–Ґ—А–Є —Б—В–∞–і–Є–Є –Љ–µ–і–ї–µ–љ–љ–Њ–≥–Њ —Б–љ–∞: —Е—А–Њ–љ–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–∞—П —А–µ–∞–ї—М–љ–Њ—Б—В—М –Є–ї–Є –≤—Б–µ–≥–Њ –ї–Є—И—М —Г—Б–ї–Њ–≤–љ–Њ–µ –і–µ–ї–µ–љ–Є–µ –њ—А–Њ—Ж–µ—Б—Б–∞ –њ–Њ—Б—В–µ–њ–µ–љ–љ–Њ–≥–Њ —Г–≥–ї—Г–±–ї–µ–љ–Є—П —Б–љ–∞?
- –Р–љ–љ–Њ—В–∞—Ж–Є—П
- –°—В–∞—В—М—П
- –°—Б—Л–ї–Ї–Є
- English

–Т–≤–µ–і–µ–љ–Є–µ
–Э–∞—Г–Ї–∞ –Њ¬†—Б–љ–µ –≤–Њ–Ј–љ–Є–Ї–ї–∞ –Є¬†—А–∞–Ј–≤–Є–≤–∞–ї–∞—Б—М –≤¬†—В–µ—Б–љ–Њ–є —Б–≤—П–Ј–Є —Б¬†–Њ—В–Ї—А—Л—В–Є—П–Љ–Є, —Г–Ї–∞–Ј—Л–≤–∞—О—Й–Є–Љ–Є –љ–∞¬†—Б—В–∞–і–Є–є–љ—Г—О –њ—А–Є—А–Њ–і—Г —Б–љ–∞. –°–љ–∞—З–∞–ї–∞ A.L. Loomis –Є¬†—Б–Њ–∞–≤—В. –Њ–њ—Г–±–ї–Є–Ї–Њ–≤–∞–ї–Є –њ—А–Є–Љ–µ—А—Л –Ј–∞–њ–Є—Б–µ–є —Н–ї–µ–Ї—В—А–Њ—Н–љ—Ж–µ—Д–∞–ї–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Њ–≥–Њ (–≠–≠–У) —Б–Є–≥–љ–∞–ї–∞, —Б–љ—П—В–Њ–≥–Њ —Б¬†–њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є¬†–≥–Њ–ї–Њ–≤—Л —Б–њ—П—Й–µ–≥–Њ —З–µ–ї–Њ–≤–µ–Ї–∞. –Ч–∞–њ–Є—Б–Є —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤–Њ–≤–∞–ї–Є –Њ¬†—В–Њ–Љ, —З—В–Њ –≤–Њ–ї–љ–Њ–≤–∞—П —Б—В—А—Г–Ї—В—Г—А–∞ –≠–≠–У –њ—А–µ—В–µ—А–њ–µ–≤–∞–µ—В –Њ–њ—А–µ–і–µ–ї–µ–љ–љ—Л–µ —Б—В–∞–і–Є–є–љ—Л–µ –Є–Ј–Љ–µ–љ–µ–љ–Є—П –≤¬†—Е–Њ–і–µ –њ–Њ–≥—А—Г–ґ–µ–љ–Є—П –≤¬†—Б–Њ–љ –Є¬†–њ–Њ –Љ–µ—А–µ –µ–≥–Њ –њ–Њ—Б–ї–µ–і—Г—О—Й–µ–≥–Њ —Г–≥–ї—Г–±–ї–µ–љ–Є—П [1]. –Ч–∞—В–µ–Љ E. Aserinsky –Є¬†N. Kleitman –і–Њ–њ–Њ–ї–љ–Є–ї–Є –і–∞–љ–љ–Њ–µ –Њ–њ–Є—Б–∞–љ–Є–µ –љ–∞–±–ї—О–і–µ–љ–Є–µ–Љ –њ–µ—А–Є–Њ–і–Є—З–µ—Б–Ї–Є –≤–Њ–Ј–љ–Є–Ї–∞—О—Й–Є—Е –≤–Ї—А–∞–њ–ї–µ–љ–Є–є –љ–µ–Њ–±—Л—З–љ–Њ–є —Д–∞–Ј—Л —Б–љ–∞, –±—Л—Б—В—А–Њ–≤–Њ–ї–љ–Њ–≤–∞—П —Б—В—А—Г–Ї—В—Г—А–∞ –Ї–Њ—В–Њ—А—Л—Е –љ–∞–њ–Њ–Љ–Є–љ–∞–ї–∞ —В–∞–Ї–Њ–≤—Г—О –њ—А–Є –і—А–µ–Љ–Њ—В–µ –Є–ї–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–Є [2]. –С–Њ–ї–µ–µ –і–µ—В–∞–ї—М–љ—Л–µ –Є–љ—Б—В—А—Г–Љ–µ–љ—В–∞–ї—М–љ—Л–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П, –Њ—Б—Г—Й–µ—Б—В–≤–ї–µ–љ–љ—Л–µ –љ–∞¬†–њ—А–Њ—В—П–ґ–µ–љ–Є–Є –њ–Њ—Б–ї–µ–і—Г—О—Й–Є—Е 15 –ї–µ—В, –њ—А–Є–≤–µ–ї–Є –Ї¬†—Б–Њ–Ј–і–∞–љ–Є—О –µ–і–Є–љ–Њ–є —Б—В–∞–љ–і–∞—А—В–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –Њ–±—К–µ–Ї—В–Є–≤–љ–Њ–≥–Њ –Њ–њ–Є—Б–∞–љ–Є—П —Б–љ–∞ –≤¬†—Д–Њ—А–Љ–µ¬†–≥–Є–њ–љ–Њ–≥—А–∞–Љ–Љ—Л. –°—Г—В—М —Н—В–Њ–≥–Њ –Њ–њ–Є—Б–∞–љ–Є—П –Ј–∞–Ї–ї—О—З–∞–µ—В—Б—П –≤¬†–і–µ–ї–µ–љ–Є–Є –Є—Б—Е–Њ–і–љ–Њ–є –Ј–∞–њ–Є—Б–Є –љ–∞¬†–Є–љ—В–µ—А–≤–∞–ї—Л, –Њ—В–љ–Њ—Б—П—Й–Є–µ—Б—П –Ї¬†—Б–Њ—Б—В–Њ—П–љ–Є—П–Љ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†—Б–љ–∞, –і–≤—Г–Љ —Д–∞–Ј–∞–Љ¬†вАУ –Љ–µ–і–ї–µ–љ–љ–Њ–≥–Њ (–Є–љ–∞—З–µ –Њ—А—В–Њ–і–Њ–Ї—Б–∞–ї—М–љ–Њ–≥–Њ) –Є¬†–±—Л—Б—В—А–Њ–≥–Њ (–Є–љ–∞—З–µ –њ–∞—А–∞–і–Њ–Ї—Б–∞–ї—М–љ–Њ–≥–Њ) —Б–љ–∞¬†вАУ –Є¬†—З–µ—В—Л—А–µ–Љ —Б—В–∞–і–Є—П–Љ –Љ–µ–і–ї–µ–љ–љ–Њ–≥–Њ —Б–љ–∞ (–њ–µ—А–≤–Њ–є, –≤—В–Њ—А–Њ–є, —В—А–µ—В—М–µ–є –Є¬†—З–µ—В–≤–µ—А—В–Њ–є) [3].
–Я–µ—А–≤–Њ–љ–∞—З–∞–ї—М–љ–Њ –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Њ–µ –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ —В–∞–Ї–Њ–є —Б–Є—Б—В–µ–Љ—Л –Њ–Ј–љ–∞—З–∞–ї–Њ –Љ–љ–Њ–≥–Њ—З–∞—Б–Њ–≤—Г—О —А–∞–±–Њ—В—Г –Њ–і–љ–Њ–≥–Њ, –∞¬†–ї—Г—З—И–µ –і–≤—Г—Е –Њ–њ—Л—В–љ—Л—Е —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤¬†вАУ ¬Ђ—Б–Ї–Њ—А–µ—А–Њ–≤¬ї (–∞–љ–≥–ї. scoring¬†вАУ —Б—З–Є—В–∞—В—М¬†вАУ –њ—А–Є–Љ. —А–µ–і.). –Ъ–∞–ґ–і—Л–є –Є–Ј¬†–љ–Є—Е –і–Њ–ї–ґ–µ–љ –±—Л–ї –љ–µ–Ј–∞–≤–Є—Б–Є–Љ–Њ –Њ—В¬†–і—А—Г–≥–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–Є—В—М —Б–≤–Њ–є –≤–∞—А–Є–∞–љ—В¬†–≥–Є–њ–љ–Њ–≥—А–∞–Љ–Љ—Л. –Ш–љ–∞—З–µ¬†–≥–Њ–≤–Њ—А—П, —А–∞–Ј–±–Є—В—М –њ–Њ–ї–љ—Л–є –Љ–љ–Њ–≥–Њ—З–∞—Б–Њ–≤–Њ–є –љ–∞–±–Њ—А 20- –Є–ї–Є 30-—Б–µ–Ї—Г–љ–і–љ—Л—Е —Н–њ–Њ—Е –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞ –љ–∞¬†–Є–љ—В–µ—А–≤–∞–ї—Л, –Њ–±–Њ–Ј–љ–∞—З–µ–љ–љ—Л–µ –Ї–∞–Ї –Њ–њ—А–µ–і–µ–ї–µ–љ–љ–Њ–µ —Б–Њ—Б—В–Њ—П–љ–Є–µ (–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ –Є–ї–Є —Б–Њ–љ), —Д–∞–Ј–∞ —Б–љ–∞ (–Љ–µ–і–ї–µ–љ–љ–∞—П –Є–ї–Є –±—Л—Б—В—А–∞—П) –Є¬†—Б—В–∞–і–Є—П –Љ–µ–і–ї–µ–љ–љ–Њ–≥–Њ —Б–љ–∞ (–њ–µ—А–≤–∞—П, –≤—В–Њ—А–∞—П, —В—А–µ—В—М—П –Є–ї–Є —З–µ—В–≤–µ—А—В–∞—П).
–Ш—Б—В–Њ—А–Є—П —Н—В–Њ–є –њ—А–Њ—Б—В–Њ–є —Б–Є—Б—В–µ–Љ—Л —Г–і–Є–≤–Є—В–µ–ї—М–љ–∞ —В–µ–Љ, —З—В–Њ, –љ–µ—Б–Љ–Њ—В—А—П –љ–∞¬†–≤–µ—Б—М –і–∞–ї—М–љ–µ–є—И–Є–є –њ—А–Њ–≥—А–µ—Б—Б –љ–∞—Г–Ї–Є –Њ¬†—Б–љ–µ –Є¬†–Ї–Њ–Љ–њ—М—О—В–µ—А–Є–Ј–∞—Ж–Є—О –µ–≥–Њ –ї–∞–±–Њ—А–∞—В–Њ—А–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є, –Њ–љ–∞ –і–Њ —Б–Є—Е –њ–Њ—А –≤—Е–Њ–і–Є—В –≤¬†—З–Є—Б–ї–Њ –Њ–±—П–Ј–∞—В–µ–ї—М–љ—Л—Е –њ—А–Њ—Ж–µ–і—Г—А –љ–∞—Г—З–љ–Њ–≥–Њ –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Њ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П —Б–љ–∞. –Ґ–∞–Ї–∞—П –њ—А–Њ—Ж–µ–і—Г—А–∞ —Б—З–Є—В–∞–µ—В—Б—П –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л–Љ —Г—Б–ї–Њ–≤–Є–µ–Љ –і–ї—П –њ–Њ—Б–ї–µ–і—Г—О—Й–µ–є –Њ–±—А–∞–±–Њ—В–Ї–Є –Є¬†–њ—Г–±–ї–Є–Ї–∞—Ж–Є–Є —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤ –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –ї—О–±–Њ–≥–Њ –љ–∞—Г—З–љ–Њ–≥–Њ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞ –њ–Њ¬†–Є–Ј—Г—З–µ–љ–Є—О —Б–љ–∞ –Є¬†–њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –ї—О–±–Њ–≥–Њ –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Њ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –µ–≥–Њ —А–∞—Б—Б—В—А–Њ–є—Б—В–≤. –Ґ–Њ–ї—М–Ї–Њ —Б—А–∞–≤–љ–Є—В–µ–ї—М–љ–Њ –љ–µ–і–∞–≤–љ–Њ –±—Л–ї–∞ –њ—А–µ–і–њ—А–Є–љ—П—В–∞ —Б–∞–Љ–∞—П —А–∞–і–Є–Ї–∞–ї—М–љ–∞—П –Ј–∞ –њ—П—В–Є–і–µ—Б—П—В–Є–ї–µ—В–љ—О—О –Є—Б—В–Њ—А–Є—О —Н—В–Њ–є —Б—В–∞–љ–і–∞—А—В–љ–Њ–є –Ї–ї–∞—Б—Б–Є—Д–Є–Ї–∞—Ж–Є–Є –њ–Њ–њ—Л—В–Ї–∞ –µ–µ —А–µ–≤–Є–Ј–Є–Є [4, 5]. –Ю–љ–∞ –њ–Њ¬†–±–Њ–ї—М—И–Њ–Љ—Г —Б—З–µ—В—Г —Б–≤–µ–ї–∞—Б—М –ї–Є—И—М –Ї¬†–Њ–±—К–µ–і–Є–љ–µ–љ–Є—О —В—А–µ—В—М–µ–є –Є¬†—З–µ—В–≤–µ—А—В–Њ–є —Б—В–∞–і–Є–є, —З—В–Њ, –Ї—Б—В–∞—В–Є —Б–Ї–∞–Ј–∞—В—М, –Є¬†—В–∞–Ї –љ–∞¬†–њ—А–Њ—В—П–ґ–µ–љ–Є–Є –Љ–љ–Њ–≥–Є—Е –њ—А–µ–і—И–µ—Б—В–≤—Г—О—Й–Є—Е –ї–µ—В —Г–ґ–µ –і–µ–ї–∞–ї–Њ—Б—М –≤¬†–±–Њ–ї—М—И–Є–љ—Б—В–≤–µ —Д—Г–љ–і–∞–Љ–µ–љ—В–∞–ї—М–љ—Л—Е –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є. –≠—В–∞ –Њ–±—К–µ–і–Є–љ–µ–љ–љ–∞—П —Б—В–∞–і–Є—П –Њ–±—Л—З–љ–Њ –Є–Љ–µ–љ–Њ–≤–∞–ї–∞—Б—М –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤—Л–Љ —Б–љ–Њ–Љ (—В–∞–Ї–ґ–µ –Є—Б–њ–Њ–ї—М–Ј—Г–µ—В—Б—П —В–µ—А–Љ–Є–љ ¬Ђ–і–µ–ї—М—В–∞-—Б–Њ–љ¬ї¬†вАУ –њ—А–Є–Љ. —А–µ–і.), –∞¬†—В–µ–њ–µ—А—М —В–µ—А–Љ–Є–љ ¬Ђ—В—А–µ—В—М—П —Б—В–∞–і–Є—П¬ї –њ–Њ–њ—А–Њ—Б—В—Г –њ—А–µ–≤—А–∞—В–Є–ї—Б—П –≤¬†—Б–Є–љ–Њ–љ–Є–Љ –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–≥–Њ —Б–љ–∞. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –Њ–±—Б—Г–ґ–і–∞–ї–∞—Б—М –Є–і–µ—П –≤–≤–µ–і–µ–љ–Є—П –љ–Њ–≤–Њ–є —Б—В–∞–і–Є–Є —Б–Њ–љ–ї–Є–≤–Њ—Б—В–Є, –Ї–Њ—В–Њ—А–∞—П –і–Њ–ї–ґ–љ–∞ –±—Л–ї–∞ —З–∞—Б—В–Є—З–љ–Њ –њ–µ—А–µ–Ї—А—Л–≤–∞—В—М—Б—П —Б¬†–њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–µ–є —Б–љ–∞ –Є¬†—Б–Њ—Б—В–Њ—П–љ–Є–µ–Љ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –љ–Њ¬†—Н—В–Њ –њ—А–µ–і–ї–Њ–ґ–µ–љ–Є–µ –±—Л–ї–Њ –≤¬†–Ї–Њ–љ—Ж–µ –Ї–Њ–љ—Ж–Њ–≤ –Њ—В–≤–µ—А–≥–љ—Г—В–Њ [4]. –Ь–µ–ґ–і—Г —В–µ–Љ —Б¬†—Г—Б—В–љ–Њ–є –Є–ї–Є –њ–µ—З–∞—В–љ–Њ–є –Ї—А–Є—В–Є–Ї–Њ–є –≤¬†–∞–і—А–µ—Б —Б—В–∞–і–Є–є–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л –Њ–њ–Є—Б–∞–љ–Є—П —Б–љ–∞ –љ–µ¬†–≤—Л—Б—В—Г–њ–∞–ї —А–∞–Ј–≤–µ —З—В–Њ —В–Њ–ї—М–Ї–Њ –ї–µ–љ–Є–≤—Л–є [6]. –Э–µ¬†–≤–і–∞–≤–∞—П—Б—М –≤–Њ –≤—Б–µ —А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–љ—Л–µ –∞—Б–њ–µ–Ї—В—Л —Н—В–Њ–є –Ї—А–Є—В–Є–Ї–Є, –Њ—В–Љ–µ—В–Є–Љ –ї–Є—И—М –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –≤–∞–ґ–љ—Л—Е –і–ї—П –њ–Њ—Б–ї–µ–і—Г—О—Й–µ–≥–Њ –Є–Ј–ї–Њ–ґ–µ–љ–Є—П –љ–µ–і–Њ—Б—В–∞—В–Ї–Њ–≤ —Б—В–∞–і–Є–є–љ–Њ–є –Ї–ї–∞—Б—Б–Є—Д–Є–Ї–∞—Ж–Є–Є —Б–љ–∞ –Є¬†—В–µ —Б–ї–µ–і—Б—В–≤–Є—П, –Ї¬†–Ї–Њ—В–Њ—А—Л–Љ —Н—В–Є –љ–µ–і–Њ—Б—В–∞—В–Ї–Є –њ—А–Є–≤–Њ–і—П—В.
