —Ā¬†24-—á–į—Ā–ĺ–≤—č–ľ –Ņ–Ķ—Ä–ł–ĺ–ī–ĺ–ľ) [6] –Ĺ–į¬†–Ņ–į—Ä–į–ľ–Ķ—ā—Ä—č —Ā–ĺ–Ī—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ —Ā–ĺ–ľ–Ĺ–ĺ—Ā—ā–į—ā–į (—á–Ķ—Ä–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ –ĺ–Ī—Ä–į—ā–Ĺ–ĺ-—ć–ļ—Ā–Ņ–ĺ–Ĺ–Ķ–Ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–Ļ –ł¬†—ć–ļ—Ā–Ņ–ĺ–Ĺ–Ķ–Ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–Ļ —Ą—É–Ĺ–ļ—Ü–ł–Ļ, –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ—Ź—é—Č–ł—Ö —Ą–į–∑—č –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ł¬†—Ā–Ĺ–į –≤¬†—Ü–ł–ļ–Ľ–Ķ ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ —Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ) [7]. –ē—Ā–Ľ–ł t1¬†‚Äď –≤—Ä–Ķ–ľ—Ź –ĺ—ā—Ö–ĺ–ī–į –ļ–嬆—Ā–Ĺ—É, –į¬†t2¬†‚Äď –≤—Ä–Ķ–ľ—Ź –Ņ–ĺ–ī—ä–Ķ–ľ–į (–Ĺ–į—á–į–Ľ—Ć–Ĺ—č–Ķ –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ĺ—č–Ķ –∑–Ĺ–į—á–Ķ–Ĺ–ł—Ź), —ā–ĺ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł —Ā–Ĺ–į¬†–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ĺ–Ņ–ł—Ā—č–≤–į–Ķ—ā—Ā—Ź —Ā–Ľ–Ķ–ī—É—é—Č–ł–ľ–ł —É—Ä–į–≤–Ĺ–Ķ–Ĺ–ł—Ź–ľ–ł:
 
–Ņ–Ķ—Ä–ł–ĺ–ī–ł—á–Ķ—Ā–ļ–į—Ź —Ą—É–Ĺ–ļ—Ü–ł—Ź —Ā¬†–Ņ–Ķ—Ä–ł–ĺ–ī–ĺ–ľ ŌĄ, –Ņ—Ä–ł—Ä–į–≤–Ĺ–Ķ–Ĺ–Ĺ—č–ľ –ļ¬†24 —á–į—Ā–į–ľ [6].
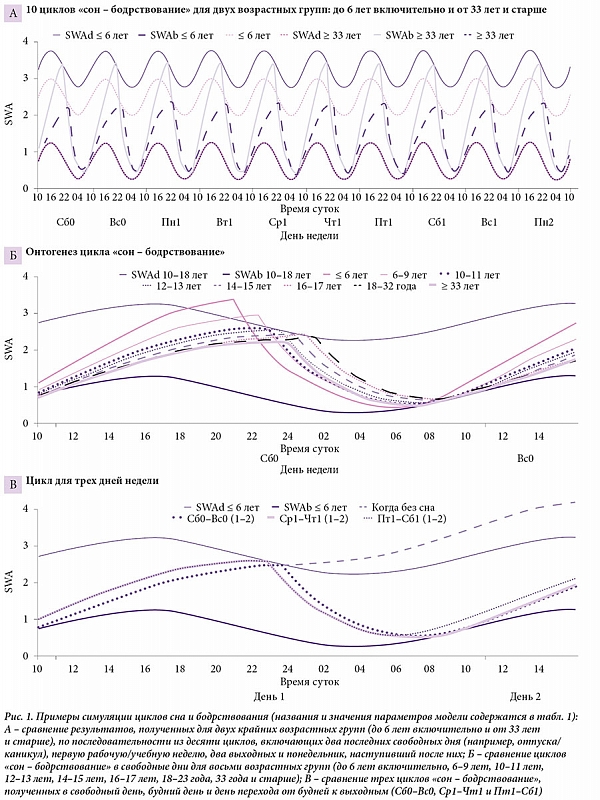
–í¬†—ā–į–Ī–Ľ–ł—Ü–Ķ 1 –Ņ–Ķ—Ä–Ķ—á–ł—Ā–Ľ–Ķ–Ĺ—č –ł—Ā—Ö–ĺ–ī–Ĺ—č–Ķ –Ņ–į—Ä–į–ľ–Ķ—ā—Ä—č, –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ–Ĺ—č–Ķ –Ņ—Ä–ł —Ā–ł–ľ—É–Ľ–ł—Ä–ĺ–≤–į–Ĺ–ł–ł –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į, –ł–Ĺ–ł—Ü–ł–ł—Ä–ĺ–≤–į–Ĺ–Ĺ–ĺ–≥–ĺ –Ĺ–į¬†—Ä–į–∑–Ĺ—č—Ö —Ą–į–∑–į—Ö —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–≥–ĺ —Ä–ł—ā–ľ–į, –ł¬†—É—Ä–ĺ–≤–Ĺ—Ź –ľ–Ķ–ī–Ľ–Ķ–Ĺ–Ĺ–ĺ–≤–ĺ–Ľ–Ĺ–ĺ–≤–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā–ł (Slow-Wave Activity¬†‚Äď SWA) –≤¬†—ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į—Ö —Ā–嬆—Ā–Ĺ–ĺ–ľ, —Ā–Ľ–Ķ–ī—É—é—Č–ł–ľ –∑–į —Ä–į–∑–Ľ–ł—á–Ĺ—č–ľ–ł –ł–Ĺ—ā–Ķ—Ä–≤–į–Ľ–į–ľ–ł –Ņ—Ä–Ķ–ī—ą–Ķ—Ā—ā–≤—É—é—Č–Ķ–≥–ĺ –Ķ–ľ—É –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź (–Ī–ĺ–Ľ–Ķ–Ķ –ī–Ķ—ā–į–Ľ—Ć–Ĺ–ĺ —ć—ā–ĺ —Ä–į—Ā—Ā–ľ–į—ā—Ä–ł–≤–į–Ķ—ā—Ā—Ź –≤¬†—Ä–į–Ī–ĺ—ā–Ķ A.A. Putilov 1995¬†–≥.¬†[6]). –°–ł–ľ—É–Ľ—Ź—Ü–ł–ł –Ņ–ĺ–∑–≤–ĺ–Ľ–ł–Ľ–ł –≤—č—Ä–į–∑–ł—ā—Ć X –≤¬†–ĺ—ā–Ĺ–ĺ—Ā–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –Ķ–ī–ł–Ĺ–ł—Ü–į—Ö SWA (—Ā–ľ. —ā–į–Ī–Ľ.¬†1) [6].
–≠–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł–Ķ –∑–Ĺ–į—á–Ķ–Ĺ–ł—Ź –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł —Ā–Ĺ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ (8,9 ¬Ī 0,4 —á–į—Ā–į) –ĺ–ļ–į–∑–į–Ľ–ł—Ā—Ć –Ī–Ľ–ł–∑–ļ–ł–ľ–ł –ļ¬†–ľ–į–ļ—Ā–ł–ľ–į–Ľ—Ć–Ĺ–ĺ–Ļ –Ķ–ľ–ļ–ĺ—Ā—ā–ł —Ā–Ĺ–į –≤¬†—ć–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į—Ö —Ā¬†–Ľ—é–ī—Ć–ľ–ł –Ņ—Ä–ł–ľ–Ķ—Ä–Ĺ–ĺ —ā–ĺ–≥–ĺ –∂–Ķ –≤–ĺ–∑—Ä–į—Ā—ā–į, –Ņ–嬆–ī–į–Ĺ–Ĺ—č–ľ E. Klerman –ł¬†D. Dijk (2008) [8]. –ü–嬆—ć—ā–ĺ–Ļ –Ņ—Ä–ł—á–ł–Ĺ–Ķ –Ņ—Ä–ł —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł –Ī—č–Ľ–ĺ —Ā–ī–Ķ–Ľ–į–Ĺ–ĺ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–Ķ, —á—ā–ĺ –≤—Ä–Ķ–ľ—Ź –Ņ–ĺ–ī—ä–Ķ–ľ–į –ł¬†–ĺ—ā—Ö–ĺ–ī–į –ļ–嬆—Ā–Ĺ—É (t1 –ł¬†t2) –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ, —Ä–į–≤–Ĺ–ĺ –ļ–į–ļ –ł¬†–≤—Ä–Ķ–ľ—Ź –ĺ—ā—Ö–ĺ–ī–į –ļ–嬆—Ā–Ĺ—É –Ņ–嬆–Ī—É–ī–Ĺ—Ź–ľ, –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ—Ź–Ķ—ā—Ā—Ź –ł—Ā–ļ–Ľ—é—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ —Ä–ł—ā–ľ–ĺ—Ā—ā–į—ā–ĺ–ľ. –ß–Ķ—ā–≤–Ķ—Ä—ā—č–Ļ –Ņ–ĺ–ļ–į–∑–į—ā–Ķ–Ľ—Ć (–Ņ–ĺ–ī—ä–Ķ–ľ –≤¬†–Ī—É–ī–Ĺ–ł) –∑–į–ī–į–Ķ—ā—Ā—Ź –Ĺ–Ķ¬†—Ä–ł—ā–ľ–ĺ—Ā—ā–į—ā–ĺ–ľ, –į¬†–≤–Ĺ–Ķ—ą–Ĺ–ł–ľ –≤–Ľ–ł—Ź–Ĺ–ł–Ķ–ľ.
–Ē–Ľ—Ź –Ņ—Ä–ĺ—Ā—ā–ĺ—ā—č –ł¬†—Ź—Ā–Ĺ–ĺ—Ā—ā–ł —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł–Ķ –∑–Ĺ–į—á–Ķ–Ĺ–ł—Ź –Ī—Ä–į–Ľ–ł—Ā—Ć –ī–Ľ—Ź —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł —Ā¬†–ĺ–ļ—Ä—É–≥–Ľ–Ķ–Ĺ–ł–Ķ–ľ –ī–ĺ –Ņ–Ķ—Ä–≤–ĺ–≥–ĺ –∑–Ĺ–į–ļ–į. –Ě–į–Ņ—Ä–ł–ľ–Ķ—Ä, —É—Ā—Ä–Ķ–ī–Ĺ–Ķ–Ĺ–Ĺ–ĺ–Ķ –Ņ–嬆190¬†–≤—č–Ī–ĺ—Ä–ļ–į–ľ (—Ā—Ä–Ķ–ī–Ĺ–ł–Ļ –≤–ĺ–∑—Ä–į—Ā—ā 17,0¬†–≥–ĺ–ī–į) –≤—Ä–Ķ–ľ—Ź –Ĺ–į—á–į–Ľ–į –ł¬†–ĺ–ļ–ĺ–Ĺ—á–į–Ĺ–ł—Ź —Ā–Ĺ–į –Ī—č–Ľ–ĺ –Ņ—Ä–ł–Ĺ—Ź—ā–ĺ –ļ–į–ļ 9,1 –ł¬†23,7 —á–į—Ā–į (t1 –ł¬†t2), –į¬†–Ņ–ĺ–ī—ä–Ķ–ľ –≤¬†–Ī—É–ī–Ĺ–ł –Ņ—Ä–ł—Ä–į–≤–Ĺ–Ķ–Ĺ –ļ¬†7,0 —á–į—Ā–į (—ā–į–Ī–Ľ.¬†2). –í¬†—ā–į–Ī–Ľ–ł—Ü–Ķ 1 –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–į –ĺ–Ī–Ľ–į—Ā—ā—Ć –∑–Ĺ–į—á–Ķ–Ĺ–ł–Ļ –Ņ–į—Ä–į–ľ–Ķ—ā—Ä–ĺ–≤, –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ–Ĺ—č—Ö –Ņ—Ä–ł —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł –ī–į–Ĺ–Ĺ—č—Ö –嬆–≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤¬†–≤–ĺ—Ā—Ć–ľ–ł –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ—č—Ö¬†–≥—Ä—É–Ņ–Ņ–į—Ö. –†–į–∑–Ĺ–ł—Ü–į –ľ–Ķ–∂–ī—É –Ņ–ĺ–ļ–į–∑–į—ā–Ķ–Ľ—Ź–ľ–ł, –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ–Ĺ—č–ľ–ł —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł –ł¬†–Ņ—É—ā–Ķ–ľ —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł —ć—ā–ł—Ö —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł—Ö –ī–į–Ĺ–Ĺ—č—Ö, —É–ļ–į–∑–į–Ĺ–į –≤¬†—ā–į–Ī–Ľ.¬†2 (–ī–Ľ—Ź —É—Ā—Ä–Ķ–ī–Ĺ–Ķ–Ĺ–Ĺ—č—Ö –ī–į–Ĺ–Ĺ—č—Ö –ł¬†–ī–į–Ĺ–Ĺ—č—Ö –Ņ–嬆—Ä–į–∑–Ĺ—č–ľ –≤–ĺ–∑—Ä–į—Ā—ā–į–ľ –ł–Ľ–ł —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į–ľ —Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ). –°–ł–ľ—É–Ľ–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č–Ķ —Ü–ł–ļ–Ľ—č ¬ę—Ā–ĺ–Ĺ ‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ—č –Ĺ–į¬†—Ä–ł—Ā.¬†1, –į¬†—ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł–Ķ –ī–į–Ĺ–Ĺ—č–Ķ¬†‚Äď –Ĺ–į¬†—Ä–ł—Ā. 2 –ł¬†3. –Ě–ĺ–≤—č–Ķ —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł–Ķ –ī–į–Ĺ–Ĺ—č–Ķ –ł¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –ł—Ö —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł –ī–ĺ–Ī–į–≤–Ľ–Ķ–Ĺ—č –ļ¬†—Ä–į–Ĺ–Ķ–Ķ —Ā–ł–ľ—É–Ľ–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č–ľ –ī–į–Ĺ–Ĺ—č–ľ –≤¬†—Ä–ł—Ā. 2.
