ะะบะธัะปะธัะตะปัะฝัะน ัััะตัั ะธย ะผัะถัะบะพะต ะฑะตัะฟะปะพะดะธะตย โ ะฒะทะฐะธะผะพัะฒัะทะฐะฝะฝัะต ะฟะฐะฝะดะตะผะธะธ XXI ะฒ. ะกะพะฒัะตะผะตะฝะฝัะต ัะฐัะผะฐะบะพัะตัะฐะฟะตะฒัะธัะตัะบะธะต ะฒะพะทะผะพะถะฝะพััะธ ะฟะฐัะพะณะตะฝะตัะธัะตัะบะพะน ะบะพััะตะบัะธะธ ะฝะฐัััะตะฝะธะน ัะฟะตัะผะฐัะพะณะตะฝะตะทะฐ ะฟัะตะฟะฐัะฐัะฐะผะธ L-ะบะฐัะฝะธัะธะฝะฐ/ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ
- ะะฝะฝะพัะฐัะธั
- ะกัะฐััั
- ะกััะปะบะธ
- English
![ะ ะธั. 1. ะะพะปะพัะพะน ััะตัะณะพะปัะฝะธะบ ะพะบัะธะดะฐัะธะฒะฝะพะณะพ ะฑะฐะปะฐะฝัะฐ ะฒย ะฝะพัะผะต [6]](/upload/iblock/67c/spermaktin1.jpg)
![ะ ะธั. 2. ะะพะดัะปััะธั ัะตะดะพะบั-ะฟะพัะตะฝัะธะฐะปะฐ ะฑะธะพะปะพะณะธัะตัะบะพะน ััะตะดั ะฟัะธ ัะธะทะธะพะปะพะณะธัะตัะบะพะผ ะธย ะฟะฐัะพะปะพะณะธัะตัะบะพะผ ะพะบะธัะปะธัะตะปัะฝะพะผ ัััะตััะต [7]](/upload/iblock/8df/spermaktin2.jpg)
![ะ ะธั. 3. ะะพะปะธะพัะณะฐะฝะฝะฐั ะผัะปััะธัะฐะบัะพัะฝะฐั ะฟะฐัะพะปะพะณะธั, ะธะฝะดััะธััะตะผะฐั ะฟะฐัะพะปะพะณะธัะตัะบะธะผ ะะก [8]](/upload/iblock/048/spermaktin3.jpg)
![ะ ะธั. 4. ะญัะธะพะปะพะณะธัะตัะบะธะต ัะฐะบัะพัั ัะฟะตัะผะฐะปัะฝะพะณะพ ะพะบะธัะปะธัะตะปัะฝะพะณะพ ัััะตััะฐ [33]](/upload/iblock/814/spermaktin4.jpg)
ะะฒะตะดะตะฝะธะต
ะกะพะณะปะฐัะฝะพ ะพะฟัะตะดะตะปะตะฝะธั ะัะตะผะธัะฝะพะน ะพัะณะฐะฝะธะทะฐัะธะธ ะทะดัะฐะฒะพะพั ัะฐะฝะตะฝะธั, ะฟะพะฝััะธะต ยซะทะดะพัะพะฒัะน ัะตะปะพะฒะตะบยป ะฟะพะดัะฐะทัะผะตะฒะฐะตั, ะฝะฐััะดั ัย ะฟัะธั ะธัะตัะบะธะผ ะธย ัะพัะธะฐะปัะฝัะผ ะทะดะพัะพะฒัะตะผ, ัะตะฟัะพะดัะบัะธะฒะฝะพะต. ะกย ัะพะถะฐะปะตะฝะธะตะผ ะฟัะธั ะพะดะธััั ะบะพะฝััะฐัะธัะพะฒะฐัั ะฝะตะฑะปะฐะณะพะฟะพะปััะธะต ะฒัะตั ััะตั ัะบะฐะทะฐะฝะฝัั ะฟะฐัะฐะผะตััะพะฒ ะฒย XXI ะฒ. ะะฐะธะฑะพะปััะตะต ะพะฟะฐัะตะฝะธะต ะฒัะทัะฒะฐัั ัะพะฒัะตะผะตะฝะฝัะต ะฝะตะณะฐัะธะฒะฝัะต ัะตะฝะดะตะฝัะธะธ, ั ะฐัะฐะบัะตัะธะทัััะธะต ัะพััะพัะฝะธะต ัะตะฟัะพะดัะบัะธะฒะฝะพะน ััะฝะบัะธะธ. ะย ะ ะพััะธะธ ัะพั ัะฐะฝัะตััั ะพะดะธะฝ ะธะทย ัะฐะผัั ะฒััะพะบะธั ะฟะพะบะฐะทะฐัะตะปะตะน ัะตะผะตะนะฝะพะณะพ ะฑะตัะฟะปะพะดะธั, ะดะพััะธะณะฐััะธะน 19โ20% (ะฒย ะะฒัะพะฟะต ะพะบะพะปะพ 15%). ะกะปะพะถะธะฒัะฐััั ัะธััะฐัะธั ะผะพะถะตั ะฟัะธะฒะตััะธ ะบย ัััะตััะฒะตะฝะฝะพะผั ัะพะบัะฐัะตะฝะธั ัะธัะปะตะฝะฝะพััะธ ะฝะฐัะตะปะตะฝะธั ัััะฐะฝั ะฒย ะฑะปะธะถะฐะนัะธะต ะดะตัััะธะปะตัะธั, ะตัะปะธ ะฝะตย ะฑัะฐัั ะฒย ัะฐััะตั ัะพัั ะฝะฐัะตะปะตะฝะธั ะทะฐ ััะตั ะธะผะผะธะณัะฐัะธะธ.
ะ ะพะปั ะผัะถัะบะพะณะพ ะฑะตัะฟะปะพะดะธั ะฝะฐัะฐะปะฐ ะฟะตัะตะพัะผััะปะธะฒะฐัััั ัะพะปัะบะพ ัะตะนัะฐั, ะบะพะณะดะฐ ะดะพะปั ยซะผัะถัะบะพะณะพยป ัะฐะบัะพัะฐ ะฒย ััััะบัััะต ัะตะผะตะนะฝะพะณะพ ะฑะตัะฟะปะพะดะธั ััะฐะปะฐ ะบะฐัะฐัััะพัะธัะตัะบะธ ัะฐััะธ ะธย ะดะพััะธะณะปะฐ 50%. ะัะธ ััะพะผ ะผัะถัะบะพะต ะฑะตัะฟะปะพะดะธะต ะฝะตะฟะพะปะฝะพัะตะฝะฝะพ ะดะธะฐะณะฝะพััะธััะตััั ะธย ะปะตัะธััั. ะะตัะผะพััั ะฝะฐย ะผะฝะพะถะตััะฒะพ ะฟัะธัะธะฝ ะฝะฐัััะตะฝะธั ะผัะถัะบะพะน ัะตััะธะปัะฝะพััะธ, ะฒัะต ัะพัะผั ะผัะถัะบะพะณะพ ะฑะตัะฟะปะพะดะธั ะธะผะตัั ะพะฑัะธะต ัะฝะธะฒะตััะฐะปัะฝัะต ะบะปะตัะพัะฝะพ-ะผะพะปะตะบัะปััะฝัะต ะผะตั ะฐะฝะธะทะผั ัะตะฐะปะธะทะฐัะธะธ ะฝะตะณะฐัะธะฒะฝัั ัััะตะบัะพะฒ ััะธะพะปะพะณะธัะตัะบะธั ัะฐะบัะพัะพะฒ ะฝะฐย ะฒัะตั ััะฐะฟะฐั ัะฟะตัะผะฐัะพะณะตะฝะตะทะฐ. ะย ะบะฐัะตััะฒะต ะพะดะฝะพะณะพ ะธะทย ัะฝะธะฒะตััะฐะปัะฝัั ะฟะฐัะพะณะตะฝะตัะธัะตัะบะธั ัะฐะบัะพัะพะฒ ัะฐััะผะฐััะธะฒะฐะตััั ะพะบะธัะปะธัะตะปัะฝัะน ัััะตัั (ะะก). ะกะฟะพัะพะฑะฝะพััั ะะก ะทะฐะฟััะบะฐัั ะผัะปััะธัะฐะบัะพัะฝัั ะฟะพะปะธะพัะณะฐะฝะฝัั ะฟะฐัะพะปะพะณะธั, ะฒะบะปััะฐั ัะตะฟัะพะดัะบัะธะฒะฝัะต ะฝะฐัััะตะฝะธั ะบะฐะบ ัย ะผัะถัะธะฝ, ัะฐะบ ะธย ั ะถะตะฝัะธะฝ, ะพะฑััะปะพะฒะปะธะฒะฐะตั ะผะตะดะธัะธะฝัะบัั ะฝะฐััะฝะพ-ะฟัะฐะบัะธัะตัะบัั ะทะฝะฐัะธะผะพััั ััะพะน ะฟัะพะฑะปะตะผั [1].
ะะบะธัะปะธัะตะปัะฝัะน ัััะตัั: ะพะฑัะธะต ะฟะฐัะพัะธะทะธะพะปะพะณะธัะตัะบะธะต ะผะตั ะฐะฝะธะทะผั, ะฟะพะฝััะธะต ะพย ัะธะทะธะพะปะพะณะธัะตัะบะพะผ ะธย ะฟะฐัะพะปะพะณะธัะตัะบะพะผ ะพะบะธัะปะธัะตะปัะฝะพะผ ัััะตััะต
ะะบะธัะปะธัะตะปัะฝะพ-ะฒะพัััะฐะฝะพะฒะธยญัะตะปัะฝัะต ัะตะฐะบัะธะธ (ะะะ ), ะธะปะธ ัะตะดะพะบั-ัะตะฐะบัะธะธ (ะพัย ะฐะฝะณะป. reduction oxidationย โ ะฒะพัััะฐะฝะพะฒะปะตะฝะธะต/ะพะบะธัะปะตะฝะธะต),ย โ ะบะปััะตะฒัะต ัะธะทะธะพะปะพะณะธัะตัะบะธะต ะฟัะพัะตััั, ะฝะตะพะฑั ะพะดะธะผัะต ะดะปั ะถะธะทะฝะตะดะตััะตะปัะฝะพััะธ ะปัะฑะพะณะพ ะถะธะฒะพะณะพ ะพัะณะฐะฝะธะทะผะฐ. ะญะฝะตัะณะธั, ะพัะฒะพะฑะพะถะดะฐััะฐััั ะฒย ั ะพะดะต ััะธั ัะตะฐะบัะธะน, ัะฐัั ะพะดัะตััั ะฝะฐย ะฟะพะดะดะตัะถะฐะฝะธะต ะฒัะตั ะฟะฐัะฐะผะตััะพะฒย ะณะพะผะตะพยญััะฐะทะฐ ะธย ะพะฑะตัะฟะตัะตะฝะธะต ะฝะพัะผะฐะปัะฝะพะณะพ ััะฝะบัะธะพะฝะธัะพะฒะฐะฝะธั ะฒัะตั ััััะบัััะฝัั ัะปะตะผะตะฝัะพะฒ ะบะปะตัะพะบ ะธย ัะบะฐะฝะตะน [1].
ะกย ั ะธะผะธัะตัะบะพะน ัะพัะบะธ ะทัะตะฝะธั ะะะ ย โ ััะพ ะฒัััะตัะฝะพ-ะฟะฐัะฐะปะปะตะปัะฝัะต ั ะธะผะธัะตัะบะธะต ัะตะฐะบัะธะธ. ะะฝะธ ะฟัะพัะตะบะฐัั ัย ะธะทะผะตะฝะตะฝะธะตะผ ััะตะฟะตะฝะตะน ะพะบะธัะปะตะฝะธั ะฐัะพะผะพะฒ (ะธะพะฝะพะฒ) ัะตะฐะณะธััััะธั ะฒะตัะตััะฒ. ะะทะผะตะฝะตะฝะธะต ัะตะฐะปะธะทัะตััั ะฟััะตะผ ะฟะตัะตัะฐัะฟัะตะดะตะปะตะฝะธั ัะปะตะบััะพะฝะพะฒ ะผะตะถะดั ะฐัะพะผะพะผ-ะพะบะธัะปะธัะตะปะตะผ (ะฐะบัะตะฟัะพัะพะผ) ะธย ะฐัะพะผะพะผ-ะฒะพัััะฐะฝะพะฒะธัะตะปะตะผ (ะดะพะฝะพัะพะผ). ะย ะฐััะพะฑะฝะพะผ ะพัะณะฐะฝะธะทะผะต ัะตะปะพะฒะตะบะฐ ะบะปััะตะฒัะผ ะพะบะธัะปะธัะตะปะตะผ ะฒััััะฟะฐะตั ะบะธัะปะพัะพะด, ะฟะพััะพะผั ะะะ ัะพััะฐะฒะปััั ัััั ะฝะพัะผะฐะปัะฝะพะน ัะธะทะธะพะปะพะณะธะธ. ะคะธะทะธะพะปะพะณะธัะตัะบะธะน ะะกย โ ะพะดะฝะพ ะธะทย ััะฝะดะฐะผะตะฝัะฐะปัะฝัั ััะปะพะฒะธะน ะฝะพัะผะฐะปัะฝะพะณะพ ััะฝะบัะธะพะฝะธัะพะฒะฐะฝะธั ะฒัะตั ะถะธะฒัั ะฐััะพะฑะฝัั ะบะปะตัะพะบ ะธย ัะธััะตะผ. ะะบะพะปะพ 95% ะฒัะตะณะพ ะฟะพััะตะฑะปัะตะผะพะณะพ ะบะธัะปะพัะพะดะฐ ะฒย ะบะปะตัะบะต ะฒะพัััะฐะฝะฐะฒะปะธะฒะฐะตััั ะฒย ะผะธัะพั ะพะฝะดัะธัั ะดะพ ะฒะพะดั ะฒย ะฟัะพัะตััะต ะพะบะธัะปะธัะตะปัะฝะพะณะพ ัะพััะพัะธะปะธัะพะฒะฐะฝะธั ะธย ัะธะฝัะตะทะฐ ะะขะค. ะััะฐะปัะฝัะต 5% ะฒย ัะตะทัะปััะฐัะต ัะฐะทะปะธัะฝัั ัะตะฐะบัะธะน ะฝะตะธะทะฑะตะถะฝะพ ะฟัะตะฒัะฐัะฐัััั ะฒย ะฐะบัะธะฒะฝัะต ัะพัะผั ะบะธัะปะพัะพะดะฐ, ะธะปะธ ัะฒะพะฑะพะดะฝัะต ัะฐะดะธะบะฐะปั (ะกะ ) [2, 3].