–Ґ—А–∞–і–Є—Ж–Є–Њ–љ–љ–Њ —Б–Њ–љ –Њ–њ—А–µ–і–µ–ї—П—О—В –Ї–∞–Ї –њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Њ–µ —Б–Њ—Б—В–Њ—П–љ–Є–µ —Ж–µ–ї–Њ–≥–Њ –Њ—А–≥–∞–љ–Є–Ј–Љ–∞. –Ф–ї—П –≤—Л–і–µ–ї–µ–љ–Є—П —Б—В–∞–і–Є–є —Б–љ–∞ –Є—Б–њ–Њ–ї—М–Ј—Г—О—В—Б—П –і–∞–љ–љ—Л–µ, –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ —Б¬†–Њ–і–љ–Њ–≥–Њ –Є–Ј¬†—Г—З–∞—Б—В–Ї–Њ–≤ –љ–∞¬†–њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є¬†–≥–Њ–ї–Њ–≤—Л –Є–ї–Є –і–∞–ґ–µ –Є–љ—Л–µ, —З–µ–Љ –≠–≠–У, –њ–Њ–ї–Є—Б–Њ–Љ–љ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Є–µ —Б–Є–≥–љ–∞–ї—Л. –Я—А–Є–Љ–µ—А–∞–Љ–Є –Љ–Њ–≥—Г—В —Б–ї—Г–ґ–Є—В—М —Н–ї–µ–Ї—В—А–Њ–Њ–Ї—Г–ї–Њ–≥—А–∞–Љ–Љ–∞ –Є¬†—Н–ї–µ–Ї—В—А–Њ–Љ–Є–Њ–≥—А–∞–Љ–Љ–∞, –Ї–Њ—В–Њ—А—Л–µ –Ј–∞–њ–Є—Б—Л–≤–∞—О—В—Б—П –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ —Б¬†–≠–≠–У –і–ї—П –≤—Л—П–≤–ї–µ–љ–Є—П —Д–∞–Ј—Л –±—Л—Б—В—А–Њ–≥–Њ —Б–љ–∞, –∞¬†—В–∞–Ї–ґ–µ –і–∞–љ–љ—Л–µ —Н–ї–µ–Ї—В—А–Њ–Њ–Ї—Г–ї–Њ–≥—А–∞—Д–Є–Є –њ–ї—О—Б –Ј–∞–њ–Є—Б—М –≠–≠–У, –њ–Њ–ї—Г—З–µ–љ–љ–∞—П –Њ—В¬†–Ї–Њ–љ–Ї—А–µ—В–љ–Њ–є (–Њ–±—Л—З–љ–Њ –Ј–∞—В—Л–ї–Њ—З–љ–Њ–є) –Њ–±–ї–∞—Б—В–Є, –Ї–Њ—В–Њ—А—Л–µ –Є—Б–њ–Њ–ї—М–Ј—Г—О—В—Б—П –і–ї—П —А–∞–Ј–≥—А–∞–љ–Є—З–µ–љ–Є—П –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†–њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞. –Ю–і–љ–∞–Ї–Њ —Б–њ–µ–Ї—В—А–∞–ї—М–љ—Л–є –∞–љ–∞–ї–Є–Ј –≠–≠–У, –Ј–∞–њ–Є—Б–∞–љ–љ–Њ–є —Б¬†—Н–ї–µ–Ї—В—А–Њ–і–Њ–≤, —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–љ—Л—Е –љ–∞¬†—А–∞–Ј–љ—Л—Е —Г—З–∞—Б—В–Ї–∞—Е¬†–≥–Њ–ї–Њ–≤—Л (–љ–∞–њ—А–Є–Љ–µ—А, –њ–µ—А–µ–і¬≠–љ–µ–Љ –Є¬†–Ј–∞—В—Л–ї–Њ—З–љ–Њ–Љ), —Г–Ї–∞–Ј—Л–≤–∞–µ—В –љ–∞¬†—В–Њ–њ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї—Г—О –њ–Њ—Б—В–µ–њ–µ–љ–љ–Њ—Б—В—М –Є¬†–љ–µ–Њ–і–љ–Њ—А–Њ–і–љ–Њ—Б—В—М –Є–Ј–Љ–µ–љ–µ–љ–Є–є –њ–∞—В—В–µ—А–љ–Њ–≤ –≠–≠–У, –∞¬†—В–∞–Ї–ґ–µ –љ–∞¬†—В–Њ, —З—В–Њ —Б–Њ–љ –љ–µ¬†–Њ–±—П–Ј–∞—В–µ–ї—М–љ–Њ –љ–∞—З–Є–љ–∞–µ—В—Б—П –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ –≤–Њ –≤—Б–µ—Е —Г—З–∞—Б—В–Ї–∞—Е –Ї–Њ—А—Л¬†–≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ [7, 8]. –С–Њ–ї–µ–µ —В–Њ–≥–Њ, —Б–Њ–љ –Є¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ –љ–µ¬†—П–≤–ї—П—О—В—Б—П –≤–Ј–∞–Є–Љ–љ–Њ –Є—Б–Ї–ї—О—З–∞—О—Й–Є–Љ–Є —Б–Њ—Б—В–Њ—П–љ–Є—П–Љ–Є, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г —Е–∞—А–∞–Ї—В–µ—А–љ—Л–µ –і–ї—П –Ї–∞–ґ–і–Њ–≥–Њ –Є–Ј¬†—Н—В–Є—Е —Б–Њ—Б—В–Њ—П–љ–Є–є –≤–Њ–ї–љ–Њ–≤—Л–µ –њ–∞—В—В–µ—А–љ—Л –≠–≠–У –Љ–Њ–≥—Г—В —Б–Њ—Б—Г—Й–µ—Б—В–≤–Њ–≤–∞—В—М –і–Њ—Б—В–∞—В–Њ—З–љ–Њ –і–ї–Є—В–µ–ї—М–љ–Њ–µ –≤—А–µ–Љ—П [7вАУ9]. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –≤–Њ–Ј–Љ–Њ–ґ–љ–∞ –і–Є—Б—Б–Њ—Ж–Є–∞—Ж–Є—П –Љ–µ–ґ–і—Г —Н—В–Є–Љ–Є –њ–∞—В—В–µ—А–љ–∞–Љ–Є –Є¬†–њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–Љ–Є –Є–ї–Є —Б—Г–±—К–µ–Ї—В–Є–≤–љ—Л–Љ–Є –Ї—А–Є—В–µ—А–Є—П–Љ–Є –љ–∞—З–∞–ї–∞ —Б–љ–∞ [10вАУ12].
–Я—А–Є–Љ–µ–љ–µ–љ–Є–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ—Л—Е –Ї–Њ–Љ–њ—М—О—В–µ—А–Є–Ј–Є—А–Њ–≤–∞–љ–љ—Л—Е –Љ–µ—В–Њ–і–Њ–≤ –Њ–±—А–∞–±–Њ—В–Ї–Є –≠–≠–У, –≤¬†–њ–µ—А–≤—Г—О –Њ—З–µ—А–µ–і—М —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞, –Њ–±—Л—З–љ–Њ –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞–µ—В –њ–Њ—Б—В–µ–њ–µ–љ–љ—Л–µ, –∞¬†–љ–µ —А–µ–Ј–Ї–Є–µ –Є–Ј–Љ–µ–љ–µ–љ–Є—П —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–є –Ї–Њ–Љ–њ–Њ–Ј–Є—Ж–Є–Є –≠–≠–У-—Б–Є–≥–љ–∞–ї–∞ –≤¬†–њ—А–Њ—Ж–µ—Б—Б–µ –њ–Њ–≥—А—Г–ґ–µ–љ–Є—П –≤¬†—Б–Њ–љ –Є¬†–≤ —Е–Њ–і–µ –µ–≥–Њ –њ–Њ—Б–ї–µ–і—Г—О—Й–µ–≥–Њ —Г–≥–ї—Г–±–ї–µ–љ–Є—П. –Ш–љ—Л–Љ–Є —Б–ї–Њ–≤–∞–Љ–Є, —Н—В–Є –Љ–µ—В–Њ–і—Л –љ–µ¬†–≤—Л—П–≤–ї—П—О—В —З–µ—В–Ї–Њ –Њ—З–µ—А—З–µ–љ–љ—Л—Е¬†–≥—А–∞–љ–Є—Ж, –Њ—В–і–µ–ї—П—О—Й–Є—Е –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ –Њ—В¬†—Б–љ–∞, –∞¬†—В–∞–Ї–ґ–µ –Њ–і–љ—Г —Б—В–∞–і–Є—О –Љ–µ–і–ї–µ–љ–љ–Њ–≥–Њ —Б–љ–∞ –Њ—В¬†–і—А—Г–≥–Њ–є [13вАУ15]. –Т¬†—З–∞—Б—В–љ–Њ—Б—В–Є, –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –њ–Њ–Ј–≤–Њ–ї—П–µ—В –Њ–њ–Є—Б–∞—В—М –і–Є–љ–∞–Љ–Є–Ї—Г –љ–∞–Є–±–Њ–ї–µ–µ –Љ–Њ—Й–љ–Њ–≥–Њ —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–≥–Њ –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–∞ –≠–≠–У-—Б–Є–≥–љ–∞–ї–∞¬†вАУ –і–µ–ї—М—В–∞-–≤–Њ–ї–љ (—З–∞—Б—В–Њ—В–∞ 0,5вАУ4,5 –У—Ж) –Є–ї–Є –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є. –Ю–љ–∞ –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ —Б–љ–Є–ґ–µ–љ–∞ –≤–Њ –≤—А–µ–Љ—П –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –љ–Њ¬†–Њ–±–љ–∞—А—Г–ґ–Є–≤–∞–µ—В –љ–µ–њ—А–µ—А—Л–≤–љ—Л–є —А–Њ—Б—В —Б—А–∞–Ј—Г –њ–Њ—Б–ї–µ –њ–µ—А–µ—Е–Њ–і–∞ –Ї–Њ¬†—Б–љ—Г –Є¬†–Ј–∞—В–µ–Љ –≤¬†–њ—А–Њ—Ж–µ—Б—Б–µ –µ–≥–Њ –і–∞–ї—М–љ–µ–є—И–µ–≥–Њ —Г–≥–ї—Г–±–ї–µ–љ–Є—П. –†–Њ—Б—В –љ–∞—З–Є–љ–∞–µ—В—Б—П —Б—А–∞–Ј—Г –њ–Њ—Б–ї–µ —Б–Љ–µ–љ—Л –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П (–Њ–±–Њ–Ј–љ–∞—З–∞–µ—В—Б—П –Ї–∞–Ї W) –Ї–Њ—А–Њ—В–Ї–Є–Љ –њ–µ—А–µ—Е–Њ–і–љ—Л–Љ –Є–љ—В–µ—А–≤–∞–ї–Њ–Љ –њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞ (N1). –Ч–∞—В–µ–Љ —А–Њ—Б—В –њ—А–Њ–і–Њ–ї–ґ–∞–µ—В—Б—П –≤–Њ –≤—А–µ–Љ—П –±–Њ–ї–µ–µ –і–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ –Є–љ—В–µ—А–≤–∞–ї–∞ –≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–Є (N2) –Є¬†–і–Њ—Б—В–Є–≥–∞–µ—В –њ–Є–Ї–∞ –љ–∞¬†–Є–љ—В–µ—А–≤–∞–ї–µ —В—А–µ—В—М–µ–є —Б—В–∞–і–Є–Є (N3). –≠—В—Г –њ–Њ—Б–ї–µ–і–љ—О—О —Б—В–∞–і–Є—О –љ–∞–Ј–≤–∞–ї–Є –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤—Л–Љ —Б–љ–Њ–Љ –Є–Ј-–Ј–∞ –і–Њ–Љ–Є–љ–Є—А–Њ–≤–∞–љ–Є—П –≤—Л—Б–Њ–Ї–Њ–∞–Љ–њ–ї–Є—В—Г–і–љ—Л—Е –љ–Є–Ј–Ї–Њ—З–∞—Б—В–Њ—В–љ—Л—Е –≤–Њ–ї–љ (–±–Њ–ї–µ–µ 75 –Љ–Т, 0,5вАУ2,0 –У—Ж). –Я—А–Њ–Є–Ј–≤–Њ–ї—М–љ—Л–µ –њ–Њ—А–Њ–≥–Њ–≤—Л–µ –Ї—А–Є—В–µ—А–Є–Є, –Є—Б–њ–Њ–ї—М–Ј—Г–µ–Љ—Л–µ –і–ї—П –≤—Л—П–≤–ї–µ–љ–Є—П —Н—В–Њ–є —Б—В–∞–і–Є–Є, —Г–њ–Њ–Љ–Є–љ–∞—О—В —В—А–Є —В–∞–Ї–Є—Е –њ–Њ—А–Њ–≥–∞ (—З–Є—Б–ї–µ–љ–љ—Л–є, —З–∞—Б—В–Њ—В–љ—Л–є –Є¬†–∞–Љ–њ–ї–Є—В—Г–і–љ—Л–є). –С–Њ–ї–µ–µ 20% –і–∞–љ–љ–Њ–є 20- –Є–ї–Є 30-—Б–µ–Ї—Г–љ–і–љ–Њ–є —Н–њ–Њ—Е–Є —Б–љ–∞ –і–Њ–ї–ґ–љ–Њ –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞—В—М –∞–Ї—В–Є–≤–љ–Њ—Б—В—М < 2 –У—Ж –∞–Љ–њ–ї–Є—В—Г–і–Њ–є > 75¬†–Љ–Ї–Т [3]. –Ь–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –љ–µ¬†—Г–љ–Є–Ї–∞–ї—М–љ–∞ –і–ї—П —В—А–µ—В—М–µ–є —Б—В–∞–і–Є–Є. –Ю–љ–∞ –Њ–±—Л—З–љ–Њ –њ—А–Є—Б—Г—В—Б—В–≤—Г–µ—В –≤–Њ –≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞, –Є¬†–љ–∞–Њ–±–Њ—А–Њ—В, —Б–≤–Њ–є—Б—В–≤–∞ –≠–≠–У, –њ–Њ¬†–Ї–Њ—В–Њ—А—Л–Љ –≤—Л—П–≤–ї—П—О—В –≤—В–Њ—А—Г—О —Б—В–∞–і–Є—О (—Б–Њ–љ–љ—Л–µ –≤–µ—А–µ—В–µ–љ–∞ –Є¬†–Ъ-–Ї–Њ–Љ–њ–ї–µ–Ї—Б—Л), –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞—О—В—Б—П –Є¬†–≤ —В—А–µ—В—М–µ–є —Б—В–∞–і–Є–Є —Б–љ–∞ [16]. –Я—А–∞–Ї—В–Є–Ї–∞ –њ–Њ–Ї–∞–Ј–∞–ї–∞, —З—В–Њ —В–∞–Ї–Є–µ –Ї—А–Є—В–µ—А–Є–Є —Б–ї–Њ–ґ–љ–Њ –њ—А–Є–Љ–µ–љ—П—В—М –і–∞–ґ–µ –њ—А–Є –Є–Ј—Г—З–µ–љ–Є–Є —Б–љ–∞ –Ј–і–Њ—А–Њ–≤–Њ–≥–Њ —З–µ–ї–Њ–≤–µ–Ї–∞, —В–∞–Ї —З—В–Њ –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ—Б—В—М –≤¬†–њ–Њ–і–Њ–±–љ–Њ–Љ —Б—Г–±—К–µ–Ї—В–Є–≤–љ–Њ–Љ –Њ—Ж–µ–љ–Є–≤–∞–љ–Є–Є –њ–Њ—А–Њ–≥–Њ–≤ –Љ–Њ–ґ–µ—В —Б—В–∞—В—М –њ—А–Є—З–Є–љ–Њ–є —Б–ї–∞–±–Њ–є —Б—В–µ–њ–µ–љ–Є —Б–Њ–≥–ї–∞—Б–Њ–≤–∞–љ–љ–Њ—Б—В–Є –Љ–µ–ґ–і—Г —А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ–Є —А–∞–±–Њ—В—Л —А–∞–Ј–љ—Л—Е ¬Ђ—Б–Ї–Њ—А–µ—А–Њ–≤¬ї [17, 18].