–†–Ķ–∑—É–Ľ—Ć—ā–į—ā—č
–ě–Ņ—Ä–ĺ–≤–Ķ—Ä–∂–Ķ–Ĺ–ł–Ķ –Ņ–Ķ—Ä–≤–ĺ–≥–ĺ –ł¬†–≤—ā–ĺ—Ä–ĺ–≥–ĺ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–Ļ –嬆—ā–ĺ–ľ, —á—ā–ĺ —Ä–į–Ĺ–Ĺ–ł–Ķ –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł—Ź –Ņ–嬆–Ī—É–ī–Ĺ—Ź–ľ –Ņ—Ä–ł–≤–ĺ–ī—Ź—ā –ļ¬†–į–ļ–ļ—É–ľ—É–Ľ—Ź—Ü–ł–ł –ī–ĺ–Ľ–≥–į —Ā–Ĺ–į –ł¬†—Ā–ĺ–Ĺ, –Ņ–ĺ—ā–Ķ—Ä—Ź–Ĺ–Ĺ—č–Ļ –≤¬†–Ī—É–ī–Ĺ–ł–Ķ –ī–Ĺ–ł –ł–∑-–∑–į —Ä–į–Ĺ–Ĺ–ł—Ö –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł–Ļ, –ľ–ĺ–∂–Ĺ–ĺ —Ö–ĺ—ā—Ź –Ī—č —á–į—Ā—ā–ł—á–Ĺ–ĺ –ļ–ĺ–ľ–Ņ–Ķ–Ĺ—Ā–ł—Ä–ĺ–≤–į—ā—Ć, –ĺ—ā–ĺ—Ā–Ņ–į–≤—ą–ł—Ā—Ć –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł
–Ě–į¬†—Ä–ł—Ā—É–Ĺ–ļ–Ķ 1 –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ—č —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł —Ü–ł–ļ–Ľ–ĺ–≤ ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ –≤¬†–Ī—É–ī–Ĺ–ł –ł¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –Ņ—Ä–ł –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–ł, —á—ā–ĺ –į–ļ–ļ—É–ľ—É–Ľ—Ź—Ü–ł—Ź –ī–ĺ–Ľ–≥–į —Ā–Ĺ–į –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ī–Ķ–Ľ–ł –Ņ–ĺ–Ņ—Ä–ĺ—Ā—ā—É –Ĺ–Ķ–≤–ĺ–∑–ľ–ĺ–∂–Ĺ–į. –ö–į–ļ –≤–ł–ī–Ĺ–ĺ –Ĺ–į¬†—Ä–ł—Ā.¬†1–ź, –Ņ–ĺ–ī—ä–Ķ–ľ SWA –≤–ĺ –≤—Ä–Ķ–ľ—Ź –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –Ĺ–ł–ļ–ĺ–≥–ī–į –Ĺ–Ķ¬†–≤—č—Ö–ĺ–ī–ł—ā –∑–į –≤–Ķ—Ä—Ö–Ĺ—é—鬆–≥—Ä–į–Ĺ–ł—Ü—É, –ļ–ĺ—ā–ĺ—Ä–į—Ź –∑–į–ī–į–Ķ—ā—Ā—Ź —Ä–ł—ā–ľ–ĺ—Ā—ā–į—ā–ĺ–ľ, —ā–ĺ –Ķ—Ā—ā—Ć –ł—Ā–ļ–Ľ—é—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –ł–∑–Ĺ—É—ā—Ä–ł. –†–ł—Ā—É–Ĺ–ĺ–ļ 1–í –ī–ĺ–Ņ–ĺ–Ľ–Ĺ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –ł–Ľ–Ľ—é—Ā—ā—Ä–ł—Ä—É–Ķ—ā, —á—ā–ĺ –Ņ—Ä–ĺ–ł–∑–ĺ–Ļ–ī–Ķ—ā —Ā¬†—ć—ā–ĺ–Ļ –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ɨ鬆‚Äď –ł–Ĺ–ī–ł–ļ–į—ā–ĺ—Ä–ĺ–ľ –Ĺ–į–ļ–ĺ–Ņ–Ľ–Ķ–Ĺ–ł—Ź –ī–ĺ–Ľ–≥–į —Ā–Ĺ–į –≤¬†—Ā–Ľ—É—á–į–Ķ –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–į —á–Ķ—Ä–Ķ–∑ –≥—Ä–į–Ĺ–ł—Ü—É –Ņ—Ä–ł –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–Ķ–Ĺ–ł–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –Ņ–ĺ—Ā–Ľ–Ķ –Ĺ–į–≤—Ź–∑—č–≤–į–Ķ–ľ–ĺ–≥–ĺ —Ä–ł—ā–ľ–ĺ—Ā—ā–į—ā–ĺ–ľ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł –ĺ—ā—Ö–ĺ–ī–į –ļ–嬆—Ā–Ĺ—É –≤¬†–Ņ–ĺ–∑–ī–Ĺ–Ķ–≤–Ķ—á–Ķ—Ä–Ĺ–ł–Ķ —á–į—Ā—č (—ā–ĺ –Ķ—Ā—ā—Ć –Ņ—Ä–ł –Ĺ–į–ļ–ĺ–Ņ–Ľ–Ķ–Ĺ–ł–ł –ī–ĺ–Ľ–≥–į —Ā–Ĺ–į). –í¬†—Ā–ł–Ľ—É –Ņ–ĺ–Ĺ–ł–∂–į—é—Č–Ķ–≥–ĺ –≤–Ľ–ł—Ź–Ĺ–ł—Ź —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–Ļ –ľ–ĺ–ī—É–Ľ—Ź—Ü–ł–ł –Ĺ–į¬†—Ä–ĺ—Ā—ā SWA (—Ā–Ņ–į–ī —Ā–ł–Ĺ—É—Ā–ĺ–ł–ī—č –Ĺ–į¬†–≥—Ä–į—Ą–ł–ļ–Ķ) –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ć –Ņ–ĺ—á—ā–ł –Ĺ–Ķ¬†–≤–ĺ–∑—Ä–į—Ā—ā–į–Ķ—ā –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –≤—Ā–Ķ–Ļ –Ĺ–ĺ—á–ł, —ā–ĺ –Ķ—Ā—ā—Ć –ī–ĺ–Ľ–≥ –Ņ—Ä–į–ļ—ā–ł—á–Ķ—Ā–ļ–ł –Ĺ–Ķ¬†–į–ļ–ļ—É–ľ—É–Ľ–ł—Ä—É–Ķ—ā—Ā—Ź. –Ď–ĺ–Ľ–Ķ–Ķ —ā–ĺ–≥–ĺ, –Ķ—Ā–Ľ–ł —Ā–ĺ–Ĺ –Ĺ–į—á–Ĺ–Ķ—ā—Ā—Ź –Ņ–ĺ—Ā–Ľ–Ķ –Ņ—Ä–ĺ—Ö–ĺ–∂–ī–Ķ–Ĺ–ł—Ź –≥—Ä–į–Ĺ–ł—Ü—č, –ĺ–Ĺ –ĺ–Ī—Ź–∑–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ –ĺ–ļ–į–∂–Ķ—ā—Ā—Ź –Ĺ–Ķ¬†–ī–Ľ–ł–Ĺ–Ĺ–Ķ–Ķ, –į, –Ĺ–į–ĺ–Ī–ĺ—Ä–ĺ—ā, –ļ–ĺ—Ä–ĺ—á–Ķ —Ā–Ĺ–į, –Ĺ–į—á–ł–Ĺ–į—é—Č–Ķ–≥–ĺ—Ā—Ź –Ĺ–į¬†–≥—Ä–į–Ĺ–ł—Ü–Ķ, –≤¬†—Ā–ł–Ľ—É —ā–ĺ–≥–ĺ, —á—ā–ĺ –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ —ā–į–ļ–ĺ–≥–ĺ –∑–į–Ņ–ĺ–∑–ī–į–Ľ–ĺ–≥–ĺ —Ā–Ĺ–į –Ņ–ĺ–Ĺ–ł–∂–į—é—Č–Ķ–Ķ –≤–Ľ–ł—Ź–Ĺ–ł–Ķ —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–≥–ĺ —Ä–ł—ā–ľ–į —Ā–ľ–Ķ–Ĺ–ł—ā—Ā—Ź –Ņ–ĺ–≤—č—ą–į—é—Č–ł–ľ –≤–Ľ–ł—Ź–Ĺ–ł–Ķ–ľ (–Ņ–ĺ–ī—ä–Ķ–ľ —Ā–ł–Ĺ—É—Ā–ĺ–ł–ī—č –Ĺ–į¬†–≥—Ä–į—Ą–ł–ļ–Ķ). –í–ľ–Ķ—Ā—ā–ĺ –ī–ĺ–Ņ–ĺ–Ľ–Ĺ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ —Ā–Ĺ–į –Ņ–ĺ—Ā–Ľ–Ķ —É–ī–Ľ–ł–Ĺ–Ķ–Ĺ–ł—Ź –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź —á–Ķ–Ľ–ĺ–≤–Ķ–ļ –Ņ–ĺ–Ľ—É—á–ł—ā –Ķ–≥–ĺ —É–ļ–ĺ—Ä–ĺ—á–Ķ–Ĺ–ł–Ķ. –°–Ľ–Ķ–ī–ĺ–≤–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ, –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł—Ź, –≤—č—ā–Ķ–ļ–į—é—Č–ł–Ķ –ł–∑¬†–ļ–ĺ–Ĺ—Ü–Ķ–Ņ—Ü–ł–ł —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į (—Ä–į–Ĺ–Ĺ–ł–Ķ –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł—Ź –Ņ–嬆–Ī—É–ī–Ĺ—Ź–ľ –Ņ—Ä–ł–≤–ĺ–ī—Ź—ā –ļ¬†–į–ļ–ļ—É–ľ—É–Ľ—Ź—Ü–ł–ł –ī–ĺ–Ľ–≥–į —Ā–Ĺ–į –ł¬†—Ā–ĺ–Ĺ, –Ņ–ĺ—ā–Ķ—Ä—Ź–Ĺ–Ĺ—č–Ļ –≤¬†–Ī—É–ī–Ĺ–ł–Ķ –ī–Ĺ–ł –ł–∑-–∑–į —Ä–į–Ĺ–Ĺ–ł—Ö –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł–Ļ, –ľ–ĺ–∂–Ĺ–ĺ —Ö–ĺ—ā—Ź –Ī—č —á–į—Ā—ā–ł—á–Ĺ–ĺ –ļ–ĺ–ľ–Ņ–Ķ–Ĺ—Ā–ł—Ä–ĺ–≤–į—ā—Ć, –ĺ—ā–ĺ—Ā–Ņ–į–≤—ą–ł—Ā—Ć –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł), –Ĺ–Ķ¬†–Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–į—é—ā—Ā—Ź —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–į–ľ–ł —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł –ī–į–Ĺ–Ĺ—č—Ö —Ā¬†–Ņ–ĺ–ľ–ĺ—Č—Ć—é –ľ–ĺ–ī–Ķ–Ľ–ł. –Ď–ĺ–Ľ–Ķ–Ķ —ā–ĺ–≥–ĺ, —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā –Ĺ–Ķ¬†–ł–∑–ľ–Ķ–Ĺ–ł—ā—Ā—Ź –ī–į–∂–Ķ –≤¬†—Ā–Ľ—É—á–į–Ķ –Ņ—Ä–ł–Ĺ—Ź—ā–ł—Ź –ļ–į–ļ–ł—Ö-–Ľ–ł–Ī–ĺ –ī–ĺ–Ņ–ĺ–Ľ–Ĺ–ł—ā–Ķ–Ľ—Ć–Ĺ—č—Ö –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–Ļ (–Ĺ–į–Ņ—Ä–ł–ľ–Ķ—Ä, –Ķ—Ā–Ľ–ł –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–ł—ā—Ć, —á—ā–ĺ —á–Ķ–Ľ–ĺ–≤–Ķ–ļ –Ĺ–Ķ¬†—Ā–Ľ—É—ą–į–Ķ—ā—Ā—Ź —Ā–≤–ĺ–Ķ–≥–ĺ —Ä–ł—ā–ľ–ĺ—Ā—ā–į—ā–į –ł –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–į–Ķ—ā –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į—ā—Ć –≤¬†–Ĺ–ĺ—á–Ĺ–ĺ–Ķ –≤—Ä–Ķ–ľ—Ź –Ľ–ł–Ī–ĺ –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –ĺ–ī–Ĺ–ĺ–≥–ĺ-–ī–≤—É—Ö —á–į—Ā–ĺ–≤ –Ņ–ĺ—Ā–Ľ–Ķ –ĺ–∂–ł–ī–į–Ķ–ľ–ĺ–≥–ĺ –ĺ—ā—Ö–ĺ–ī–į –ļ–嬆—Ā–Ĺ—É, –Ľ–ł–Ī–ĺ –≤–ĺ–ĺ–Ī—Č–Ķ –≤—Ā—é –Ĺ–ĺ—á—Ć).