ะย ะฝะพัะผะต ัะตะปัะน ััะด ะฑะธะพั ะธะผะธัะตัะบะธั ะฟัะพัะตััะพะฒ, ะฟัะพัะตะบะฐััะธั ะฒย ะฐััะพะฑะฝัั ะพัะณะฐะฝะธะทะผะฐั , ัะพะฟััะถะตะฝ ัย ะตััะตััะฒะตะฝะฝัะผ ะพะฑัะฐะทะพะฒะฐะฝะธะตะผ ะกะ . ะะดะฝะฐะบะพ ะกะ , ะพะฑัะฐะทัััะธะตัั ะฒย ั ะพะดะต ะะะ , ัััะตััะฒะตะฝะฝะพ ัะฐะทะปะธัะฐัััั ะฟัะตะถะดะต ะฒัะตะณะพ ะฟะพย ััะฝะบัะธะพะฝะฐะปัะฝะพะน ะฝะฐะณััะทะบะต. ะขะฐะบ, ะพะดะฝะธ (ะฟะตัะฒะธัะฝัะต) ะกะ ะฟะพะปะตะทะฝั ะดะปั ะพัะณะฐะฝะธะทะผะฐ, ะฟะพัะบะพะปัะบั ััะฐััะฒััั ะฒย ัะตะปะพะผ ัะฟะตะบััะต ะฝะตะพะฑั ะพะดะธะผัั ะดะปั ะฝะพัะผะฐะปัะฝะพะน ะถะธะทะฝะตะดะตััะตะปัะฝะพััะธ ัะธะทะธะพะปะพะณะธัะตัะบะธั ัะตะฐะบัะธะน:
- ัะตะณัะปััะธะธ ะบะปะตัะพัะฝัั ะฟัะพัะตััะพะฒ (ะบะปะตัะพัะฝะพะต ะดะตะปะตะฝะธะต, ะดัั ะฐะฝะธะต) ัะตัะตะท ะทะฐะฒะธัะธะผัะน ะพัย ะฐะบัะธะฒะฝัั ัะพัะผ ะบะธัะปะพัะพะดะฐ ัะธะณะฝะฐะปะธะฝะณ;
- ะพะฑะตัะฟะตัะตะฝะธะธ ะฑะฐะบัะตัะธัะธะดะฝะพะณะพ ะธย ะพะฝะบะพััะฐัะธัะตัะบะพะณะพ ัััะตะบัะพะฒ;
- ะฐะบัะธะฒะฐัะธะธ ะธะผะผัะฝะฝัั ัะตะฐะบัะธะน ะปะตะนะบะพัะธัะพะฒ;
- ะพะบะฐะทะฐะฝะธะธ ะฟัะพัะธะฒะพะฒะพัะฟะฐะปะธัะตะปัะฝะพะณะพ ัะธััะตะผะฝะพะณะพ ะธย ะปะพะบะฐะปัะฝะพะณะพ ะพัะฒะตัะฐ ะธย ั.ะด.
ะะฐะธะฑะพะปะตะต ะทะฝะฐัะธะผั ะดะปั ะพัะณะฐะฝะธะทะผะฐ ัะฐะบะธะต ะฟะตัะฒะธัะฝัะต ะกะ , ะบะฐะบ ััะฟะตัะพะบัะธะดะฝัะน ะฐะฝะธะพะฝ-ัะฐะดะธะบะฐะป (O2-),ย ะณะธะดัะพะบัะธะปัะฝัะน ัะฐะดะธะบะฐะป (โขะะ), ะฟะตัะตะบะธัั ะฒะพะดะพัะพะดะฐ (H2O2), ัะธะฝะณะปะตัะฝัะน ะบะธัะปะพัะพะด (1O2),ย ะณะธะฟะพั ะปะพัะฝะฐั ะบะธัะปะพัะฐ (HOCl), ะพะบัะธะด ะฐะทะพัะฐ (NO.), ะฟะตัะพะบัะธะฝะธััะธั (ONOO-) [1โ3]. ะัะพัะธัะฝัะต ะกะ , ะฒย ะพัะปะธัะธะต ะพัย ะฟะตัะฒะธัะฝัั , ะฝะตย ะฒัะฟะพะปะฝััั ัะธะทะธะพะปะพะณะธัะตัะบะธ ะฟะพะปะตะทะฝัั ััะฝะบัะธะน, ะฝะฐะฟัะพัะธะฒ, ะพะฝะธ ัะฐะทัััะธัะตะปัะฝะพ ะดะตะนััะฒััั ะฝะฐย ะบะปะตัะพัะฝัะต ััััะบัััั, ัััะตะผััั ะพัะฝััั ัะปะตะบััะพะฝั ัย ยซะฟะพะปะฝะพัะตะฝะฝัั ยป ะผะพะปะตะบัะป, ะฒัะปะตะดััะฒะธะต ัะตะณะพ ยซะฟะพัััะฐะดะฐะฒัะฐัยป ะผะพะปะตะบัะปะฐ ัะฐะผะฐ ััะฐะฝะพะฒะธััั ัะปะฐะฑัะผ ะกะ (ััะตัะธัะฝัะผ) [1โ3].
ะย ะฝะพัะผะต ะบะปะตัะบะธ ะธะผะตัั ะพัะพะฑัั ัะธััะตะผั ะทะฐัะธัั (ะฐะฝัะธะพะบัะธะดะฐะฝัยญะฝัั) ะพัย ะพะฑัะฐะทัััะธั ัั ะฒย ะฟัะพัะตััะต ะถะธะทะฝะตะดะตััะตะปัะฝะพััะธ ะกะ . ะะฝัะธะพะบัะธะดะฐะฝัะฝะฐั ัะธััะตะผะฐ ะพะฑัะฐะทะพะฒะฐะฝะฐ ะฝะธะทะบะพะผะพะปะตะบัะปััะฝัะผะธ ะฐะฝัะธะพะบัะธะดะฐะฝัะฐะผะธ ะธย ัะฟะตัะธะฐะปะธะทะธัะพะฒะฐะฝะฝัะผะธ ะฐะฝัะธะพะบัะธะดะฐะฝัะฝัะผะธ ัะตัะผะตะฝัะฐะผะธ. ะย ะบะปััะตะฒัะผ ะฐะฝัะธะพะบัะธะดะฐะฝัะฝัะผ ัะตัะผะตะฝัะฐะผ ะพัะฝะพััััั ัะตัะผะตะฝัั ัะฟะตัะธะฐะปะธะทะธัะพะฒะฐะฝะฝะพะน ัะตัะผะตะฝัะฝะพะน ัะธััะตะผั, ะฒย ะบะพัะพััั ะฒั ะพะดัั ััะฟะตัะพะบัะธะด-ะดะธัะผััะฐะทะฐ, ะบะฐัะฐะปะฐะทะฐ ะธย ะณะปััะฐัะธะพะฝ-ะฟะตัะพะบัะธะดะฐะทะฐ. ะะฝะธ ะบะฐัะฐะปะธะทะธัััั ัะตะฐะบัะธะธ, ะฒย ัะตะทัะปััะฐัะต ะบะพัะพััั ะกะ ะธย ะฟะตัะตะบะธัะธ ะฟัะตะฒัะฐัะฐัััั ะฒย ะฝะตะฐะบัะธะฒะฝัะต ัะพะตะดะธะฝะตะฝะธั. ะัะพะผะต ัะพะณะพ, ัััะตััะฒััั ะฝะตัะฟะตัะธะฐะปะธะทะธัะพะฒะฐะฝะฝัะต ัะตัะผะตะฝัะฝัะต ัะธััะตะผั ะฟะพย ะธะฝะฐะบัะธะฒะฐัะธะธ ัะฒะพะฑะพะดะฝัั ัะฐะดะธะบะฐะปะพะฒ, ะฟัะตะดััะฐะฒะปะตะฝะฝัะต ะฝะธะทะบะพะผะพะปะตะบัะปััะฝัะผะธ ะฐะฝัะธะพะบัะธะดะฐะฝัะฐะผะธ โ ะฒะธัะฐะผะธะฝะฐะผะธ ะ, ะ, ะ, ะก, D, ััะตัะพะธะดะฝัะผะธย ะณะพัะผะพะฝะฐะผะธ, ัะปะฐะฒะพะฝะพะธะดะฐะผะธ, ะฟะพะปะธัะตะฝะพะปะฐะผะธ (ะฒะธัะฐะผะธะฝะพะผ ะ , ะบะพัะฝะทะธะผะพะผ Q10, ะธะปะธ ัะฑะธั ะธะฝะพะฝะพะผ), ัะธะพะป-ะดะธััะปััะธะดะฝะพะน ัะธััะตะผะพะน ะฝะฐย ะพัะฝะพะฒะตย ะณะปััะฐัะธะพะฝะฐ (ะฒย ัะฐััะฝะพััะธ, ะฐะปััะฐ-ะปะธะฟะพะตะฒะพะน ะบะธัะปะพัะพะน), ะฐัะพะผะฐัะธัะตัะบะธะผะธ ัะพะตะดะธะฝะตะฝะธัะผะธ, ะผะพัะตะฒะพะน ะบะธัะปะพัะพะน, ัะฐััะธะฝะพะผ, ะบะฐัะฝะพะทะธะฝะพะผ, ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะพะผ, ะฐัะตัะธะปัะธััะตะธะฝะพะผ, ั ะตะปะฐัะพัะฐะผะธ ะธะพะฝะพะฒ ะถะตะปะตะทะฐ, ัะธะฝะบะพะผ, ัะตะปะตะฝะพะผ ะธย ะดั. [4, 5].
ะะฝะพะณะพัะธัะปะตะฝะฝัะต ะบะพะผะฟะพะฝะตะฝัั ะบะปะตัะพัะฝะพะน ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะพะน ัะธััะตะผั, ะธะฝะณะธะฑะธััั ะธะทะฑััะพัะฝะพััั ะกะ , ะฟะพะดะดะตัะถะธะฒะฐัั ะะก ะฝะฐย ัะธะทะธะพะปะพะณะธัะตัะบะธ ะฐะดะตะบะฒะฐัะฝะพะผ ััะพะฒะฝะต. ยซะคะธะทะธะพะปะพะณะธัะฝะพัััยป ัะพั ัะฐะฝัะตััั ะดะพ ัะตั ะฟะพั, ะฟะพะบะฐ ัะพะฑะปัะดะฐะตััั ะฟัะธะฝัะธะฟ ะทะพะปะพัะพะณะพ ััะตัะณะพะปัะฝะธะบะฐ ะพะบัะธะดะฐัะธะฒะฝะพะณะพ ะฑะฐะปะฐะฝัะฐ. ะขะพะปัะบะพ ะดะธะฝะฐะผะธัะตัะบะพะต ัะฐะฒะฝะพะฒะตัะธะต ะผะตะถะดั ััะพะฒะฝะตะผ ะฟัะพะดัะบัะธะธ ะกะ , ะฐะบัะธะฒะฝะพัััั ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะพะน ัะธััะตะผั ะธย ะฝะพัะผะฐะปัะฝัะผ ััะฝะบัะธะพะฝะธัะพะฒะฐะฝะธะตะผ ััะฐะฝัะผะธััะตัะฝัั (ะฟะตัะตะดะฐััะธั ะฑะธะพะปะพะณะธัะตัะบะธะต ัะธะณะฝะฐะปั) ะฑะธะพะผะพะปะตะบัะป ะผะพะถะตั ะพะฑะตัะฟะตัะธัั ะฑะธะพะปะพะณะธัะตัะบัั ะฑะตะทะพะฟะฐัะฝะพััั ะบะปะตัะบะธ ะธย ะฒัะตะณะพ ะพัะณะฐะฝะธะทะผะฐ ะฒย ัะตะปะพะผ (ัะธั.ย 1) [6]. ะัะธ ะฝะฐัััะตะฝะธะธ ะทะพะปะพัะพะณะพ ััะตัะณะพะปัะฝะธะบะฐ ะพะบัะธะดะฐัะธะฒะฝะพะณะพ ะฑะฐะปะฐะฝัะฐ (ะณะธะฟะตัะฟัะพะดัะบัะธะธ ะกะ ะธ/ะธะปะธ ะฝะตะดะพััะฐัะพัะฝะพะน ัะบะพัะพััะธ ะธั ะธะฝะฐะบัะธะฒะฐัะธะธ, ะดะตัะธัะธัะต/ะธััะพัะตะฝะธะธ ัะฐะบัะพัะพะฒ ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะพะน ัะธััะตะผั ะธะปะธ ะธั ัะพัะตัะฐะฝะธั) ยซัะธะทะธะพะปะพะณะธัะตัะบะธะนยป ะะก ะฑััััะพ ะธย ะฝะตะทะฐะผะตัะฝะพ ะฟะตัะตั ะพะดะธั ะฒย ะธะทะฑััะพัะฝะพ ะฒััะฐะถะตะฝะฝัะน ะฟะฐัะพะปะพะณะธัะตัะบะธะน (ัะธั.ย 2) [7].
ะะฐัะพะปะพะณะธัะตัะบะธะน ะะก ะฟะพัะฐะถะฐะตั ะฟัะฐะบัะธัะตัะบะธ ะฒัะต ััััะบัััั ะพัะณะฐะฝะธะทะผะฐ, ะฒะบะปััะฐั ะะะ, ะฑะตะปะบะธ ะธย ะปะธะฟะธะดั ะผะตะผะฑัะฐะฝ. ะะฐย ะบะปะตัะพัะฝะพ-ัะบะฐะฝะตะฒะพะผ ััะพะฒะฝะต ะพะฝ ะฟัะพัะฒะปัะตััั ัะฐะทะปะธัะฝัะผะธ ะฟะฐัะพะปะพะณะธัะตัะบะธะผะธ ะฟัะพัะตััะฐะผะธ:
- ะฝะฐัััะตะฝะธะตะผย ะณะพะผะตะพััะฐะทะฐ, ะฒย ัะฐััะฝะพััะธ ะดะธัะฑะฐะปะฐะฝัะพะผ ะผะตะถะดั ะฟัะพ- ะธย ะฟัะพัะธะฒะพะฒะพัะฟะฐะปะธัะตะปัะฝัะผะธ ัะธัะพยญะบะธะฝะฐะผะธ (ั ัะพะฝะธัะตัะบะพะต ัะธััะตะผะฝะพะต ะฒะพัะฟะฐะปะตะฝะธะต);
- ะธัะตะผะธะตะน (ัะฝะดะพัะตะปะธะฐะปัะฝะพะน ะดะธัััะฝะบัะธะตะน); ย ะณะธะฟะพะบัะธะตะน (ะผะตะผะฑัะฐะฝะพะฟะฐัะธะธ ะฒัะปะตะดััะฒะธะต ะฐะบัะธะฒะฐัะธะธ ะฟะตัะตะบะธัะฝะพะณะพ ะพะบะธัะปะตะฝะธั ะปะธะฟะธะดะพะฒ ะผะตะผะฑัะฐะฝ ะบะปะตัะพะบ);
- ะฐะฟะพะฟัะพะทะพะผ ะธย ะฝะตะบัะพะทะพะผ ะบะปะตัะพะบ;
- ะฝะฐัััะตะฝะธะตะผ ะบะปะตัะพัะฝะพะน ัะตัะตะฟัะธะธ ะธย ะฟะตััะตะฟัะธะธ (ะฐัะตัะปะตะบัะธั ะธย ะณะธะฟะพัะตัะปะตะบัะธั ะบะปะตัะบะธ);
- ะฒะตะณะตัะฐัะธะฒะฝะพ-ะผะตะดะธะฐัะพัะฝะพะน ะดะธัััะฝะบัะธะตะน ะบะปะตัะบะธ;
- ัะฝะตัะณะตัะธัะตัะบะธะผะธ ะธย ะผะตัะฐะฑะพะปะธัะตัะบะธะผะธ ะฝะฐัััะตะฝะธัะผะธ (ะผะธัะพั ะพะฝะดัะธะฐะปัะฝะฐั ะดะธัััะฝะบัะธั) ะธย ะดั.