–Ф–µ–є—Б—В–≤–Є—В–µ–ї—М–љ–Њ, –љ–µ–≤—Л—Б–Њ–Ї–∞—П —Б—В–µ–њ–µ–љ—М —Б–Њ–≥–ї–∞—Б–Є—П –±—Л–ї–∞ –Њ–±–љ–∞—А—Г–ґ–µ–љ–∞ –њ—А–Є –Ї–ї–∞—Б—Б–Є—Д–Є–Ї–∞—Ж–Є–Є —Б–Њ—Б–µ–і–љ–Є—Е —Б—В–∞–і–Є–є (W/N1, N1/N2 –Є¬†N2/N3), –њ—А–Є—З–µ–Љ –Њ—Б–Њ–±–µ–љ–љ–Њ —Б–ї–∞–±—Л–є —А–µ–Ј—Г–ї—М—В–∞—В –±—Л–ї –≤—Л—П–≤–ї–µ–љ –њ—А–Є —А–∞–Ј–≥—А–∞–љ–Є—З–µ–љ–Є–Є –њ–µ—А–≤–Њ–є –Є¬†—В—А–µ—В—М–µ–є —Б—В–∞–і–Є–є. –Т¬†—З–∞—Б—В–љ–Њ—Б—В–Є, —Б—А–∞–≤–љ–µ–љ–Є—П —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤ –њ—П—В–Є ¬Ђ—Б–Ї–Њ—А–µ—А–Њ–≤¬ї –Њ–±–љ–∞—А—Г–ґ–Є–ї–Є, —З—В–Њ –њ—А–Њ—Ж–µ–љ—В –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–≥–Њ —Б–Њ–≥–ї–∞—Б–Є—П –Љ–µ–ґ–і—Г –љ–Є–Љ–Є (–њ—П—В—М –Є–Ј¬†–њ—П—В–Є) –њ—А–Є –Ї–ї–∞—Б—Б–Є—Д–Є–Ї–∞—Ж–Є–Є –њ–µ—А–≤–Њ–є –Є¬†—В—А–µ—В—М–µ–є —Б—В–∞–і–Є–є —Б–Њ—Б—В–∞–≤–Є–ї —В–Њ–ї—М–Ї–Њ 3,6 –Є¬†18,6% —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ [18]. –Ґ–∞–Ї–Њ–≥–Њ —А–Њ–і–∞ –њ—А–Њ–±–ї–µ–Љ—Л –Љ–Њ–≥—Г—В —Б—В–∞—В—М –њ—А–Є—З–Є–љ–Њ–є –љ–µ–љ–∞–і–µ–ґ–љ–Њ–є –Њ—Ж–µ–љ–Ї–Є –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Б—В–∞–і–Є–є —Б–љ–∞, –≤–Ї–ї—О—З–∞—П –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є —Б–Њ–љ [17]. –Т–∞–ґ–љ–Њ –Њ—В–Љ–µ—В–Є—В—М –Њ–і–Є–љ —Д–∞–Ї—В. –Ф–Њ –љ–µ–і–∞–≤–љ–µ–≥–Њ –≤—А–µ–Љ–µ–љ–Є –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є —Б–Њ–љ –Є¬†–Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–ї–Є—Б—М –≤¬†–Њ—Б–љ–Њ–≤–љ–Њ–Љ –Ї–∞–Ї –њ–Њ–ї–µ–Ј–љ—Л–µ –Є–љ–і–Є–Ї–∞—В–Њ—А—Л (–Љ–∞—А–Ї–µ—А—Л)¬†–≥–ї—Г–±–Є–љ—Л —Б–љ–∞ –Є¬†–µ–≥–Њ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є. –Ю–і–љ–∞–Ї–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ—Б–ї–µ–і–љ–Є—Е –ї–µ—В –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ –Є–Ј-–Ј–∞ —Г–љ–Є–Ї–∞–ї—М–љ–Њ–є –ґ–Є–Ј–љ–µ–љ–љ–Њ–є —Д—Г–љ–Ї—Ж–Є–Є –Є–Ј–Љ–µ—А–µ–љ–Є–µ –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–≥–Њ —Б–љ–∞ –Є¬†–Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –Љ–Њ–ґ–µ—В –њ—А–µ–і—Б—В–∞–≤–ї—П—В—М —Б–њ–µ—Ж–Є–∞–ї—М–љ—Л–є –Є–љ—В–µ—А–µ—Б –і–ї—П –Є–Ј—Г—З–µ–љ–Є—П –љ–Њ—А–Љ–∞–ї—М–љ–Њ–≥–Њ –Є¬†–њ–∞—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–≥–Њ —Б–љ–∞. –Т¬†—З–∞—Б—В–љ–Њ—Б—В–Є, –±—Л–ї–∞ –≤—Л—П–≤–ї–µ–љ–∞ –Є—Е —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–∞—П —А–Њ–ї—М –і–ї—П –љ–Њ—А–Љ–∞–ї—М–љ–Њ–≥–Њ –њ—А–Њ—Ж–µ—Б—Б–∞ –Ј–∞–њ–Њ–Љ–Є–љ–∞–љ–Є—П –Є¬†–Њ–±—Г—З–µ–љ–Є—П [19вАУ23]. –Т¬†—Б–≤—П–Ј–Є —Б¬†—В–µ–Љ —З—В–Њ –љ–∞–ї–Є—З–Є–µ –Є¬†—Ж–µ–ї–Њ—Б—В–љ–Њ—Б—В—М –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–≥–Њ —Б–љ–∞ —Б–≤—П–Ј–∞–љ—Л —Б–Њ¬†—Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М—О –Ї¬†—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—О –Є¬†—Б–Њ—Е—А–∞–љ–µ–љ–Є—О —Б–ї–µ–і–Њ–≤ –њ–∞–Љ—П—В–Є, –љ–µ–і–Њ—Б—В–∞—В–Њ–Ї –Є–Љ–µ–љ–љ–Њ —Н—В–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞ –Љ–Њ–ґ–µ—В –Њ–±—К—П—Б–љ–Є—В—М –љ–µ–Ї–Њ—В–Њ—А—Л–µ –Ї–Њ–≥–љ–Є—В–Є–≤–љ—Л–µ –љ–∞—А—Г—И–µ–љ–Є—П –њ—А–Є –±–µ—Б—Б–Њ–љ–љ–Є—Ж–µ¬†–≤¬†–њ–Њ–ґ–Є–ї–Њ–Љ –≤–Њ–Ј—А–∞—Б—В–µ [24, 25]. –°–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ, –±–Њ–ї–µ–µ –љ–∞–і–µ–ґ–љ–Њ–µ –Њ–њ—А–µ–і–µ–ї–µ–љ–Є–µ¬†–≥—А–∞–љ–Є—Ж –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–≥–Њ —Б–љ–∞ —Б—В–∞–љ–Њ–≤–Є—В—Б—П –Њ—Б–Њ–±–µ–љ–љ–Њ –∞–Ї—В—Г–∞–ї—М–љ—Л–Љ –і–ї—П –Њ—Ж–µ–љ–Ї–Є –µ–≥–Њ –≤–ї–Є—П–љ–Є—П –љ–∞¬†—В–∞–Ї–Є–µ –љ–∞—А—Г—И–µ–љ–Є—П.
–Э–µ—Б–Љ–Њ—В—А—П –љ–∞¬†—В–Њ —З—В–Њ —Б—В–∞–і–Є–є–љ–∞—П –Ї–ї–∞—Б—Б–Є—Д–Є–Ї–∞—Ж–Є—П –і–∞–µ—В –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –і–Њ–≤–Њ–ї—М–љ–Њ —Н–Ї–Њ–љ–Њ–Љ–љ–Њ–≥–Њ –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ –Њ–њ–Є—Б–∞–љ–Є—П —Б–љ–∞ –њ—Г—В–µ–Љ –≤—Л–і–µ–ї–µ–љ–Є—П –љ–µ–±–Њ–ї—М—И–Њ–≥–Њ –Ї–Њ–ї–Є—З–µ—Б—В–≤–∞ –і–∞-–Є–ї–Є-–љ–µ—В-–њ–µ—А–µ–Љ–µ–љ–љ—Л—Е, –Є–Љ–µ–љ—Г–µ–Љ—Л—Е —Б—В–∞–і–Є—П–Љ–Є, –Њ–љ–∞ –њ–Њ–ї–љ–Њ—Б—В—М—О –Є–≥–љ–Њ—А–Є—А—Г–µ—В –≤–Њ–њ—А–Њ—Б –Њ¬†–љ–µ–є—А–Њ–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–Љ –Ј–љ–∞—З–µ–љ–Є–Є –Є¬†–≤–∞–ї–Є–і–љ–Њ—Б—В–Є —В–∞–Ї–Њ–≥–Њ –њ–Њ–і—А–∞–Ј–і–µ–ї–µ–љ–Є—П [6]. –Т–Њ–Ј–љ–Є–Ї–∞–µ—В –≤–Њ–њ—А–Њ—Б: –Љ–Њ–ґ–љ–Њ –ї–Є —Б–≤–µ—Б—В–Є —Б–њ–µ–Ї—В—А–∞–ї—М–љ—Л–µ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є—П –≠–≠–У-—Б–Є–≥–љ–∞–ї–∞ –Ї¬†—В–∞–Ї–Њ–Љ—Г –ґ–µ —Н–Ї–Њ–љ–Њ–Љ–Є—З–љ–Њ–Љ—Г –Њ–њ–Є—Б–∞–љ–Є—О —Б–љ–∞, –Ї–∞–Ї–Њ–µ –і–∞–µ—В –µ–≥–Њ —Б—В–∞–і–Є–є–љ–∞—П –Ї–ї–∞—Б—Б–Є—Д–Є–Ї–∞—Ж–Є—П, –љ–Њ¬†–≤¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П –њ–Њ–ї—Г—З–Є—В—М –љ–µ–є—А–Њ–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є –Њ—Б–Љ—Л—Б–ї–µ–љ–љ–Њ–µ –Њ–њ–Є—Б–∞–љ–Є–µ? –Т¬†—А—П–і–µ –њ—А–µ–і—Л–і—Г—Й–Є—Е –њ—Г–±–ї–Є–Ї–∞—Ж–Є–є –Љ—Л –њ—А–µ–і—Б—В–∞–≤–Є–ї–Є —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ—Л–µ –і–Њ–Ї–∞–Ј–∞—В–µ–ї—М—Б—В–≤–∞ —В–Њ–≥–Њ, —З—В–Њ –∞–љ–∞–ї–Є–Ј –≠–≠–У-—Б–њ–µ–Ї—В—А–∞ –Љ–µ—В–Њ–і–Њ–Љ¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –њ–Њ–Ј–≤–Њ–ї—П–µ—В —Н–Ї–Њ–љ–Њ–Љ–Є—З–љ–Њ –њ—А–µ–і—Б—В–∞–≤–Є—В—М —В–∞–Ї–Њ–є —Б–њ–µ–Ї—В—А –≤¬†–≤–Є–і–µ –љ–µ–±–Њ–ї—М—И–Њ–≥–Њ –љ–∞–±–Њ—А–∞ –Ј–љ–∞—З–µ–љ–Є–є –љ–µ–Ј–∞–≤–Є—Б–Є–Љ—Л—Е (–Њ—А—В–Њ–≥–Њ–љ–∞–ї—М–љ—Л—Е –Є¬†–љ–µ–Ї–Њ—А—А–µ–ї–Є—А–Њ–≤–∞–љ–љ—Л—Е) –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В [26вАУ33]. –С—Л–ї–∞ —В–∞–Ї–ґ–µ –≤—Л—Б–Ї–∞–Ј–∞–љ–∞¬†–≥–Є–њ–Њ—В–µ–Ј–∞, —З—В–Њ, –≤–Њ-–њ–µ—А–≤—Л—Е, —В—А–∞–і–Є—Ж–Є–Њ–љ–љ—Л–µ —Б–њ–µ–Ї—В—А–∞–ї—М–љ—Л–µ —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є –≠–≠–У-—Б–Є–≥–љ–∞–ї–∞ (–Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –Є¬†–∞–Ї—В–Є–≤–љ–Њ—Б—В—М –≤¬†–∞–ї—М—Д–∞-, —В–µ—В–∞-, —Б–Є–≥–Љ–∞- –Є¬†–њ—А–Њ—З–Є—Е –і–Є–∞–њ–∞–Ј–Њ–љ–∞—Е) –Њ—В—А–∞–ґ–∞—О—В –Ї–Њ–Љ–±–Є–љ–Є—А–Њ–≤–∞–љ–љ–Њ–µ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ –љ–µ—Б–Ї–Њ–ї—М–Ї–Є—Е –Њ–њ–њ–Њ–љ–Є—А—Г—О—Й–Є—Е –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ —А–µ–≥—Г–ї—П—Ж–Є–Є —Ж–Є–Ї–ї–∞ ¬Ђ—Б–Њ–љ¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬ї. –Т–Њ-–≤—В–Њ—А—Л—Е, –Є–Ј–≤–ї–µ—З–µ–љ–Є–µ¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В —Н—В–Њ–≥–Њ —Б–њ–µ–Ї—В—А–∞ –њ–Њ–Ј–≤–Њ–ї—П–µ—В –Њ–±–љ–∞—А—Г–ґ–Є—В—М –≠–≠–У-–њ–Њ–і–њ–Є—Б–Є —Н—В–Є—Е –Њ—В–і–µ–ї—М–љ—Л—Е –Њ–њ–њ–Њ–љ–Є—А—Г—О—Й–Є—Е –њ—А–Њ—Ж–µ—Б—Б–Њ–≤, –∞¬†–љ–µ –Є—Е —Б–Њ–≤–Љ–µ—Б—В–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ [26вАУ28].
–Ш–і–µ—П –∞–љ—В–∞–≥–Њ–љ–Є—Б—В–Є—З–µ—Б–Ї–Њ–є –њ—А–Є—А–Њ–і—Л —Е—А–Њ–љ–Њ—А–µ–≥—Г–ї—П—Ж–Є–Є –њ–Њ–ї–љ–Њ—Б—В—М—О —Б–Њ–≥–ї–∞—Б—Г–µ—В—Б—П —Б¬†–љ–µ–є—А–Њ–љ–∞—Г—З–љ—Л–Љ–Є –Њ—В–Ї—А—Л—В–Є—П–Љ–Є, –њ—А–µ–і–њ–Њ–ї–∞–≥–∞—О—Й–Є–Љ–Є, —З—В–Њ —А–µ–≥—Г–ї—П—Ж–Є—П —Ж–Є–Ї–ї–∞ ¬Ђ—Б–Њ–љ¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬ї –Ј–∞–Ї–ї—О—З–∞–µ—В—Б—П –≤–Њ –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є–Є –Њ–њ–њ–Њ–љ–Є—А—Г—О—Й–Є—Е –њ—А–Њ—Ж–µ—Б—Б–Њ–≤. –Ю–і–љ–Є —П–≤–ї—П—О—В—Б—П –њ—А–Њ–Љ–Њ—В–Њ—А–∞–Љ–Є —Б–љ–∞ –Є¬†–Є–љ–≥–Є–±–Є—В–Њ—А–∞–Љ–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –∞¬†–і—А—Г–≥–Є–µ, –љ–∞–Њ–±–Њ—А–Њ—В, –њ—А–Њ–Љ–Њ—В–Њ—А–∞–Љ–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†–Є–љ–≥–Є–±–Є—В–Њ—А–∞–Љ–Є —Б–љ–∞ –Є–ї–Є –Њ—В–і–µ–ї—М–љ—Л—Е –µ–≥–Њ —Д–∞–Ј [34вАУ36]. –Т¬†—З–∞—Б—В–љ–Њ—Б—В–Є, –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П —Г–Ї–∞–Ј—Л–≤–∞—О—В –љ–∞¬†—В–Њ, —З—В–Њ –≤–µ–љ—В—А–Њ–ї–∞—В–µ—А–∞–ї—М–љ—Л–µ –њ—А–µ–Њ–њ—В–Є—З–µ—Б–Ї–Є–µ —П–і—А–∞¬†–≥–Є–њ–Њ—В–∞–ї–∞–Љ—Г—Б–∞ —Б–Њ–і–µ—А–ґ–∞—В –Є¬†–љ–µ–є—А–Њ–љ—Л-–њ—А–Њ–Љ–Њ—В–Њ—А—Л —Б–љ–∞, –Є¬†–љ–µ–є—А–Њ–љ—Л-–њ—А–Њ–Љ–Њ—В–Њ—А—Л –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –Ї–Њ—В–Њ—А—Л–µ –Ї–Њ–љ—В—А–Њ–ї–Є—А—Г—О—В —Б–Љ–µ–љ—Г —Б–Њ—Б—В–Њ—П–љ–Є–є —Б–љ–∞¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –∞¬†—В–∞–Ї–ґ–µ —З–µ—А–µ–і–Њ–≤–∞–љ–Є–µ –Љ–µ–і–ї–µ–љ–љ–Њ–є –Є¬†–±—Л—Б—В—А–Њ–є —Д–∞–Ј —Б–љ–∞ [35].