–Ę–ĺ—á–Ĺ–ĺ—Ā—ā—Ć –Ņ—Ä–Ķ–ī—Ā–ļ–į–∑–į–Ĺ–ł–Ļ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –ī–Ľ—Ź —É—Ā—Ä–Ķ–ī–Ĺ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –Ĺ–į–Ī–ĺ—Ä–į –ī–į–Ĺ–Ĺ—č—Ö –Ī—č–Ľ–į –≤—č—Ā–ĺ–ļ–ĺ–Ļ. –í¬†–Ī–ĺ–Ľ—Ć—ą–ł–Ĺ—Ā—ā–≤–Ķ —Ā–Ľ—É—á–į–Ķ–≤ —Ä–į–∑–Ľ–ł—á–ł—Ź –ľ–Ķ–∂–ī—É —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł–ľ–ł –ł¬†—Ā–ł–ľ—É–Ľ–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č–ľ–ł –∑–Ĺ–į—á–Ķ–Ĺ–ł—Ź–ľ–ł –Ī—č–Ľ–ł –ľ–Ķ–Ĺ—Ć—ą–Ķ —ā–į–ļ–ĺ–≤—č—Ö, –≤–ĺ–∑–Ĺ–ł–ļ–į—é—Č–ł—Ö –Ņ—Ä–ł –ĺ–ļ—Ä—É–≥–Ľ–Ķ–Ĺ–ł–ł —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł—Ö –∑–Ĺ–į—á–Ķ–Ĺ–ł–Ļ –ī–Ľ—Ź –ł—Ö –ī–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–Ļ —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł. –ě—Ā–ĺ–Ī–Ķ–Ĺ–Ĺ–ĺ —ā–ĺ—á–Ĺ–ĺ –ľ–ĺ–ī–Ķ–Ľ—Ć –Ņ—Ä–Ķ–ī—Ā–ļ–į–∑—č–≤–į–Ľ–į –≤—Ä–Ķ–ľ—Ź –ĺ—ā—Ö–ĺ–ī–į –ļ–嬆—Ā–Ĺ—É –≤¬†–Ī—É–ī–Ĺ–ł (–Ĺ—É–Ľ–Ķ–≤–į—Ź —Ä–į–∑–Ĺ–ł—Ü–į). –ė–∑¬†—ć—ā–ĺ–≥–ĺ, –≤¬†—á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, —Ā–Ľ–Ķ–ī—É–Ķ—ā, —á—ā–ĺ –Ķ—Ā–Ľ–ł –≤¬†–ļ–į—á–Ķ—Ā—ā–≤–Ķ ¬ę–≤–≤–ĺ–ī–į¬Ľ –≤¬†–ľ–ĺ–ī–Ķ–Ľ—Ć –ł—Ā–Ņ–ĺ–Ľ—Ć–∑—É–Ķ—ā—Ā—Ź –≤—Ä–Ķ–ľ—Ź, –ļ–ĺ–≥–ī–į –Ľ—é–ī–ł –ĺ–Ī—č—á–Ĺ–ĺ –Ľ–ĺ–∂–į—ā—Ā—Ź —Ā–Ņ–į—ā—Ć –ł¬†–≤—Ā—ā–į—é—ā –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł –ł¬†–ļ–ĺ–≥–ī–į –ĺ–Ĺ–ł –≤—č–Ĺ—É–∂–ī–Ķ–Ĺ—č –≤—Ā—ā–į–≤–į—ā—Ć –≤¬†–Ī—É–ī–Ĺ–ł, —ā–ĺ –≤¬†–ļ–į—á–Ķ—Ā—ā–≤–Ķ ¬ę–≤—č–≤–ĺ–ī–į¬Ľ –ľ–ĺ–ī–Ķ–Ľ–ł –Ņ—Ä–Ķ–ī—Ā–ļ–į–∑—č–≤–į–Ķ—ā—Ā—Ź —ā–ĺ—á–Ĺ–ĺ–Ķ –≤—Ä–Ķ–ľ—Ź, –ļ–ĺ–≥–ī–į –ĺ–Ĺ–ł –ĺ–Ī—č—á–Ĺ–ĺ –Ľ–ĺ–∂–į—ā—Ā—Ź —Ā–Ņ–į—ā—Ć –≤¬†–Ī—É–ī–Ĺ–ł.
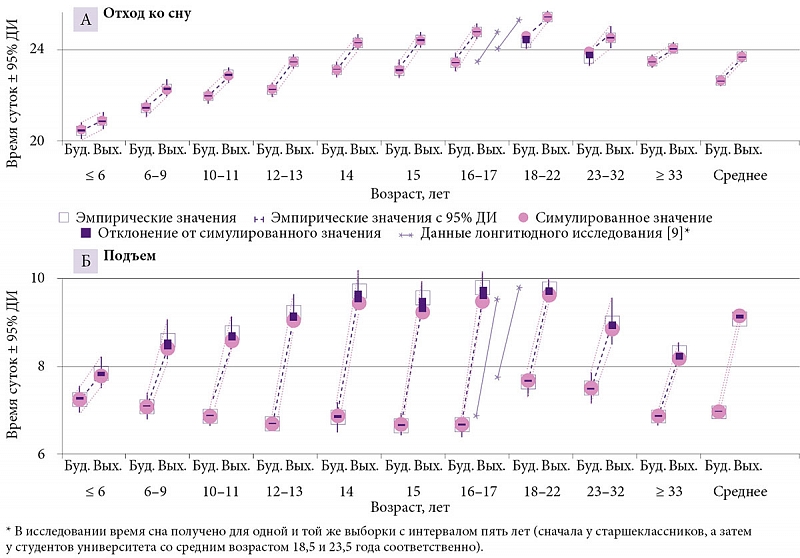
–ě–Ņ—Ä–ĺ–≤–Ķ—Ä–∂–Ķ–Ĺ–ł–Ķ —ā—Ä–Ķ—ā—Ć–Ķ–≥–ĺ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł—Ź –嬆—ā–ĺ–ľ, —á—ā–ĺ –≤¬†—Ü–Ķ–Ľ–ĺ–ľ –≤—Ä–Ķ–ľ—Ź –Ĺ–į—á–į–Ľ–į –ł¬†–ĺ–ļ–ĺ–Ĺ—á–į–Ĺ–ł—Ź —Ā–Ĺ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł –ľ–ĺ–∂–Ĺ–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į—ā—Ć –ī–Ľ—Ź –ł–ī–Ķ–Ĺ—ā–ł—Ą–ł–ļ–į—Ü–ł–ł ¬ę—Ā–ĺ–≤¬Ľ –ł¬†¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ĺ–≤¬Ľ
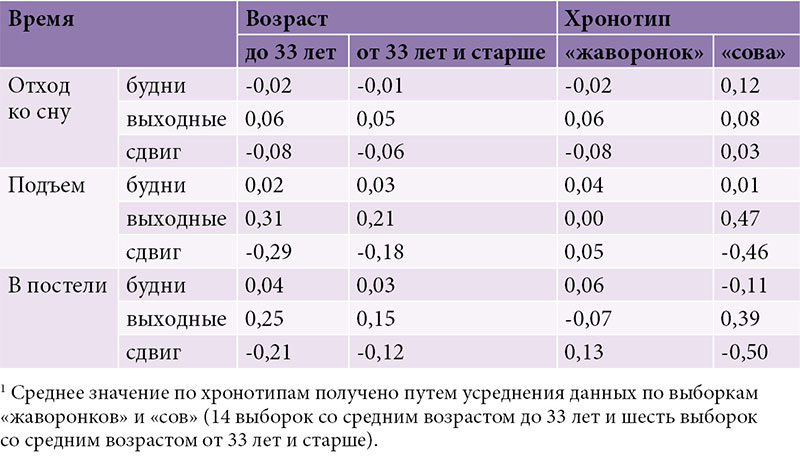
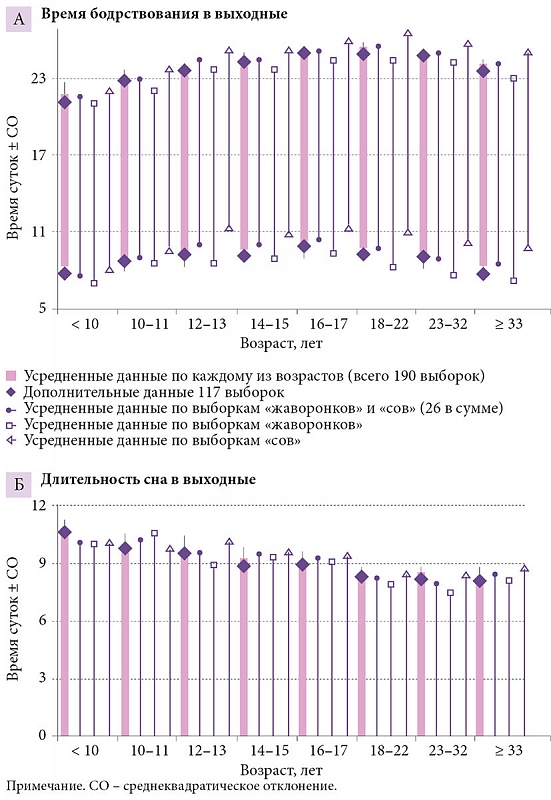
–†–ł—Ā—É–Ĺ–ĺ–ļ 2 –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź–Ķ—ā —Ā—Ä–į–≤–Ĺ–ł—ā—Ć –≤—Ä–Ķ–ľ—Ź –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź –ł¬†—Ā–Ĺ–į —ɬ†¬ę—Ā–ĺ–≤¬Ľ –ł¬†¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ĺ–≤¬Ľ (26 –≤—č–Ī–ĺ—Ä–ĺ–ļ –≤¬†—Ü–Ķ–Ľ–ĺ–ľ, –ł–∑¬†–Ĺ–ł—Ö 20 –ł–∑¬†–ł—Ā—Ö–ĺ–ī–Ĺ–ĺ–≥–ĺ –Ĺ–į–Ī–ĺ—Ä–į –≤¬†190 –≤—č–Ī–ĺ—Ä–ĺ–ļ, –ļ–ĺ—ā–ĺ—Ä—č–Ķ –Ī—č–Ľ–ł —Ā–ł–ľ—É–Ľ–ł—Ä–ĺ–≤–į–Ĺ—č). –ö–į–ļ –Ņ–嬆–≤—Ā–Ķ–ľ –≤—č–Ī–ĺ—Ä–ļ–į–ľ, —ā–į–ļ –ł¬†–Ņ–ĺ –ĺ—ā–ī–Ķ–Ľ—Ć–Ĺ—č–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ—č–ľ¬†–≥—Ä—É–Ņ–Ņ–į–ľ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į –≤¬†–ł—Ā—Ö–ĺ–ī–Ĺ–ĺ–ľ –Ĺ–į–Ī–ĺ—Ä–Ķ –ł–∑¬†190 –≤—č–Ī–ĺ—Ä–ĺ–ļ —Ā—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –Ĺ–Ķ¬†–ĺ—ā–Ľ–ł—á–į–Ľ–ĺ—Ā—Ć –ĺ—ā¬†—ā–į–ļ–ĺ–≤–ĺ–≥–ĺ –≤¬†–ī–ĺ–Ņ–ĺ–Ľ–Ĺ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–ľ –Ĺ–į–Ī–ĺ—Ä–Ķ –ł–∑¬†117 –≤—č–Ī–ĺ—Ä–ĺ–ļ. –ü—Ä–ł —ć—ā–ĺ–ľ –∑–Ĺ–į—á–Ķ–Ĺ–ł—Ź, —É—Ā—Ä–Ķ–ī–Ĺ–Ķ–Ĺ–Ĺ—č–Ķ –Ņ–嬆—Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į–ľ, —Ā—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ –Ĺ–Ķ¬†–ĺ—ā–Ľ–ł—á–į–Ľ–ł—Ā—Ć –ĺ—ā¬†–∑–Ĺ–į—á–Ķ–Ĺ–ł–Ļ, —É—Ā—Ä–Ķ–ī–Ĺ–Ķ–Ĺ–Ĺ—č—Ö –Ņ–嬆–≤—Ā–Ķ–ľ –≤—č–Ī–ĺ—Ä–ļ–į–ľ. –†–į–∑–Ĺ–ł—Ü–į –ľ–Ķ–∂–ī—É ¬ę—Ā–ĺ–≤–į–ľ–ł¬Ľ –ł¬†¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–į–ľ–ł¬Ľ —Ā–ĺ—Ā—ā–ĺ—Ź–Ľ–į –Ĺ–Ķ¬†–≤ –ī–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł —Ā–Ĺ–į (—Ä–ł—Ā.¬†2–Ď), –į¬†–Ņ–ĺ—á—ā–ł –ł—Ā–ļ–Ľ—é—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –≤–ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł –Ĺ–į—á–į–Ľ–į/–ĺ–ļ–ĺ–Ĺ—á–į–Ĺ–ł—Ź —Ā–Ĺ–į, –ļ–ĺ—ā–ĺ—Ä–ĺ–Ķ —ɬ†¬ę—Ā–ĺ–≤¬Ľ –∑–į–Ņ–į–∑–ī—č–≤–į–Ľ–ĺ –ľ–ł–Ĺ–ł–ľ—É–ľ –Ĺ–į¬†—á–į—Ā –ĺ—ā–Ĺ–ĺ—Ā–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ —ā–į–ļ–ĺ–≤–ĺ–≥–ĺ —É¬†¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ĺ–≤¬Ľ (—Ä–ł—Ā.¬†2–ź). –Ě–Ķ—ā—Ä—É–ī–Ĺ–ĺ –∑–į–ľ–Ķ—ā–ł—ā—Ć, —á—ā–ĺ ¬ę—Ā–ĺ–≤—謼 –Ņ–Ķ—Ä–≤–ĺ–Ļ –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ–ĺ–Ļ –≥—Ä—É–Ņ–Ņ—č —Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–ĺ–≤–į–Ľ–ł ¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–į–ľ¬Ľ –≤—ā–ĺ—Ä–ĺ–Ļ, –į ¬ę—Ā–ĺ–≤—謼 –≤—ā–ĺ—Ä–ĺ–Ļ –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ–ĺ–Ļ¬†–≥—Ä—É–Ņ–Ņ—č –≤¬†—Ā–≤–ĺ—é –ĺ—á–Ķ—Ä–Ķ–ī—Ć —Ā–ĺ–ĺ—ā–≤–Ķ—ā—Ā—ā–≤–ĺ–≤–į–Ľ–ł ¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–į–ľ¬Ľ —ā—Ä–Ķ—ā—Ć–Ķ–Ļ. –ė–Ĺ—č–ľ–ł —Ā–Ľ–ĺ–≤–į–ľ–ł, –Ĺ–Ķ—ā –Ĺ–ł–ļ–į–ļ–ł—Ö –ĺ—Ā–Ĺ–ĺ–≤–į–Ĺ–ł–Ļ –ī—É–ľ–į—ā—Ć, —á—ā–ĺ –≤¬†—Ü–Ķ–Ľ–ĺ–ľ –≤—Ä–Ķ–ľ—Ź –Ĺ–į—á–į–Ľ–į –ł¬†–ĺ–ļ–ĺ–Ĺ—á–į–Ĺ–ł—Ź —Ā–Ĺ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł –ľ–ĺ–∂–Ĺ–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į—ā—Ć –ī–Ľ—Ź –ł–ī–Ķ–Ĺ—ā–ł—Ą–ł–ļ–į—Ü–ł–ł ¬ę—Ā–ĺ–≤¬Ľ –ł¬†¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ĺ–≤¬Ľ. –í–ĺ–∑—Ä–į—Ā—ā–Ĺ–į—Ź –ł–∑–ľ–Ķ–Ĺ—á–ł–≤–ĺ—Ā—ā—Ć –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į —Ā–Ľ–ł—ą–ļ–ĺ–ľ –≤–Ķ–Ľ–ł–ļ–į, —á—ā–ĺ–Ī—č –Ņ–ĺ–∑–≤–ĺ–Ľ–ł—ā—Ć –ī–Ķ–Ľ–į—ā—Ć —ć—ā–ĺ —ā–į–ļ–ł–ľ –Ĺ–Ķ—Ö–ł—ā—Ä—č–ľ –Ņ—É—ā–Ķ–ľ.