ะัะธ ะฟะฐัะพะปะพะณะธัะตัะบะพะผ ะะก ะฒย ะบะปะตัะบะต ะฝะฐะบะฐะฟะปะธะฒะฐัััั ะบะธัะปัะต ะฟัะพะดัะบัั ะดะตะณัะฐะดะฐัะธะธ (ะฝะฐัััะตะฝะธะต ะบะปะตัะพัะฝะพะณะพ ัะตะดะพะบั-ะฟะพัะตะฝัะธะฐะปะฐ), ััะพ ะฒะตะดะตั ะบย ัะดะฒะธะณั ัะ ะฑะธะพะปะพะณะธัะตัะบะธั ะถะธะดะบะพััะตะน ะธย ัะธัะพะฟะปะฐะทะผั ะบะปะตัะพะบ ะฒย ะบะธัะปัั ััะพัะพะฝั (ะทะฐะบะธัะปะตะฝะธั, ะธะปะธ ะฐัะธะดะพะทั). ะย ะธัะพะณะต ััะธย ะณะพะผะตะพยญััะฐัะธัะตัะบะธะต ะฝะฐัััะตะฝะธั ะฟัะธะฒะพะดัั ัะฝะฐัะฐะปะฐ ะบย ััะฝะบัะธะพะฝะฐะปัะฝะพะน, ะฐย ะฟัะธ ะดะปะธัะตะปัะฝะพะน ะฟะตััะธััะตะฝัะธะธ ะธ/ะธะปะธ ะฟัะพะณัะตััะธัะพะฒะฐะฝะธะธ ะฟะฐัะพะปะพะณะธัะตัะบะพะณะพ ะะกย โ ะธย ะบ ะพัะณะฐะฝะธัะตัะบะพะน ะบะปะตัะพัะฝะพ-ัะบะฐะฝะตะฒะพะน ะฟะฐัะพะปะพะณะธะธ. ะะฝะฐ ะฟัะตะดะพะฟัะตะดะตะปัะตั ัะฐะทะฒะธัะธะต ะฟะพะปะธะพัะณะฐะฝะฝัั ะธย ะผัะปััะธัะฐะบัะพัะฝัั ัะธััะตะผะฝัั ะฝะฐัััะตะฝะธะน, ะฒะบะปััะฐั ัะตะฟัะพะดัะบัะธะฒะฝัะต ะฟะพัะตัะธ (ัะธั.ย 3) [8].
ะกะปะตะดัะตั ะทะฐะผะตัะธัั, ััะพ ะฑะพะปััะธะฝััะฒะพ ะบะปะตัะพะบ ัะฟะพัะพะฑะฝั ะฟะตัะตะฝะพัะธัั ัะผะตัะตะฝะฝัะน ะะก ะฑะปะฐะณะพะดะฐัั ัะพะผั, ััะพ ะพะฑะปะฐะดะฐัั ัะตะฟะฐัะฐัะธะฒะฝะพะน ัะธััะตะผะพะน, ะฒััะฒะปัััะตะน ะธย ัะดะฐะปัััะตะน ะฟะพะฒัะตะถะดะตะฝะฝัะต ะพะบะธัะปะตะฝะธะตะผ ะผะพะปะตะบัะปั, ะบะพัะพััะต ะทะฐัะตะผ ะทะฐะผะตะฝััััั. ะัะพะผะต ัะพะณะพ, ะบะปะตัะบะธ ะผะพะณัั ะฟะพะฒััะธัั ะฐะฝัะธะพะบัะธะดะฐะฝัะฝัั ะทะฐัะธัั ะฒย ะพัะฒะตั ะฝะฐย ััะธะปะตะฝะธะต ะะก. ะะดะฝะฐะบะพ ะฟัะธ ะฟะฐัะพะปะพะณะธัะตัะบะพะผ ะะก ะฒัะต ะผะพะปะตะบัะปั ะถะธะฒัั ะพัะณะฐะฝะธะทะผะพะฒ (ะปะธะฟะธะดั, ะฑะตะปะบะธ, ะฝัะบะปะตะธะฝะพะฒัะต ะบะธัะปะพัั ะธย ัะณะปะตะฒะพะดั) ััะฐะฝะพะฒัััั ะฟะพัะตะฝัะธะฐะปัะฝัะผะธ ะผะธัะตะฝัะผะธ ะพะบะธัะปะธัะตะปัะฝะพะณะพ ะฟะพะฒัะตะถะดะตะฝะธั [9โ11]. ะะตะทะฐะฒะธัะธะผะพ ะพัย ะฒััะฐะถะตะฝะฝะพััะธ ะธย ะฟัะธัะพะดั ะะก ะฟะตัะฒัะน ัะดะฐั ัะฐัะต ะฒัะตะณะพ ะฟัะธะฝะธะผะฐัั ะฒััะพะบะพัะฟะตัะธะฐะปะธะทะธัะพะฒะฐะฝะฝัะต ะธย ะฝะฐะธะฑะพะปะตะต ัะฝะตัะณะพะฟะพััะตะฑะปัััะธะต ะบะปะตัะบะธ. ะะฝะธ ััะฒััะฒะธัะตะปัะฝั ะดะฐะถะต ะบย ะผะธะฝะธะผะฐะปัะฝะพะผั ะฝะฐัััะตะฝะธั ะฟัะพัะตััะพะฒ ะผะธัะพั ะพะฝะดัะธะฐะปัะฝะพะณะพ ัะธะฝัะตะทะฐ ัะฝะตัะณะธะธ ะฒัะปะตะดััะฒะธะต ะะก, ะพัะตะฝั ะฑััััะพ ะธย ะทะฐะบะพะฝะพะผะตัะฝะพ ะฟัะธะฒะพะดััะตะผั ะบย ะธะทะผะตะฝะตะฝะธั ะบะปะตัะพัะฝะพะณะพ ะผะตัะฐะฑะพะปะธะทะผะฐ ะธย ัะฝะตัะณะตัะธัะตัะบะพะณะพ ะพะฑะผะตะฝะฐ. ะขะฐะบะธะผะธ ะบะปะตัะบะฐะผะธ ะฒย ะพัะณะฐะฝะธะทะผะต ัะตะปะพะฒะตะบะฐ ัะฒะปััััั ะฝะตะนัะพะฝั (ะฝะตัะฒะฝะฐั ัะธััะตะผะฐ), ะผะธะพัะธัั (ะผััะตัะฝะฐั ัะธััะตะผะฐ) ะธย ะฟะพะปะพะฒัะต ะบะปะตัะบะธ (ัะตะฟัะพะดัะบัะธะฒะฝะฐั ัะธััะตะผะฐ), ะฐย ัะฐะบะถะต ะฑััััะพ ะพะฑะฝะพะฒะปัััะธะตัั ะบะปะตัะบะธ ะบัะพะฒะธ ะธย ะบะพะถะฝะพะณะพ ะฟะพะบัะพะฒะฐ [8, 12]. ะะพั ะฟะพัะตะผั ะดะฐะถะต ะฝะตะทะฝะฐัะธัะตะปัะฝัะน ะฟะฐัะพะปะพะณะธัะตัะบะธะน ะะก ัะฟะพัะพะฑะตะฝ ะฟัะธะฒะตััะธ ะบย ัััะตััะฒะตะฝะฝะพะผั ัะฝะธะถะตะฝะธั ัะตััะธะปัะฝะพััะธ ัย ะฟัะฐะบัะธัะตัะบะธ ะทะดะพัะพะฒัั ะผัะถัะธะฝ [13โ15].
ะะบะธัะปะธัะตะปัะฝัะน ัััะตัั ะธย ัะฟะตัะผะฐัะพะณะตะฝะตะท ะฒย ะฝะพัะผะต ะธย ะฟัะธ ะผัะถัะบะพะผ ะฑะตัะฟะปะพะดะธะธ
ะย ะพัะฝะพะฒะฝัะผ ัะธะทะธะพะปะพะณะธัะตัะบะธะผ ะบะปะตัะพัะฝัะผ ะธััะพัะฝะธะบะฐะผ ะกะ ะฒย ััะบัะปััะต ะพัะฝะพััััั, ะบะฐะบ ะฝะธย ะฟะฐัะฐะดะพะบัะฐะปัะฝะพ, ัะฐะผะธ ัะฟะตัะผะฐัะพะทะพะธะดั, ะบะพัะพััะต ัย ัะฐะผัั ัะฐะฝะฝะธั ััะฐะฟะพะฒ ัะฟะตัะผะฐัะพะณะตะฝะตะทะฐ ัะฟะพัะพะฑะฝั ะฒััะฐะฑะฐััะฒะฐัั ะฝะตะฑะพะปััะธะต ะบะพะปะธัะตััะฒะฐ ะฐะบัะธะฒะฝัั ัะพัะผ ะบะธัะปะพัะพะดะฐ, ะฟัะตะดััะฐะฒะปัััะธั ัะพะฑะพะน ะฝะตย ััะพ ะธะฝะพะต ะบะฐะบ ะกะ [16]. ะญัะธ ะฐะบัะธะฒะฝัะต ะผะพะปะตะบัะปั ะฒะพะฒะปะตัะตะฝั ะฒย ะฟัะพัะตััั ะบะพะฝะดะตะฝัะฐัะธะธ ั ัะพะผะฐัะธะฝะฐ ะะะ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะธย ัะตะณัะปะธัััั ะบะพะปะธัะตััะฒะพ ะทะฐัะพะดััะตะฒัั ะบะปะตัะพะบ ะฟััะตะผ ะธะฝะดัะบัะธะธ ะฐะฟะพะฟัะพะทะฐ ะธะปะธ ะฟัะพะปะธัะตัะฐัะธะธ ะผะฐัะตัะธะฝัะบะธั ัะฟะตัะผะฐัะพะณะตะฝะฝัั ะบะปะตัะพะบย โ ัะฟะตัะผะฐัะพะณะพะฝะธะน [17]. ะย ะทัะตะปะพะผ ััะบัะปััะต ะฐะบัะธะฒะฝัะต ัะพัะผั ะบะธัะปะพัะพะดะฐ, ะธะปะธ ะกะ , ะธะณัะฐัั ะฒะฐะถะฝัั ัะพะปั ะฒย ะพะฑะตัะฟะตัะตะฝะธะธ ะฐะบัะพัะพะผะฐะปัะฝะพะน ัะตะฐะบัะธะธ, ััะฐะฑะธะปัะฝะพััะธ ะผะธัะพั ะพะฝะดัะธะฐะปัะฝะพะน ะะะ ะธย ะฟะพะดะฒะธะถะฝะพััะธ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ. ะะฝะธ ัะฐะบะถะต ะผะพะณัั ััะฝะบัะธะพะฝะธัะพะฒะฐัั ะบะฐะบ ัะธะณะฝะฐะปัะฝัะต ะผะพะปะตะบัะปั (ะฒัะพัะธัะฝัะต ะฟะพััะตะดะฝะธะบะธ ะบะปะตัะพัะฝะพะน ัะธะณะฝะฐะปะธะทะฐัะธะธ).
ะะทะฒะตััะฝะพ ะฝะตัะบะพะปัะบะพ ะผะตั ะฐะฝะธะทะผะพะฒ ะพะฑัะฐะทะพะฒะฐะฝะธั ะกะ ะฒย ะฝะพัะผะฐะปัะฝะพะผ ััะบัะปััะต. ะะตัะฒัะน ัะฒัะทะฐะฝ ัย ะฟัะธัััััะฒะธะตะผ ัะตัะผะตะฝัะฐัะธะฒะฝะพะณะพ ะบะพะผะฟะปะตะบัะฐ ัย ััะฐััะธะตะผ HAะะค-ะพะบัะธะดะฐะทั ะฒย ะฝะฐััะถะฝะพะน ะบะปะตัะพัะฝะพะน ะผะตะผะฑัะฐะฝะต ัะฟะตัะผะฐัะพะทะพะธะดะฐ, ะฒัะพัะพะนย โ ัย ัะฐะฑะพัะพะน ัะฐัะฟะพะปะพะถะตะฝะฝัั ะฒย ั ะฒะพััะพะฒะพะน ัะฐััะธ ัะฟะตัะผะฐัะพะทะพะธะดะฐ ะผะธัะพั ะพะฝะดัะธะน, ะบะพัะพััะต ะฟัะพะฟััะบะฐัั ัะปะตะบััะพะฝั ะธะทย ะดัั ะฐัะตะปัะฝะพะน ัะตะฟะธ ัย ััะฐััะธะตะผ ะะะ-ะพะบัะธะดะพัะตะดัะบัะฐะทั (ะดะธะฐัะพัะฐะทั). ะะพัะปะตะดะฝัั ัะตัะฝะพ ัะฒัะทะฐะฝะฐ ัย ะบัะฐะฝัะธะฝะพะบัะธะดะฐะทะพะน, ะบะพัะพัะฐั ะธะผะตะตััั ะบะฐะบ ะฒย ัะฟะตัะผะฐัะพะทะพะธะดะฐั , ัะฐะบ ะธย ะฒ ัะตะผะตะฝะฝะพะน ะฟะปะฐะทะผะต ะธย ัะฐะบะถะต ัะฟะพัะพะฑะฝะฐ ะธะฝะดััะธัะพะฒะฐัั ัะธะฝัะตะท ะฝะตะฑะพะปััะพะณะพ ะบะพะปะธัะตััะฒะฐ ัะฟะตัะผะฐะปัะฝัั ะกะ [18]. ะะปะตัะบะธ ััะบัะปััะฐ ัย ะฝะฐัััะตะฝะฝะพะน ะผะพััะพะปะพะณะธะตะน, ะฟัะตะธะผััะตััะฒะตะฝะฝะพ ัย ัะธัะพะฟะปะฐะทะผะฐัะธัะตัะบะธะผะธ ะฐะฝะพะผะฐะปะธัะผะธ, ัะบะฐะทัะฒะฐััะธะผะธ ะฝะฐย ะธั ะฝะตะทัะตะปะพััั ะธย ัะฝะธะถะตะฝะฝัะน ะฟะพัะตะฝัะธะฐะป ัะพะถะดะฐะตะผะพััะธ, ะฟัะพะธะทะฒะพะดัั ะฑะพะปััะต ะกะ , ัะตะผ ัะฟะตัะผะฐัะพะทะพะธะดั ัย ะฝะพัะผะฐะปัะฝะพะน ะผะพััะพะปะพะณะธัะตัะบะพะน ััััะบัััะพะน [19, 20]. ะกััะตััะฒัะตั ัะฐะบะถะต ัะฐะทะฝะธัะฐ ะผะตะถะดั ะบะพะปะธัะตััะฒะพะผ ะกะ , ะฟัะพะดััะธััะตะผัั ัะฟะตัะผะฐัะพะทะพะธะดะฐะผะธ ะฝะฐย ัะฐะทะฝัั ััะฐะดะธัั ัะพะทัะตะฒะฐะฝะธั [21].
ะัะต ะพะดะธะฝ ะธััะพัะฝะธะบ ะกะ ะฒย ััะบัะปััะตย โ ะปะตะนะบะพัะธัั, ะบะพัะพััะต ะฒย ัะธะทะธะพะปะพะณะธัะตัะบะธั ััะปะพะฒะธัั ะฟัะพะธะทะฒะพะดัั ะธั ะดะพ 1000 ัะฐะท ะฑะพะปััะต, ัะตะผ ัะฟะตัะผะฐัะพะทะพะธะดั. ะขะฐะบะพะน ะฒััะพะบะธะน ััะพะฒะตะฝั ะฟัะพะดัะบัะธะธ ะกะ ะฝะตะพะฑั ะพะดะธะผ ะดะปั ะทะฐัะธัั ะบะปะตัะพะบ ะพัย ะธะฝัะตะบัะธะน ะธย ะฒะพัะฟะฐะปะตะฝะธั. ะะบัะธะฒะธัะพะฒะฐะฝะฝัะต ะปะตะนะบะพัะธัั ะธะฝัะธะปัััะธัััั ะฟะพัะฐะถะตะฝะฝัะน ะพัะณะฐะฝ, ะฒัะดะตะปัั ะฑะพะปััะธะต ะบะพะปะธัะตััะฒะฐ ะกะ , ััะพ ะฒะตะดะตั ะบย ะพะณัะฐะฝะธัะตะฝะธั ะธย ะปะธะบะฒะธะดะฐัะธะธ ะฑะพะปััะธะฝััะฒะฐ ะธะฝัะตะบัะธะพะฝะฝัั ะฐะณะตะฝัะพะฒ [22]. ะะพะฒััะตะฝะฝะพะต ะบะพะปะธัะตััะฒะพ ะปะตะนะบะพัะธัะพะฒ (ะปะตะนะบะพัะธัะพัะฟะตัะผะธั) ะฝะฐย ัะพะฝะต ะดะพะบะฐะทะฐะฝะฝะพะณะพ ะฝะฐะปะธัะธั ะธะฝัะตะบัะธะพะฝะฝัั ะฐะณะตะฝัะพะฒ ะฒย ััะบัะปััะต (ะฑะฐะบัะตัะธะพัะฟะตัะผะธะธ) ะดะพััะพะฒะตัะฝะพ ะฐััะพัะธะธััะตััั ัย ัั ัะดัะตะฝะธะตะผ ะพะฟะปะพะดะพัะฒะพััััะตะน ัะฟะพัะพะฑะฝะพััะธ ััะบัะปััะฐ, ะฒย ัะฐััะฝะพััะธ ะฟัะธ ั ัะพะฝะธัะตัะบะธั ะธะฝัะตะบัะธะพะฝะฝัั ะฟัะพััะฐัะธัะฐั [23].