–¶–µ–ї—М
–Я—А–Њ–≤–µ—А–Ї–∞¬†–≥–Є–њ–Њ—В–µ–Ј—Л –Њ¬†—В–Њ–Љ, —З—В–Њ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –Њ–њ–Є—Б—Л–≤–∞—В—М –ї—О–±—Л–µ –≠–≠–У-–Ј–∞–њ–Є—Б–Є —Б–љ–∞ –≤¬†–≤–Є–і–µ –њ–Њ—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М–љ–Њ—Б—В–Є –љ–µ–±–Њ–ї—М—И–Њ–≥–Њ —З–Є—Б–ї–∞ –і–∞-–Є–ї–Є-–љ–µ—В-–њ–µ—А–µ–Љ–µ–љ–љ—Л—Е, –љ–∞–Ј—Л–≤–∞–µ–Љ—Л—Е —Б—В–∞–і–Є—П–Љ–Є, –Њ–±—К—П—Б–љ—П–µ—В—Б—П —В–∞–Ї–Є–Љ —Б–≤–Њ–є—Б—В–≤–Њ–Љ –Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤ —А–µ–≥—Г–ї—П—Ж–Є–Є —Б–љ–∞¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –Ї–∞–Ї —А–µ—Ж–Є–њ—А–Њ–Ї–љ—Л–µ –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є—П –Љ–µ–ґ–і—Г –њ—А–Њ—Ж–µ—Б—Б–∞–Љ–Є-–њ—А–Њ–Љ–Њ—В–Њ—А–∞–Љ–Є —Б–љ–∞ –Є¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –Ї–Њ—В–Њ—А—Л–µ –Є–љ–≥–Є–±–Є—А—Г—О—В –і—А—Г–≥ –і—А—Г–≥–∞ –њ—А–Є —Б–Љ–µ–љ–µ –Њ–і–љ–Њ–≥–Њ —Б–Њ—Б—В–Њ—П–љ–Є—П/—Б—Г–±—Б–Њ—Б—В–Њ—П–љ–Є—П —Б–љ–∞¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –і—А—Г–≥–Є–Љ.
–Ь–∞—В–µ—А–Є–∞–ї –Є¬†–Љ–µ—В–Њ–і—Л
–Ф–ї—П –њ—А–Њ–≤–µ—А–Ї–Є¬†–≥–Є–њ–Њ—В–µ–Ј—Л –±—Л–ї–Є –њ—А–Њ–∞–љ–∞–ї–Є–Ј–Є—А–Њ–≤–∞–љ—Л –і–∞–љ–љ—Л–µ –і–≤—Г—Е —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–Њ–≤.
–Я–µ—А–≤—Л–є —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В –≤–Ї–ї—О—З–∞–ї –≠–≠–У-–Ј–∞–њ–Є—Б–Є —Б–љ–∞ –Є¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П 14 –ґ–µ–љ—Й–Є–љ. –Ъ–∞–ґ–і–∞—П –Є–Ј¬†–љ–Є—Е –њ—А–Є–љ—П–ї–∞ —Г—З–∞—Б—В–Є–µ –≤¬†–і–≤—Г—Е –Є–і–µ–љ—В–Є—З–љ—Л—Е —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞—Е. –£—З–∞—Б—В–љ–Є—Ж–∞ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞ –љ–∞—Е–Њ–і–Є–ї–∞—Б—М –≤¬†–ї–∞–±–Њ—А–∞—В–Њ—А–Є–Є –∞–і–∞–њ—В–∞—Ж–Є–Њ–љ–љ—Г—О –љ–Њ—З—М, –≤¬†—Б–ї–µ–і—Г—О—Й—Г—О –љ–Њ—З—М –њ—А–Њ–≤–Њ–і–Є–ї–∞—Б—М –њ–Њ–ї–Є—Б–Њ–Љ–љ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–∞—П –Ј–∞–њ–Є—Б—М —Б–љ–∞. –°–≤–µ—В –≤—Л–Ї–ї—О—З–∞–ї—Б—П –≤¬†23 —З–∞—Б–∞, –Є¬†–і–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М —Б–љ–∞ –љ–µ¬†–Њ–≥—А–∞–љ–Є—З–Є–≤–∞–ї–∞—Б—М. –Т¬†–њ–Њ—Б–ї–µ–і—Г—О—Й–Є–µ —Б—Г—В–Ї–Є —Г—З–∞—Б—В–љ–Є—Ж–µ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞ –њ—А–µ–і–ї–∞–≥–∞–ї–Њ—Б—М –Ј–∞—Б–љ—Г—В—М –≤¬†—В–µ—З–µ–љ–Є–µ 20 –Љ–Є–љ—Г—В –≤¬†10, 12, 14, 16, 18, 23, 01, 03 –Є¬†05 —З–∞—Б–Њ–≤.
–Т—В–Њ—А–Њ–є —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В –≤–Ї–ї—О—З–∞–ї –≠–≠–У-–Ј–∞–њ–Є—Б–Є —Б–љ–∞ –Є¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П 18 –Љ–Њ–ї–Њ–і—Л—Е –Љ—Г–ґ—З–Є–љ. –Ф–µ–≤—П—В—М —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–Њ–≤ —Б¬†–і–≤—Г–Љ—П —Г—З–∞—Б—В–љ–Є–Ї–∞–Љ–Є –±—Л–ї–Њ –њ—А–Њ–≤–µ–і–µ–љ–Њ –≤¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ–Њ–Љ –Ї–Њ–Љ–њ–ї–µ–Ї—Б–µ –≤¬†–Є–љ—В–µ—А–≤–∞–ї–µ –Љ–µ–ґ–і—Г —Г—В—А–Њ–Љ —Б—Г–±–±–Њ—В—Л –Є¬†—Г—В—А–Њ–Љ –њ–Њ–љ–µ–і–µ–ї—М–љ–Є–Ї–∞. –Ю–±–∞ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–ї–Є –≤¬†—Б—Г–±–±–Њ—В—Г –і–Њ 23 —З–∞—Б–Њ–≤, –∞¬†–Ј–∞—В–µ–Љ –Њ–і–Є–љ –Є–Ј¬†–љ–Є—Е —Б–њ–∞–ї –і–Њ 06 —З–∞—Б–Њ–≤ —Г—В—А–∞, –∞¬†–≤—В–Њ—А–Њ–є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–ї –і–Њ —В–Њ–≥–Њ –ґ–µ —З–∞—Б–∞. –°–ї–µ–і—Г—О—Й–Є–µ 24 —З–∞—Б–∞ –≤–Ї–ї—О—З–∞–ї–Є 12 —Г–ї—М—В—А–∞–і–Є–∞–љ–љ—Л—Е —Ж–Є–Ї–ї–Њ–≤ ¬Ђ—Б–Њ–љ¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬ї, —Б–Њ—Б—В–Њ—П—Й–Є—Е –Є–Ј¬†20-–Љ–Є–љ—Г—В–љ–Њ–є –њ–Њ–њ—Л—В–Ї–Є —Б–љ–∞ –Є¬†100 –Љ–Є–љ—Г—В –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П. –≠–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ—Л–µ –њ—А–Њ—В–Њ–Ї–Њ–ї—Л –±–Њ–ї–µ–µ –њ–Њ–і—А–Њ–±–љ–Њ –Њ–њ–Є—Б–∞–љ—Л –≤¬†–њ—А–µ–і—Л–і—Г—Й–µ–є –њ—Г–±–ї–Є–Ї–∞—Ж–Є–Є [26].
–Ч–∞–њ–Є—Б—М —Б–љ–∞ –≤–µ–ї–∞—Б—М —Б¬†–њ–Њ–Љ–Њ—Й—М—О –≤–Њ—Б—М–Љ–Є–Ї–∞–љ–∞–ї—М–љ–Њ–≥–Њ –њ–Њ–ї–Є–≥—А–∞—Д–∞ Medicor (EEG8S, Micromed, Hungary). –Ю–љ–∞ –≤–Ї–ї—О—З–∞–ї–∞ –Ј–∞–њ–Є—Б—М –њ–Њ–і–±–Њ—А–Њ–і–Њ—З–љ–Њ–є —Н–ї–µ–Ї—В—А–Њ–Љ–Є–Њ–≥—А–∞–Љ–Љ—Л (–Њ–і–Є–љ –Ї–∞–љ–∞–ї), —Н–ї–µ–Ї—В—А–Њ–Њ–Ї—Г–ї–Њ–≥—А–∞–Љ–Љ—Л (–і–≤–∞ –Ї–∞–љ–∞–ї–∞) –Є¬†–≠–≠–У (–њ—П—В—М –Ї–∞–љ–∞–ї–Њ–≤). –Ю–±—А–∞–±–∞—В—Л–≤–∞–ї–Є—Б—М –і–∞–љ–љ—Л–µ –Њ—В–≤–µ–і–µ–љ–Є—П Cz-A1 (–≤–µ—А—И–Є–љ–∞¬†–≥–Њ–ї–Њ–≤—Л –Є¬†–Љ–Њ—З–Ї–∞ –ї–µ–≤–Њ–≥–Њ —Г—Е–∞). –Ф–ї—П —Д–Є–Ї—Б–∞—Ж–Є–Є —Н–ї–µ–Ї—В—А–Њ–і–Њ–≤ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–∞—Б—М –њ–∞—Б—В–∞ Ten20 (Nicolet Biomedical, Madison, Wisconsin, USA). –Ч–∞–њ–Є—Б–Є —А–∞—Б—И–Є—Д—А–Њ–≤—Л–≤–∞–ї–Є—Б—М –і–≤—Г–Љ—П –љ–µ–Ј–∞–≤–Є—Б–Є–Љ—Л–Љ–Є ¬Ђ—Б–Ї–Њ—А–µ—А–∞–Љ–Є¬ї –њ–Њ¬†—Б—В–∞–љ–і–∞—А—В–љ—Л–Љ –Ї—А–Є—В–µ—А–Є—П–Љ [3]. –≠–≠–У-—Б–Є–≥–љ–∞–ї —Д–Є–ї—М—В—А–Њ–≤–∞–ї—Б—П, –Њ—Ж–Є—Д—А–Њ–≤—Л–≤–∞–ї—Б—П –Є¬†–Ј–∞–њ–Є—Б—Л–≤–∞–ї—Б—П –љ–∞¬†–і–Є—Б–Ї –Ї–Њ–Љ–њ—М—О—В–µ—А–∞. –£—З–∞—Б—В–Ї–Є –Ј–∞–њ–Є—Б–Є —Б¬†–∞—А—В–µ—Д–∞–Ї—В–∞–Љ–Є –≤—Л—П–≤–ї—П–ї–Є—Б—М –љ–∞¬†–њ—П—В–Є—Б–µ–Ї—Г–љ–і–љ—Л—Е –Є–љ—В–µ—А–≤–∞–ї–∞—Е, –Ј–∞—В–µ–Љ –∞–±—Б–Њ–ї—О—В–љ—Л–µ —Б–њ–µ–Ї—В—А–∞–ї—М–љ—Л–µ –Љ–Њ—Й–љ–Њ—Б—В–Є (¬µV2) –≤—Л—З–Є—Б–ї—П–ї–Є—Б—М –і–ї—П –≤—Б–µ—Е –Њ—Б—В–∞–ї—М–љ—Л—Е –њ—П—В–Є—Б–µ–Ї—Г–љ–і–љ—Л—Е –Є–љ—В–µ—А–≤–∞–ї–Њ–≤ —Б¬†–њ–Њ–Љ–Њ—Й—М—О –±—Л—Б—В—А–Њ–≥–Њ –њ—А–µ–Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –§—Г—А—М–µ. –Ф–∞–ї—М–љ–µ–є—И–∞—П —А–µ–і—Г–Ї—Ж–Є—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є –≤–Ї–ї—О—З–∞–ї–∞ —Г—Б—А–µ–і–љ–µ–љ–Є–µ –Ј–љ–∞—З–µ–љ–Є–є –љ–∞¬†—Б–Њ—Б–µ–і–љ–Є—Е —З–∞—Б—В–Њ—В–∞—Е –≤¬†–њ—А–µ–і–µ–ї–∞—Е –Њ–і–љ–Њ–≥–µ—А—Ж–µ–≤–Њ–≥–Њ –і–Є–∞–њ–∞–Ј–Њ–љ–∞ –Є¬†—Г—Б—А–µ–і–љ–µ–љ–Є–µ –њ–Њ–ї—Г—З–µ–љ–љ—Л—Е —В–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ –Ј–љ–∞—З–µ–љ–Є–є —Б–њ–µ–Ї—В—А–∞ –љ–∞¬†–Њ–і–љ–Њ–Љ–Є–љ—Г—В–љ—Л—Е –Є–љ—В–µ—А–≤–∞–ї–∞—Е. –Ч–љ–∞—З–µ–љ–Є—П –≤¬†–і–Є–∞–њ–∞–Ј–Њ–љ–µ –Њ—В¬†1 –і–Њ 16 –У—Ж –±—Л–ї–Є –Ї–Њ–љ–≤–µ—А—В–Є—А–Њ–≤–∞–љ—Л –≤¬†–љ–∞—В—Г—А–∞–ї—М–љ–Њ-–ї–Њ–≥–∞—А–Є—Д–Љ–Є—З–µ—Б–Ї—Г—О —И–Ї–∞–ї—Г –Є¬†–Њ–±—А–∞–±–Њ—В–∞–љ—Л –Љ–µ—В–Њ–і–Њ–Љ¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В. –Ы–Њ–≥-—В—А–∞–љ—Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–љ—Л–µ –Ј–љ–∞—З–µ–љ–Є—П –і–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ–Њ —Г—Б—А–µ–і–љ—П–ї–Є—Б—М –≤¬†–Є–љ—В–µ—А–≤–∞–ї–µ 4 –У—Ж, —З—В–Њ –њ—А–Є–Љ–µ—А–љ–Њ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г–µ—В –і–Є–∞–њ–∞–Ј–Њ–љ–∞–Љ –і–µ–ї—М—В–∞-, —В–µ—В–∞-, –∞–ї—М—Д–∞- –Є¬†—Б–Є–≥–Љ–∞-–∞–Ї—В–Є–≤–љ–Њ—Б—В–Є (1вАУ4, 5вАУ8, 9вАУ12 –Є¬†13вАУ16 –У—Ж —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ). –Ш–Ј¬†—Б–њ–µ–Ї—В—А–∞ —В–∞–Ї–Њ–≥–Њ —З–∞—Б—В–Њ—В–љ–Њ–≥–Њ –і–Є–∞–њ–∞–Ј–Њ–љ–∞ –Є–Ј–≤–ї–µ–Ї–∞–ї–Є—Б—М —З–µ—В—Л—А–µ¬†–≥–ї–∞–≤–љ—Л–µ –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л. –Ф–ї—П –њ–Њ–ї—Г—З–µ–љ–Є—П —З–µ—В—Л—А–µ—Е –Ј–љ–∞—З–µ–љ–Є–є¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В 16 –Ј–љ–∞—З–µ–љ–Є–є —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–є –Љ–Њ—Й–љ–Њ—Б—В–Є —Б—Г–Љ–Љ–Є—А–Њ–≤–∞–ї–Є—Б—М –њ–Њ—Б–ї–µ –Є—Е –Њ–њ—В–Є–Љ–∞–ї—М–љ–Њ–≥–Њ –≤–Ј–≤–µ—И–Є–≤–∞–љ–Є—П –≤¬†—Б–Њ–Њ—В–≤–µ—В—Б—В–≤–Є–Є —Б¬†–Є—Е –љ–∞–≥—А—Г–Ј–Ї–∞–Љ–Є –љ–∞¬†–Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Г. –Ф–µ—В–∞–ї—М–љ—Л–µ –Њ–њ–Є—Б–∞–љ–Є—П –Љ–µ—В–Њ–і–Њ–ї–Њ–≥–Є–Є –Њ–±—А–∞–±–Њ—В–Ї–Є –≠–≠–У-—Б–њ–µ–Ї—В—А–∞ –Љ–µ—В–Њ–і–Њ–Љ¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –±—Л–ї–Є –Њ–њ—Г–±–ї–Є–Ї–Њ–≤–∞–љ—Л —А–∞–љ–µ–µ [26вАУ33].