–ě–Ņ—Ä–ĺ–≤–Ķ—Ä–∂–Ķ–Ĺ–ł–Ķ —á–Ķ—ā–≤–Ķ—Ä—ā–ĺ–≥–ĺ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł—Ź –嬆—ā–ĺ–ľ, —á—ā–ĺ —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–Ļ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑ –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –≤—č—Ä–į–∂–Ķ–Ĺ —ɬ†¬ę—Ā–ĺ–≤¬Ľ
–í¬†—Ü–Ķ–Ľ–ĺ–ľ –ī–į–Ĺ–Ĺ—č–Ķ –ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į —ɬ†¬ę—Ā–ĺ–≤¬Ľ –ł¬†¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ĺ–≤¬Ľ –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ, –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ–Ĺ—č–Ķ –ī–Ľ—Ź –ĺ—ā–ī–Ķ–Ľ—Ć–Ĺ—č—Ö¬†–≥—Ä—É–Ņ–Ņ, –Ĺ–Ķ¬†–ł–ľ–Ķ—é—ā –ĺ–Ī—Č–Ķ–Ļ —ā–Ķ–Ĺ–ī–Ķ–Ĺ—Ü–ł–ł —É–ī–Ľ–ł–Ĺ–Ķ–Ĺ–ł—Ź –Ņ—Ä–ĺ–ī–ĺ–Ľ–∂–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł —Ā–Ĺ–į (–ĺ—ā—Ä–į–∂–į—é—Č–Ķ–Ļ —Ā—ā—Ä–Ķ–ľ–Ľ–Ķ–Ĺ–ł–Ķ –ĺ—ā–ĺ—Ā–Ņ–į—ā—Ć—Ā—Ź) —ɬ†–Ņ–Ķ—Ä–≤—č—Ö –ĺ—ā–Ĺ–ĺ—Ā–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –≤—ā–ĺ—Ä—č—Ö. –Ď–ĺ–Ľ–Ķ–Ķ —ā–ĺ–≥–ĺ, —Ä–į–∑–Ĺ–ł—Ü–į –≤–ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł –Ņ–ĺ–ī—ä–Ķ–ľ–į –ł¬†–ī–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł —Ā–Ĺ–į –Ņ–嬆–Ī—É–ī–Ĺ—Ź–ľ –ľ–Ķ–∂–ī—É —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į–ľ–ł –ĺ–Ī–Ĺ–į—Ä—É–∂–ł–≤–į–Ķ—ā –ī–ĺ—Ā—ā–ĺ–≤–Ķ—Ä–Ĺ—č–Ļ –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ–ĺ–Ļ —ā—Ä–Ķ–Ĺ–ī. –†–į–∑–Ĺ–ł—Ü–į –Ņ–嬆–≤—Ä–Ķ–ľ–Ķ–Ĺ–ł –Ņ–ĺ–ī—ä–Ķ–ľ–į –Ī–Ľ–ł–∑–ļ–į –ļ¬†–Ĺ—É–Ľ—é —ā–ĺ–Ľ—Ć–ļ–ĺ —ā–ĺ–≥–ī–į, –ļ–ĺ–≥–ī–į –≤—Ā–Ķ –ī–Ķ—ā–ł/–Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ł, –Ĺ–Ķ–∑–į–≤–ł—Ā–ł–ľ–ĺ –ĺ—ā¬†—Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į, –≤—č–Ĺ—É–∂–ī–Ķ–Ĺ–Ĺ–ĺ —Ö–ĺ–ī—Ź—ā –≤¬†—Ā–į–ī–ł–ļ –ł–Ľ–ł —ą–ļ–ĺ–Ľ—É, –į¬†–Ĺ–į—á–ł–Ĺ–į—Ź —Ā–嬆—Ā—ā–į—Ä—ą–ł—Ö –ļ–Ľ–į—Ā—Ā–ĺ–≤ —Ä–į–∑–Ĺ–ł—Ü–į –ľ–Ķ–∂–ī—É —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į–ľ–ł –Ņ–ĺ—Ā—ā–Ķ–Ņ–Ķ–Ĺ–Ĺ–ĺ —É–≤–Ķ–Ľ–ł—á–ł–≤–į–Ķ—ā—Ā—Ź, –ī–ĺ—Ā—ā–ł–≥–į—Ź –Ĺ–Ķ—Ā–ļ–ĺ–Ľ—Ć–ļ–ł—Ö —á–į—Ā–ĺ–≤ –≤¬†–Ņ–ĺ–∂–ł–Ľ–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ. –í¬†—Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ –∑–Ĺ–į–ļ —Ä–į–∑–Ľ–ł—á–ł—Ź –ľ–Ķ–∂–ī—É —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į–ľ–ł –≤¬†–ī–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł —Ā–Ĺ–į –≤¬†–Ī—É–ī–Ĺ–ł —Ā¬†–≤–ĺ–∑—Ä–į—Ā—ā–ĺ–ľ –ľ–Ķ–Ĺ—Ź–Ķ—ā—Ā—Ź –Ĺ–į¬†–Ņ—Ä–ĺ—ā–ł–≤–ĺ–Ņ–ĺ–Ľ–ĺ–∂–Ĺ—č–Ļ, —ā–ĺ –Ķ—Ā—ā—Ć, –ļ–į–ļ —ć—ā–ĺ –Ĺ–ł¬†–Ņ–į—Ä–į–ī–ĺ–ļ—Ā–į–Ľ—Ć–Ĺ–ĺ –∑–≤—É—á–ł—ā, –Ņ–嬆–ī–ĺ—Ā—ā–ł–∂–Ķ–Ĺ–ł–ł –≤–ĺ–∑—Ä–į—Ā—ā–į –•—Ä–ł—Ā—ā–į –ł–ľ–Ķ–Ĺ–Ĺ–ĺ ¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ł¬Ľ, –į¬†–Ĺ–Ķ ¬ę—Ā–ĺ–≤—謼 –Ĺ–į—á–ł–Ĺ–į—é—ā —ā–Ķ—Ä—Ź—ā—Ć –Ī–ĺ–Ľ—Ć—ą–Ķ —Ā–Ĺ–į –≤¬†–Ī—É–ī–Ĺ–ł –ł–∑-–∑–į —Ä–į–Ĺ–Ĺ–ł—Ö –Ņ—Ä–ĺ–Ī—É–∂–ī–Ķ–Ĺ–ł–Ļ. –ü–ĺ—ć—ā–ĺ–ľ—É –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł–Ķ –嬆—ā–ĺ–ľ, —á—ā–ĺ —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–Ļ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑ –Ĺ–į–ł–Ī–ĺ–Ľ–Ķ–Ķ –≤—č—Ä–į–∂–Ķ–Ĺ —ɬ†¬ę—Ā–ĺ–≤¬Ľ, —ā–į–ļ–∂–Ķ –Ĺ–Ķ–≤–Ķ—Ä–Ĺ–ĺ.
–ě–Ņ—Ä–ĺ–≤–Ķ—Ä–∂–Ķ–Ĺ–ł–Ķ –Ņ—Ź—ā–ĺ–≥–ĺ –ł¬†—ą–Ķ—Ā—ā–ĺ–≥–ĺ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–Ļ –嬆—ā–ĺ–ľ, —á—ā–ĺ –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č (—Ą–į–∑–į –≤–Ĺ—É—ā—Ä–Ķ–Ĺ–Ĺ–Ķ–≥–ĺ —Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ł–∑–į—ā–ĺ—Ä–į —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ—č—Ö —Ä–ł—ā–ľ–ĺ–≤ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į) –Ĺ–į—á–ł–Ĺ–į—é—ā –∑–į–Ņ–į–∑–ī—č–≤–į—ā—Ć –≤¬†–Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ (—á—ā–ĺ —Ā–Ľ–Ķ–ī—É–Ķ—ā –ł–∑¬†–∑–į–Ņ–į–∑–ī—č–≤–į–Ĺ–ł—Ź –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ) –ł¬†–≤ –ī–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–ľ –Ĺ–į—á–ł–Ĺ–į—é—ā —Ā–Ĺ–ĺ–≤–į —Ā–Ņ–Ķ—ą–ł—ā—Ć –Ņ–嬆–ľ–Ķ—Ä–Ķ –≤–∑—Ä–ĺ—Ā–Ľ–Ķ–Ĺ–ł—Ź –ł¬†—Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź
–í¬†—Ü–Ķ–Ľ–ĺ–ľ —Ä–į–∑–Ľ–ł—á–ł–Ķ –ľ–Ķ–∂–ī—É —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į–ľ–ł –ĺ—ā—Ā—É—ā—Ā—ā–≤—É–Ķ—ā, –į¬†–ļ–į–∂—É—Č–Ķ–Ķ—Ā—Ź –ĺ—á–Ķ–≤–ł–ī–Ĺ—č–ľ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł–Ķ –嬆—Ā—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–ľ —Ä–į–∑–Ľ–ł—á–ł–ł –≤–ĺ–∑–Ĺ–ł–ļ–į–Ķ—ā –ł–∑-–∑–į —ā–ĺ–≥–ĺ, —á—ā–ĺ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź, –ļ–į–ļ –Ņ—Ä–į–≤–ł–Ľ–ĺ, –Ņ—Ä–ĺ–≤–ĺ–ī—Ź—ā—Ā—Ź –Ĺ–į¬†–Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–į—Ö. –£¬†–Ĺ–ł—Ö —ć—ā–ĺ —Ź–≤–Ľ–Ķ–Ĺ–ł–Ķ –į—Ā—Ā–ĺ—Ü–ł–ł—Ä—É–Ķ—ā—Ā—Ź —Ā¬†–Ĺ–į–ł–Ī–ĺ–Ľ—Ć—ą–ł–ľ–ł –∂–ł–∑–Ĺ–Ķ–Ĺ–Ĺ—č–ľ–ł —ā—Ä—É–ī–Ĺ–ĺ—Ā—ā—Ź–ľ–ł, –Ņ–ĺ—Ā–ļ–ĺ–Ľ—Ć–ļ—É –≤¬†—Ā—ā–į—Ä—ą–ł—Ö –ļ–Ľ–į—Ā—Ā–į—Ö —Ā–ĺ–Ĺ —Ā–ī–≤–ł–≥–į–Ķ—ā—Ā—Ź –Ĺ–į¬†–Ņ–ĺ–∑–ī–Ĺ–Ķ–Ķ –≤—Ä–Ķ–ľ—Ź, –Ŗ嬆–≤—Ā–Ķ —ą–ļ–ĺ–Ľ—Ć–Ĺ–ł–ļ–ł –≤—č–Ĺ—É–∂–ī–Ķ–Ĺ—č –Ņ—Ä–ł—Ö–ĺ–ī–ł—ā—Ć –≤¬†—ą–ļ–ĺ–Ľ—É –ļ¬†–ĺ–ī–Ĺ–ĺ–ľ—É –ł¬†—ā–ĺ–ľ—É –∂–Ķ —á–į—Ā—É, –Ĺ–Ķ–∑–į–≤–ł—Ā–ł–ľ–ĺ –ĺ—ā¬†–Ņ—Ä–Ķ–ī–Ņ–ĺ—á–ł—ā–į–Ķ–ľ–ĺ–≥–ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į.