ะะดะฝะฐะบะพ ะปะตะนะบะพัะธัะพัะฟะตัะผะธั ะผะพะถะตั ะฑััั ัะฐะบะถะต ัะตะทัะปััะฐัะพะผ ะฒะพะทะดะตะนััะฒะธั ัะฐะบัะพัะพะฒ ะฝะตะธะฝัะตะบัะธะพะฝะฝะพะน ะฟัะธัะพะดั, ะฝะฐะฟัะธะผะตั ะดะปะธัะตะปัะฝะพะณะพ ัะตะบััะฐะปัะฝะพะณะพ ะฒะพะทะดะตัะถะฐะฝะธั ะธะปะธ ยซะบะพะผะฟัะตััะธะพะฝะฝะพะนยป ะฟะฐัะพะปะพะณะธะธ ะพัะณะฐะฝะพะฒ ะผะพัะพะฝะบะธ (ะฒะฐัะธะบะพัะตะปะต,ย ะณะธะดัะพัะตะปะต, ะฟะฐั ะพะฒะพ-ะผะพัะพะฝะพัะฝัะตย ะณััะถะธ). ะย ัะฐะบะธั ัะปััะฐัั ะฟัะธ ะผะธะบัะพะฑะธะพะปะพะณะธัะตัะบะธั ะธััะปะตะดะพะฒะฐะฝะธัั ััะบัะปััะฐ ะธะฝัะตะบัะธะพะฝะฝัะน ะฐะณะตะฝั ัะฐัะต ะฒัะตะณะพ ะฝะตย ะบัะปััะธะฒะธััะตััั [24โ27].
ะย ัะฟะตัะผะพะฟะปะฐะทะผะต ัััะตััะฒัะตั ัะฟะตัะผะฐะปัะฝะฐั ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะฐั ัะธััะตะผะฐ ะทะฐัะธัั. ะะฝะฐ ัะพััะพะธั ะบะฐะบ ะธะทย ัะตัะผะตะฝัะฝัั , ัะฐะบ ะธย ะฝะตัะตัะผะตะฝัะฝัั ัะฐะบัะพัะพะฒ ะธย ะฝะธะทะบะพะผะพะปะตะบัะปััะฝัั ัะพะตะดะธะฝะตะฝะธะน ัย ะฐะฝัะธะพะบัะธะดะฐะฝัะฝัะผ ะฟะพัะตะฝัะธะฐะปะพะผ ะธย ะพะฑะตัะฟะตัะธะฒะฐะตั ัะพั ัะฐะฝะตะฝะธะต ะพะฟัะธะผะฐะปัะฝะพะณะพ ะพะบัะธะดะฐัะธะฒะฝะพะณะพ ะฑะฐะปะฐะฝัะฐ ััะบัะปััะฐ. ะะปะฐะฒะฝัะน ะบะพะผะฟะพะฝะตะฝั ัะฟะตัะผะฐะปัะฝะพะน ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะพะน ัะธััะตะผั ะฝะฐะทัะฒะฐะตััั ยซัะตัะผะตะฝัะฐัะธะฒะฝะพะน ััะธะฐะดะพะนยป, ัะฐะบ ะบะฐะบ ัะพััะพะธั ะธะทย ััะตั ะบะปััะตะฒัั ัะตัะผะตะฝัะพะฒ-ะฐะฝัะธะพะบัะธะดะฐะฝัะพะฒ: ััะฟะตัะพะบัะธะดะดะธัะผััะฐะทั (ะกะะ), ะบะฐัะฐะปะฐะทั ะธย ะณะปััะฐัะธะพะฝ-ะฟะตัะพะบัะธะดะฐะทั [13].
ะกะะย โ ะผะตัะฐะปะป-ัะพะดะตัะถะฐัะธะต ัะตัะผะตะฝัั, ะบะฐัะฐะปะธะทะธััััะธะต ะดะธัะผััะฐัะธั ัะตะฐะบัะธะธ ััะฟะตัะพะบัะธะดะฝะพะณะพ ะฐะฝะธะพะฝะฐ ะธย ะฟัะธัััััะฒัััะธะต ะบะฐะบ ะฒะฝัััะธ ะบะปะตัะบะธ, ัะฐะบ ะธย ะฒะฝะต ะตะต. ะะฒะต ะฒะฝัััะธะบะปะตัะพัะฝัะต ัะพัะผั ะกะะ ะฟะตัะฒะพะณะพ ัะธะฟะฐ ัะพะดะตัะถะฐั ะผะตะดั ะธย ัะธะฝะบ (Cu-ะกะะ ะธย Zn-ะกะะ) ะธย ะปะพะบะฐะปะธะทััััั ะฒย ะพัะฝะพะฒะฝะพะผ ะฒย ัะธัะพะฟะปะฐะทะผะต ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ. ะะฝัััะธะบะปะตัะพัะฝะฐั ัะพัะผะฐ ะกะะ ะฒัะพัะพะณะพ ัะธะฟะฐ ัะพะดะตัะถะธั ะผะฐัะณะฐะฝะตั (Mn-ะกะะ) ะธย ะปะพะบะฐะปะธะทัะตััั ะฟัะตะธะผััะตััะฒะตะฝะฝะพ ะฒย ะฐะบัะธะฒะฝัั ัะตะฝััะฐั ะผะธัะพั ะพะฝะดัะธะน ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ. ะคะพัะผั ะกะะ ััะตััะตะณะพ ัะธะฟะฐ (ะฒะฝะตะบะปะตัะพัะฝัะต) ะผะพะณัั ะฝะฐั ะพะดะธัััั ะบะฐะบ ะฒย ัะฒะพะฑะพะดะฝะพะผ ะฒะธะดะต, ัะฐะบ ะธย ะฒ ัะฒัะทะฐะฝะฝะพะผ ัย ะฟะพะปะธัะฐั ะฐัะธะดะฐะผะธ ัะพััะพัะฝะธะธ. ะะพย ััััะบัััะต ะกะะ ััะตััะตะณะพ ัะธะฟะฐ ะฟะพั ะพะถะฐ ะฝะฐย ะกะะ ะฒัะพัะพะณะพ ัะธะฟะฐ, ะฝะพย ะฒะผะตััะพ ะผะฐัะณะฐะฝัะฐ ัะพะดะตัะถะธั ะฒย ะฐะบัะธะฒะฝะพะผ ัะตะฝััะต ะผะตะดั ะธย ัะธะฝะบ [28]. ะััะพะบะฐั ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะฐั ะฐะบัะธะฒะฝะพััั ัะฟะตัะผะพะฟะปะฐะทะผั ะฒย ะฝะพัะผะต ะฝะฐย 75% ะพะฑะตัะฟะตัะธะฒะฐะตััั ะกะะ ะฟะตัะฒะพะณะพ ัะธะฟะฐ ะธย ะฝะฐ 25%ย โ ะกะะ ััะตััะตะณะพ ัะธะฟะฐ, ะบะพัะพััะต ัะบะพัะตะต ะฒัะตะณะพ ัะธะฝัะตะทะธัััััั ะฒย ะฟัะตะดััะฐัะตะปัะฝะพะน ะถะตะปะตะทะต [29].
ะะฐัะฐะปะฐะทะฐ ััะบัะปััะฐ ะบะฐัะฐะปะธะทะธััะตั ัะฐะทะปะพะถะตะฝะธะต ะฟะตัะพะบัะธะดะฐ ะฒะพะดะพัะพะดะฐ ะฝะฐย ะผะพะปะตะบัะปััะฝัะน ะบะธัะปะพัะพะด ะธย ะฒะพะดั. ะฅะฐัะฐะบัะตัะฝะฐั ะพัะพะฑะตะฝะฝะพััั ััััะบัััั ะบะฐัะฐะปะฐะทัย โ ะฝะฐะปะธัะธะต ัะธััะตะผัย ะณะตะผะฐ ัย ัะตะฝััะฐะปัะฝะพ ัะฐัะฟะพะปะพะถะตะฝะฝัะผ ะฐัะพะผะพะผ ะถะตะปะตะทะฐ. ะััะพะบะฐั ะฐะบัะธะฒะฝะพััั ะบะฐัะฐะปะฐะทั ะฑัะปะฐ ะฒััะฒะปะตะฝะฐ ะฒย ะฟะตัะพะบัะธัะพะผะฐั , ะผะธัะพั ะพะฝะดัะธัั , ัะฝะดะพะฟะปะฐะทะผะฐัะธัะตัะบะพะผ ัะตัะธะบัะปัะผะต ะธย ัะธัะพะทะพะปะต ะผะฝะพะณะธั ัะธะฟะพะฒ ะบะปะตัะพะบ ะธย ัะบะฐะฝะตะน, ะฒะบะปััะฐั ัะตะผะตะฝะฝัั ะฟะปะฐะทะผั, ะฒย ะบะพัะพัะพะน ะตะต ะธััะพัะฝะธะบะพะผ ัะฒะปัะตััั ัะตะบัะตั ะฟัะตะดััะฐัะตะปัะฝะพะน ะถะตะปะตะทั [30].
ะขัะตัะธะน ัะฟะตัะธะฐะปะธะทะธัะพะฒะฐะฝะฝัะน ัะตัะผะตะฝัย โย ะณะปััะฐัะธะพะฝ-ะฟะตัะพะบัะธะดะฐะทะฐ ะบะฐัะฐะปะธะทะธััะตั ะฟัะพัะตััั ะฟัะตะฒัะฐัะตะฝะธั ะฟะตัะตะบะธัะธ ะฒะพะดะพัะพะดะฐ ะธย ะพัะณะฐะฝะธัะตัะบะธั ะฟะตัะพะบัะธะดะพะฒ, ะฒะบะปััะฐั ะฟะตัะพะบัะธะดั ัะพััะพะปะธะฟะธะดะพะฒ [28]. ะย ะฐะบัะธะฒะฝะพะผ ัะตะฝััะตย ะณะปััะฐัะธะพะฝ-ะฟะตัะพะบัะธะดะฐะทั ัะฐัะฟะพะปะฐะณะฐะตััั ัะตะปะตะฝ ะฒย ัะพัะผะต ัะตะปะตะฝะพัะธััะตะธะฝะฐ. ะคะตัะผะตะฝั ัะบัะฟัะตััะธััะตััั ะฟัะตะธะผััะตััะฒะตะฝะฝะพ ะฒย ะผะธัะพั ะพะฝะดัะธะฐะปัะฝะพะน ะผะฐััะธัะต ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ, ั ะพัั ัััะตััะฒัะตั ะธย ะตะณะพ ัะดะตัะฝะฐั ัะพัะผะฐ, ะทะฐัะธัะฐััะฐั ะะะ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะพัย ะะก [31]. ะะพย ะธะผะตััะธะผัั ะดะฐะฝะฝัะผ, ะธััะพัะฝะธะบะพะผย ะณะปััะฐัะธะพะฝ-ะฟะตัะพะบัะธะดะฐะทั ะฒย ัะฟะตัะผะพะฟะปะฐะทะผะต, ะบะฐะบ ะธย ะดััะณะธั ะฐะฝัะธะพะบัะธะดะฐะฝัะฝัั ัะฟะตัะธะฐะปะธะทะธัะพะฒะฐะฝะฝัั ัะตัะผะตะฝัะพะฒ, ัะฒะปัะตััั ะฟัะตะดััะฐัะตะปัะฝะฐั ะถะตะปะตะทะฐ [32].
ะัะพะผะต ัะพะณะพ, ะฒย ัะฟะตัะผะพะฟะปะฐะทะผะต ะฒย ะฝะพัะผะต ะฟัะธัััััะฒััั ะฝะตัะฟะตัะธะฐะปะธะทะธัะพะฒะฐะฝะฝัะต ัะตัะผะตะฝัั-ะฐะฝัะธะพะบัะธะดะฐะฝัั, ะบะพัะพััะต ะฒะทะฐะธะผะพะดะตะนััะฒััั ัะพย ัะฟะตัะธะฐะปะธะทะธัะพะฒะฐะฝะฝะพะน ยซัะตัะผะตะฝัะฝะพะน ััะธะฐะดะพะนยป ััะบัะปััะฐ ะธย ะฑะตะท ะบะพัะพััั ะฟะพะปะฝะพัะตะฝะฝะฐั ัะฐะฑะพัะฐ ัะฟะตัะผะฐะปัะฝะพะน ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะพะน ัะธััะตะผั ะฒย ัะตะปะพะผ ะฝะตะฒะพะทะผะพะถะฝะฐ:ย ะณะปััะฐัะธะพะฝ, ะฟะฐะฝัะพัะตะฝะพะฒะฐั ะบะธัะปะพัะฐ, ะบะพัะฝะทะธะผ Q10, L-ะบะฐัะฝะธัะธะฝ, ะฒะธัะฐะผะธะฝั A, E, C ะธย B, ะผะธะฝะตัะฐะปั (ัะธะฝะบ, ัะตะปะตะฝ, ั ัะพะผ, ะผะตะดั) [14, 33โ35].