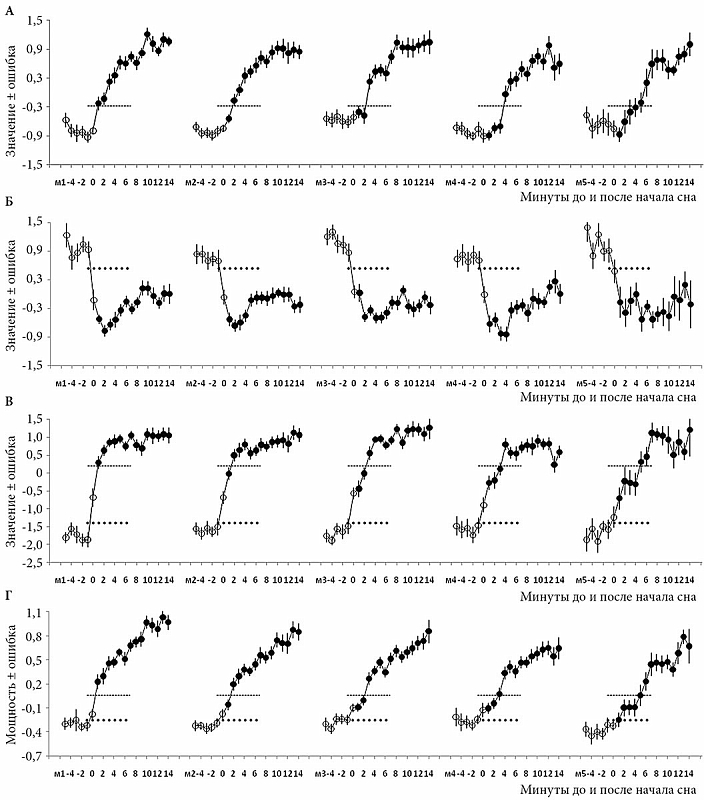
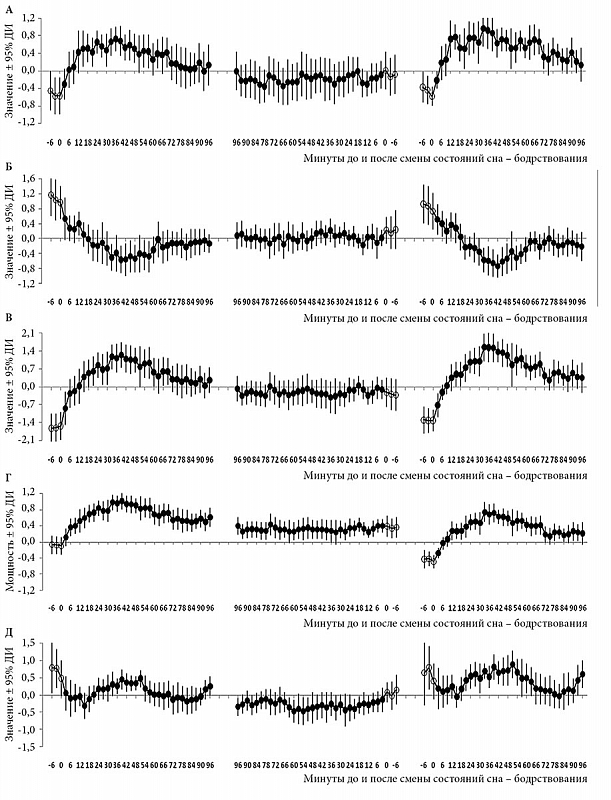
–†–µ–Ј—Г–ї—М—В–∞—В—Л —В–∞–Ї–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –і–∞–љ–љ—Л—Е –Њ—В–Њ–±—А–∞–ґ–µ–љ—Л –љ–∞¬†—А–Є—Б. 1вАУ3. –Я–µ—А–≤—Л–µ –і–≤–∞ —А–Є—Б—Г–љ–Ї–∞ –Њ—Б–љ–Њ–≤–∞–љ—Л –љ–∞¬†–і–∞–љ–љ—Л—Е –њ–Њ¬†–Љ–љ–Њ–≥–Њ–Ї—А–∞—В–љ—Л–Љ 20-–Љ–Є–љ—Г—В–љ—Л–Љ –њ–Њ–њ—Л—В–Ї–∞–Љ —Б–љ–∞ (–≤—В–Њ—А–Њ–є –Є¬†–њ–µ—А–≤—Л–є —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В—Л –Є¬†–≤—В–Њ—А–Њ–є —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ), –∞¬†—В—А–µ—В–Є–є —А–Є—Б—Г–љ–Њ–Ї –Њ—Б–љ–Њ–≤–∞–љ –љ–∞¬†–і–∞–љ–љ—Л—Е –і–≤—Г–Ї—А–∞—В–љ—Л—Е –Ј–∞–њ–Є—Б–µ–є –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞ (–њ–µ—А–≤—Л–є —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В). –Ъ—А–Є–≤—Л–µ –Ј–љ–∞—З–µ–љ–Є–є¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В (–У–Ъ1, –У–Ъ2, –У–Ъ1-2 –Є¬†–У–Ъ4) –Є¬†—В—А–∞–і–Є—Ж–Є–Њ–љ–љ–Њ–≥–Њ –≠–≠–У-–Є–љ–і–µ–Ї—Б–∞ (–Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М) —Г–Ї–∞–Ј–∞–љ—Л –ї–Є–±–Њ —Б¬†–Њ—И–Є–±–Ї–Њ–є —Б—А–µ–і–љ–µ–≥–Њ (–Њ—И–Є–±–Ї–∞; —А–Є—Б.¬†1), –ї–Є–±–Њ —Б¬†–і–Њ–≤–µ—А–Є—В–µ–ї—М–љ—Л–Љ –Є–љ—В–µ—А–≤–∞–ї–Њ–Љ (–Ф–Ш; —А–Є—Б.¬†2 –Є¬†3). –Ю–љ–Є —А–∞—Б—Б—З–Є—В–∞–љ—Л –њ—Г—В–µ–Љ —Г—Б—А–µ–і–љ–µ–љ–Є—П —Б–љ–∞—З–∞–ї–∞ –њ–Њ¬†–Ї–∞–ґ–і–Њ–Љ—Г —Г—З–∞—Б—В–љ–Є–Ї—Г —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞, –∞¬†–њ–Њ—В–Њ–Љ –і–ї—П –≤—Б–µ–є¬†–≥—А—Г–њ–њ—Л —Г—З–∞—Б—В–љ–Є–Ї–Њ–≤ –Њ–і–љ–Њ–≥–Њ (—А–Є—Б.¬†1 –Є¬†3) –Є–ї–Є –і–≤—Г—Е —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–Њ–≤ (—А–Є—Б.¬†2).
–†–µ–Ј—Г–ї—М—В–∞—В—Л
–†–Є—Б—Г–љ–Њ–Ї 1 –Є–ї–ї—О—Б—В—А–Є—А—Г–µ—В —А–µ–Ј—Г–ї—М—В–∞—В—Л –њ–Њ–і—А–Њ–±–љ–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–Є –Љ–µ–і–ї–µ–љ–љ–Њ–≥–Њ —Б–љ–∞ (¬Ђ–і—А–µ–Љ–Њ—В–љ—Л–є¬ї —Б–Њ–љ) –≤–Њ –≤—А–µ–Љ—П 20-–Љ–Є–љ—Г—В–љ—Л—Е –њ–Њ–њ—Л—В–Њ–Ї –Ј–∞—Б–љ—Г—В—М. –≠—В–∞ —Б—В–∞–і–Є—П –Њ–±—Л—З–љ–Њ —П–≤–ї—П–µ—В—Б—П –Ї—А–∞—В–Ї–Њ–≤—А–µ–Љ–µ–љ–љ—Л–Љ –њ–µ—А–µ—Е–Њ–і–љ—Л–Љ —Н—В–∞–њ–Њ–Љ –Љ–µ–ґ–і—Г –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ–Љ –Є¬†–±–Њ–ї–µ–µ¬†–≥–ї—Г–±–Њ–Ї–Њ–є —Б—В–∞–і–Є–µ–є ¬Ђ–љ–∞—Б—В–Њ—П—Й–µ–≥–Њ¬ї —Б–љ–∞ (–≤—В–Њ—А–Њ–є). –Ч–љ–∞—З–µ–љ–Є—П –њ–µ—А–≤–Њ–є (–У–Ъ1) –Є¬†–≤—В–Њ—А–Њ–є (–У–Ъ2)¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –≠–≠–У-—Б–њ–µ–Ї—В—А–∞, –∞¬†—В–∞–Ї–ґ–µ –Є—Е —А–∞–Ј–љ–Є—Ж—Л (–У–Ъ1-2) –Є¬†–Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є¬†вАУ —В—А–∞–і–Є—Ж–Є–Њ–љ–љ–Њ–≥–Њ –≠–≠–У-–Є–љ–і–µ–Ї—Б–∞¬†вАУ –±—Л–ї–Є —А–∞—Б—Б—З–Є—В–∞–љ—Л –і–ї—П –њ—П—В–Є —А–∞–Ј–љ—Л—Е –Ј–∞–і–µ—А–ґ–µ–Ї –љ–∞—З–∞–ї–∞ –≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞ (–Љ1вАУ–Љ5) –љ–∞¬†20-–Љ–Є–љ—Г—В–љ–Њ–Љ –Є–љ—В–µ—А–≤–∞–ї–µ –Њ—В¬†–Љ–Є–љ—Г—В—Л -5 –і–Њ +14 –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –љ—Г–ї–µ–≤–Њ–є¬†(0) –Љ–Є–љ—Г—В—Л –љ–∞—З–∞–ї–∞ —Б–љ–∞. –Ґ–Њ—З–µ—З–љ—Л–µ –Є¬†–њ—Г–љ–Ї—В–Є—А–љ—Л–µ –ї–Є–љ–Є–Є –њ–Њ–Ї–∞–Ј—Л–≤–∞—О—В –њ—А–Є–Љ–µ—А–љ—Г—О¬†–≥—А–∞–љ–Є—Ж—Г, –Њ—В–і–µ–ї—П—О—Й—Г—О –њ–µ—А–≤—Г—О —Б—В–∞–і–Є—О —Б–љ–∞ –Њ—В¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†–≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞. –Р–љ–∞–ї–Є–Ј —Б—В—А—Г–Ї—В—Г—А—Л –≠–≠–У-—Б–њ–µ–Ї—В—А–∞ –Љ–µ—В–Њ–і–Њ–Љ¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –≤—Л—П–≤–ї—П–µ—В –љ–µ–є—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї—Г—О –њ—А–Є—А–Њ–і—Г —Н—В–Њ–є —Б—В–∞–і–Є–Є. –Ю–љ–∞ —Б–ї—Г–ґ–Є—В ¬Ђ–љ–µ–є—В—А–∞–ї—М–љ–Њ–є –њ–Њ–ї–Њ—Б–Њ–є¬ї, —А–∞–Ј–і–µ–ї—П—О—Й–µ–є ¬Ђ–њ—А–Њ—В–Є–≤–Њ–±–Њ—А—Б—В–≤—Г—О—Й–Є–µ¬ї –і—А–∞–є–≤—Л –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†—Б–љ–∞. –Ч–љ–∞—З–µ–љ–Є–µ –≤—В–Њ—А–Њ–є¬†–≥–ї–∞–≤–љ–Њ–є –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л (–њ—А–µ–і–њ–Њ–ї–∞–≥–∞–µ–Љ–Њ–≥–Њ –Љ–∞—А–Ї–µ—А–∞ –і—А–∞–є–≤–∞ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П) —А–µ–Ј–Ї–Њ —Б–љ–Є–ґ–∞–µ—В—Б—П –≤—Б–µ–≥–і–∞ –љ–∞¬†–≥—А–∞–љ–Є—Ж–µ —Б¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ–Љ (—А–Є—Б.¬†1–С), –∞¬†–Ј–љ–∞—З–µ–љ–Є–µ –њ–µ—А–≤–Њ–є –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л (–њ—А–µ–і–њ–Њ–ї–∞–≥–∞–µ–Љ–Њ–≥–Њ –Љ–∞—А–Ї–µ—А–∞ –і—А–∞–є–≤–∞ —Б–љ–∞) —А–µ–Ј–Ї–Њ –≤–Њ–Ј—А–∞—Б—В–∞–µ—В –≤—Б–µ–≥–і–∞ –љ–∞¬†–≥—А–∞–љ–Є—Ж–µ —Б–Њ¬†–≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–µ–є (—А–Є—Б.¬†1–Р). –Э–∞¬†–≥—А–∞—Д–Є–Ї–µ —А–∞–Ј–љ–Є—Ж—Л –Ј–љ–∞—З–µ–љ–Є–є —Н—В–Є—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В (—А–Є—Б.¬†1–Т) —Е–Њ—А–Њ—И–Њ –≤–Є–і–љ—Л –і–≤–µ ¬Ђ—Б—В—Г–њ–µ–љ—М–Ї–Є¬ї, —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г—О—Й–Є–µ —Б—В–∞–і–Є–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†–≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞, —В–Њ–≥–і–∞ –Ї–∞–Ї –њ—А–Њ–Љ–µ–ґ—Г—В–Њ–Ї –Љ–µ–ґ–і—Г –љ–Є–Љ–Є –Ј–∞–љ–Є–Љ–∞–µ—В –њ–µ—А–≤–∞—П —Б—В–∞–і–Є—П. –£–≤–µ–ї–Є—З–µ–љ–Є–µ –Њ—В¬†–Њ–і–љ–Њ–є –і–Њ –њ—П—В–Є –Љ–Є–љ—Г—В –≤–µ–і–µ—В –Ї¬†—В–Њ—З–љ–Њ —В–∞–Ї–Њ–Љ—Г –ґ–µ —Г–≤–µ–ї–Є—З–µ–љ–Є—О –њ—А–Њ–Љ–µ–ґ—Г—В–Ї–∞ –Љ–µ–ґ–і—Г ¬Ђ—Б—В—Г–њ–µ–љ—П–Љ–Є¬ї. –Э–∞–њ—А–Њ—В–Є–≤, –µ—Б–ї–Є –∞–љ–∞–ї–Є–Ј–Є—А—Г–µ—В—Б—П –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М, –Є–Ј–Љ–µ—А–µ–љ–љ–∞—П –Ї–∞–Ї –Љ–Њ—Й–љ–Њ—Б—В—М –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –≤¬†–Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–Љ –і–Є–∞–њ–∞–Ј–Њ–љ–µ –≠–≠–У-—Б–њ–µ–Ї—В—А–∞ (—А–Є—Б.¬†1–У), —В–∞–Ї–Њ–є —З–µ—В–Ї–Њ–є ¬Ђ—Б—В—Г–њ–µ–љ—З–∞—В–Њ–є¬ї —Б—В—А—Г–Ї—В—Г—А—Л –љ–µ¬†–Њ–±–љ–∞—А—Г–ґ–Є–≤–∞–µ—В—Б—П. –Я–Њ—Б–ї–µ –Є–љ–Є—Ж–Є–∞—Ж–Є–Є —Б–љ–∞ —Н—В–Њ—В —В—А–∞–і–Є—Ж–Є–Њ–љ–љ–Њ –Є—Б–њ–Њ–ї—М–Ј—Г–µ–Љ—Л–є –Љ–∞—А–Ї–µ—А ¬Ђ–≤—Л–њ–ї–∞—В—Л –і–Њ–ї–≥–∞ —Б–љ–∞¬ї –њ—А–Њ—Б—В–Њ –њ–Њ—Б—В–µ–њ–µ–љ–љ–Њ ¬Ђ—А–∞—Б—В–∞–µ—В¬ї –њ–Њ¬†–Љ–µ—А–µ —Г–≥–ї—Г–±–ї–µ–љ–Є—П —Б–љ–∞.