–†–ł—Ā—É–Ĺ–ĺ–ļ 3 –ł–Ľ–Ľ—é—Ā—ā—Ä–ł—Ä—É–Ķ—ā –≤–Ľ–ł—Ź–Ĺ–ł–Ķ —Ä–į–Ĺ–Ĺ–Ķ–≥–ĺ –Ĺ–į—á–į–Ľ–į —É—Ä–ĺ–ļ–ĺ–≤ –≤¬†—ą–ļ–ĺ–Ľ–Ķ –Ĺ–į¬†—Ä–į–∑–Ľ–ł—á–ł—Ź –ľ–Ķ–∂–ī—É –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ķ–ľ —Ā–Ĺ–į –≤¬†–Ī—É–ī–Ĺ–ł –ł¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ. –Ě–į¬†–≥—Ä–į–Ĺ–ł—Ü–Ķ 18‚Äď22-–Ľ–Ķ—ā–Ĺ–Ķ–≥–ĺ –≤–ĺ–∑—Ä–į—Ā—ā–į –≤¬†–≥—Ä–į—Ą–ł–ļ–ł —Ā–Ņ–Ķ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ –ī–ĺ–Ī–į–≤–Ľ–Ķ–Ĺ—č –ī–į–Ĺ–Ĺ—č–Ķ (–≤¬†–≤–ł–ī–Ķ –ī–≤—É—Ö –ĺ—ā–ī–Ķ–Ľ—Ć–Ĺ—č—Ö –Ľ–ł–Ĺ–ł–Ļ) –Ľ–ĺ–Ĺ–≥–ł—ā—é–ī–Ĺ–ĺ–≥–ĺ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł—Ź, –≤—č–Ņ–ĺ–Ľ–Ĺ–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ M.¬†Urner –ł¬†—Ā–ĺ–į–≤—ā. (2009) [9], –≤¬†–ļ–ĺ—ā–ĺ—Ä–ĺ–ľ –∑–Ĺ–į—á–Ķ–Ĺ–ł—Ź –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –Ī—č–Ľ–ł –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ—č –ī–Ľ—Ź –ĺ–ī–Ĺ–ĺ–Ļ –ł¬†—ā–ĺ–Ļ –∂–Ķ –≤—č–Ī–ĺ—Ä–ļ–ł —Ā¬†–ł–Ĺ—ā–Ķ—Ä–≤–į–Ľ–ĺ–ľ –Ņ—Ź—ā—Ć –Ľ–Ķ—ā. –°–Ĺ–į—á–į–Ľ–į —É—á–į—Ā—ā–Ĺ–ł–ļ–ł –ĺ–Ņ—Ä–ĺ—Ā–į –Ī—č–Ľ–ł —Ā—ā–į—Ä—ą–Ķ–ļ–Ľ–į—Ā—Ā–Ĺ–ł–ļ–į–ľ–ł, –į¬†–Ņ–ĺ—ā–ĺ–ľ —Ā—ā—É–ī–Ķ–Ĺ—ā–į–ľ–ł —É–Ĺ–ł–≤–Ķ—Ä—Ā–ł—ā–Ķ—ā–į. –≠—ā–ł –ī–į–Ĺ–Ĺ—č–Ķ –Ĺ–į–Ņ–ĺ–ľ–ł–Ĺ–į—é—ā —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č, –Ņ–ĺ–Ľ—É—á–Ķ–Ĺ–Ĺ—č–Ķ –Ņ—Ä–ł —É—Ā—Ä–Ķ–ī–Ĺ–Ķ–Ĺ–ł–ł –≤—Ā–Ķ—Ö –≤—č–Ī–ĺ—Ä–ĺ–ļ –Ņ–嬆–ī–≤—É–ľ —Ā–ĺ—Ā–Ķ–ī—Ā—ā–≤—É—é—Č–ł–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ—č–ľ¬†–≥—Ä—É–Ņ–Ņ–į–ľ. –í–ł–ī–ł–ľ–ĺ, –ĺ–Ī—Č–į—Ź —ā–Ķ–Ĺ–ī–Ķ–Ĺ—Ü–ł—Ź —ā–į–ļ–ĺ–≤–į, —á—ā–ĺ, –ĺ—Ā–≤–ĺ–Ī–ĺ–ī–ł–≤—ą–ł—Ā—Ć –ĺ—ā¬†–ī–ł–ļ—ā–į—ā–į —ą–ļ–ĺ–Ľ—č, –ľ–ĺ–Ľ–ĺ–ī—č–Ķ –Ľ—é–ī–ł –Ĺ–į—á–ł–Ĺ–į–Ľ–ł –≤—Ā—ā–į–≤–į—ā—Ć –≤¬†–Ī—É–ī–Ĺ–ł –≤¬†—Ā—Ä–Ķ–ī–Ĺ–Ķ–ľ –Ĺ–į¬†—á–į—Ā –Ņ–ĺ–∑–∂–Ķ, —á–Ķ–ľ –≤–ĺ –≤—Ä–Ķ–ľ—Ź —É—á–Ķ–Ī—č –≤¬†—Ā—ā–į—Ä—ą–ł—Ö –ļ–Ľ–į—Ā—Ā–į—Ö. –Ě–į¬†—Ä–ł—Ā—É–Ĺ–ļ–Ķ 3 —ć—ā–ĺ—ā —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ, –į¬†–Ĺ–Ķ –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł –ĺ–Ī—É—Ā–Ľ–ĺ–≤–Ľ–Ķ–Ĺ–Ĺ—č–Ļ —Ā–ī–≤–ł–≥ –∑–į–ľ–Ķ—ā–Ĺ–ĺ –ł—Ā–ļ–į–∂–į–Ķ—ā –≤¬†—Ü–Ķ–Ľ–ĺ–ľ –Ņ–Ľ–į–≤–Ĺ–ĺ–Ķ –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ–ĺ–Ķ –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł–Ķ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ, –Ŗ嬆–Ņ–ĺ–ī–ĺ–Ī–Ĺ–į—Ź –į–Ĺ–ĺ–ľ–į–Ľ–ł—Ź –Ĺ–Ķ¬†–Ņ–ĺ–ľ–Ķ—ą–į–Ľ–į –ĺ–Ņ—Ä–Ķ–ī–Ķ–Ľ–ł—ā—Ć –ĺ–Ī—Č–ł–Ķ –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ—č–Ķ —ā–Ķ–Ĺ–ī–Ķ–Ĺ—Ü–ł–ł –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł—Ź –Ņ–į—Ä–į–ľ–Ķ—ā—Ä–ĺ–≤ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–ĺ–≤ —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į —Ā¬†–Ņ–ĺ–ľ–ĺ—Č—Ć—é –ľ–ĺ–ī–Ķ–Ľ–ł (1).
–ö–į–ļ –ł–Ľ–Ľ—é—Ā—ā—Ä–ł—Ä—É–Ķ—ā —Ä–ł—Ā. 1–Ď, —Ā–ł–ľ—É–Ľ—Ź—Ü–ł—Ź –Ī—č–Ľ–į –Ņ—Ä–ĺ–≤–Ķ–ī–Ķ–Ĺ–į —Ā¬†–ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–Ķ–ľ, —á—ā–ĺ –Ĺ–ł¬†–ĺ–ī–ł–Ĺ –ł–∑¬†–Ņ–į—Ä–į–ľ–Ķ—ā—Ä–ĺ–≤ —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–Ļ –ľ–ĺ–ī—É–Ľ—Ź—Ü–ł–ł –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –Ĺ–Ķ¬†–ľ–Ķ–Ĺ—Ź–Ķ—ā—Ā—Ź –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –≤—Ā–Ķ–Ļ –∂–ł–∑–Ĺ–ł. –í¬†—á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–į—Ź —Ą–į–∑–į –ĺ—Ā—ā–į–Ķ—ā—Ā—Ź –ĺ–ī–Ĺ–ĺ–Ļ –ł¬†—ā–ĺ–Ļ –∂–Ķ. –Ď–ĺ–Ľ–Ķ–Ķ —ā–ĺ–≥–ĺ, –Ī—č–Ľ–ĺ —ā–į–ļ–∂–Ķ –Ņ—Ä–ł–Ĺ—Ź—ā–ĺ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł–Ķ, —á—ā–ĺ SWA –ĺ—Ā—ā–į–Ķ—ā—Ā—Ź –Ĺ–Ķ–ł–∑–ľ–Ķ–Ĺ–Ĺ–ĺ–Ļ –≤¬†–Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ (—Ö–ĺ—ā—Ź –Ĺ–į¬†—Ā–į–ľ–ĺ–ľ –ī–Ķ–Ľ–Ķ –ĺ–Ĺ–į —Ā–Ĺ–ł–∂–į–Ķ—ā—Ā—Ź –ł¬†–≤ –ī–Ķ—ā—Ā—ā–≤–Ķ, –ł¬†–≤ –Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ, –ł¬†–Ņ–ĺ –ľ–Ķ—Ä–Ķ –ī–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–≥–ĺ –≤–∑—Ä–ĺ—Ā–Ľ–Ķ–Ĺ–ł—Ź, –į¬†–Ņ–ĺ—ā–ĺ–ľ –ł¬†—Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź). –Ę–į–ļ–ł–Ķ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł—Ź –Ņ–ĺ–∑–≤–ĺ–Ľ–ł–Ľ–ł –Ņ—Ä–ĺ–ī–Ķ–ľ–ĺ–Ĺ—Ā—ā—Ä–ł—Ä–ĺ–≤–į—ā—Ć, —á—ā–ĺ –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ—č–Ķ —Ā–ī–≤–ł–≥–ł –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –Ņ—Ä–ł –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–Ķ –ĺ—ā¬†–ī–Ķ—ā—Ā–ļ–ĺ–≥–ĺ –ļ¬†–≤–∑—Ä–ĺ—Ā–Ľ–ĺ–ľ—É –≤–ĺ–∑—Ä–į—Ā—ā—É –ľ–ĺ–∂–Ĺ–ĺ –ĺ–Ī—ä—Ź—Ā–Ĺ–ł—ā—Ć –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł–Ķ–ľ –≤—Ā–Ķ–≥–ĺ –ĺ–ī–Ĺ–ĺ–≥–ĺ –Ņ–į—Ä–į–ľ–Ķ—ā—Ä–į –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–į —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ¬†‚Äď –Ņ–ĺ—Ā—ā–ĺ—Ź–Ĺ–Ĺ–ĺ–Ļ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł –ī–Ľ—Ź —Ą–į–∑—č –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł—Ź, —ā–ĺ –Ķ—Ā—ā—Ć –ī–Ľ—Ź —Ą–į–∑—č –Ņ–ĺ–ī—ä–Ķ–ľ–į SWA (Tb). –Ē—Ä—É–≥–ł–Ķ –Ņ–ĺ–ī—Ä–ĺ–Ī–Ĺ–ĺ—Ā—ā–ł –ľ–ĺ–ī–Ķ–Ľ–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ—č—Ö —Ä–į–∑–Ľ–ł—á–ł–Ļ –ľ–ĺ–∂–Ĺ–ĺ –Ĺ–į–Ļ—ā–ł –≤¬†—Ä–į–Ī–ĺ—ā–Ķ A. Putilov –ł¬†E. Verevkin [4]. –Ē–Ľ—Ź —Ü–Ķ–Ľ–Ķ–Ļ –ī–į–Ĺ–Ĺ–ĺ–Ļ —Ā—ā–į—ā—Ć–ł –≤–į–∂–Ĺ–ĺ –ĺ—ā–ľ–Ķ—ā–ł—ā—Ć, —á—ā–ĺ –≤–ĺ–∑–ľ–ĺ–∂–Ĺ–ĺ—Ā—ā—Ć —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ—č—Ö –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł–Ļ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –Ī–Ķ–∑ –Ņ–ĺ—Ā—ā—É–Ľ–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź –ļ–į–ļ–ł—Ö-–Ľ–ł–Ī–ĺ –Ņ–į—Ä–į–Ľ–Ľ–Ķ–Ľ—Ć–Ĺ—č—Ö –≤–ĺ–∑—Ä–į—Ā—ā–Ĺ—č—Ö –ł–∑–ľ–Ķ–Ĺ–Ķ–Ĺ–ł–Ļ —Ą–į–∑—č —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–≥–ĺ –≤–Ľ–ł—Ź–Ĺ–ł—Ź –Ĺ–į¬†—Ā–ĺ–Ĺ –Ĺ–į—Ö–ĺ–ī–ł—ā—Ā—Ź –≤¬†–Ņ—Ä–ĺ—ā–ł–≤–ĺ—Ä–Ķ—á–ł–ł —Ā¬†–ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł—Ź–ľ–ł –į–≤—ā–ĺ—Ä–ĺ–≤ –ļ–ĺ–Ĺ—Ü–Ķ–Ņ—Ü–ł–ł —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į –嬆—ā–ĺ–ľ, —á—ā–ĺ –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č (—Ą–į–∑–į –≤–Ĺ—É—ā—Ä–Ķ–Ĺ–Ĺ–Ķ–≥–ĺ —Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ł–∑–į—ā–ĺ—Ä–į —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ—č—Ö —Ä–ł—ā–ľ–ĺ–≤ –ĺ—Ä–≥–į–Ĺ–ł–∑–ľ–į) –Ĺ–į—á–ł–Ĺ–į—é—ā –∑–į–Ņ–į–∑–ī—č–≤–į—ā—Ć –≤¬†–Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ (—á—ā–ĺ —Ā–Ľ–Ķ–ī—É–Ķ—ā –ł–∑¬†–∑–į–Ņ–į–∑–ī—č–≤–į–Ĺ–ł—Ź –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ), –Ŗ嬆–∑–į—ā–Ķ–ľ –Ņ—Ä–ł–Ĺ–ł–ľ–į—é—ā—Ā—Ź —Ā–Ĺ–ĺ–≤–į —Ā–Ņ–Ķ—ą–ł—ā—Ć –Ņ–嬆–ľ–Ķ—Ä–Ķ –ī–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–≥–ĺ –≤–∑—Ä–ĺ—Ā–Ľ–Ķ–Ĺ–ł—Ź –ł¬†—Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź.