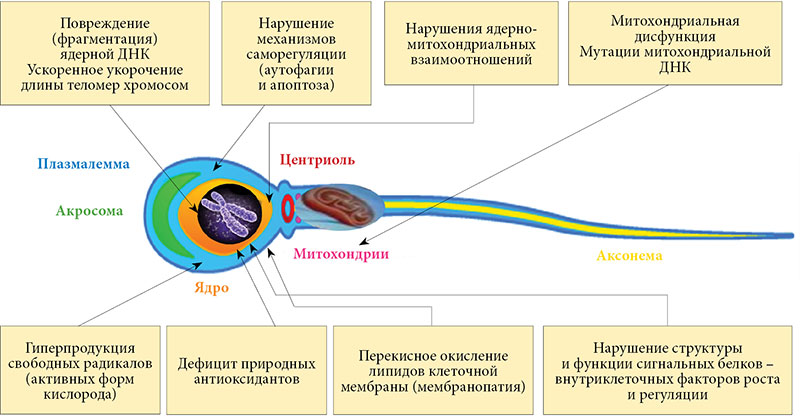
ะขะฐะบะธะผ ะพะฑัะฐะทะพะผ, ัะฟะตัะผะฐัะพะณะตะฝะตะท ะดะฐะถะต ัย ะทะดะพัะพะฒะพะณะพ ะผัะถัะธะฝั ะฝะตะธะทะฑะตะถะฝะพ ัะพะฟัะพะฒะพะถะดะฐะตััั ะพะฑัะฐะทะพะฒะฐะฝะธะตะผ ัะธะทะธะพะปะพะณะธัะตัะบะธ ะผะธะฝะธะผะฐะปัะฝะพะณะพ ะบะพะปะธัะตััะฒะฐ ะกะ , ะฝะตะพะฑั ะพะดะธะผัั ะดะปั ะฐะดะตะบะฒะฐัะฝะพะณะพ ะฟัะพัะตะบะฐะฝะธั ะฒัะตั ััะฐะฟะพะฒ ะพะฑัะฐะทะพะฒะฐะฝะธั ะธย ัะพะทัะตะฒะฐะฝะธั ะฟะพะปะพะฒัั ะบะปะตัะพะบ. ะะพะฝััะพะปั ะทะฐ ยซัะธะทะธะพะปะพะณะธัะฝะพััััยป ัะฟะตัะผะฐะปัะฝะพะณะพ ะะก ะพัััะตััะฒะปัะตั ัะพะพัะฒะตัััะฒัััะฐั ัะฟะตัะผะฐะปัะฝะฐั ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะฐั ัะธััะตะผะฐ, ะบะพัะพัะฐั ะพะดะฝะพะฒัะตะผะตะฝะฝะพ ะพะฑะตัะฟะตัะธะฒะฐะตั ะพะฟัะธะผะฐะปัะฝัะน ััะพะฒะตะฝั ะทะฐัะธัั ะพัย ะปัะฑะพะน ะธะทะฑััะพัะฝะพะน ะฟัะพะดัะบัะธะธ ะธะปะธ ััะตะทะผะตัะฝะพะน ะฐะบัะธะฒะฝะพััะธ ะพะฑัะฐะทัััะธั ัั ะฒย ััะบัะปััะต ะกะ . ะคัะฝะบัะธะพะฝะฐะปัะฝะฐั ะฝะตะดะพััะฐัะพัะฝะพััั ะปัะฑะพะณะพ ะธะทย ะบะพะผะฟะพะฝะตะฝัะพะฒ ัะฟะตัะผะฐะปัะฝะพะน ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะพะน ัะธััะตะผั ะผะพะถะตั ะฒัะทะฒะฐัั ัะฝะธะถะตะฝะธะต ะพะฑัะตะน ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะพะน ัะฟะพัะพะฑะฝะพััะธ ััะบัะปััะฐ. ะะฐะถะต ะฟัะธ ัะธะทะธะพะปะพะณะธัะตัะบะพะผ ััะพะฒะฝะต ะพะฑัะฐะทะพะฒะฐะฝะธั ะกะ ััะพ ะทะฝะฐะผะตะฝัะตั ัะฐะทะฒะธัะธะต ัะถะต ะฟะฐัะพะปะพะณะธัะตัะบะพะณะพ ัะฟะตัะผะฐะปัะฝะพะณะพ ะะกย โ ัะฝะธะฒะตััะฐะปัะฝะพะณะพ ะฟะฐัะพะณะตะฝะตัะธัะตัะบะพะณะพ ะผะตั ะฐะฝะธะทะผะฐ ะผัะถัะบะพะณะพ ะฑะตัะฟะปะพะดะธั ะฝะตะทะฐะฒะธัะธะผะพ ะพัย ะตะณะพ ะฟัะธัะธะฝั (ัะธั.ย 4) [33].
ะย ะฝะฐััะพััะตะต ะฒัะตะผั ะธะทะฒะตััะฝั ะผะฝะพะณะธะต ะบะปะตัะพัะฝะพ-ะผะพะปะตะบัะปััะฝัะต ะผะตั ะฐะฝะธะทะผั, ะฟะพััะตะดััะฒะพะผ ะบะพัะพััั ะฟะฐัะพะปะพะณะธัะตัะบะธะน ะะก ะฝะตะณะฐัะธะฒะฝะพ ะฒะปะธัะตั ะฝะฐย ะฒัะต ััะฐะฟั ัะฟะตัะผะฐัะพะณะตะฝะตะทะฐ, ะพัย ะฐะบัะธะฒะฐัะธะธ ัะฟะตัะผะฐัะพะณะพะฝะธะน ะดะพ ะพะบะพะฝัะฐัะตะปัะฝะพะณะพ ัะพะทัะตะฒะฐะฝะธั ัะฟะตัะผะธะตะฒ, ัััะตััะฒะตะฝะฝะพ ัะฝะธะถะฐั ะผัะถัะบัั ัะตััะธะปัะฝะพััั (ัะธั.ย 5) [36, 37].
ะะตัะผะพััั ะฝะฐย ัะฐะบะพะต ะผะฝะพะณะพะพะฑัะฐะทะธะต ะฒะฐัะธะฐะฝัะพะฒ ัััะตัั-ะธะฝะดััะธัะพะฒะฐะฝะฝัั ะบะปะตัะพัะฝัั ะฝะฐัััะตะฝะธะน, ะฝะฐะธะฑะพะปะตะต ะบัะธัะธัะตัะบะธะต ะฟะฐัะพะปะพะณะธัะตัะบะธะต ะธะทะผะตะฝะตะฝะธั, ะพะฑััะปะพะฒะปะตะฝะฝัะต ะฝะตะณะฐัะธะฒะฝัะผ ะฒะพะทะดะตะนััะฒะธะตะผ ะฟะฐัะพะปะพะณะธัะตัะบะพะณะพ ะะก ะฝะฐย ะฟะพะปะพะฒัะต ะบะปะตัะบะธ ะธย ะดะพััะพะฒะตัะฝะพ ัะฝะธะถะฐััะธะต ะพะฟะปะพะดะพัะฒะพัััััั ัะฟะพัะพะฑะฝะพััั ััะบัะปััะฐ, ะฟัะพะธัั ะพะดัั ะฒย ะผะตะผะฑัะฐะฝะฐั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ (ะผะตะผะฑัะฐะฝะพะฟะฐัะธั), ะผะธัะพั ะพะฝะดัะธัั (ะผะธัะพั ะพะฝะดัะธะฐะปัะฝะฐั ะดะธัััะฝะบัะธั ะธย ะผััะฐัะธะธ ะผะธัะพั ะพะฝะดัะธะฐะปัะฝะพะน ะะะ) ะธย ะณะตะฝะตัะธัะตัะบะพะผ ะฐะฟะฟะฐัะฐัะต ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ (ะผััะฐัะธะธ ะธย ััะฐะณะผะตะฝัะฐัะธั ัะดะตัะฝะพะน ะะะ) [38โ45].
ะกะพะฒัะตะผะตะฝะฝัะต ะฒะพะทะผะพะถะฝะพััะธ ะฟะฐัะพะณะตะฝะตัะธัะตัะบะพะน ะบะพััะตะบัะธะธ ะฟะฐัะพะปะพะณะธัะตัะบะพะณะพ ะพะบะธัะปะธัะตะปัะฝะพะณะพ ัััะตััะฐ ะฟัะธ ะผัะถัะบะพะผ ะฑะตัะฟะปะพะดะธะธ
ะย ะฝะฐััะพััะตะต ะฒัะตะผั ะฝะธย ัย ะบะพะณะพ ะฝะตย ะฒัะทัะฒะฐะตั ัะพะผะฝะตะฝะธะน ะฝะตะพะฑั ะพะดะธะผะพััั ะฟัะพะฒะตะดะตะฝะธั ะฑะฐะทะพะฒะพะน ะผะตัะฐะฑะพะปะธัะตัะบะพะน ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะพะน ัะตัะฐะฟะธะธ ะผัะถัะบะพะณะพ ะฑะตัะฟะปะพะดะธั ะฝะตะทะฐะฒะธัะธะผะพ ะพัย ะฝะตะฟะพััะตะดััะฒะตะฝะฝะพะน ะตะณะพ ะฟัะธัะธะฝั (ััะพะปะพะณะธัะตัะบะพะน, ัะฝะดะพะบัะธะฝะพะปะพะณะธัะตัะบะพะน, ะธะผะผัะฝะพะฒะพัะฟะฐะปะธัะตะปัะฝะพะน ะธะปะธ ะฟัะธ ะธั ัะพัะตัะฐะฝะธะธ), ะฐย ัะตะผ ะฑะพะปะตะต ะฟัะธ ะฝะตััะพัะฝะตะฝะฝัั (ะธะดะธะพะฟะฐัะธัะตัะบะธั ) ัะพัะผะฐั . ะัะพะผะต ัะพะณะพ, ัะฐะผะพ ะฟะพย ัะตะฑะต ะฟะพะฝััะธะต ยซะธะดะธะพะฟะฐัะธัะตัะบะพะต ะผัะถัะบะพะต ะฑะตัะฟะปะพะดะธะตยป ะฒะพะฒัะต ะฝะตย ะพะทะฝะฐัะฐะตั ะพััััััะฒะธั ัย ะผัะถัะธะฝั ัะฒะฝัั ะธะปะธ ัะบััััั ย ะณะพัะผะพะฝะฐะปัะฝะพ-ะผะตัะฐะฑะพะปะธัะตัะบะธั ะฟัะธัะธะฝ ะฝะฐัััะตะฝะธั ัะตะฟัะพะดัะบัะธะฒะฝะพะน ััะฝะบัะธะธ, ัะพะปั ะบะพัะพััั ัะฒะฝะพ ะฝะตะดะพะพัะตะฝะธะฒะฐะตััั. ะย ััะธะผ ะฟัะธัะธะฝะฐะผ ะพัะฝะพััััั ะฒัะต ัะปััะฐะธ ยซะฝะตะพะฑัััะฝะธะผะพะนยป ัย ัะพัะบะธ ะทัะตะฝะธั ััะพะปะพะณะฐ-ะฐะฝะดัะพะปะพะณะฐ ะฟะฐัะพัะฟะตัะผะธะธ ัย ะผะพะปะพะดัั ะผัะถัะธะฝ ัย ะพะถะธัะตะฝะธะตะผ, ะธะฝััะปะธะฝะพัะตะทะธััะตะฝัะฝะพัััั, ัะฐั ะฐัะฝัะผ ะดะธะฐะฑะตัะพะผ, ะผะตัะฐะฑะพะปะธัะตัะบะธะผ ัะธะฝะดัะพะผะพะผ, ะดะตัะธัะธัะพะผ ะฒะธัะฐะผะธะฝะฐ D,ย ะณะธะฟะพะณะพะฝะฐะดะธะทะผะพะผ, ะฝะฐัััะตะฝะธัะผะธ ัะฝะฐ (ะดะตัะธัะธั ะผะตะปะฐัะพะฝะธะฝะฐ) ะธย ะดััะณะธะผะธย ะณะพัะผะพะฝะฐะปัะฝะพ-ะผะตัะฐะฑะพะปะธัะตัะบะธะผะธ ัะธััะตะผะฝัะผะธ ะฝะฐัััะตะฝะธัะผะธ. ะฃะบะฐะทะฐะฝะฝัะต ะฝะฐัััะตะฝะธั ะผะพะณัั ะธะฝะดััะธัะพะฒะฐัั ัะธััะตะผะฝัะน ะธย ะปะพะบะฐะปัะฝัะน (ัะฟะตัะผะฐะปัะฝัะน) ะะก ะธย ะพะฑััะปะพะฒะปะธะฒะฐัั ะตะณะพ ะฝะตะธะทะฑะตะถะฝะพะต ะฟัะพะณัะตััะธัะพะฒะฐะฝะธะต ะฟัะธ ะพััััััะฒะธะธ ัะฒะพะตะฒัะตะผะตะฝะฝะพะน ะดะธะฐะณะฝะพััะธะบะธ ะธย ะผะฐะบัะธะผะฐะปัะฝะพ ัะฐะฝะฝะตะน ะฟะฐัะพะณะตะฝะตัะธัะตัะบะพะน ะบะพััะตะบัะธะธ.
ะัะธ ะฟะพะดะพะทัะตะฝะธะธ ะฝะฐย ัะบะพะฑั ะธะดะธะพะฟะฐัะธัะตัะบะพะต ะฑะตัะฟะปะพะดะธะต ะฒะฐะถะฝะพ ัะฐััะผะฐััะธะฒะฐัั ะผะตัะฐะฑะพะปะธะทะผ ะธย ะณะพะผะตะพััะฐะท ัะธััะตะผะฝะพ, ะฐย ะฝะต ัะฐะทะดะตะปััั ะพัะณะฐะฝะธะทะผ ะฝะฐย ะฟะพะปะพะฒัั ัะธััะตะผั ะธย ยซัะบัััะฐะณะตะฝะธัะฐะปัะฝััยป ะฟะฐัะพะปะพะณะธั [46โ51].
ะกะพะณะปะฐัะฝะพ ะฟะพัะปะตะดะฝะธะผ ะฟัะฑะปะธะบะฐัะธัะผ ะธย ะผะตัะฐะฐะฝะฐะปะธะทะฐะผ ะฟัะธะผะตะฝะตะฝะธะต ะฟัะตะฟะฐัะฐัะพะฒ ัย ะฐะฝัะธะพะบัะธะดะฐะฝัะฝัะผ ัััะตะบัะพะผ ะฒย ะบะฐัะตััะฒะต ัะผะฟะธัะธัะตัะบะพะน ัะตัะฐะฟะธะธ ะผัะถัะบะพะณะพ ะฑะตัะฟะปะพะดะธั ะธย ะฒ ัะฐะผะบะฐั ะฟะพะดะณะพัะพะฒะบะธ ะบย ะฟัะพัะพะบะพะปั ัะบัััะฐะบะพัะฟะพัะฐะปัะฝะพะณะพ ะพะฟะปะพะดะพัะฒะพัะตะฝะธั (ะญะะ) ะพะฑะพัะฝะพะฒะฐะฝะฝะพ ะธย ะฟะพะฒััะฐะตั ะฒะตัะพััะฝะพััั ะดะพััะธะถะตะฝะธั ะฑะตัะตะผะตะฝะฝะพััะธ ะธย ัะพะถะดะตะฝะธั ัะตะฑะตะฝะบะฐ ะฒย ะฟะฐัะต [52โ55]. ะย ัะฐะฑะปะธัะต ะฟัะธะฒะตะดะตะฝั ัะพะปัะบะพ ัะต ะปะตะบะฐัััะฒะตะฝะฝัะต ะธย ะฑะธะพะปะพะณะธัะตัะบะธ ะฐะบัะธะฒะฝัะต ะฒะตัะตััะฒะฐ, ะฟะพะปะพะถะธัะตะปัะฝัะน ัััะตะบั ะบะพัะพััั ัะถะต ะดะพะบะฐะทะฐะฝ ะธย ะบะพัะพััะต ัะตะบะพะผะตะฝะดััััั ะบย ะฟัะธะผะตะฝะตะฝะธั ะฒย ะบะฐัะตััะฒะต ัะผะฟะธัะธัะตัะบะพะน ัะตัะฐะฟะธะธ ะฑะตัะฟะปะพะดะธั ะธย ะฟะพะดะณะพัะพะฒะบะธ ะผัะถัะธะฝั ะบย ะฟัะพัะพะบะพะปั ะญะะ [56โ85].