–†–Є—Б—Г–љ–Њ–Ї 2 –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В —А–µ–Ј—Г–ї—М—В–∞—В—Л, –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ –і–ї—П —Б–Њ—Б—В–Њ—П–љ–Є—П –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†–≤—Б–µ—Е —В—А–µ—Е —Б—В–∞–і–Є–є –Љ–µ–і–ї–µ–љ–љ–Њ–≥–Њ —Б–љ–∞ –≤¬†–њ—А–µ–і–µ–ї–∞—Е 20-–Љ–Є–љ—Г—В–љ–Њ–≥–Њ –Є–љ—В–µ—А–≤–∞–ї–∞ –њ–Њ–њ—Л—В–Њ–Ї —Б–љ–∞. –Ч–љ–∞—З–µ–љ–Є—П¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В (–њ–µ—А–≤–Њ–є, –≤—В–Њ—А–Њ–є, –Є—Е —А–∞–Ј–љ–Є—Ж—Л –Є¬†—З–µ—В–≤–µ—А—В–Њ–є) –Є¬†—В—А–∞–і–Є—Ж–Є–Њ–љ–љ—Л–µ –Є–љ–і–µ–Ї—Б—Л –±—Л–ї–Є —А–∞—Б—Б—З–Є—В–∞–љ—Л –љ–∞¬†—В—А–µ—Е –і–≤—Г—Е—Б—В–∞–і–Є–є–љ—Л—Е –Є–љ—В–µ—А–≤–∞–ї–∞—Е (–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†–њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞, –њ–µ—А–≤–Њ–є –Є¬†–≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–є —Б–љ–∞ –Є¬†–≤—В–Њ—А–Њ–є –Є¬†—В—А–µ—В—М–µ–є —Б—В–∞–і–Є–є —Б–љ–∞). –Ґ—А–∞–і–Є—Ж–Є–Њ–љ–љ—Л–µ –≠–≠–У-–Є–љ–і–µ–Ї—Б—Л, —В–∞–Ї–Є–µ –Ї–∞–Ї –Љ–Њ—Й–љ–Њ—Б—В—М –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –Љ–Њ–Ј–≥–∞ –≤¬†–і–µ–ї—М—В–∞-, –∞–ї—М—Д–∞-, —В–µ—В–∞- –Є¬†—Б–Є–≥–Љ–∞-–і–Є–∞–њ–∞–Ј–Њ–љ–∞—Е –≠–≠–У-—Б–њ–µ–Ї—В—А–∞, –∞¬†—В–∞–Ї–ґ–µ –Є—Е –Њ—В–љ–Њ—И–µ–љ–Є—П, —В–∞–Ї–Є–µ –Ї–∞–Ї —А–∞–Ј–љ–Є—Ж–∞ –∞–ї—М—Д–∞- –Є¬†—В–µ—В–∞-–Љ–Њ—Й–љ–Њ—Б—В–Є, –Љ–Њ–≥—Г—В –і–Њ–≤–Њ–ї—М–љ–Њ –±—Л—Б—В—А–Њ –Љ–µ–љ—П—В—М—Б—П –љ–∞¬†–≥—А–∞–љ–Є—Ж–∞—Е –њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞, —Е–Њ—В—П –Є¬†–љ–µ —В–∞–Ї —Б—В—А–µ–Љ–Є—В–µ–ї—М–љ–Њ, –Ї–∞–Ї –Ј–љ–∞—З–µ–љ–Є—П –њ–µ—А–≤–Њ–є (–У–Ъ1) –Є¬†–≤—В–Њ—А–Њ–є (–У–Ъ2)¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –≠–≠–У-—Б–њ–µ–Ї—В—А–∞. –Ю–і–љ–∞–Ї–Њ —В–∞–Ї–Є–µ –Є–Ј–Љ–µ–љ–µ–љ–Є—П —В—А–∞–і–Є—Ж–Є–Њ–љ–љ—Л—Е –Є–љ–і–µ–Ї—Б–Њ–≤ –љ–µ¬†–Њ–±–љ–∞—А—Г–ґ–Є–≤–∞—О—В—Б—П –љ–∞¬†–≥—А–∞–љ–Є—Ж–µ –Љ–µ–ґ–і—Г –≤—В–Њ—А–Њ–є –Є¬†—В—А–µ—В—М–µ–є —Б—В–∞–і–Є—П–Љ–Є —Б–љ–∞, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г –Њ–љ–∞ –Њ–њ—А–µ–і–µ–ї—П–µ—В—Б—П –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ —Г—Б–ї–Њ–≤–љ—Л–Љ–Є –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ—Л–Љ–Є –Ї—А–Є—В–µ—А–Є—П–Љ–Є (—З–∞—Б—В–Њ—В—Л, –Ї–Њ–ї–Є—З–µ—Б—В–≤–∞ –Є¬†–∞–Љ–њ–ї–Є—В—Г–і—Л –Љ–µ–і–ї–µ–љ–љ—Л—Е –≤–Њ–ї–љ). –Т¬†–њ—А–Њ—В–Є–≤–Њ–≤–µ—Б —Н—В–Є–Љ –Є–љ–і–µ–Ї—Б–∞–Љ –Ј–љ–∞—З–µ–љ–Є–µ —З–µ—В–≤–µ—А—В–Њ–є (–У–Ъ4)¬†–≥–ї–∞–≤–љ–Њ–є –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л –≠–≠–У-—Б–њ–µ–Ї—В—А–∞ –љ–∞—З–Є–љ–∞–µ—В —Б–≤–Њ–є –±—Л—Б—В—А—Л–є —А–Њ—Б—В¬†–≥–і–µ-—В–Њ –≤–±–ї–Є–Ј–Є —Н—В–Њ–є¬†–≥—А–∞–љ–Є—Ж—Л (—А–Є—Б.¬†2–У). –Ш–Ј–Љ–µ–љ–µ–љ–Є–µ –Ј–љ–∞–Ї–∞ –Њ–і–љ–Њ–є –Є–Ј¬†–Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –≤¬†—А–∞–є–Њ–љ–µ¬†–≥—А–∞–љ–Є—Ж—Л –Љ–µ–ґ–і—Г —Б—В–∞–і–Є—П–Љ–Є (–≤—В–Њ—А–Њ–є¬†вАУ –і–ї—П¬†–≥—А–∞–љ–Є—Ж—Л –Љ–µ–ґ–і—Г –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ–Љ –Є¬†–њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–µ–є, –њ–µ—А–≤–Њ–є¬†вАУ –Љ–µ–ґ–і—Г –њ–µ—А–≤–Њ–є –Є¬†–≤—В–Њ—А–Њ–є —Б—В–∞–і–Є—П–Љ–Є –Є¬†—З–µ—В–≤–µ—А—В–Њ–є¬†вАУ –Љ–µ–ґ–і—Г –≤—В–Њ—А–Њ–є –Є¬†—В—А–µ—В—М–µ–є —Б—В–∞–і–Є—П–Љ–Є). –Я—А–Є—З–µ–Љ —В–∞–Ї–Њ–µ –Њ–њ—А–µ–і–µ–ї–µ–љ–Є–µ¬†–≥—А–∞–љ–Є—Ж—Л —Б—В–∞–і–Є–Є —Г–ґ–µ –љ–µ¬†—П–≤–ї—П–µ—В—Б—П —Г—Б–ї–Њ–≤–љ—Л–Љ, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г¬†–≥—А–∞–љ–Є—Ж–∞ —Г–Ї–∞–Ј—Л–≤–∞–µ—В –љ–∞¬†–љ–µ–є—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є –Њ—Б–Љ—Л—Б–ї–µ–љ–љ—Л–µ ¬Ђ—Б—В—Г–њ–µ–љ—З–∞—В—Л–µ¬ї –Є–Ј–Љ–µ–љ–µ–љ–Є—П —Г—А–Њ–≤–љ—П —Е—А–Њ–љ–Њ—А–µ–≥—Г–ї—П—В–Њ—А–љ—Л—Е –њ—А–Њ—Ж–µ—Б—Б–Њ–≤. –Т¬†—А–µ–Ј—Г–ї—М—В–∞—В–µ –њ–Њ—П–≤–ї—П–µ—В—Б—П –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –Њ–±—К–µ–Ї—В–Є–≤–љ–Њ –Њ—Ж–µ–љ–Є–≤–∞—В—М –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М –Ї–∞–ґ–і–Њ–є –Є–Ј¬†—Б—В–∞–і–Є–є –њ—Г—В–µ–Љ –њ—А–Є–Љ–µ–љ–µ–љ–Є—П –Ї—А–Є—В–µ—А–Є—П –њ–µ—А–µ—Б–µ—З–µ–љ–Є—П –љ—Г–ї—П –Ј–љ–∞—З–µ–љ–Є–µ–Љ –≤—В–Њ—А–Њ–є –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л (—А–Є—Б.¬†2–С), –њ–µ—А–≤–Њ–є –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л –Є–ї–Є –Є—Е —А–∞–Ј–љ–Є—Ж–µ–є (—А–Є—Б.¬†2–Р –Є¬†–Т) –Є¬†–Ј–љ–∞—З–µ–љ–Є–µ–Љ —З–µ—В–≤–µ—А—В–Њ–є –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л (—А–Є—Б.¬†2–У). –Ч–љ–∞—З–µ–љ–Є–µ –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л –Љ–µ–љ—П–µ—В—Б—П –ї–Є–±–Њ —Б¬†–њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ–≥–Њ –љ–∞¬†–Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ–Њ–µ (–У–Ъ2), –ї–Є–±–Њ —Б¬†–Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ–Њ–≥–Њ –љ–∞¬†–њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ–µ (–У–Ъ1, –У–Ъ1-2 –Є¬†–У–Ъ4).
–†–Є—Б—Г–љ–Њ–Ї 3 —Б—А–∞–≤–љ–Є–≤–∞–µ—В –Љ–∞—А–Ї–µ—А—Л –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ —А–µ–≥—Г–ї—П—Ж–Є–Є —Б–љ–∞ –≤¬†–љ–∞—З–∞–ї–µ –Є¬†–Ї–Њ–љ—Ж–µ –Њ–±—Л—З–љ–Њ–≥–Њ –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞. –°—З–Є—В–∞–µ—В—Б—П, —З—В–Њ –њ–Њ—Н—В–∞–њ–љ–Њ–µ —Б–љ–Є–ґ–µ–љ–Є–µ —Б—А–µ–і–љ–µ–≥–Њ —Г—А–Њ–≤–љ—П –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –≤¬†—З–µ—А–µ–і–µ —Г–ї—М—В—А–∞–і–Є–∞–љ–љ—Л—Е —Ж–Є–Ї–ї–Њ–≤ —Б–љ–∞ –Њ—В—А–∞–ґ–∞–µ—В –њ—А–Њ—Ж–µ—Б—Б ¬Ђ–≤—Л–њ–ї–∞—В—Л –і–Њ–ї–≥–∞ —Б–љ–∞¬ї, –љ–∞–Ї–Њ–њ–ї–µ–љ–љ–Њ–≥–Њ –≤–Њ –≤—А–µ–Љ—П –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П. –Ч–љ–∞—З–µ–љ–Є—П –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є (—А–Є—Б.¬†3–У) –Є¬†–Ј–љ–∞—З–µ–љ–Є—П¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В¬†вАУ –У–Ъ1 (—А–Є—Б.¬†3–Р), –У–Ъ2 (—А–Є—Б.¬†3–С), –У–Ъ1-2 (—А–Є—Б.¬†3–Т) –Є¬†–У–Ъ4 (—А–Є—Б.¬†3–Ф)¬†вАУ –±—Л–ї–Є —А–∞—Б—Б—З–Є—В–∞–љ—Л –і–ї—П 100-–Љ–Є–љ—Г—В–љ—Л—Е –Є–љ—В–µ—А–≤–∞–ї–Њ–≤ –≤¬†–љ–∞—З–∞–ї–µ –Є¬†–Ї–Њ–љ—Ж–µ –Њ–±—Л—З–љ–Њ–≥–Њ –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞. –Ф–ї—П —В–Њ–≥–Њ —З—В–Њ–±—Л –Є–ї–ї—О—Б—В—А–Є—А–Њ–≤–∞—В—М —А–∞–Ј–ї–Є—З–Є—П –Љ–µ–ґ–і—Г —Н—В–Є–Љ–Є –Є–љ—В–µ—А–≤–∞–ї–∞–Љ–Є, –±—Л–ї–∞ –і–Њ–њ–Њ–ї–љ–Є—В–µ–ї—М–љ–Њ —А–∞—Б—Б—З–Є—В–∞–љ–∞ —А–∞–Ј–љ–Є—Ж–∞ –Љ–µ–ґ–і—Г –Ј–љ–∞—З–µ–љ–Є—П–Љ–Є, –њ–Њ–ї—Г—З–µ–љ–љ—Л–Љ–Є –≤¬†–љ–∞—З–∞–ї–µ –Є¬†–Ї–Њ–љ—Ж–µ —Б–љ–∞. –°—Е–Њ–і—Б—В–≤–Њ –Љ–µ–ґ–і—Г –і–Є–љ–∞–Љ–Є–Ї–Њ–є –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є (—А–Є—Б.¬†3–У) –Є¬†–і–Є–љ–∞–Љ–Є–Ї–Њ–є –У–Ъ1-2 (—А–Є—Б.¬†3–Т) –љ–∞¬†–≤—Б–µ—Е —В—А–µ—Е –Є–љ—В–µ—А–≤–∞–ї–∞—Е –њ–Њ–Ј–≤–Њ–ї—П–µ—В –њ—А–µ–і–њ–Њ–ї–∞–≥–∞—В—М, —З—В–Њ –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –Њ—В—А–∞–ґ–∞–µ—В —Б—Г–Љ–Љ–∞—А–љ—Л–є –≤–Ї–ї–∞–і –і—А–∞–є–≤–Њ–≤ —Б–љ–∞ –Є¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –≤¬†–њ—А–Њ—Ж–µ—Б—Б —А–µ–≥—Г–ї—П—Ж–Є–Є ¬Ђ–≤—Л–њ–ї–∞—В—Л –і–Њ–ї–≥–∞ —Б–љ–∞¬ї. –Ф–ї—П —В–Њ–≥–Њ —З—В–Њ–±—Л –Є–љ–Є—Ж–Є–Є—А–Њ–≤–∞—В—М —Н—В–Њ—В –њ—А–Њ—Ж–µ—Б—Б –≤¬†—Д–Њ—А–Љ–µ —А–Њ—Б—В–∞ –У–Ъ1 (–њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ –њ—А–µ–і—Б—В–∞–≤–ї—П—О—Й–µ–є –і—А–∞–є–≤ —Б–љ–∞) —Б¬†–Љ–Є–љ–Є–Љ–∞–ї—М–љ–Њ–≥–Њ –Ј–љ–∞—З–µ–љ–Є—П –љ–∞¬†–≥—А–∞–љ–Є—Ж–µ –њ–µ—А–≤–Њ–є –Є¬†–≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–є –і–Њ –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ–≥–Њ —Г—А–Њ–≤–љ—П –≤¬†—В—А–µ—В—М–µ–є —Б—В–∞–і–Є–Є –њ–µ—А–≤–Њ–≥–Њ —Ж–Є–Ї–ї–∞ —Б–љ–∞ (—А–Є—Б.¬†3–Р), –Ј–љ–∞—З–µ–љ–Є–µ –У–Ъ2 (–њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ –њ—А–µ–і—Б—В–∞–≤–ї—П—О—Й–µ–є –і—А–∞–є–≤ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П) –і–Њ–ї–ґ–љ–Њ –Ј–∞—А–∞–љ–µ–µ —А–µ–Ј–Ї–Њ —Г–Љ–µ–љ—М—И–Є—В—М—Б—П –љ–∞¬†–≥—А–∞–љ–Є—Ж–µ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†–њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–Є (—А–Є—Б.¬†3–С). –Э–∞—З–∞–ї–Њ –њ–Њ—Б–ї–µ–і—Г—О—Й–µ–≥–Њ —Н—В–∞–њ–∞ –љ–∞–Є–±–Њ–ї–µ–µ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ–є ¬Ђ–≤—Л–њ–ї–∞—В—Л¬ї —Б–Њ–≤–њ–∞–і–∞–µ—В —Б¬†–љ–∞—З–∞–ї–Њ–Љ —В—А–µ—В—М–µ–є —Б—В–∞–і–Є–Є —Б–љ–∞. –Ґ–∞–Ї–Њ–µ –љ–∞—З–∞–ї–Њ –Љ–∞—А–Ї–Є—А—Г–µ—В—Б—П –љ–∞—З–∞–ї–Њ–Љ –њ–Њ–і—К–µ–Љ–∞ –У–Ъ4 —Б¬†–Љ–Є–љ–Є–Љ–∞–ї—М–љ–Њ–≥–Њ –Ј–љ–∞—З–µ–љ–Є—П –љ–∞¬†–Є–љ—В–µ—А–≤–∞–ї–µ –≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–Є (—А–Є—Б.¬†3–Ф). –≠—В–Њ –љ–∞—З–∞–ї–Њ —В–∞–Ї–ґ–µ —Б–Њ–≤–њ–∞–і–∞–µ—В —Б¬†–љ–∞—З–∞–ї–Њ–Љ –і–∞–ї—М–љ–µ–є—И–µ–≥–Њ —Б–њ–∞–і–∞ –У–Ъ2 (—А–Є—Б.¬†3–С), —В–Њ–≥–і–∞ –Ї–∞–Ї –Ј–љ–∞—З–µ–љ–Є–µ –У–Ъ1 –Њ—Б—В–∞–µ—В—Б—П —Б—В–∞–±–Є–ї—М–љ–Њ –≤—Л—Б–Њ–Ї–Є–Љ –і–Њ —Б–∞–Љ–Њ–≥–Њ –Ї–Њ–љ—Ж–∞ —В—А–µ—В—М–µ–є —Б—В–∞–і–Є–Є (—А–Є—Б.¬†3–Р). –Т–Њ –≤—А–µ–Љ—П –њ–Њ—Б–ї–µ–і—Г—О—Й–µ–≥–Њ –њ–µ—А–µ—Е–Њ–і–∞ –Њ—В¬†–Љ–µ–і–ї–µ–љ–љ–Њ–≥–Њ (–Њ—А—В–Њ–і–Њ–Ї—Б–∞–ї—М–љ–Њ–≥–Њ) —Б–љ–∞ –Ї¬†–њ–Њ—Б–ї–µ–і—Г—О—Й–µ–є —Д–∞–Ј–µ –±—Л—Б—В—А–Њ–≥–Њ (–њ–∞—А–∞–і–Њ–Ї—Б–∞–ї—М–љ–Њ–≥–Њ) —Б–љ–∞ –≤¬†–Ї–Њ–љ—Ж–µ –њ–µ—А–≤–Њ–≥–Њ —Г–ї—М—В—А–∞–і–Є–∞–љ–љ–Њ–≥–Њ —Ж–Є–Ї–ї–∞ —Б–љ–∞ –≤—Б–µ –Ј–љ–∞—З–µ–љ–Є—П –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –љ–∞—З–Є–љ–∞—О—В ¬Ђ–і—А—Г–ґ–љ–Њ¬ї —Б—В—А–µ–Љ–Є—В—М—Б—П –Ї¬†–Ј–љ–∞—З–µ–љ–Є—О, –±–ї–Є–Ј–Ї–Њ–Љ—Г –Ї¬†–љ—Г–ї—О. –Э–∞–Ї–Њ–љ–µ—Ж, –њ–Њ—Б–ї–µ —З–µ—А–µ–і—Л —Г–ї—М—В—А–∞–і–Є–∞–љ–љ—Л—Е —Ж–Є–Ї–ї–Њ–≤ —Б–љ–∞ –њ—А–Є–Љ–µ—А–љ–Њ —В–∞–Ї–Њ–µ –ґ–µ —Б–Њ–Њ—В–≤–µ—В—Б—В–≤—Г—О—Й–µ–µ –љ—Г–ї—О –Ј–љ–∞—З–µ–љ–Є–µ –і–Њ—Б—В–Є–≥–∞–µ—В—Б—П –≤—Б–µ–Љ–Є –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В–∞–Љ–Є –љ–∞¬†—Д–Є–љ–∞–ї—М–љ–Њ–Љ —Н—В–∞–њ–µ –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞ (—А–Є—Б.¬†3–Р, –С, –Ф), –Ї–Њ–≥–і–∞ ¬Ђ–і–Њ–ї–≥ —Б–љ–∞¬ї –њ–Њ–ї–љ–Њ—Б—В—М—О ¬Ђ–≤—Л–њ–ї–∞—З–µ–љ¬ї.