–ě–Ņ—Ä–ĺ–≤–Ķ—Ä–∂–Ķ–Ĺ–ł–Ķ —Ā–Ķ–ī—Ć–ľ–ĺ–≥–ĺ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł—Ź –嬆—ā–ĺ–ľ, —á—ā–ĺ –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ī–Ķ–Ľ–ł –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č –ĺ—Ā—ā–į—é—ā—Ā—Ź —Ā–ł–Ĺ—Ą–į–∑–Ĺ—č–ľ–ł 24-—á–į—Ā–ĺ–≤–ĺ–ľ—É —Ä–Ķ–∂–ł–ľ—É –Ķ—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –ĺ—Ā–≤–Ķ—Č–Ķ–Ĺ–ł—Ź
–†–į–∑–Ľ–ł—á–ł—Ź —Ą–į–∑—č —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–Ļ –ľ–ĺ–ī—É–Ľ—Ź—Ü–ł–ł –Ī—č–Ľ–ł –Ņ–ĺ—Ā—ā—É–Ľ–ł—Ä–ĺ–≤–į–Ĺ—č —ā–ĺ–Ľ—Ć–ļ–ĺ –Ņ—Ä–ł —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł —Ä–į–∑–Ľ–ł—á–ł–Ļ –≤–ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –ľ–Ķ–∂–ī—É ¬ę—Ā–ĺ–≤–į–ľ–ł¬Ľ –ł¬†¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–į–ľ–ł¬Ľ (–Ņ–ĺ–ī—Ä–ĺ–Ī–Ĺ–ĺ—Ā—ā–ł –ľ–ĺ–∂–Ĺ–ĺ –Ĺ–į–Ļ—ā–ł –≤¬†—Ä–į–Ī–ĺ—ā–Ķ A. Putilov –ł¬†—Ā–ĺ–į–≤—ā. [5]). –ú–ĺ–ī–Ķ–Ľ–ł—Ä–ĺ–≤–į–Ĺ–ł–Ķ –Ņ—Ä–Ķ–ī—Ā–ļ–į–∑—č–≤–į–Ķ—ā –ĺ—ā—Ā—É—ā—Ā—ā–≤–ł–Ķ –ļ–į–ļ–ł—Ö-–Ľ–ł–Ī–ĺ —Ā—É—Č–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ—č—Ö —Ä–į–∑–Ľ–ł—á–ł–Ļ –Ņ–嬆–ī–Ľ–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ—Ā—ā–ł —Ā–Ĺ–į –ľ–Ķ–∂–ī—É –ī–≤—É–ľ—Ź —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į–ľ–ł. –ö–į–ļ –≤–ł–ī–Ĺ–ĺ –ł–∑¬†—ā–į–Ī–Ľ. 2, —Ä–į—Ā—Ö–ĺ–∂–ī–Ķ–Ĺ–ł–Ķ –ľ–Ķ–∂–ī—É —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł–ľ–ł –ł¬†—Ā–ł–ľ—É–Ľ–ł—Ä–ĺ–≤–į–Ĺ–Ĺ—č–ľ–ł –∑–Ĺ–į—á–Ķ–Ĺ–ł—Ź–ľ–ł –Ī—č–Ľ–ĺ –ľ–ł–Ĺ–ł–ľ–į–Ľ—Ć–Ĺ—č–ľ –ī–Ľ—Ź ¬ę–∂–į–≤–ĺ—Ä–ĺ–Ĺ–ļ–ĺ–≤¬Ľ, —ā–ĺ–≥–ī–į –ļ–į–ļ ¬ę—Ā–ĺ–≤—謼 —É–ļ–į–∑—č–≤–į–Ľ–ł –Ī–ĺ–Ľ–Ķ–Ķ –Ņ–ĺ–∑–ī–Ĺ–Ķ–Ķ –≤—Ä–Ķ–ľ—Ź –Ņ–ĺ–ī—ä–Ķ–ľ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ, —á–Ķ–ľ –≤—Ä–Ķ–ľ—Ź, –Ņ—Ä–Ķ–ī—Ā–ļ–į–∑–į–Ĺ–Ĺ–ĺ–Ķ –ľ–ĺ–ī–Ķ–Ľ—Ć—é. –ü—Ä–ł —ć—ā–ĺ–ľ —Ä–į—Ā—Ö–ĺ–∂–ī–Ķ–Ĺ–ł–Ķ –Ī—č–Ľ–ĺ –Ī–ĺ–Ľ–Ķ–Ķ –≤—č—Ä–į–∂–Ķ–Ĺ–Ĺ—č–ľ —ɬ†–Ľ–ł—Ü –ľ–ĺ–Ľ–ĺ–∂–Ķ 33¬†–Ľ–Ķ—ā. –ö—Ä–ĺ–ľ–Ķ —ā–ĺ–≥–ĺ, –ļ–į–ļ –≤–ł–ī–Ĺ–ĺ –Ĺ–į¬†—Ä–ł—Ā. 3, –Ī–ĺ–Ľ–Ķ–Ķ –Ņ–ĺ–∑–ī–Ĺ–Ķ–Ķ –≤—Ä–Ķ–ľ—Ź –Ņ–ĺ–ī—ä–Ķ–ľ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –Ņ–嬆—Ā—Ä–į–≤–Ĺ–Ķ–Ĺ–ł—é —Ā¬†—ā–Ķ–ľ, —á—ā–ĺ –Ņ—Ä–Ķ–ī—Ā–ļ–į–∑–į–Ľ–į –ľ–ĺ–ī–Ķ–Ľ—Ć, –Ĺ–į–Ī–Ľ—é–ī–į–Ľ–ĺ—Ā—Ć –ł¬†–≤ —Ā—ā–į—Ä—ą–Ķ–ľ –Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤–ĺ–ľ –≤–ĺ–∑—Ä–į—Ā—ā–Ķ. –ě–ī–Ĺ–ĺ –ł–∑¬†–≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č—Ö –ĺ–Ī—ä—Ź—Ā–Ĺ–Ķ–Ĺ–ł–Ļ –ĺ–Ī–Ĺ–į—Ä—É–∂–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ —Ä–į—Ā—Ö–ĺ–∂–ī–Ķ–Ĺ–ł—Ź (–嬆–ī—Ä—É–≥–ł—Ö —Ä–į—Ā—Ö–ĺ–∂–ī–Ķ–Ĺ–ł—Ź—Ö –ľ–ĺ–∂–Ĺ–ĺ –Ņ—Ä–ĺ—á–Ķ—Ā—ā—Ć –≤¬†—Ä–į–Ī–ĺ—ā–į—Ö A. Putilov –ł¬†E.¬†Verevkin [4], A. Putilov [5])¬†‚Äď —Ā–ī–≤–ł–≥ —Ą–į–∑—č —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–≥–ĺ —Ä–ł—ā–ľ–į –Ņ–ĺ–ī –≤–Ľ–ł—Ź–Ĺ–ł–Ķ–ľ –ł—Ā–ļ—É—Ā—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –≤–Ķ—á–Ķ—Ä–Ĺ–Ķ–≥–ĺ –ĺ—Ā–≤–Ķ—Č–Ķ–Ĺ–ł—Ź —ɬ†–Ľ—é–ī–Ķ–Ļ —Ā¬†–Ņ–ĺ–∑–ī–Ĺ–ł–ľ –ĺ—ā—Ö–ĺ–ī–ĺ–ľ –ļ–嬆—Ā–Ĺ—É, —á—ā–ĺ –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź–Ķ—ā —ć—ā–ł–ľ –Ľ—é–ī—Ź–ľ —Ā–Ņ–į—ā—Ć –ī–ĺ–Ľ—Ć—ą–Ķ –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł (—Ā–≤–Ķ—ā –≤—č–∑—č–≤–į–Ķ—ā –∑–į–Ņ–į–∑–ī—č–≤–į–Ĺ–ł–Ķ —Ą–į–∑—č, –Ĺ–Ķ¬†—É—á—ā–Ķ–Ĺ–Ĺ–ĺ–Ķ –≤¬†—Ā–ł–ľ—É–Ľ—Ź—Ü–ł–ł). –ó–į—ā–Ķ–ľ –≤¬†—Ä–į–Ī–ĺ—á–ł–Ķ/—É—á–Ķ–Ī–Ĺ—č–Ķ –ī–Ĺ–ł —Ą–į–∑–į –ľ–ĺ–∂–Ķ—ā —Ā–ī–≤–ł–≥–į—ā—Ć—Ā—Ź –Ĺ–į¬†—ā—É –∂–Ķ –≤–Ķ–Ľ–ł—á–ł–Ĺ—É –≤–Ņ–Ķ—Ä–Ķ–ī –ł–∑-–∑–į –≤–Ľ–ł—Ź–Ĺ–ł—Ź —É—ā—Ä–Ķ–Ĺ–Ĺ–Ķ–≥–ĺ –ĺ—Ā–≤–Ķ—Č–Ķ–Ĺ–ł—Ź, —á—ā–ĺ –≤–Ņ–ĺ–Ľ–Ĺ–Ķ –≤–Ķ—Ä–ĺ—Ź—ā–Ĺ–ĺ –ł–∑-–∑–į —Ä–į–Ĺ–Ĺ–Ķ–≥–ĺ –Ņ–ĺ–ī—ä–Ķ–ľ–į. –ü–ĺ—ć—ā–ĺ–ľ—É –ī–Ľ—Ź –Ľ—é–ī–Ķ–Ļ —Ā¬†–Ņ–ĺ–∑–ī–Ĺ–ł–ľ –ĺ—ā—Ö–ĺ–ī–ĺ–ľ –ļ–嬆—Ā–Ĺ—É –ľ–ĺ–∂–Ķ—ā –Ī—č—ā—Ć –Ĺ–Ķ–≤–Ķ—Ä–Ĺ—č–ľ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł–Ķ –嬆—ā–ĺ–ľ, —á—ā–ĺ –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ī–Ķ–Ľ–ł –Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł–Ķ —á–į—Ā—č –ĺ—Ā—ā–į—é—ā—Ā—Ź —Ā–ł–Ĺ—Ą–į–∑–Ĺ—č–ľ–ł 24-—á–į—Ā–ĺ–≤–ĺ–ľ—É —Ä–Ķ–∂–ł–ľ—É –Ķ—Ā—ā–Ķ—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ–≥–ĺ –ĺ—Ā–≤–Ķ—Č–Ķ–Ĺ–ł—Ź. –ė¬†—ć—ā–ł –Ľ—é–ī–ł –ľ–ĺ–≥—É—ā –Ķ–∂–Ķ–Ĺ–Ķ–ī–Ķ–Ľ—Ć–Ĺ–ĺ –ł—Ā–Ņ—č—ā—č–≤–į—ā—Ć –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑ –≤¬†–Ķ–≥–ĺ —Ā–į–ľ–ĺ–Ļ –ĺ–Ī—č—á–Ĺ–ĺ–Ļ —Ą–ĺ—Ä–ľ–Ķ.
–ě–Ņ—Ä–ĺ–≤–Ķ—Ä–∂–Ķ–Ĺ–ł–Ķ –≤–ĺ—Ā—Ć–ľ–ĺ–≥–ĺ –ī–ĺ–Ņ—É—Č–Ķ–Ĺ–ł—Ź –嬆—ā–ĺ–ľ, —á—ā–ĺ —ā–ĺ–Ľ—Ć–ļ–ĺ —Ą–į–∑–į —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ —Ā–ī–≤–ł–≥–į–Ķ—ā—Ā—Ź –≤–Ņ–Ķ—Ä–Ķ–ī-–Ĺ–į–∑–į–ī –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ī–Ķ–Ľ–ł
–ö–į–∑–į–Ľ–ĺ—Ā—Ć –Ī—č, –Ņ—Ä–Ķ–ī–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–ł–Ķ –嬆—ā–ĺ–ľ, —á—ā–ĺ —ā–ĺ–Ľ—Ć–ļ–ĺ —Ą–į–∑–į —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ —Ā–ī–≤–ł–≥–į–Ķ—ā—Ā—Ź –≤–Ņ–Ķ—Ä–Ķ–ī-–Ĺ–į–∑–į–ī –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ī–Ķ–Ľ–ł, —ā—Ä—É–ī–Ĺ–ĺ –ĺ–Ņ—Ä–ĺ–≤–Ķ—Ä–≥–Ĺ—É—ā—Ć. –ě–ī–Ĺ–į–ļ–ĺ –ĺ–Ĺ–ĺ –ĺ—Ā–Ĺ–ĺ–≤–į–Ĺ–ĺ –ł—Ā–ļ–Ľ—é—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –Ĺ–į¬†—Ą–į–ļ—ā–Ķ, —á—ā–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į —Ā–ī–≤–ł–≥–į–Ķ—ā—Ā—Ź —Ā–Ĺ–į—á–į–Ľ–į –Ĺ–į¬†—Ä–į–Ĺ–Ĺ–ł–Ķ —á–į—Ā—č –≤¬†–Ī—É–ī–Ĺ–ł, –į¬†–Ņ–ĺ—ā–ĺ–ľ –Ĺ–į¬†–Ņ–ĺ–∑–ī–Ĺ–ł–Ķ —á–į—Ā—č –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ. –ú–ĺ–ī–Ķ–Ľ—Ć –ĺ—ā—Ä–į–∂–į–Ķ—ā –Ĺ–Ķ¬†—ā–ĺ–Ľ—Ć–ļ–ĺ –≤—Ä–Ķ–ľ—Ź —Ā–Ĺ–į, –Ŗ嬆–ł –ī–ł–Ĺ–į–ľ–ł–ļ—É —Ä–ł—ā–ľ–ł—á–Ķ—Ā–ļ–ĺ–≥–ĺ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–į, –Ľ–Ķ–∂–į—Č–Ķ–≥–ĺ –≤¬†–ĺ—Ā–Ĺ–ĺ–≤–Ķ —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ. –ē–≥–ĺ –ł–Ĺ–ī–ł–ļ–į—ā–ĺ—Ä–ĺ–ľ —Ā—á–ł—ā–į—é—ā—Ā—Ź –ļ–ĺ–Ľ–Ķ–Ī–į–Ĺ–ł—Ź SWA –≤¬†—ć—ā–ĺ–ľ —Ü–ł–ļ–Ľ–Ķ¬†‚Äď –ĺ–Ī—Ä–į—ā–Ĺ–ĺ-—ć–ļ—Ā–Ņ–ĺ–Ĺ–Ķ–Ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–Ļ —Ä–ĺ—Ā—ā –ł¬†—ć–ļ—Ā–Ņ–ĺ–Ĺ–Ķ–Ĺ—Ü–ł–į–Ľ—Ć–Ĺ—č–Ļ —Ā–Ņ–į–ī (—Ā–ľ.¬†—Ä–ł—Ā.¬†1). –Ě–į¬†—Ä–ł—Ā—É–Ĺ–ļ–Ķ 1–í –≤–ł–ī–Ĺ–ĺ, —á—ā–ĺ –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –≤—Ā–Ķ–Ļ –Ĺ–Ķ–ī–Ķ–Ľ–ł –ľ–Ķ–Ĺ—Ź–Ķ—ā—Ā—Ź —ā–ĺ–Ľ—Ć–ļ–ĺ —Ą–ĺ—Ä–ľ–į –≤–ĺ–Ľ–Ĺ—č —Ü–ł–ļ–Ľ–į –ļ–ĺ–Ľ–Ķ–Ī–į–Ĺ–ł–Ļ SWA. –ē—Ā–Ľ–ł –≤¬†–ļ–į—á–Ķ—Ā—ā–≤–Ķ –Ķ–≥–ĺ —Ą–į–∑—č –≤—č–Ī—Ä–į—ā—Ć —É—Ä–ĺ–≤–Ķ–Ĺ—Ć SWA –Ĺ–į–ļ–į–Ĺ—É–Ĺ–Ķ –Ņ–ĺ–ī—ä–Ķ–ľ–į –≤¬†–Ī—É–ī–Ĺ–ł, —ā–ĺ –Ķ—Ā—ā—Ć —Ā–Ķ–ľ—Ć —á–į—Ā–ĺ–≤ —É—ā—Ä–į (—Ą–į–∑–į¬†‚Äď —ć—ā–ĺ –Ľ—é–Ī–į—Ź –Ņ—Ä–ĺ–ł–∑–≤–ĺ–Ľ—Ć–Ĺ–ĺ –≤—č–Ī—Ä–į–Ĺ–Ĺ–į—Ź —ā–ĺ—á–ļ–į —Ü–ł–ļ–Ľ–į), —ā–ĺ —ć—ā–į —É—ā—Ä–Ķ–Ĺ–Ĺ—Ź—Ź —Ą–į–∑–į –ĺ–ļ–į–∑—č–≤–į–Ķ—ā—Ā—Ź –Ņ—Ä–į–ļ—ā–ł—á–Ķ—Ā–ļ–ł –ĺ–ī–ł–Ĺ–į–ļ–ĺ–≤–ĺ–Ļ –Ņ–嬆—É—Ä–ĺ–≤–Ĺ—é SWA –ł¬†–≤ –Ľ—é–Ī–ĺ–ľ –ł–∑¬†—Ü–ł–ļ–Ľ–ĺ–≤. –ė–Ĺ—č–ľ–ł —Ā–Ľ–ĺ–≤–į–ľ–ł, —ɬ†–Ņ–ĺ–ī–į–≤–Ľ—Ź—é—Č–Ķ–≥–ĺ –Ī–ĺ–Ľ—Ć—ą–ł–Ĺ—Ā—ā–≤–į –Ľ—é–ī–Ķ–Ļ –ĺ–Ĺ–į –Ĺ–Ķ¬†—Ā–ī–≤–ł–≥–į–Ķ—ā—Ā—Ź –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ –Ĺ–Ķ–ī–Ķ–Ľ–ł.