L-ะบะฐัะฝะธัะธะฝ/ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝ: ัะธะทะธะพะปะพะณะธัะตัะบะฐั ัะพะปั ะฒย ะบะปะตัะพัะฝะพะผ ะผะตัะฐะฑะพะปะธะทะผะต
L-ะบะฐัะฝะธัะธะฝ, ะธะปะธ 3-ะฐะผะธะฝะพะผะฐัะปัะฝะฐั ะบะธัะปะพัะฐ,ย โ ะฟัะธัะพะดะฝะพะต ัะพะตะดะธะฝะตะฝะธะต, ะฐย ัะฐะบะถะต ะฟะพะปะธะฒะธัะฐะผะธะฝะฝะพะต ะฒะตัะตััะฒะพ, ะฝะตะพะฑั ะพะดะธะผะพะต ะดะปั ะฝะพัะผะฐะปัะฝะพะณะพ ะผะตัะฐะฑะพะปะธะทะผะฐ ะฟัะฐะบัะธัะตัะบะธ ะฒัะตั ะบะปะตัะพะบ ะพัะณะฐะฝะธะทะผะฐ. L-ะบะฐัะฝะธัะธะฝ ะฐะบัะธะฒะฝะพ ััะฐััะฒัะตั ะฒย ะบะปะตัะพัะฝะพะผ ะผะตัะฐะฑะพะปะธะทะผะต ะถะธัะฝัั ะบะธัะปะพั, ะฑัะดััะธ ัะฝะธะบะฐะปัะฝัะผ ะฟะตัะตะฝะพััะธะบะพะผ ะดะปะธะฝะฝะพัะตะฟะพัะตัะฝัั ะถะธัะฝัั ะบะธัะปะพั ะธะทย ะบะปะตัะพัะฝะพะณะพ ัะธัะพะทะพะปั ัะตัะตะท ะผะตะผะฑัะฐะฝั ะผะธัะพั ะพะฝะดัะธะน ะฒย ะบัะธะฟัั ะธั ะผะฐััะธะบัะฐ,ย ะณะดะต ะฟัะพะธัั ะพะดะธั ะธั ะดะฐะปัะฝะตะนัะธะน ะผะตัะฐะฑะพะปะธะทะผ, ะฝะฐะฟัะฐะฒะปะตะฝะฝัะน ะฝะฐย ัะธะฝัะตะท ัะฝะตัะณะธะธ (ะะขะค). ะัะฝะพะฒะฝัะต ััะฐะฟั ัะธะฝัะตะทะฐ ะะขะค ะฒย ะผะธัะพั ะพะฝะดัะธัั : ะผะพะฑะธะปะธะทะฐัะธั ะฐัะตัะธะป-ะะพะ (ะบะฐัะฝะธัะธะฝะพะฒัะน ัะธะบะป), ัะธะบะป ััะธะบะฐัะฑะพะฝะพะฒัั ะบะธัะปะพั (ัะธะบะป ะัะตะฑัะฐ) ะธย ะฟะตัะตะฝะพั ัะปะตะบััะพะฝะพะฒ ะฒย ะพะบะธัะปะธัะตะปัะฝะพะต ัะพััะพัะธะปะธัะพะฒะฐะฝะธะต (ะดัั ะฐัะตะปัะฝะฐั ัะตะฟั). ะะฐัะฝะธัะธะฝะพะฒัะน ัะธะบะปย โ ะฝะฐัะฐะปัะฝัะน ะฒะฐะถะฝะตะนัะธะน ััะฐะฟ ัะฝะตัะณะตัะธัะตัะบะพะณะพ ะผะตัะฐะฑะพะปะธะทะผะฐ ะบะปะตัะบะธ. ะะผะตะฝะฝะพ L-ะบะฐัะฝะธัะธะฝ ััะฐะฝัะฟะพััะธััะตั ะฝะตะพะฑั ะพะดะธะผัะต ะดะปั ะดะฐะปัะฝะตะนัะตะณะพ ยซัะณะพัะฐะฝะธัยป ะฒย ัะธะบะปะต ะัะตะฑัะฐ ะถะธัะฝัะต ะบะธัะปะพัั ะฒย ะผะฐััะธะบั ะผะธัะพั ะพะฝะดัะธะน,ย ะณะดะต ะฐะบัะธะฒะฝะพ ะฒะทะฐะธะผะพะดะตะนััะฒัะตั ัย ะฐัะตัะธะป-ะะพะ. ะย ัะตะทัะปััะฐัะต ะฟัะพะธัั ะพะดะธั ะฐัะตัะธะปะธัะพะฒะฐะฝะธะต L-ะบะฐัะฝะธัะธะฝะฐ (ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝ) ะธย ะพะฑัะฐะทัะตััั ะะพะย โ ะฒะฐะถะฝะตะนัะธะน ะบะพ-ัะตัะผะตะฝั ัะธะบะปะฐ ะัะตะฑัะฐ. ะัะธ ััะพะผ ะฒย ะผะธัะพั ะพะฝะดัะธัั ะดะพะปะถะฝะพ ะพะฑัะทะฐัะตะปัะฝะพ ัะพั ัะฐะฝััััั ะดะธะฝะฐะผะธัะตัะบะพะต ะผะตัะฐะฑะพะปะธัะตัะบะพะต ัะฐะฒะฝะพะฒะตัะธะต ะฒย ัะธััะตะผะต ยซะฐัะตัะธะป-ะะพะ + L-ะบะฐัะฝะธัะธะฝ <=> ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝ + ะะพะยป. ะญัะฐ ัะธััะตะผะฐย โ ะพะดะฝะฐ ะธะทย ะพัะฝะพะฒะฝัั ะฐัะตัะธะปัะฝัั ะฑััะตัะฝัั ัะธััะตะผ ะบะปะตัะบะธ, ะฟะพะดะดะตัะถะธะฒะฐััะธั ัะตะณะตะฝะตัะฐัะธั ะผะธัะพั ะพะฝะดัะธะน [86].
ะกะฐะผัะต ะฒััะพะบะธะต ะบะพะฝัะตะฝััะฐัะธะธ L-ะบะฐัะฝะธัะธะฝะฐ ะฒย ะผัะถัะบะพะน ัะตะฟัะพะดัะบัะธะฒะฝะพะน ัะธััะตะผะต ะพะฑะฝะฐััะถะตะฝั ะฒย ะฟัะธะดะฐัะบะฐั ัะธัะตะบ (ะฒย 2000 ัะฐะท ะฒััะต ะตะณะพ ะบะพะฝัะตะฝััะฐัะธะธ ะฒย ัะตะปัะฝะพะน ะบัะพะฒะธ) [87โ89]. ะััะพะบะธะน ััะพะฒะตะฝั ะถะธัะฝัั ะบะธัะปะพั ะฒย ะฟัะธะดะฐัะบะฐั ัะธัะตะบย โ ัะตะทัะปััะฐั ะฐะบัะธะฒะฝะพะณะพ ัะตะบัะตัะพัะฝะพะณะพ ะฟัะพัะตััะฐ. ะย ะธััะปะตะดะพะฒะฐะฝะธัั ะฟะพะบะฐะทะฐะฝะฐ ะฟะพะปะพะถะธัะตะปัะฝะฐั ัะฒัะทั ะผะตะถะดั ะฝะฐัะฐะปัะฝัะผ ะดะฒะธะถะตะฝะธะตะผ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ, ะฟะพะฒััะตะฝะฝัะผ ััะพะฒะฝะตะผ ะถะธัะฝัั ะบะธัะปะพั ะฒย ะฟัะธะดะฐัะบะฐั ัะธัะตะบ ะธย ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะพะผ ะฒย ััะบัะปััะต [90โ93]. ะขะฐะบะธะผ ะพะฑัะฐะทะพะผ, ะพัย ะฝะพัะผะฐะปัะฝะพะณะพ ััะพะฒะฝั L-ะบะฐัะฝะธัะธะฝะฐ ะธย ะพะฑัะฐะทัััะตะณะพัั ะธะทย ะฝะตะณะพ ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ัััะตััะฒะตะฝะฝะพ ะทะฐะฒะธัะธั ัะฝะตัะณะตัะธัะตัะบะธะน ะบะปะตัะพัะฝัะน ะพะฑะผะตะฝ. ะัะธ ััะพะผ ัะพะปัะบะพ ะพะบะพะปะพ 30% L-ะบะฐัะฝะธัะธะฝะฐ ะฒย ะพัะณะฐะฝะธะทะผะต ะธะผะตะตั ัะฝะดะพะณะตะฝะฝะพะต ะฟัะพะธัั ะพะถะดะตะฝะธะต, ะฐย 70% ะฟะพัััะฟะฐะตั ัย ะฟะธัะตะน. ะะผะตะฝะฝะพ ะฐะปะธะผะตะฝัะฐัะฝัะน ะดะตัะธัะธั L-ะบะฐัะฝะธัะธะฝะฐ ะฒะพ ะผะฝะพะณะพะผ ะฟัะตะดะพะฟัะตะดะตะปัะตั ะฒะพะทะผะพะถะฝะพััั ัะฐะทะฒะธัะธั ะฝะฐัััะตะฝะธะน ะบะปะตัะพัะฝะพะณะพ ัะฝะตัะณะตัะธัะตัะบะพะณะพ ะพะฑะผะตะฝะฐ ะฒย ะปัะฑะพะผ ะฒะพะทัะฐััะต ะธย ะฟัะฐะบัะธัะตัะบะธ ะฟัะธ ะปัะฑะพะผ ะทะฐะฑะพะปะตะฒะฐะฝะธะธ, ะฒะบะปััะฐั ะฑะตัะฟะปะพะดะธะต [94].
ะะปะธัะฝะธะต ะฟัะตะฟะฐัะฐัะพะฒ L-ะบะฐัะฝะธัะธะฝะฐ/ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะฝะฐย ะผะฐะบัะพั ะฐัะฐะบัะตัะธััะธะบะธ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ
ะ ะตะทัะปััะฐัั ะผะฝะพะณะพัะธัะปะตะฝะฝัั ัะบัะฟะตัะธะผะตะฝัะฐะปัะฝัั ะธััะปะตะดะพะฒะฐะฝะธะน, ะฟะพะดัะฒะตัะถะดะตะฝะฝัะต ะฑะพะปััะธะผ ะบะปะธะฝะธัะตัะบะธะผ ะพะฟััะพะผ, ัะฒะธะดะตัะตะปัััะฒััั ะพย ะฒััะพะบะพะน ัััะตะบัะธะฒะฝะพััะธ ะฟัะธะผะตะฝะตะฝะธั L-ะบะฐัะฝะธัะธะฝะฐ/ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะฒย ะบะพะฝัะตัะฒะฐัะธะฒะฝะพะน ัะตัะฐะฟะธะธ ะผัะถัะบะพะณะพ ะฑะตัะฟะปะพะดะธั, ะฒะบะปััะฐั ะธะดะธะพะฟะฐัะธัะตัะบัั ะฟะฐัะพะทะพะพัะฟะตัะผะธั. ะะฐะฝะฝะฐั ะบะพะผะฑะธะฝะฐัะธั ะดะพััะพะฒะตัะฝะพ ะฟัะธะฒะพะดะธั ะบย ัะฒะตะปะธัะตะฝะธั ะพะฑัะตะณะพ ะบะพะปะธัะตััะฒะฐ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะธย ะผะพััะพะปะพะณะธัะตัะบะธ ะฝะพัะผะฐะปัะฝัั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะฒย ััะบัะปััะต, ะฟะพะฒััะตะฝะธั ะธั ะพะฑัะตะน ะธย ะฟัะพะณัะตััะธะฒะฝะพะน ะฟะพะดะฒะธะถะฝะพััะธ (ะทะฐ ััะตั ัะปัััะตะฝะธั ะผะธัะพั ะพะฝะดัะธะฐะปัะฝะพะณะพ ัะธะฝัะตะทะฐ ัะฝะตัะณะธะธ, ะฝะตะพะฑั ะพะดะธะผะพะน ะดะปั ะฐะบัะธะฒะฝะพะน ะบะธะฝะตัะธะบะธ), ะฐย ัะฐะบะถะต ัะฟะพัะพะฑััะฒัะตั ะฟะพะฒััะตะฝะธั ยซะฒัะถะธะฒะฐะตะผะพััะธยป ะฟะพะปะพะฒัั ะบะปะตัะพะบ [95].
ะย ััะดะต ะดะฒะพะนะฝัั ัะปะตะฟัั ะบะพะฝััะพะปะธััะตะผัั ะบะปะธะฝะธัะตัะบะธั ะธััะปะตะดะพะฒะฐะฝะธะน ะฒััะฒะปะตะฝะฐ ะดะพััะพะฒะตัะฝะฐั ะฟะพะปะพะถะธัะตะปัะฝะฐั ัะฒัะทั ะผะตะถะดั ััะพะฒะฝัะผะธ L-ะบะฐัะฝะธัะธะฝะฐ, ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะธย ะฟะพะดะฒะธะถะฝะพัััั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ัย ะฑะตัะฟะปะพะดะฝัั ะผัะถัะธะฝ. ะัะธ ััะพะผ ัะฒัะทั ะฑัะปะฐ ะฒััะฐะถะตะฝะฝะตะต ัย ะผัะถัะธะฝ ัย ะฑะพะปะตะต ะฝะธะทะบะพะน ะธัั ะพะดะฝะพะน ะฟะพะดะฒะธะถะฝะพัััั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ [60, 96].
G. Balercia ะธย ัะพะฐะฒั. (2005) ะพัะตะฝะธะปะธ ะฒะปะธัะฝะธะต L-ะบะฐัะฝะธัะธะฝะฐ, ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะธย ะธั ะบะพะผะฑะธะฝะฐัะธะธ ะฝะฐย ะบะธะฝะตัะธะบั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะธย ะพะฑััั ัะฟะพัะพะฑะฝะพััั ะฟะพะณะปะพัะตะฝะธั ัะฒะพะฑะพะดะฝัั ัะฐะดะธะบะฐะปะพะฒ ะบะธัะปะพัะพะดะฐ. ะย ัะฐะฝะดะพะผะธะทะธัะพะฒะฐะฝะฝะพะผ ะดะฒะพะนะฝะพะผ ัะปะตะฟะพะผ ะบะพะฝััะพะปะธััะตะผะพะผ ะธััะปะตะดะพะฒะฐะฝะธะธ ะฟัะธะฝัะปะธ ััะฐััะธะต 60 ะผัะถัะธะฝ ัย ะธะดะธะพะฟะฐัะธัะตัะบะพะน ะฐััะตะฝะพัะตัะฐัะพะทะพะพัะฟะตัะผะธะตะน. ะ ะตะทัะปััะฐัั ัะตััะธะผะตัััะฝะพะน ัะตัะฐะฟะธะธ ะฟะพะบะฐะทะฐะปะธ ะฒััะพะบัั ัััะตะบัะธะฒะฝะพััั ะฟัะตะฟะฐัะฐัะพะฒ L-ะบะฐัะฝะธัะธะฝะฐ ะธย ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ. ะะพ ะฒัะตะผั ัะตัะฐะฟะธะธ ะฒย ะฟะฐัะฐั ,ย ะณะดะต ะผัะถัะธะฝั ะฟัะธะฝะธะผะฐะปะธ L-ะบะฐัะฝะธัะธะฝ, ะฑัะปะพ ะทะฐัะตะณะธัััะธัะพะฒะฐะฝะพ ะดะตะฒััั ะฑะตัะตะผะตะฝะฝะพััะตะน ะธย ะตัะต ะฟััั ะฑะตัะตะผะตะฝะฝะพััะตะน ะฒย ะฟะฐัะฐั ,ย ะณะดะต ะผัะถัะธะฝั ะฟะพะปััะฐะปะธ ะบะพะผะฑะธะฝะธัะพะฒะฐะฝะฝัั ัะตัะฐะฟะธั L-ะบะฐัะฝะธัะธะฝะพะผ ะธย ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะพะผ [97].