–Ю–±—Б—Г–ґ–і–µ–љ–Є–µ —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤
–Э–∞—Г—З–љ–Њ–µ –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Њ–µ –Њ–њ–Є—Б–∞–љ–Є–µ —Б–љ–∞ –љ–µ–Є–Ј–Љ–µ–љ–љ–Њ –≤–Ї–ї—О—З–∞–µ—В —Б–Њ–Ј–і–∞–љ–Є–µ¬†–≥–Є–њ–љ–Њ–≥—А–∞–Љ–Љ—Л, –Ї–Њ—В–Њ—А–∞—П —Б–≤–Њ–і–Є—В –≤—Б–µ —А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–љ—Л–µ –њ–∞—В—В–µ—А–љ—Л –≠–≠–У- –Є¬†–і—А—Г–≥–Є—Е –≤—Е–Њ–і—П—Й–Є—Е –≤¬†–њ–Њ–ї–Є—Б–Њ–Љ–љ–Њ–≥—А–∞–Љ–Љ—Г —Б–Є–≥–љ–∞–ї–Њ–≤ –Ї¬†–љ–µ–±–Њ–ї—М—И–Њ–Љ—Г —З–Є—Б–ї—Г –і–∞-–Є–ї–Є-–љ–µ—В-–њ–µ—А–µ–Љ–µ–љ–љ—Л—Е, –Њ–±–Њ–Ј–љ–∞—З–µ–љ–љ—Л—Е –Ї–∞–Ї –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ –Є–ї–Є –Њ–њ—А–µ–і–µ–ї–µ–љ–љ–∞—П —Д–∞–Ј–∞ –Є–ї–Є —Б—В–∞–і–Є—П —Б–љ–∞. –Э–∞¬†–њ—А–Њ—В—П–ґ–µ–љ–Є–Є –≤—Б–µ–є –Є—Б—В–Њ—А–Є–Є –љ–∞—Г—З–љ–Њ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П —Б–љ–∞ –Њ–і–љ–Њ–є –Є–Ј¬†–љ–µ—А–µ—И–µ–љ–љ—Л—Е –љ–∞—Г—З–љ—Л—Е –њ—А–Њ–±–ї–µ–Љ –Њ—Б—В–∞–µ—В—Б—П –Ј–∞–і–∞—З–∞ –Ј–∞–Љ–µ–љ—Л —В–∞–Ї–Њ–≥–Њ —Д–Њ—А–Љ–∞–ї—М–љ–Њ–≥–Њ –њ–Њ–і—Е–Њ–і–∞ –±–Њ–ї–µ–µ –µ—Б—В–µ—Б—В–≤–µ–љ–љ—Л–Љ –Љ–µ—В–Њ–і–Њ–Љ, –Ї–Њ—В–Њ—А—Л–є –њ—А–µ–і–њ–Њ–ї–∞–≥–∞–µ—В –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –љ–µ–є—А–Њ—Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є –Њ—Б–Љ—Л—Б–ї–µ–љ–љ–Њ–є –і–µ–Љ–∞—А–Ї–∞—Ж–Є–Є¬†–≥—А–∞–љ–Є—Ж –Љ–µ–ґ–і—Г —Б—В–∞–і–Є—П–Љ–Є —Б–љ–∞. –Ґ–∞–Ї–Њ–є –Љ–µ—В–Њ–і –љ—Г–ґ–љ–Њ –љ–µ¬†—В–Њ–ї—М–Ї–Њ —А–∞–Ј—А–∞–±–Њ—В–∞—В—М, –љ–Њ¬†–Є¬†—В–µ–Њ—А–µ—В–Є—З–µ—Б–Ї–Є –Њ–±–Њ—Б–љ–Њ–≤–∞—В—М. –Ч–∞ –Њ—Б–љ–Њ–≤—Г –њ—А–µ–і–µ–ї—М–љ–Њ –њ—А–Њ—Б—В–Њ–≥–Њ, –љ–Њ¬†–≤¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П –Њ–±—К–µ–Ї—В–Є–≤–љ–Њ–≥–Њ –Є¬†—В–µ–Њ—А–µ—В–Є—З–µ—Б–Ї–Є –Њ–±–Њ—Б–љ–Њ–≤–∞–љ–љ–Њ–≥–Њ –њ–Њ–і—Е–Њ–і–∞ –Ї¬†–Њ–њ—А–µ–і–µ–ї–µ–љ–Є—О¬†–≥—А–∞–љ–Є—Ж –Љ–µ–ґ–і—Г —Б—В–∞–і–Є—П–Љ–Є —Б–љ–∞ –Љ–Њ–ґ–љ–Њ –≤–Ј—П—В—М —В–µ–Њ—А–µ—В–Є—З–µ—Б–Ї–Є–µ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є—П –Њ–±¬†—Г—З–∞—Б—В–Є–Є –Њ–њ–њ–Њ–љ–Є—А—Г—О—Й–Є—Е —Е—А–Њ–љ–Њ–љ–µ–є—А–Њ–±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤ —А–µ–≥—Г–ї—П—Ж–Є–Є —Б–Љ–µ–љ—Л —Б–Њ—Б—В–Њ—П–љ–Є–є –Є¬†—Б—Г–±—Б–Њ—Б—В–Њ—П–љ–Є–є —Б–љ–∞¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П. –Ґ–∞–Ї—Г—О –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –њ–Њ–і—В–≤–µ—А–ґ–і–∞—О—В —А–µ–Ј—Г–ї—М—В–∞—В—Л –∞–љ–∞–ї–Є–Ј–∞ –і–Є–љ–∞–Љ–Є–Ї–Є –Ј–љ–∞—З–µ–љ–Є–є¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В, –Ї–Њ—В–Њ—А—Л–µ, –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ, —П–≤–ї—П—О—В—Б—П –≠–≠–У-–Љ–∞—А–Ї–µ—А–∞–Љ–Є –∞–љ—В–∞–≥–Њ–љ–Є—Б—В–Є—З–µ—Б–Ї–Є—Е –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є–є –њ—А–Њ—Ж–µ—Б—Б–Њ–≤-–њ—А–Њ–Љ–Њ—В–Њ—А–Њ–≤ —Б–љ–∞ –Є¬†–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –≤¬†—Д–∞–Ј–∞—Е –њ–µ—А–µ—Е–Њ–і–∞ —Б–Њ—Б—В–Њ—П–љ–Є–є –Є¬†—Б—Г–±—Б–Њ—Б—В–Њ—П–љ–Є–є –Њ—В¬†–Њ–і–љ–Њ–≥–Њ –Ї¬†–і—А—Г–≥–Њ–Љ—Г –≤¬†—Б—Г—В–Њ—З–љ–Њ–Љ —Ж–Є–Ї–ї–µ ¬Ђ—Б–Њ–љ¬†вАУ–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬ї –Є¬†—Г–ї—М—В—А–∞–і–Є–∞–љ–љ—Л—Е —Ж–Є–Ї–ї–∞—Е —Б–љ–∞.
–°–ї–µ–і—Г–µ—В –Њ–ґ–Є–і–∞—В—М, —З—В–Њ –∞–љ–∞–ї–Є–Ј –≠–≠–У-—Б–њ–µ–Ї—В—А–∞ –Љ–µ—В–Њ–і–Њ–Љ¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –Љ–Њ–ґ–µ—В –±—Л—В—М –њ–Њ–ї–µ–Ј–µ–љ –і–ї—П –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –≤¬†–Њ–±–ї–∞—Б—В–Є –Љ–µ–і–Є—Ж–Є–љ—Л —Б–љ–∞, –њ—А–Є—З–µ–Љ –љ–µ¬†—В–Њ–ї—М–Ї–Њ —В–µ–Љ, —З—В–Њ –њ–Њ–Ј–≤–Њ–ї–Є—В —Г–Љ–µ–љ—М—И–Є—В—М —Б—Г–±—К–µ–Ї—В–Є–≤–љ—Л–µ –≤–ї–Є—П–љ–Є—П ¬Ђ—Б–Ї–Њ—А–µ—А–∞¬ї –љ–∞¬†—А–µ–Ј—Г–ї—М—В–∞—В—Л –Њ–њ–Є—Б–∞–љ–Є—П —Б–љ–∞ –≤¬†–≤–Є–і–µ¬†–≥–Є–њ–љ–Њ–≥—А–∞–Љ–Љ—Л. –°–ї–µ–і—Г–µ—В –Њ–ґ–Є–і–∞—В—М, —З—В–Њ —В–∞–Ї–Њ–є –∞–љ–∞–ї–Є–Ј –Љ–Њ–ґ–µ—В –њ–Њ–Ј–≤–Њ–ї–Є—В—М –ї—Г—З—И–µ –њ–Њ–љ—П—В—М –њ—А–Є—З–Є–љ—Л –љ–∞—А—Г—И–µ–љ–Є–є —Б–љ–∞. –Э–∞–њ—А–Є–Љ–µ—А, –≤¬†–Њ–і–љ–Њ–є –Є–Ј¬†–љ–∞—И–Є—Е –њ—Г–±–ї–Є–Ї–∞—Ж–Є–є [33] –Љ—Л –Њ–њ–Є—Б–∞–ї–Є –≤–Њ–Ј—А–∞—Б—В–љ—Л–µ –Є–Ј–Љ–µ–љ–µ–љ–Є—П –Ј–љ–∞—З–µ–љ–Є–є –і–≤—Г—Е –њ–µ—А–≤—Л—Е¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В. –Ю–Ї–∞–Ј–∞–ї–Њ—Б—М, —З—В–Њ –њ—А–Њ—Ж–µ—Б—Б –љ–Њ—А–Љ–∞–ї—М–љ–Њ–≥–Њ —Б—В–∞—А–µ–љ–Є—П —Б–љ–∞, –љ–∞–Є–±–Њ–ї–µ–µ —А–∞–љ–љ–Є–Љ –Є¬†–Њ—З–µ–≤–Є–і–љ—Л–Љ –њ—А–Є–Ј–љ–∞–Ї–Њ–Љ –Ї–Њ—В–Њ—А–Њ–≥–Њ —П–≤–ї—П–µ—В—Б—П —Б–љ–Є–ґ–µ–љ–Є–µ –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –Є¬†–Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–≥–Њ —Б–љ–∞, –Љ–Њ–ґ–љ–Њ –Њ–±—К—П—Б–љ–Є—В—М –љ–µ¬†—В–Њ–ї—М–Ї–Њ –Њ—Б–ї–∞–±–ї–µ–љ–Є–µ–Љ –і—А–∞–є–≤–∞ —Б–љ–∞ (—Б–љ–Є–ґ–µ–љ–Є–µ–Љ –Ј–љ–∞—З–µ–љ–Є—П –њ–µ—А–≤–Њ–є –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л), –љ–Њ¬†–Є¬†—Г—Б–Є–ї–µ–љ–Є–µ–Љ –і—А–∞–є–≤–∞ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П (–њ–Њ–≤—Л—И–µ–љ–Є–µ–Љ –Ј–љ–∞—З–µ–љ–Є—П –≤—В–Њ—А–Њ–є –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л) –Є–Ј-–Ј–∞ –і–µ–Ј–Є–љ–≥–Є–±–Є—А—Г—О—Й–µ–≥–Њ –≤–ї–Є—П–љ–Є—П –Њ—Б–ї–∞–±–ї–µ–љ–Є—П –і—А–∞–є–≤–∞ —Б–љ–∞. –≠—В–Њ—В —А–µ–Ј—Г–ї—М—В–∞—В –њ–Њ–Ј–≤–Њ–ї—П–µ—В –ї–µ–≥–Ї–Њ –Њ—В–≤–µ—В–Є—В—М –љ–∞¬†–≤–Њ–њ—А–Њ—Б: ¬Ђ–Я–Њ—З–µ–Љ—Г —Б—В–∞—А–µ–љ–Є–µ —Б–Њ–њ—А–Њ–≤–Њ–ґ–і–∞–µ—В—Б—П –љ–µ¬†—В–Њ–ї—М–Ї–Њ –љ–∞—А—Г—И–µ–љ–Є—П–Љ–Є —Ж–Є–Ї–ї–Є—З–љ–Њ—Б—В–Є —Б–Њ—Б—В–Њ—П–љ–Є–є —Б–љ–∞¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†–љ–∞—А—Г—И–µ–љ–Є—П–Љ–Є –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞ (—З–∞—Б—В—Л–Љ–Є –Є¬†—А–∞–љ–љ–Є–Љ–Є –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П–Љ–Є), –љ–Њ¬†–Є¬†–њ–Њ—П–≤–ї–µ–љ–Є–µ–Љ –љ–µ–Ї–Њ—В–Њ—А—Л—Е –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤ —Г¬†–њ–Њ–ґ–Є–ї—Л—Е –ї—О–і–µ–є –њ–µ—А–µ–і –Љ–Њ–ї–Њ–і—Л–Љ–Є –ї—О–і—М–Љ–Є –≤¬†—Г—Б–ї–Њ–≤–Є—П—Е –ґ–Є–Ј–љ–Є –≤¬†—Б–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–Љ –Њ–±—Й–µ—Б—В–≤–µ?¬ї –Т–µ—А–Њ—П—В–љ–µ–µ –≤—Б–µ–≥–Њ, –Є–Љ–µ–љ–љ–Њ —Г—Б–Є–ї–µ–љ–Є–µ –і—А–∞–є–≤–∞ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –њ–Њ–Ј–≤–Њ–ї—П–µ—В –њ–Њ–ґ–Є–ї—Л–Љ –ї—О–і—П–Љ —А–µ–ґ–µ –Ј–∞—Б—Л–њ–∞—В—М –љ–µ–њ—А–Њ–Є–Ј–≤–Њ–ї—М–љ–Њ –≤¬†—Б–ї—Г—З–∞–µ –Њ—Б—В—А–Њ–є –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ—Б—В–Є –±–Њ–і—А—Б—В–≤–Њ–≤–∞—В—М –њ–Њ—Б–ї–µ –і–ї–Є—В–µ–ї—М–љ–Њ–≥–Њ –њ–µ—А–Є–Њ–і–∞ –ї–Є—И–µ–љ–Є—П —Б–љ–∞ –Є–ї–Є ¬Ђ–љ–µ–і–Њ—Б—Л–њ–∞¬ї. –≠—В–Њ—В —А–µ–Ј—Г–ї—М—В–∞—В —В–∞–Ї–ґ–µ –њ–Њ–Ј–≤–Њ–ї—П–µ—В –Њ–±—К—П—Б–љ–Є—В—М –і–Њ–≤–Њ–ї—М–љ–Њ –њ—А–Њ—Б—В—Л–Љ–Є –њ—А–Є—З–Є–љ–љ–Њ-—Б–ї–µ–і—Б—В–≤–µ–љ–љ—Л–Љ–Є —Б–≤—П–Ј—П–Љ–Є –љ–∞–±–ї—О–і–∞–µ–Љ—Л–µ —Б¬†–≤–Њ–Ј—А–∞—Б—В–Њ–Љ –њ–∞—В–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Є–Ј–Љ–µ–љ–µ–љ–Є—П —Б–љ–∞. –Э–∞–њ—А–Є–Љ–µ—А, —В–∞–Ї–Є–µ –љ–∞¬†–њ–µ—А–≤—Л–є –≤–Ј–≥–ї—П–і –і–∞–ї–µ–Ї–Њ –љ–µ¬†–Њ—З–µ–≤–Є–і–љ—Л–Љ –Њ–±—А–∞–Ј–Њ–Љ —Б–≤—П–Ј–∞–љ–љ—Л–µ –і—А—Г–≥ —Б¬†–і—А—Г–≥–Њ–Љ —Б–Є–Љ–њ—В–Њ–Љ—Л, –Ї–∞–Ї –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ—Л–є —Г—А–Њ–≤–µ–љ—М –і–љ–µ–≤–љ–Њ–≥–Њ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П, –љ–µ–ґ–µ–ї–∞—В–µ–ї—М–љ–∞—П –і–љ–µ–≤–љ–∞—П —Б–Њ–љ–ї–Є–≤–Њ—Б—В—М, –њ–Њ—П–≤–ї–µ–љ–Є–µ –њ—А–Є–≤—Л—З–Ї–Є –і—А–µ–Љ–∞—В—М –і–љ–µ–Љ, –љ–µ—Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –±—Л—Б—В—А–Њ –Ј–∞—Б—Л–њ–∞—В—М, –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–∞—П –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В—М –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞, –љ–µ—Г–і–Њ–≤–ї–µ—В–≤–Њ—А–µ–љ–љ–Њ—Б—В—М —В–∞–Ї–Є–Љ —Б–љ–Њ–Љ, –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–љ–Њ—Б—В—М –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞, —З–∞—Б—В—Л–µ –љ–Њ—З–љ—Л–µ –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П, –љ–µ–ґ–µ–ї–∞—В–µ–ї—М–љ—Л–µ –њ—А–Њ–±—Г–ґ–і–µ–љ–Є—П —А–∞–љ–љ–Є–Љ —Г—В—А–Њ–Љ [33].
–Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –∞–љ–∞–ї–Є–Ј –≠–≠–У-—Б–њ–µ–Ї—В—А–∞ –Љ–µ—В–Њ–і–Њ–Љ¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –њ–Њ–Ј–≤–Њ–ї–Є—В —В–Њ—З–љ–µ–µ –Њ—Ж–µ–љ–Є–≤–∞—В—М —Г—З–∞—Б—В–Є–µ –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ —А–µ–≥—Г–ї—П—Ж–Є–Є —Ж–Є–Ї–ї–∞ ¬Ђ—Б–Њ–љ¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬ї –≤¬†—А–∞–Ј–ї–Є—З–љ—Л—Е —А–∞—Б—Б—В—А–Њ–є—Б—В–≤–∞—Е –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞ –Є¬†—Ж–Є–Ї–ї–∞ ¬Ђ—Б–Њ–љ¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬ї. –Э–∞–њ—А–Є–Љ–µ—А, –µ—Б–ї–Є –Љ–∞—А–Ї–µ—А–Њ–Љ –љ–∞—З–∞–ї–∞ –њ—А–Њ—Ж–µ—Б—Б–∞ ¬Ђ–≤—Л–њ–ї–∞—В—Л –і–Њ–ї–≥–∞ —Б–љ–∞¬ї —П–≤–ї—П–µ—В—Б—П –љ–µ¬†–љ–∞—З–∞–ї–Њ —А–Њ—Б—В–∞ –Љ–µ–і–ї–µ–љ–љ–Њ–≤–Њ–ї–љ–Њ–≤–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є —Б—А–∞–Ј—Г –њ–Њ—Б–ї–µ —Б–Љ–µ–љ—Л –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П —Б–љ–Њ–Љ, –∞¬†–љ–∞—З–∞–ї–Њ —А–Њ—Б—В–∞ –Ј–љ–∞—З–µ–љ–Є—П –њ–µ—А–≤–Њ–є –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В—Л –љ–∞¬†–≥—А–∞–љ–Є—Ж–µ –њ–µ—А–≤–Њ–є –Є¬†–≤—В–Њ—А–Њ–є —Б—В–∞–і–Є–є —Б–љ–∞ [30, 31], —В–Њ –њ—А–Є —А–∞—Б—З–µ—В–µ —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞ –љ—Г–ґ–љ–Њ –љ–µ¬†–і–Њ–±–∞–≤–ї—П—В—М –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В—М –њ–µ—А–≤–Њ–є —Б—В–∞–і–Є–Є –Ї¬†–Њ–±—Й–µ–є –і–ї–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Б–љ–∞, –∞¬†–≤—Л—З–Є—В–∞—В—М –µ–µ. –Ш–љ—В–µ—А–≤–∞–ї—Л ¬Ђ–і—А–µ–Љ–Њ—В–љ–Њ–≥–Њ¬ї —Б–љ–∞ –Ї–Њ—А–Њ—В–Ї–Є —В–Њ–ї—М–Ї–Њ –њ—А–Є –љ–Њ—А–Љ–∞–ї—М–љ–Њ–Љ —Б–љ–µ, –љ–Њ¬†–Њ–љ–Є —Г–≤–µ–ї–Є—З–µ–љ—Л –≤¬†–і–≤–∞ –Є¬†–±–Њ–ї–µ–µ —А–∞–Ј —Г¬†–њ–Њ–ґ–Є–ї—Л—Е –ї—О–і–µ–є –Є¬†–њ—А–Є –Љ–љ–Њ–≥–Є—Е —А–∞—Б—Б—В—А–Њ–є—Б—В–≤–∞—Е —Б–љ–∞ (–±–µ—Б—Б–Њ–љ–љ–Є—Ж–µ, –∞–њ–љ–Њ—Н —Б–љ–∞ –Є¬†–і—А.). –Я–Њ—Н—В–Њ–Љ—Г –Љ–Њ–ґ–љ–Њ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ —Б—Г–±—К–µ–Ї—В–Є–≤–љ—Л–µ –Њ—Й—Г—Й–µ–љ–Є—П –љ–∞—А—Г—И–µ–љ–Є–є –љ–Њ—З–љ–Њ–≥–Њ —Б–љ–∞, —Е–∞—А–∞–Ї—В–µ—А–љ—Л–µ –і–ї—П —Н—В–Є—Е –ї—О–і–µ–є –Є¬†–њ—А–Є —Н—В–Є—Е —А–∞—Б—Б—В—А–Њ–є—Б—В–≤–∞—Е, –љ–µ¬†—В–∞–Ї —Г–ґ —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –њ—А–Њ—В–Є–≤–Њ—А–µ—З–∞—В –Њ–±—К–µ–Ї—В–Є–≤–љ—Л–Љ –Њ—Ж–µ–љ–Ї–∞–Љ —Н—В–Є—Е –љ–∞—А—Г—И–µ–љ–Є–є, –Ї–∞–Ї —Н—В–Њ —З–∞—Б—В–Њ –Љ–Њ–ґ–љ–Њ –Ј–∞–Ї–ї—О—З–Є—В—М –њ–Њ¬†—А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ —А–∞—Б—З–µ—В–∞ —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є —Б–љ–∞ —В—А–∞–і–Є—Ж–Є–Њ–љ–љ—Л–Љ –Љ–µ—В–Њ–і–Њ–Љ.
–Э–∞–Ї–Њ–љ–µ—Ж, —В–∞–Ї–Њ–є –∞–љ–∞–ї–Є–Ј –Љ–Њ–ґ–µ—В –±—Л—В—М –љ–∞–њ—А–∞–≤–ї–µ–љ –љ–∞¬†–Њ–±–љ–∞—А—Г–ґ–µ–љ–Є–µ –Ї–Њ–љ–Ї—А–µ—В–љ—Л—Е –њ—А–Є—З–Є–љ –і–Є—Б—Б–Њ—Ж–Є–∞—Ж–Є–Є –Љ–µ–ґ–і—Г —А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ–Є –Њ–±—К–µ–Ї—В–Є–≤–љ–Њ–≥–Њ –Є–Ј—Г—З–µ–љ–Є—П —А–∞—Б—Б—В—А–Њ–є—Б—В–≤ —Б–љ–∞ –Є¬†–њ–Њ–≤–µ–і–µ–љ—З–µ—Б–Ї–Є–Љ–Є –Є–ї–Є —Б—Г–±—К–µ–Ї—В–Є–≤–љ—Л–Љ–Є —Б–Є–Љ–њ—В–Њ–Љ–∞–Љ–Є –µ–≥–Њ –љ–∞—А—Г—И–µ–љ–Є–є. –Т¬†–Њ—В–ї–Є—З–Є–µ –Њ—В¬†—В—А–∞–і–Є—Ж–Є–Њ–љ–љ—Л—Е –Ї—А–Є—В–µ—А–Є–µ–≤ –љ–∞—Б—В—Г–њ–ї–µ–љ–Є—П —Б–љ–∞ –Є¬†—Б–Љ–µ–љ—Л –µ–≥–Њ —Б—В–∞–і–Є–є –Ї—А–Є—В–µ—А–Є–Є, –Њ—Б–љ–Њ–≤–∞–љ–љ—Л–µ –љ–∞¬†–∞–љ–∞–ї–Є–Ј–µ –≠–≠–У-—Б–њ–µ–Ї—В—А–∞ –Љ–µ—В–Њ–і–Њ–Љ¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В, –Љ–Њ–≥—Г—В –±—Л—В—М –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ—Л –і–ї—П –Ї–ї–∞—Б—Б–Є—Д–Є–Ї–∞—Ж–Є–Є —Б—Г–±—Б–Њ—Б—В–Њ—П–љ–Є–є —Б–љ–∞ –≤¬†–ї—О–±–Њ–є –Њ–±–ї–∞—Б—В–Є –Ї–Њ—А—Л¬†–≥–Њ–ї–Њ–≤–љ–Њ–≥–Њ –Љ–Њ–Ј–≥–∞ [29]. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, —Н—В–Є –Ї—А–Є—В–µ—А–Є–Є –Љ–Њ–≥—Г—В –±—Л—В—М –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ—Л –і–ї—П –≤—Л—П–≤–ї–µ–љ–Є—П –Ї–Њ–љ–Ї—А–µ—В–љ—Л—Е —Б—Г–±—Б–Њ—Б—В–Њ—П–љ–Є–є —Б–Њ–љ–ї–Є–≤–Њ—Б—В–Є –≤¬†–њ—А–µ–і–µ–ї–∞—Е –Ї–Њ–љ—В–Є–љ—Г—Г–Љ–∞ ¬Ђ–±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ¬†вАУ—Б–Њ–љ–ї–Є–≤–Њ—Б—В—М¬ї [32]. –Ъ–∞–Ї –Є–Ј–≤–µ—Б—В–љ–Њ, –і–Њ —Б–Є—Е –њ–Њ—А¬†–≥—А–∞–і–∞—Ж–Є—П –і—А–µ–Љ–Њ—В–љ—Л—Е —Б–Њ—Б—В–Њ—П–љ–Є–є –Њ—Б—В–∞–µ—В—Б—П –Ј–∞ –њ—А–µ–і–µ–ї–∞–Љ–Є —В—А–∞–і–Є—Ж–Є–Њ–љ–љ–Њ–є —Б—В–∞–і–Є–є–љ–Њ–є –Ї–ї–∞—Б—Б–Є—Д–Є–Ї–∞—Ж–Є–Є, —Е–Њ—В—П –љ–µ–Њ–і–љ–Њ–Ї—А–∞—В–љ–Њ –≤—Л—Б–Ї–∞–Ј—Л–≤–∞–ї–Њ—Б—М –Љ–љ–µ–љ–Є–µ –Њ¬†—В–Њ–Љ, —З—В–Њ –њ–µ—А–Є–Њ–і, —А–∞–Ј–≥—А–∞–љ–Є—З–Є–≤–∞—О—Й–Є–є –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є–µ –Є¬†—Б–Њ–љ, —П–≤–ї—П–µ—В—Б—П –Ї–Њ–љ—В–Є–љ—Г—Г–Љ–Њ–Љ –і—А–µ–Љ–Њ—В–љ—Л—Е —Б—Г–±—Б–Њ—Б—В–Њ—П–љ–Є–є, –∞¬†–љ–µ –µ–і–Є–љ–Њ–є —Г–љ–Є—Д–Є—Ж–Є—А–Њ–≤–∞–љ–љ–Њ–є —Б—В–∞–і–Є–µ–є [14, 37].
–Ч–∞–Ї–ї—О—З–µ–љ–Є–µ
–†–µ–Ј—Г–ї—М—В–∞—В—Л –∞–љ–∞–ї–Є–Ј–∞ –і–Є–љ–∞–Љ–Є–Ї–Є –Ј–љ–∞—З–µ–љ–Є–є¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞—О—В —Б–Њ–≤–њ–∞–і–µ–љ–Є–µ –Љ–Њ–Љ–µ–љ—В–Њ–≤ —А–µ–Ј–Ї–Њ–≥–Њ –Є–Ј–Љ–µ–љ–µ–љ–Є—П –Ј–љ–∞—З–µ–љ–Є–є¬†–≥–ї–∞–≤–љ—Л—Е –Ї–Њ–Љ–њ–Њ–љ–µ–љ—В –≠–≠–У-—Б–њ–µ–Ї—В—А–∞ —Б¬†–њ–µ—А–µ—Е–Њ–і–∞–Љ–Є –Њ—В¬†–Њ–і–љ–Њ–є —Б—В–∞–і–Є–Є —Б–љ–∞ –Ї¬†–і—А—Г–≥–Њ–є. –£—Б–њ–µ—Е —В—А–∞–і–Є—Ж–Є–Њ–љ–љ–Њ–є —Б—В–∞–і–Є–є–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л, –Њ–±–µ—Б–њ–µ—З–Є–≤—И–µ–є –љ–∞¬†–њ–Њ–ї—Б—В–Њ–ї–µ—В–Є—П –љ–µ–њ—А–µ—А—Л–≤–љ–Њ—Б—В—М –љ–∞—Г—З–љ—Л—Е –Є¬†–Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є—Е –Њ–њ–Є—Б–∞–љ–Є–є —Б–љ–∞, –Љ–Њ–ґ–µ—В –±—Л—В—М –Њ–±—К—П—Б–љ–µ–љ —В–µ–Љ, —З—В–Њ¬†–≥—А–∞–љ–Є—Ж—Л —Б—В–∞–і–Є–є —Б–љ–∞ –њ—А–Є–Љ–µ—А–љ–Њ —Б–Њ–≤–њ–∞–і–∞—О—В —Б¬†–Љ–Њ–Љ–µ–љ—В–∞–Љ–Є, –Ї–Њ–≥–і–∞ —Б–Њ—Б—В–Њ—П–љ–Є—П —Б–љ–∞¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П –Є¬†—Б—Г–±—Б–Њ—Б—В–Њ—П–љ–Є—П —Б–љ–∞ —Б–Љ–µ–љ—П—О—В –і—А—Г–≥ –і—А—Г–≥–∞ –≤¬†—А–µ–Ј—Г–ї—М—В–∞—В–µ –∞–љ—В–∞–≥–Њ–љ–Є—Б—В–Є—З–µ—Б–Ї–Њ–≥–Њ –≤–Ј–∞–Є–Љ–Њ–і–µ–є—Б—В–≤–Є—П –Љ–µ–ґ–і—Г –њ—А–Њ—Ж–µ—Б—Б–∞–Љ–Є –њ—А–Њ–Љ–Њ—В–Њ—А–∞–Љ–Є –Є¬†–Є–љ–≥–Є–±–Є—В–Њ—А–∞–Љ–Є —Б–љ–∞¬†вАУ –±–Њ–і—А—Б—В–≤–Њ–≤–∞–љ–Є—П.
A.A. Putilov
Scientific Research Institute of Molecular Biology and Biophysics, Novosibirsk
Contact person: Arkady Aleksandrovich Putilov, putilov@ngs.ru
Origin and history of sleep science were closely associated with discoveries leading to development of the traditional system of description of night sleep episodes as the sequences of few yes-or-no variables called sleep stages. The unexpected success of this simple system provided continuity in the scientific and clinical description of the sleep process for, at least, a half of century in despite of its numerous unresolved shortcomings. This success can be explained by a tendency for coincidence of the epochs of transition between sleep stages with the intervals of switching between sub-states of the sleep-wake continuum due to the antagonistic interaction between underlying wake- and sleep-promoting processes. This explanation was supported by the present analysis of the time courses of scores on principal components of the EEG spectrum that were utilized as the EEG indicators of these processes. Particularly, it was found that the rapid zero-crossing changes in these scores usually occurred during transition between wake and sleep states and sub-states. Prospects of implementation of this approach to principal component scoring of the EEG spectrum in sleep medicine are also discussed.
–£–≤–∞–ґ–∞–µ–Љ—Л–є –њ–Њ—Б–µ—В–Є—В–µ–ї—М uMEDp!
–£–≤–µ–і–Њ–Љ–ї—П–µ–Љ –Т–∞—Б –Њ —В–Њ–Љ, —З—В–Њ –Ј–і–µ—Б—М —Б–Њ–і–µ—А–ґ–Є—В—Б—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П, –њ—А–µ–і–љ–∞–Ј–љ–∞—З–µ–љ–љ–∞—П –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –і–ї—П —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П.
–Х—Б–ї–Є –Т—Л –љ–µ —П–≤–ї—П–µ—В–µ—Б—М —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–Љ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П, –∞–і–Љ–Є–љ–Є—Б—В—А–∞—Ж–Є—П –љ–µ –љ–µ—Б–µ—В –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ—Б—В–Є –Ј–∞ –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ –њ–Њ—Б–ї–µ–і—Б—В–≤–Є—П, –≤–Њ–Ј–љ–Є–Ї—И–Є–µ –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–∞–Љ–Њ—Б—В–Њ—П—В–µ–ї—М–љ–Њ–≥–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Т–∞–Љ–Є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є —Б –њ–Њ—А—В–∞–ї–∞ –±–µ–Ј –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є –Ї–Њ–љ—Б—Г–ї—М—В–∞—Ж–Є–Є —Б –≤—А–∞—З–Њ–Љ.
–Э–∞–ґ–Є–Љ–∞—П –љ–∞ –Ї–љ–Њ–њ–Ї—Г ¬Ђ–Т–Њ–є—В–Є¬ї, –Т—Л –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В–µ, —З—В–Њ —П–≤–ї—П–µ—В–µ—Б—М –≤—А–∞—З–Њ–Љ –Є–ї–Є —Б—В—Г–і–µ–љ—В–Њ–Љ –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–≥–Њ –≤—Г–Ј–∞.