–ě–Ī—Ā—É–∂–ī–Ķ–Ĺ–ł–Ķ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–ĺ–≤
–°–ł–ľ—É–Ľ–ł—Ä–ĺ–≤–į–Ĺ–ł–Ķ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ł¬†–Ņ–ĺ –Ī—É–ī–Ĺ—Ź–ľ —Ā¬†–Ņ–ĺ–ľ–ĺ—Č—Ć—é –ľ–ĺ–ī–Ķ–Ľ–ł —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ –Ņ–ĺ–∑–≤–ĺ–Ľ—Ź–Ķ—ā —É—Ā–ĺ–ľ–Ĺ–ł—ā—Ć—Ā—Ź –≤¬†–ī–ĺ—Ā—ā–ĺ–≤–Ķ—Ä–Ĺ–ĺ—Ā—ā–ł –Ľ—é–Ī–ĺ–≥–ĺ –ł–∑¬†–Ī—č—ā—É—é—Č–ł—Ö –≤¬†–Ĺ–į—É—á–Ĺ—č—Ö –ļ—Ä—É–≥–į—Ö —Ā¬†2006¬†–≥.¬†–Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł–Ļ –嬆—Ź–≤–Ľ–Ķ–Ĺ–ł–ł —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į. –ě–Ĺ–ł –Ľ–ł—ą—Ć –Ĺ–į¬†–Ņ–Ķ—Ä–≤—č–Ļ –≤–∑–≥–Ľ—Ź–ī –ļ–į–∂—É—ā—Ā—Ź –ĺ—á–Ķ–≤–ł–ī–Ĺ—č–ľ–ł, –Ŗ嬆–ł—Ö –Ĺ–Ķ–Ľ—Ć–∑—Ź –ĺ–Ī—ä—Ź—Ā–Ĺ–ł—ā—Ć —Ā¬†–Ņ–ĺ–∑–ł—Ü–ł–Ļ —Ö—Ä–ĺ–Ĺ–ĺ–Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ĺ–Ļ —ā–Ķ–ĺ—Ä–ł–ł, –Ī–į–∑–ł—Ä—É—é—Č–Ķ–Ļ—Ā—Ź –Ĺ–į¬†–ľ–į—ā–Ķ–ľ–į—ā–ł—á–Ķ—Ā–ļ–ĺ–ľ –∑–Ĺ–į–Ĺ–ł–ł. –ü–ĺ—Ā–ļ–ĺ–Ľ—Ć–ļ—É –ī–Ľ—Ź —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–Ļ –Ī—č–Ľ–ł –ī–ĺ—Ā—ā—É–Ņ–Ĺ—č —ā–ĺ–Ľ—Ć–ļ–ĺ –ī–į–Ĺ–Ĺ—č–Ķ –嬆–≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į, –į¬†–ī–į–Ĺ–Ĺ—č–Ķ –ĺ –ľ–į—Ä–ļ–Ķ—Ä–į—Ö —Ą–į–∑—č —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–≥–ĺ —Ä–ł—ā–ľ–į –ī–Ľ—Ź –Ņ–ĺ–ī–į–≤–Ľ—Ź—é—Č–Ķ–≥–ĺ –Ī–ĺ–Ľ—Ć—ą–ł–Ĺ—Ā—ā–≤–į –≤—č–Ī–ĺ—Ä–ĺ–ļ –ĺ—ā—Ā—É—ā—Ā—ā–≤–ĺ–≤–į–Ľ–ł, –Ĺ–Ķ–ĺ–Ī—Ö–ĺ–ī–ł–ľ–ĺ –Ī—č–Ľ–ĺ –Ņ–ĺ–ī–ī–Ķ—Ä–∂–į—ā—Ć —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –ľ–ĺ–ī–Ķ–Ľ–ł—Ä–ĺ–≤–į–Ĺ–ł—Ź —ā–Ķ–ľ–ł –Ĺ–Ķ–ľ–Ĺ–ĺ–≥–ł–ľ–ł –Ľ–ł—ā–Ķ—Ä–į—ā—É—Ä–Ĺ—č–ľ–ł –ł—Ā—ā–ĺ—á–Ĺ–ł–ļ–į–ľ–ł, –≤¬†–ļ–ĺ—ā–ĺ—Ä—č—Ö –Ĺ–į—Ä—Ź–ī—É —Ā–嬆–≤—Ä–Ķ–ľ–Ķ–Ĺ–Ķ–ľ —Ā–Ĺ–į –Ņ—Ä–ł–≤–Ķ–ī–Ķ–Ĺ—č —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā—č –ł–∑–ľ–Ķ—Ä–Ķ–Ĺ–ł—Ź —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–Ļ —Ą–į–∑—č –Ĺ–į¬†–Ņ—Ä–ł–ľ–Ķ—Ä–Ķ —Ä–ł—ā–ľ–ĺ–≤ —ā–Ķ–ľ–Ņ–Ķ—Ä–į—ā—É—Ä—č —ā–Ķ–Ľ–į –ł–Ľ–ł –ľ–Ķ–Ľ–į—ā–ĺ–Ĺ–ł–Ĺ–į.
–Ě–į–Ņ—Ä–ł–ľ–Ķ—Ä, –≤¬†–Ľ–ĺ–Ĺ–≥–ł—ā—é–ī–Ĺ–ĺ–ľ –ł—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–ł S. Crowley –ł¬†—Ā–ĺ–į–≤—ā. (2014) —Ą–į–∑–į –Ĺ–į—á–į–Ľ–į —Ā–ł–Ĺ—ā–Ķ–∑–į –ľ–Ķ–Ľ–į—ā–ĺ–Ĺ–ł–Ĺ–į –ĺ—Ā—ā–į–≤–į–Ľ–į—Ā—Ć –Ĺ–Ķ–ł–∑–ľ–Ķ–Ĺ–Ĺ–ĺ–Ļ –Ņ—Ä–ł –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–Ķ –ĺ—ā¬†–ľ–Ľ–į–ī—ą–Ķ–≥–ĺ –ļ¬†—Ā—ā–į—Ä—ą–Ķ–ľ—É –Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤–ĺ–ľ—É –≤–ĺ–∑—Ä–į—Ā—ā—É, —Ö–ĺ—ā—Ź –Ņ—Ä–ł —ć—ā–ĺ–ľ –Ĺ–į–Ī–Ľ—é–ī–į–Ľ–į—Ā—Ć —ā–ł–Ņ–ł—á–Ĺ–į—Ź –ī–Ľ—Ź —ā–į–ļ–ĺ–≥–ĺ –Ņ–Ķ—Ä–Ķ—Ö–ĺ–ī–į –ļ–į—Ä—ā–ł–Ĺ–į –∑–į–Ņ–į–∑–ī—č–≤–į—é—Č–Ķ–≥–ĺ —Ā–ī–≤–ł–≥–į –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ [10]. –ö¬†—ć—ā–ĺ–ľ—É –ľ–ĺ–∂–Ĺ–ĺ –ī–ĺ–Ī–į–≤–ł—ā—Ć –ī–≤–į —É—ā–ĺ—á–Ĺ–Ķ–Ĺ–ł—Ź. –í–ĺ-–Ņ–Ķ—Ä–≤—č—Ö, –ĺ–Ī—Ä–į—ā–Ĺ—č–Ļ (–ĺ–Ņ–Ķ—Ä–Ķ–∂–į—é—Č–ł–Ļ) —Ā–ī–≤–ł–≥ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į –Ņ–嬆–ľ–Ķ—Ä–Ķ –ī–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–≥–ĺ –≤–∑—Ä–ĺ—Ā–Ľ–Ķ–Ĺ–ł—Ź –ł¬†—Ā—ā–į—Ä–Ķ–Ĺ–ł—Ź –Ĺ–į—á–ł–Ĺ–į–Ķ—ā—Ā—Ź —Ä–į–Ĺ—Ć—ą–Ķ, —á–Ķ–ľ –ĺ–Ī–Ĺ–į—Ä—É–∂–ł–≤–į—é—ā—Ā—Ź –ļ–į–ļ–ł–Ķ-–Ľ–ł–Ī–ĺ –Ņ—Ä–ł–∑–Ĺ–į–ļ–ł –ĺ–Ņ–Ķ—Ä–Ķ–∂–į—é—Č–Ķ–≥–ĺ —Ā–ī–≤–ł–≥–į —Ą–į–∑—č —Ä–ł—ā–ľ–ĺ–≤-–ľ–į—Ä–ļ–Ķ—Ä–ĺ–≤ —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–≥–ĺ —Ä–ł—ā–ľ–į, –į¬†–ĺ–Ņ–Ķ—Ä–Ķ–∂–į—é—Č–ł–Ļ —Ā–ī–≤–ł–≥ —ć—ā–ĺ–Ļ —Ą–į–∑—č, –Ĺ–į–Ī–Ľ—é–ī–į–Ķ–ľ—č–Ļ –≤¬†–≥–Ľ—É–Ī–ĺ–ļ–ĺ–Ļ —Ā—ā–į—Ä–ĺ—Ā—ā–ł, –ľ–ĺ–∂–Ķ—ā –Ī—č—ā—Ć –ĺ–Ī—ä—Ź—Ā–Ĺ–Ķ–Ĺ –ī—Ä—É–≥–ł–ľ–ł –Ņ—Ä–ł—á–ł–Ĺ–į–ľ–ł (–Ī–ĺ–Ľ–Ķ–Ķ –Ņ–ĺ–ī—Ä–ĺ–Ī–Ĺ–ĺ —ć—ā–ĺ —Ä–į—Ā—Ā–ľ–ĺ—ā—Ä–Ķ–Ĺ–ĺ –≤¬†—Ä–į–Ī–ĺ—ā–Ķ A. Putilov 2016¬†–≥.¬†[11]). –í–ĺ-–≤—ā–ĺ—Ä—č—Ö, —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ—č–Ļ –Ņ–Ķ—Ä–ł–ĺ–ī –Ĺ–Ķ¬†–ľ–Ķ–Ĺ—Ź–Ķ—ā—Ā—Ź —Ā¬†–≤–ĺ–∑—Ä–į—Ā—ā–ĺ–ľ, —á—ā–ĺ –Ī—č–Ľ–ĺ –ĺ–Ī–Ĺ–į—Ä—É–∂–Ķ–Ĺ–ĺ –ļ–į–ļ –Ņ—Ä–ł —Ā—Ä–į–≤–Ĺ–Ķ–Ĺ–ł–ł –Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤ —Ā¬†–≤–∑—Ä–ĺ—Ā–Ľ—č–ľ–ł, —ā–į–ļ –ł¬†–Ņ—Ä–ł —Ā—Ä–į–≤–Ĺ–Ķ–Ĺ–ł–ł –≤–∑—Ä–ĺ—Ā–Ľ—č—Ö —Ā¬†–Ņ–ĺ–∂–ł–Ľ—č–ľ–ł –Ľ—é–ī—Ć–ľ–ł [4, 5, 11].
–ö¬†–Ī–ĺ–Ľ—Ć—ą–ĺ–ľ—É —É–ī–ł–≤–Ľ–Ķ–Ĺ–ł—é, –Ņ—É–Ī–Ľ–ł–ļ–į—Ü–ł–ł, –≤¬†–ļ–ĺ—ā–ĺ—Ä—č—Ö —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–į—Ź —Ą–į–∑–į –ł–∑–ľ–Ķ—Ä—Ź–Ľ–į—Ā—Ć –Ī—č —Ā–Ĺ–į—á–į–Ľ–į –≤¬†–Ī—É–ī–Ĺ–ł, –į¬†–∑–į—ā–Ķ–ľ –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ, –Ņ—Ä–į–ļ—ā–ł—á–Ķ—Ā–ļ–ł –ĺ—ā—Ā—É—ā—Ā—ā–≤—É—é—ā. –ü–嬆–ļ—Ä–į–Ļ–Ĺ–Ķ–Ļ –ľ–Ķ—Ä–Ķ, –Ķ–ī–ł–Ĺ–ĺ–∂–ī—č —ć—ā–ĺ –Ī—č–Ľ–ĺ —Ā–ī–Ķ–Ľ–į–Ĺ–ĺ –≤¬†–ī–ł—Ā—Ā–Ķ—Ä—ā–į—Ü–ł–ĺ–Ĺ–Ĺ–ĺ–Ļ —Ä–į–Ī–ĺ—ā–Ķ G. Zerbini (2017), –≤¬†—á–ł—Ā–Ľ–Ķ —Ä—É–ļ–ĺ–≤–ĺ–ī–ł—ā–Ķ–Ľ–Ķ–Ļ –ļ–ĺ—ā–ĺ—Ä–ĺ–Ļ –ĺ–ļ–į–∑–į–Ľ–ł—Ā—Ć –į–≤—ā–ĺ—Ä—č —Ā—ā–į—ā—Ć–ł –Ņ–嬆—Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–ľ—É –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑—É [12]. –ź–≤—ā–ĺ—Ä –ĺ–Ī–Ĺ–į—Ä—É–∂–ł–Ľ —Ā–ī–≤–ł–≥ —Ą–į–∑—č –Ĺ–į—á–į–Ľ–į —Ā–Ķ–ļ—Ä–Ķ—Ü–ł–ł –ľ–Ķ–Ľ–į—ā–ĺ–Ĺ–ł–Ĺ–į —ɬ†–Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤ —Ā¬†–Ņ–ĺ–∑–ī–Ĺ–ł–ľ —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–ĺ–ľ –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł, –Ņ—Ä–ł—á–Ķ–ľ –ł¬†–∑–ł–ľ–ĺ–Ļ, –ł¬†–Ľ–Ķ—ā–ĺ–ľ, —á–Ķ–≥–ĺ –Ĺ–Ķ¬†–Ĺ–į–Ī–Ľ—é–ī–į–Ľ–ĺ—Ā—Ć —ɬ†–Ņ–ĺ–ī—Ä–ĺ—Ā—ā–ļ–ĺ–≤ —Ā¬†—Ä–į–Ĺ–Ĺ–ł–ľ –ł¬†–Ņ—Ä–ĺ–ľ–Ķ–∂—É—ā–ĺ—á–Ĺ—č–ľ —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–ĺ–ľ. –ü–ĺ—ć—ā–ĺ–ľ—É –Ņ—Ä–ł–≤–Ķ–ī–Ķ–Ĺ–Ĺ–ĺ–Ķ –≤—č—ą–Ķ –ĺ–Ī—ä—Ź—Ā–Ĺ–Ķ–Ĺ–ł–Ķ —Ä–į–∑–Ľ–ł—á–ł–Ļ –ľ–Ķ–∂–ī—É —ć–ľ–Ņ–ł—Ä–ł—á–Ķ—Ā–ļ–ł–ľ –ł¬†–Ņ—Ä–Ķ–ī—Ā–ļ–į–∑–į–Ĺ–Ĺ—č–ľ –≤—Ä–Ķ–ľ–Ķ–Ĺ–Ķ–ľ –Ņ–ĺ–ī—ä–Ķ–ľ–į –≤¬†–≤—č—Ö–ĺ–ī–Ĺ—č–Ķ –ī–Ĺ–ł —ɬ†–Ľ—é–ī–Ķ–Ļ —Ā¬†–Ņ–ĺ–∑–ī–Ĺ–ł–ľ —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–ĺ–ľ –ľ–ĺ–∂–Ķ—ā –Ī—č—ā—Ć –≤–Ņ–ĺ–Ľ–Ĺ–Ķ –Ņ—Ä–į–≤–ī–ĺ–Ņ–ĺ–ī–ĺ–Ī–Ĺ—č–ľ.