A. Garolla ะธย ัะพะฐะฒั. (2005) ะธะทััะฐะปะธ ัััะตะบัะธะฒะฝะพััั L-ะบะฐัะฝะธัะธะฝะฐ ัย 30 ะผัะถัะธะฝ ัย ะฐััะตะฝะพัะตัะฐัะพัะฟะตัะผะธะตะน ะฒย ัะฐะผะบะฐั ะดะฒะพะนะฝะพะณะพ ัะปะตะฟะพะณะพ ะธััะปะตะดะพะฒะฐะฝะธั. ะะฐัะธะตะฝัั ะฟะพะปััะฐะปะธ ะฟะปะฐัะตะฑะพ ะฒย ัะตัะตะฝะธะต ััะตั ะผะตัััะตะฒ, ะฐย ะทะฐัะตะผ L-ะบะฐัะฝะธัะธะฝ ะฒย ัััะพัะฝะพะน ะดะพะทะต 2ย ะณย ะตะถะตะดะฝะตะฒะฝะพ ะฒย ัะตัะตะฝะธะต ััะตั ะผะตัััะตะฒ. ะัะปะพ ะพัะผะตัะตะฝะพ ัะปัััะตะฝะธะต ะฟะพะดะฒะธะถะฝะพััะธ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะฒย ะณััะฟะฟะต ะผัะถัะธะฝ ัย ะฝะพัะผะฐะปัะฝัะผ ััะพะฒะฝะตะผย ะณะปััะฐัะธะพะฝ-ะฟะตัะพะบัะธะดะฐะทะฝะพะน ะฐะบัะธะฒะฝะพััะธ [98].
ะะพะทะธัะธะฒะฝัะต ัะตะทัะปััะฐัั ะฟัะธะผะตะฝะตะฝะธั ะบะพะผะฟะปะตะบัะฐ L-ะบะฐัะฝะธัะธะฝะฐ ะธย ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะฒย ะปะตัะตะฝะธะธ ะฑะตัะฟะปะพะดะธั ัย ะผัะถัะธะฝ ะฑัะปะธ ะฟะพะปััะตะฝั ะฒย ััะดะต ะฝะตะดะฐะฒะฝะพ ะฟัะพะฒะตะดะตะฝะฝัั ะบััะฟะฝัั ัะพััะธะนัะบะธั ะบะปะธะฝะธัะตัะบะธั ะธััะปะตะดะพะฒะฐะฝะธะน [99โ102].
ะ.ะ. ะะธั ะฐะนะปะธัะตะฝะบะพ ะธย ัะพะฐะฒั. (2014) ะดะพััะพะฒะตัะฝะพ ัััะฐะฝะพะฒะธะปะธ, ััะพ ัะตัะตะท ััะธ ะผะตัััะฐ ะฟัะธะผะตะฝะตะฝะธั ะบะพะผะฟะปะตะบัะฐ L-ะบะฐัะฝะธัะธะฝะฐ ะธย ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ (ะกะฟะตัะผะฐะบัะธะฝยฎ) ะดะพะปั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ัย ะฟะพัััะฟะฐัะตะปัะฝัะผ ะดะฒะธะถะตะฝะธะตะผ (ะ+ะ) ะธย ะดะพะปั ะผะพััะพะปะพะณะธัะตัะบะธ ะฝะพัะผะฐะปัะฝัั ัะพัะผ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ัะฒะตะปะธัะธะปะธัั ะฑะพะปะตะต ัะตะผ ะฒย ะดะฒะฐ ัะฐะทะฐ, ะบะพะฝัะตะฝััะฐัะธั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะฒย ััะบัะปััะต ะฒััะพัะปะฐ ะฒย 2,3 ัะฐะทะฐ [101].
ะ.ะ. ะะธะฝะพะณัะฐะดะพะฒ ะธย ัะพะฐะฒั. (2014) ะพัะตะฝะธะฒะฐะปะธ ะธะทะผะตะฝะตะฝะธั ะฒย ะผะพััะพะปะพะณะธะธ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะฟะพัะปะต ััะตั ะผะตัััะฝะพะณะพ ะฟัะธะผะตะฝะตะฝะธั ะบะพะผะฟะปะตะบัะฐ ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะธย L-ะบะฐัะฝะธัะธะฝะฐ ััะผะฐัะฐัะฐ ะฒย ะบะพะผะฑะธะฝะธัะพะฒะฐะฝะฝะพะน ัะตัะฐะฟะธะธ ัย ะดััะณะธะผะธ ะฟัะตะฟะฐัะฐัะฐะผะธ. ะะพะปะพะถะธัะตะปัะฝะฐั ะดะธะฝะฐะผะธะบะฐ ะฝะฐะฑะปัะดะฐะปะฐัั ัย 53,2% ะฟะฐัะธะตะฝัะพะฒ, ะทะฝะฐัะธะผัั ะธะทะผะตะฝะตะฝะธะน ะฝะตย ะฟัะพะธะทะพัะปะพ ัย 44,3% ะฟะฐัะธะตะฝัะพะฒ, ะพััะธัะฐัะตะปัะฝะฐั ะดะธะฝะฐะผะธะบะฐ ะทะฐัะธะบัะธัะพะฒะฐะฝะฐ ัย 1,5% ะฑะพะปัะฝัั . ะะพััะตะปััะธะพะฝะฝัะน ะฐะฝะฐะปะธะท ะฝะตย ะฒััะฒะธะป ะดะพััะพะฒะตัะฝะพะน ัะฒัะทะธ ะธะทะผะตะฝะตะฝะธั ะผะพััะพะปะพะณะธะธ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ัย ะพััะฐะปัะฝัะผะธ ะฟะพะบะฐะทะฐัะตะปัะผะธ ัะฟะตัะผะพะณัะฐะผะผั. ะะฝะฐะปะธะท ะฟะพย ัะพะฟะพััะฐะฒะปะตะฝะธั ะธะทะผะตะฝะตะฝะธะน ะดะพะปะธ ะผะพััะพะปะพะณะธัะตัะบะธ ะฝะพัะผะฐะปัะฝัั ัะพัะผ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ัย ัะธัะปะพะผ ะฑะตัะตะผะตะฝะฝะพััะตะน ัย ะฟะฐััะฝะตัั ะฟะฐัะธะตะฝัะพะฒ ะฟะพะบะฐะทะฐะป, ััะพ ะฒัะต ะฑะตัะตะผะตะฝะฝะพััะธ ะฝะฐัััะฟะฐะปะธ ะฟัะธ ะผะธะฝะธะผะฐะปัะฝะพะผ ะทะฝะฐัะตะฝะธะธ ะผะพััะพะปะพะณะธะธ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะฒย 8% [102].
ะะพย ะดะฐะฝะฝัะผ ะดััะณะพะณะพ ะธััะปะตะดะพะฒะฐะฝะธั, ะบัะฐัะบะพะฒัะตะผะตะฝะฝะพะต ะฟัะธะผะตะฝะตะฝะธะต L-ะบะฐัะฝะธัะธะฝะฐ ะผะพะถะตั ะฟะพะปะพะถะธัะตะปัะฝะพ ะฟะพะฒะปะธััั ะฝะฐย ะบะพะปะธัะตััะฒะพ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะธย ะฟัะธะฒะตััะธ ะบย ััะฟะตัะฝะพะน ะฑะตัะตะผะตะฝะฝะพััะธ ะฟัะธ ะฟัะพัะตะดััะต ะธะฝััะฐัะธัะพะฟะปะฐะทะผะฐัะธัะตัะบะพะน ะธะฝัะตะบัะธะธ ัะฟะตัะผะฐัะพะทะพะธะดะฐ ะฒย ัะนัะตะบะปะตัะบั [103].
ะขะฐะบะธะผ ะพะฑัะฐะทะพะผ, ะฟะพะทะธัะธะฒะฝัะต ัััะตะบัั L-ะบะฐัะฝะธัะธะฝะฐ/ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะฒย ะพัะฝะพัะตะฝะธะธ ะบะธะฝะตะผะฐัะธัะตัะบะธั ะธย ะผะฐะบัะพะผะพััะพะปะพะณะธัะตัะบะธั ั ะฐัะฐะบัะตัะธััะธะบ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ัััะฐะฝะพะฒะปะตะฝั ะดะพััะฐัะพัะฝะพ ะดะฐะฒะฝะพ ะธย ั ะพัะพัะพ ะธะทััะตะฝั ะฒย ะธััะปะตะดะพะฒะฐะฝะธัั , ะฒัะฟะพะปะฝะตะฝะฝัั ะฝะฐย ะฒััะพะบะพะผ ะดะพะบะฐะทะฐัะตะปัะฝะพะผ ััะพะฒะฝะต [59, 104, 105].
ะะปะธัะฝะธะต ะฟัะตะฟะฐัะฐัะพะฒ L-ะบะฐัะฝะธัะธะฝะฐ/ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะฝะฐย ะผะธะบัะพั ะฐัะฐะบัะตัะธััะธะบะธ (ััะฐะณะผะตะฝัะฐัะธั ะะะ) ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ
ะย ะฟะพัะปะตะดะฝะธะตย ะณะพะดั ะฑัะปะธ ะฟัะพะฒะตะดะตะฝั ะบะปะธะฝะธะบะพ-ัะบัะฟะตัะธะผะตะฝัะฐะปัะฝัะต ะธััะปะตะดะพะฒะฐะฝะธั, ะฒย ะบะพัะพััั ััะตะดะธ ะฟัะพัะธั ัััะธะฝะฝัั ััะฐะฝะดะฐััะฝัั ะฟะฐัะฐะผะตััะพะฒ ััะบัะปััะฐ (ะบะพะฝัะตะฝััะฐัะธั, ะฟะพะดะฒะธะถะฝะพััั ะธย ะผะฐะบัะพะผะพััะพะปะพะณะธั) ัะตะปะตะฝะฐะฟัะฐะฒะปะตะฝะฝะพ ะธะทััะฐะปัั ัะฟะตัะผะฐะปัะฝัะน ะะก. ะย ัะฐััะฝะพััะธ, ะพัะตะฝะธะฒะฐะปะธัั ัะฐััะพัะฐ ะธย ััะตะฟะตะฝั ะฒััะฐะถะตะฝะฝะพััะธ ััะฐะณะผะตะฝัะฐัะธะธ ะะะ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ัย ะฟะพะผะพััั ัะพะพัะฒะตัััะฒัััะธั ัะตัั-ัะธััะตะผ (TUNEL-ัะตััะฐ). ะะบะฐะทะฐะปะพัั, ััะพ ะบัะพะผะต ัะถะต ะธะทะฒะตััะฝะพะณะพ ัััะตะบัะฐย โ ัะปัััะตะฝะธะต ัะฝะตัะณะตัะธัะตัะบะพะณะพ ะพะฑะผะตะฝะฐ ะฒย ัะฟะตัะผะฐัะพะทะพะธะดะฐั (ะปะธะบะฒะธะดะฐัะธั ัะฟะตัะผะฐะปัะฝะพะน ะผะธัะพั ะพะฝะดัะธะฐะปัะฝะพะน ะดะธัััะฝะบัะธะธ), L-ะบะฐัะฝะธัะธะฝ/ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝ ะดะพะฟะพะปะฝะธัะตะปัะฝะพ ะฝะธะฒะตะปะธัะพะฒะฐะป ะฒะปะธัะฝะธะต ะฟะฐัะพะปะพะณะธัะตัะบะพะณะพ ัะฟะตัะผะฐะปัะฝะพะณะพ ะะก, ะฟะพะฒััะฐั ัััะพะนัะธะฒะพัััย ะณะตะฝะตัะธัะตัะบะพะณะพ ะฐะฟะฟะฐัะฐัะฐ (ะะะ) ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ. ะญัะพ ะฟัะพัะฒะปัะปะพัั ัะฝะธะถะตะฝะธะตะผ ัะฐััะพัั ะธย ััะตะฟะตะฝะธ ะฒััะฐะถะตะฝะฝะพััะธ ััะฐะณะผะตะฝัะฐัะธะธ ะะะ. ะขะฐะบ, ะฟะพย ัะบัะฟะตัะธะผะตะฝัะฐะปัะฝัะผ ะดะฐะฝะฝัะผ, L-ะบะฐัะฝะธัะธะฝ ะฟัะธ ะดะพะฑะฐะฒะปะตะฝะธะธ ะฒย ะพั ะปะฐะถะดะตะฝะฝัะน ััะบัะปัั ะถะธะฒะพัะฝัั ะฑะพะปะตะต ัััะตััะฒะตะฝะฝะพ (ะฝะฐย 15,6%) ัะฝะธะถะฐะป ะฒััะฐะถะตะฝะฝะพััั ะฟะพะฒัะตะถะดะตะฝะธะน ะะะ ะพะบะธัะปะธัะตะปัะฝะพะน ะฟัะธัะพะดั (ััะฐะณะผะตะฝัะฐัะธั) ะฟะพย ััะฐะฒะฝะตะฝะธั ัย ะฟะธััะฒะฐัะพะผ (ะฝะฐย 9,0% ัะพะพัะฒะตัััะฒะตะฝะฝะพ). ะญัะพ ะฟะพะทะฒะพะปัะตั ัะฐััะผะฐััะธะฒะฐัั L-ะบะฐัะฝะธัะธะฝ ะบะฐะบ ะฟะพัะตะฝัะธะฐะปัะฝัะน ัััะตะบัะธะฒะฝัะน ัะฟะตัะผะฐะปัะฝัะน ะบะพะฝัะตัะฒะฐะฝั [106]. ะย ัะบัะฟะตัะธะผะตะฝัะฐั ัะฐะบะถะต ะฟะพะบะฐะทะฐะฝะฐ ะฒััะพะบะฐั ัััะตะบัะธะฒะฝะพััั L-ะบะฐัะฝะธัะธะฝะฐ ะฒย ะทะฐัะธัะต ะพัย ะฒะพะทะดะตะนััะฒะธั ะฐะบัะธะปะฐะผะธะดะฐย โ ัะธะฝัะตัะธัะตัะบะพะณะพ ะผะฐัะตัะธะฐะปะฐ, ะบะพัะพััะน ัะธัะพะบะพ ะฟัะธะผะตะฝัะตััั ะธย ะพะฑะปะฐะดะฐะตั ะพะฟัะตะดะตะปะตะฝะฝัะผะธ ะผััะฐะณะตะฝะฝัะผะธ ัััะตะบัะฐะผะธ [107].