–í—č–≤–ĺ–ī—č
–ė–∑ —Ā–ļ–į–∑–į–Ĺ–Ĺ–ĺ–≥–ĺ —Ā–Ľ–Ķ–ī—É–Ķ—ā, —á—ā–ĺ –Ņ—Ä–Ķ–ī—Ā—ā–į–≤–Ľ–Ķ–Ĺ–ł—Ź –嬆—Ö—Ä–ĺ–Ĺ–ĺ–Ī–ł–ĺ–Ľ–ĺ–≥–ł—á–Ķ—Ā–ļ–ł—Ö –ľ–Ķ—Ö–į–Ĺ–ł–∑–ľ–į—Ö, –Ľ–Ķ–∂–į—Č–ł—Ö –≤¬†–ĺ—Ā–Ĺ–ĺ–≤–Ķ —Ź–≤–Ľ–Ķ–Ĺ–ł—Ź —Ā–ĺ—Ü–ł–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑–į, –Ĺ—É–∂–ī–į—é—ā—Ā—Ź –≤¬†–ĺ—Ā–Ĺ–ĺ–≤–į—ā–Ķ–Ľ—Ć–Ĺ–ĺ–ľ –Ņ–Ķ—Ä–Ķ—Ā–ľ–ĺ—ā—Ä–Ķ. –ü–嬆—Ā—É—ā–ł, –ī–Ľ—Ź –Ī–ĺ–Ľ—Ć—ą–ł–Ĺ—Ā—ā–≤–į –Ľ—é–ī–Ķ–Ļ —ć—ā–ĺ –Ĺ–Ķ¬†–ī–Ķ—Ā–ł–Ĺ—Ö—Ä–ĺ–Ĺ–ĺ–∑, –į¬†–Ĺ–Ķ –Ī–ĺ–Ľ–Ķ–Ķ —á–Ķ–ľ –Ī–Ķ–∑–≤–ĺ–∑–≤—Ä–į—ā–Ĺ–ĺ–Ķ —É–ļ–ĺ—Ä–ĺ—á–Ķ–Ĺ–ł–Ķ –Ņ—Ä–ł–≤—č—á–Ĺ–ĺ–≥–ĺ –≤—Ä–Ķ–ľ–Ķ–Ĺ–ł —Ā–Ĺ–į. –ö–į–ļ–ł—Ö-–Ľ–ł–Ī–ĺ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł–Ļ —ā–į–ļ–ĺ–≥–ĺ —É–ļ–ĺ—Ä–ĺ—á–Ķ–Ĺ–ł—Ź –ī–Ľ—Ź —Ü–ł—Ä–ļ–į–ī–ł–į–Ĺ–Ĺ–ĺ–≥–ĺ –ł¬†–≥–ĺ–ľ–Ķ–ĺ—Ā—ā–į—ā–ł—á–Ķ—Ā–ļ–ĺ–≥–ĺ –ļ–ĺ–ľ–Ņ–ĺ–Ĺ–Ķ–Ĺ—ā–ĺ–≤ –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ā–į —Ä–Ķ–≥—É–Ľ—Ź—Ü–ł–ł —Ü–ł–ļ–Ľ–į ¬ę—Ā–ĺ–Ŭ†‚Äď –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–Ķ¬Ľ –≤–ĺ–∑–Ĺ–ł–ļ–Ĺ—É—ā—Ć –Ņ—Ä–ĺ—Ā—ā–ĺ –Ĺ–Ķ¬†–ľ–ĺ–∂–Ķ—ā (–≤¬†—á–į—Ā—ā–Ĺ–ĺ—Ā—ā–ł, –≤¬†—Ą–ĺ—Ä–ľ–Ķ –Ĺ–į–ļ–ĺ–Ņ–Ľ–Ķ–Ĺ–ł—Ź –≤¬†—ā–Ķ—á–Ķ–Ĺ–ł–Ķ —Ä–į–Ī–ĺ—á–Ķ–Ļ/—É—á–Ķ–Ī–Ĺ–ĺ–Ļ –Ĺ–Ķ–ī–Ķ–Ľ–ł –ī–ĺ–Ľ–≥–į —Ā–Ĺ–į —Ā¬†–Ķ–≥–ĺ –ī–į–Ľ—Ć–Ĺ–Ķ–Ļ—ą–Ķ–Ļ, –Ņ—É—Ā—ā—Ć –ī–į–∂–Ķ —á–į—Ā—ā–ł—á–Ĺ–ĺ–Ļ, –ļ–ĺ–ľ–Ņ–Ķ–Ĺ—Ā–į—Ü–ł–Ķ–Ļ –≤ –≤—č—Ö–ĺ–ī–Ĺ—č–Ķ).
–ė—Ā—Ā–Ľ–Ķ–ī–ĺ–≤–į–Ĺ–ł–Ķ –≤—č–Ņ–ĺ–Ľ–Ĺ–Ķ–Ĺ–ĺ –Ņ—Ä–ł¬†—Ą–ł–Ĺ–į–Ĺ—Ā–ĺ–≤–ĺ–Ļ –Ņ–ĺ–ī–ī–Ķ—Ä–∂–ļ–Ķ –†–§–§–ė (¬ę–ě—Ü–Ķ–Ĺ–ļ–į —Ä–į–∑–Ľ–ł—á–ł–Ļ –ľ–Ķ–∂–ī—É —á–Ķ—ā—č—Ä—Ć–ľ—Ź –ļ—Ä–į–Ļ–Ĺ–ł–ľ–ł —Ö—Ä–ĺ–Ĺ–ĺ—ā–ł–Ņ–į–ľ–ł –≤¬†–Ņ–į—Ä–į–ľ–Ķ—ā—Ä–į—Ö –ļ–ĺ–Ľ–Ķ–Ī–į–Ĺ–ł–Ļ –ĺ–Ī—ä–Ķ–ļ—ā–ł–≤–Ĺ—č—Ö –Ņ–ĺ–ļ–į–∑–į—ā–Ķ–Ľ–Ķ–Ļ —Ā–ĺ–Ĺ–Ľ–ł–≤–ĺ—Ā—ā–ł –ł¬†—Ä–į–Ī–ĺ—ā–ĺ—Ā–Ņ–ĺ—Ā–ĺ–Ī–Ĺ–ĺ—Ā—ā–ł –Ņ—Ä–ł –Ņ—Ä–ĺ–Ľ–ĺ–Ĺ–≥–ł—Ä–ĺ–≤–į–Ĺ–Ĺ–ĺ–ľ –Ī–ĺ–ī—Ä—Ā—ā–≤–ĺ–≤–į–Ĺ–ł–ł¬Ľ, ‚ĄĖ¬†19-013-00424-–į).
–ź–≤—ā–ĺ—Ä –Ņ—Ä–ł–∑–Ĺ–į—ā–Ķ–Ľ–Ķ–Ĺ –ļ.–Ī.–Ĺ. –ē–≤–≥–Ķ–Ĺ–ł—é –ď–Ķ–ĺ—Ä–≥–ł–Ķ–≤–ł—á—É –í–Ķ—Ä–Ķ–≤–ļ–ł–Ĺ—É –∑–į –≤—č–Ņ–ĺ–Ľ–Ĺ–Ķ–Ĺ–ł–Ķ –≤¬†–Ņ—Ä–ĺ–≥—Ä–į–ľ–ľ–Ķ Microsoft Excel —Ā–ł–ľ—É–Ľ—Ź—Ü–ł–Ļ –ī–Ľ—Ź –ī–į–Ĺ–Ĺ–ĺ–Ļ —Ā—ā–į—ā—Ć–ł.
A.A. Putilov, DBSci, PhD
Institute of Higher Nervous Activity and Neurophysiology of the Russian Academy of Sciences, Moscow
Federal Research Centre for Fundamental and Translational Medicine, Novosibirsk
Contact person: Arcady A. Putilov, putilov@ngs.ru
Purpose. People believe that, by extending sleep on the weekend, they can compensate its reduction during the preceding weekdays. This belief, however, has never been supported with simulations based on a model of sleep-wake regulation. Therefore, such a model was applied to debunk the myth of catchup weekend sleep along with other myths around the phenomenon of ‚Äėsocial jet lag‚Äô (a misalignment between work hours and preferred sleep times governed by the body clocks entrained to the 24-hour light-dark cycle rather than to any social time cue, e.g., work hours).
Material and methods. To simulate sleep times reported in the literature for 190 samples (mean ages vary from 0.5 to 60 years), A. Putilov (1995) version of the classical two-process model of sleep-wake regulation S. Daan et al. (1984) was used, and additional 117 samples were collected from the most recent publications to confirm the results.
Results. The empirical analysis of sleep times and their simulations suggested that sleep debt cannot be accumulated after early morning awakenings on weekdays, and sleep lost on weekdays due to such scheduled early wakeups cannot be repaid; in general, clock times for weekend sleep cannot be solely used to distinguish between ‚Äėlarks‚Äô and ‚Äėowls‚Äô, and ‚Äėsocial jet lag‚Äô cannot be more prominent in ‚Äėowls‚Äô than ‚Äėlarks‚Äô the circadian phase cannot delay in adolescents following the delay of their sleep times, and it cannot advance back despite advancing sleep times in middle-aged adults; a sleep phase determined by level of electroencephalogram slow wave activity in the morning hours remains rather stable throughout the week, while the circadian phase in ‚Äėowls‚Äô can be shifted back-and-forth relative to natural light regimen throughout the week.
Conclusion. It seems that the chronobiologial mechanisms underlying ‚Äėsocial jet lag‚Äô require rethinking.
–£–≤–į–∂–į–Ķ–ľ—č–Ļ –Ņ–ĺ—Ā–Ķ—ā–ł—ā–Ķ–Ľ—Ć uMEDp!
–£–≤–Ķ–ī–ĺ–ľ–Ľ—Ź–Ķ–ľ –í–į—Ā –ĺ —ā–ĺ–ľ, —á—ā–ĺ –∑–ī–Ķ—Ā—Ć —Ā–ĺ–ī–Ķ—Ä–∂–ł—ā—Ā—Ź –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł—Ź, –Ņ—Ä–Ķ–ī–Ĺ–į–∑–Ĺ–į—á–Ķ–Ĺ–Ĺ–į—Ź –ł—Ā–ļ–Ľ—é—á–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ –ī–Ľ—Ź —Ā–Ņ–Ķ—Ü–ł–į–Ľ–ł—Ā—ā–ĺ–≤ –∑–ī—Ä–į–≤–ĺ–ĺ—Ö—Ä–į–Ĺ–Ķ–Ĺ–ł—Ź.
–ē—Ā–Ľ–ł –í—č –Ĺ–Ķ —Ź–≤–Ľ—Ź–Ķ—ā–Ķ—Ā—Ć —Ā–Ņ–Ķ—Ü–ł–į–Ľ–ł—Ā—ā–ĺ–ľ –∑–ī—Ä–į–≤–ĺ–ĺ—Ö—Ä–į–Ĺ–Ķ–Ĺ–ł—Ź, –į–ī–ľ–ł–Ĺ–ł—Ā—ā—Ä–į—Ü–ł—Ź –Ĺ–Ķ –Ĺ–Ķ—Ā–Ķ—ā –ĺ—ā–≤–Ķ—ā—Ā—ā–≤–Ķ–Ĺ–Ĺ–ĺ—Ā—ā–ł –∑–į –≤–ĺ–∑–ľ–ĺ–∂–Ĺ—č–Ķ –ĺ—ā—Ä–ł—Ü–į—ā–Ķ–Ľ—Ć–Ĺ—č–Ķ –Ņ–ĺ—Ā–Ľ–Ķ–ī—Ā—ā–≤–ł—Ź, –≤–ĺ–∑–Ĺ–ł–ļ—ą–ł–Ķ –≤ —Ä–Ķ–∑—É–Ľ—Ć—ā–į—ā–Ķ —Ā–į–ľ–ĺ—Ā—ā–ĺ—Ź—ā–Ķ–Ľ—Ć–Ĺ–ĺ–≥–ĺ –ł—Ā–Ņ–ĺ–Ľ—Ć–∑–ĺ–≤–į–Ĺ–ł—Ź –í–į–ľ–ł –ł–Ĺ—Ą–ĺ—Ä–ľ–į—Ü–ł–ł —Ā –Ņ–ĺ—Ä—ā–į–Ľ–į –Ī–Ķ–∑ –Ņ—Ä–Ķ–ī–≤–į—Ä–ł—ā–Ķ–Ľ—Ć–Ĺ–ĺ–Ļ –ļ–ĺ–Ĺ—Ā—É–Ľ—Ć—ā–į—Ü–ł–ł —Ā –≤—Ä–į—á–ĺ–ľ.
–Ě–į–∂–ł–ľ–į—Ź –Ĺ–į –ļ–Ĺ–ĺ–Ņ–ļ—É ¬ę–í–ĺ–Ļ—ā–ł¬Ľ, –í—č –Ņ–ĺ–ī—ā–≤–Ķ—Ä–∂–ī–į–Ķ—ā–Ķ, —á—ā–ĺ —Ź–≤–Ľ—Ź–Ķ—ā–Ķ—Ā—Ć –≤—Ä–į—á–ĺ–ľ –ł–Ľ–ł —Ā—ā—É–ī–Ķ–Ĺ—ā–ĺ–ľ –ľ–Ķ–ī–ł—Ü–ł–Ĺ—Ā–ļ–ĺ–≥–ĺ –≤—É–∑–į.