ะญะบัะฟะตัะธะผะตะฝัั ัย ััะบัะปััะพะผ ัะฐะผัะพะฒ ะบััั ัะฒะธะดะตัะตะปัััะฒััั ะพย ัะพะผ, ััะพ ะฟัะตะดะฒะฐัะธัะตะปัะฝะพะต ะฟัะธะผะตะฝะตะฝะธะต L-ะบะฐัะฝะธัะธะฝะฐ ะดะพััะพะฒะตัะฝะพ ะฟะพะฒััะฐะตั ัััะพะนัะธะฒะพััั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะบย ะฒะพะทะดะตะนััะฒะธั ัะฐะทะปะธัะฝัั ะปะตะบะฐัััะฒะตะฝะฝัั ัะตััะธะบัะปััะฝัั ัะพะบัะธะฝะพะฒ, ะฒย ัะฐััะฝะพััะธ ะพะฝะบะพะปะพะณะธัะตัะบะธั ะฟัะตะฟะฐัะฐัะพะฒ. ะญัะพั ัะฐะบั ะบัะฐะนะฝะต ะฒะฐะถะตะฝ ะดะปั ัะตัะตะฝะธั ะฐะบััะฐะปัะฝะพะน ะฟัะพะฑะปะตะผั ัะพั ัะฐะฝะตะฝะธั ัะตััะธะปัะฝะพััะธ ัย ะผัะถัะธะฝ ัย ะพะฝะบะพะปะพะณะธัะตัะบะพะน ะฟะฐัะพะปะพะณะธะตะน, ะฟะพัะบะพะปัะบั ะฟัะธะตะผ ั ะธะผะธะพะฟัะตะฟะฐัะฐัะพะฒ ะฒย ะฒะธะดะต ะผะพะฝะพัะตัะฐะฟะธะธ ะธะปะธ ะบะฐะบ ะบะพะผะฟะพะฝะตะฝัะฐ ะบะพะผะฑะธะฝะธัะพะฒะฐะฝะฝะพะน ัะตัะฐะฟะธะธ ะฝะตะธะทะฑะตะถะฝะพ ัะพะฟัะพะฒะพะถะดะฐะตััั ัะตะฟัะพะดัะบัะธะฒะฝัะผะธ ะฟะพัะตััะผะธ, ะฝะตัะตะดะบะพ ะฝะตะพะฑัะฐัะธะผัะผะธ. ะขะฐะบ, R.E. Cabral ะธย ัะพะฐะฒั. (2014) ะฒย ัะบัะฟะตัะธะผะตะฝัะฐะปัะฝะพะน ัะฐะฑะพัะต ะฟะพะบะฐะทะฐะปะธ ะดะพััะฐัะพัะฝะพ ะฒััะพะบะธะน ะฟัะพัะตะบัะธะฒะฝัะน ัััะตะบั L-ะบะฐัะฝะธัะธะฝะฐ ะฒย ะพัะฝะพัะตะฝะธะธ ัะฐััะพ ะฟัะธะผะตะฝัะตะผะพะณะพ ะฒย ะบะปะธะฝะธัะตัะบะพะน ะฟัะฐะบัะธะบะต ะผะพัะฝะพะณะพ ัะธัะพััะฐัะธะบะฐ ะดะพะบัะพััะฑะธัะธะฝะฐ. ะงะตัะตะท 64 ะดะฝั ัะบัะฟะตัะธะผะตะฝัะฐ ัะฐััะพัะฐ ััะฐะณะผะตะฝัะฐัะธะธ ะะะ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ, ะพัะตะฝะตะฝะฝะพะน ะฟะพย TUNEL-ัะตััั, ะพะบะฐะทะฐะปะฐัั ะดะพััะพะฒะตัะฝะพ ะฝะธะถะต ัย ะบััั, ะปะตัะตะฝะฝัั L-ะบะฐัะฝะธัะธะฝะพะผ/ะดะพะบัะพััะฑะธัะธะฝะพะผ, ะฟะพย ััะฐะฒะฝะตะฝะธั ัย ะบัััะฐะผะธ, ะฟะพะปััะฐะฒัะธะผะธ ัะพะปัะบะพ ะดะพะบัะพััะฑะธัะธะฝ [108].
ะะพะฑะฐะฒะปะตะฝะธะต ะบย ะบัะธะพะบะพะฝัะตัะฒะธััััะตะน ััะตะดะต L-ะบะฐัะฝะธัะธะฝะฐ ะฒย ะดะพะทะต 1โ2ย ะณย ะฟัะธะฒะพะดะธะปะพ ะบย ะทะฝะฐัะธัะตะปัะฝะพะผั ัะปัััะตะฝะธั ะฟะฐัะฐะผะตััะพะฒ ัะตะปะพะฒะตัะตัะบะพะณะพ ััะบัะปััะฐ ะฟะพัะปะต ะพััะฐะธะฒะฐะฝะธั ะฟะพย ััะฐะฒะฝะตะฝะธั ัย ะพะฑัะฐะทัะฐะผะธ ะบะพะฝััะพะปัะฝะพะนย ะณััะฟะฟั. ะย ะพัะฝะพะฒะฝะพะนย ะณััะฟะฟะต ะฑัะปะธ ะปัััะต ะฟะพะบะฐะทะฐัะตะปะธ ะพะฑัะตะน ะธย ะฟัะพะณัะตััะธะฒะฝะพะน ะฟะพะดะฒะธะถะฝะพััะธ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ, ะฐย ัะฐะบะถะต ะธั ะถะธะทะฝะตัะฟะพัะพะฑะฝะพััะธ [109โ111]. ะย ะดััะณะพะผ ะธััะปะตะดะพะฒะฐะฝะธะธ ะดะพะฑะฐะฒะปะตะฝะธะต L-ะบะฐัะฝะธัะธะฝะฐ ะดะพ ะฟัะพัะตััะฐ ะบัะธะพยญะบะพะฝัะตัะฒะฐัะธะธ ัััะตััะฒะตะฝะฝะพ ัะฝะธะทะธะปะพ ััะตะฟะตะฝั ะบัะธะพะฟะพะฒัะตะถะดะตะฝะธะน ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะบะฐะบ ะฒย ะฐััะตะฝะพะทะพะพัะฟะตัะผะธัะตัะบะธั , ัะฐะบ ะธย ะฒ ะฝะพัะผะพะทะพะพัะฟะตัะผะธัะตัะบะธั ะพะฑัะฐะทัะฐั ัะฟะตัะผั, ะฟัะธ ััะพะผ ะทะฐัะธัะฝัะน ัััะตะบั ะฑัะป ะฒััะฐะถะตะฝะฝะตะต ะธะผะตะฝะฝะพ ะฟัะธ ะฐััะตะฝะพะทะพะพัะฟะตัะผะธะธ [112].
ะัะปะพ ะฟะพะบะฐะทะฐะฝะพ, ััะพ ะดะพะฟะพะปะฝะธัะตะปัะฝะพะต ะฝะฐะทะฝะฐัะตะฝะธะต ะฟะพะปะธะฒะธัะฐะผะธะฝะฝัั ะฟัะตะฟะฐัะฐัะพะฒ, ัะพะดะตัะถะฐัะธั L-ะบะฐัะฝะธัะธะฝ, ะฒย ัะตัะตะฝะธะต ััะตั ะผะตัััะตะฒ ะฟะพัะปะต ะพะฟะตัะฐัะธะฒะฝะพะณะพ ะปะตัะตะฝะธั ะฒะฐัะธะบะพัะตะปะต ะฟะตัะฒะพะน ััะตะฟะตะฝะธ ะฟะพะทะฒะพะปัะปะพ ัะผะตะฝััะธัั ัะฐััะพัั ััะฐะณะผะตะฝัะฐัะธะธ ะะะ ะฝะฐย 22,1โ34,3% ะธย ัะปัััะธัั ะฟะพะดะฒะธะถะฝะพััั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ ะฒย ััะตะดะฝะตะผ ะฝะฐย 31,3% ะฝะฐย ัะพะฝะต ัะฒะตะปะธัะตะฝะธั ะพะฑัะตะณะพ ะบะพะปะธัะตััะฒะฐ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ [113โ115].
ะะฐะบะพะฒั ะฒะพะทะผะพะถะฝัะต ะผะตั ะฐะฝะธะทะผั ะทะฐัะธัะฝะพะณะพ ัััะตะบัะฐ L-ะบะฐัะฝะธัะธะฝะฐ/ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะฒย ะพัะฝะพัะตะฝะธะธ ะะะ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ? ะะดะฝะธะผ ะธะทย ะฝะธั ะผะพะถะตั ะฑััั ัััะฐะฝะพะฒะปะตะฝะฝะฐั ัะฟะพัะพะฑะฝะพััั L-ะบะฐัะฝะธัะธะฝะฐ/ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ัััะตััะฒะตะฝะฝะพ ัะฝะธะถะฐัั ััะพะฒะตะฝั ะฟัะพะฒะพัะฟะฐะปะธัะตะปัะฝัั ัะธัะพะบะธะฝะพะฒ ะธย ะฟัะพะดัะบัะพะฒ ะฟะตัะตะบะธัะฝะพะณะพ ะพะบะธัะปะตะฝะธั ะปะธะฟะธะดะพะฒ, ะฒย ัะฐััะฝะพััะธ ะผะฐะปะพะฝะดะธะฐะปัะดะตะณะธะดะฐ, ะฒย ัะฟะตัะผะพะฟะปะฐะทะผะต. ะย ัะตะทัะปััะฐัะต ะดะพััะพะฒะตัะฝะพ ัะฝะธะถะฐะตััั ััะตะฟะตะฝั ะฒััะฐะถะตะฝะฝะพััะธ ัะธัะพะบะธะฝะพะฒะพะณะพ ะฒะพัะฟะฐะปะตะฝะธั ะธย ัะฐััะพัะฐ ััะฐะณะผะตะฝัะฐัะธะธ ะะะ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ, ัะฒะตะปะธัะธะฒะฐะตััั ะบะพะฝัะตะฝััะฐัะธั ะฟัะธัะพะดะฝัั ะฐะฝัะธะพะบัะธะดะฐะฝัะพะฒ (ะฒย ัะฐััะฝะพััะธ, ะพะบัะธะดะฐ ะฐะทะพัะฐ NO) [116]. ะััะณะธะผ ะฟะพัะตะฝัะธะฐะปัะฝัะผ ะะะ-ะฟัะพัะตะบัะธะฒะฝัะผ ะผะตั ะฐะฝะธะทะผะพะผ ะผะพะถะตั ะฑััั ัะฟะพัะพะฑะฝะพััั L-ะบะฐัะฝะธัะธะฝะฐ/ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะพะฑะตัะฟะตัะธะฒะฐัั ะฟะตัะฒะธัะฝัั ะฐะฝัะธะพะบัะธะดะฐะฝัะฝัั ะทะฐัะธัั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ, ะฝะธะฒะตะปะธััั ะฝะตะณะฐัะธะฒะฝัะต ัััะตะบัั ะะก ะฒย ะบะปะตัะพัะฝะพะผ ัะฝะดะพยญะฟะปะฐะทะผะฐัะธัะตัะบะพะผ ัะตัะธะบัะปัะผะต ะทะฐ ััะตั ัะฝะธะถะตะฝะธั ะบะพะฝัะตะฝััะฐัะธะธ ัะฒะพะฑะพะดะฝัั ะถะธัะฝัั ะบะธัะปะพั ะธย ะธะทะฑััะพัะฝะพะณะพ ะบะปะตัะพัะฝะพะณะพ ะฐัะตัะธะป-ะบะพัะฝะทะธะผะฐ ะย [117, 118].
ะัะตะฟะฐัะฐัั L-ะบะฐัะฝะธัะธะฝะฐ ะธย ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ ะฒย ัััะธะฝะฝะพะน ัะพััะธะนัะบะพะน ะฟัะฐะบัะธะบะต
ะะพั ัะถะต ะฟะพััะธ 20 ะปะตั ะฒย ะ ะพััะธะธ ัะธัะพะบะพ ะฟัะธะผะตะฝัะตััั ะพัะธะณะธะฝะฐะปัะฝัะน ะบะพะผะฟะปะตะบั, ัะพััะพััะธะน ะธะทย ะบะพะผะฑะธะฝะฐัะธะธ L-ะบะฐัะฝะธัะธะฝะฐ ััะผะฐัะฐัะฐ ะธย ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝะฐ, ะพะฑะพะณะฐัะตะฝะฝะพะน ัััะบัะพะทะพะน ะธย ะปะธะผะพะฝะฝะพะน ะบะธัะปะพัะพะน (ะกะฟะตัะผะฐะบัะธะฝยฎ). ะะพะผะฟะปะตะบั ะพะบะฐะทัะฒะฐะตั ะผะตัะฐะฑะพะปะธัะตัะบะพะต ะดะตะนััะฒะธะต, ััะธะผัะปะธััััะตะต ัะฟะตัะผะฐัะพะณะตะฝะตะท, ะฟะพะฒััะฐะตั ะบะพะฝัะตะฝััะฐัะธั ะธย ะพะฑัะตะต ะบะพะปะธัะตััะฒะพ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ, ัะปัััะฐะตั ะธั ะฟะพะดะฒะธะถะฝะพััั ะธย ะผะพััะพะปะพะณะธั (ัะพัะผั, ัััะพะตะฝะธะต ะธย ะดััะณะธะต ะฟะพะบะฐะทะฐัะตะปะธ).
L-ะบะฐัะฝะธัะธะฝะฐ ััะผะฐัะฐั ะธย ะฐัะตัะธะป-L-ะบะฐัะฝะธัะธะฝ ะพะฑะตัะฟะตัะธะฒะฐัั ะฝะพัะผะฐะปัะฝะพะต ัะพะทัะตะฒะฐะฝะธะต ะธย ัะฒะตะปะธัะธะฒะฐัั ะฟะพะดะฒะธะถะฝะพััั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ. ะัะธ ััะพะผ L-ะบะฐัะฝะธัะธะฝะฐ ััะผะฐัะฐั ะปัััะต, ัะตะผ L-ะบะฐัะฝะธัะธะฝ, ะฟัะพะฝะธะบะฐะตั ัะตัะตะท ะผะตะผะฑัะฐะฝั ะฒย ะผะฐััะธะบั ะผะธัะพั ะพะฝะดัะธะน, ะฑัะดััะธ ัะฒัะทะฐะฝะฝัะผ ัย ะถะธัะฝัะผะธ ะบะธัะปะพัะฐะผะธ. ะะพััะพะผั L-ะบะฐัะฝะธัะธะฝะฐ ััะผะฐัะฐั ะธะฝัะตะฝัะธัะธัะธััะตั ะพะฑะผะตะฝะฝัะต ะฟัะพัะตััั (ะฒะบะปััะฐั ัะฝะตัะณะตัะธัะตัะบะธะต) ะฒย ะผัะถัะบะธั ะฟะพะปะพะฒัั ะบะปะตัะบะฐั , ะพะฑะตัะฟะตัะธะฒะฐะตั ะฟะพะปะฝะพัะตะฝะฝะพะต ัะพะทัะตะฒะฐะฝะธะต ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ, ััะฐะฑะธะปะธะทะธััั ะธั ะบะปะตัะพัะฝัะต ะผะตะผะฑัะฐะฝั, ัะฝะธะถะฐะตั ัะฐััะพัั ััะฐะณะผะตะฝัะฐัะธะธ ะะะ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ.
ะคััะบัะพะทะฐย โ ะพัะฝะพะฒะฝะพะน ะธััะพัะฝะธะบ ัะฝะตัะณะธะธ ะดะปั ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ, ะพะบะฐะทัะฒะฐะตั ะฟะพัะตะฝัะธััััะตะต ะฒะปะธัะฝะธะต ะฝะฐย ัะตััะธะปัะฝัะต (ะพะฟะปะพะดะพัะฒะพััััะธะต) ัะฒะพะนััะฒะฐ ััะบัะปััะฐ. ะะธะผะพะฝะฝะฐั ะบะธัะปะพัะฐ ัะฟะพัะพะฑััะฒัะตั ัะฐะทะถะธะถะตะฝะธั ัะตะผะตะฝะฝะพะน ะถะธะดะบะพััะธ ะธย ะฐะบัะธะฒะฐัะธะธย ะณะธะฐะปััะพะฝะธะดะฐะทัย โ ัะตัะผะตะฝัะฐ, ะพะฑะตัะฟะตัะธะฒะฐััะตะณะพ ะฟัะพะฝะธะบะฝะพะฒะตะฝะธะต ัะฟะตัะผะฐัะพะทะพะธะดะฐ ะฒย ัะนัะตะบะปะตัะบั [119].
ะกะฟะตัะผะฐะบัะธะฝยฎ ะฟัะพัะตะป ัะตะปัะน ััะด ะดะพะบะฐะทะฐัะตะปัะฝัั ะบะปะธะฝะธะบะพ-ะปะฐะฑะพัะฐัะพัะฝัั ะธัะฟััะฐะฝะธะน ะฒย ะ ะพััะธะธ, ะฟะพััะพะผั ัะตะบะพะผะตะฝะดะพะฒะฐะฝ ะะธะณะพะน ัะฟะตัะธะฐะปะธััะพะฒ ะผัะถัะบะพะน ัะตะฟัะพะดัะบัะธะธ ะดะปั ะบะพััะตะบัะธะธ ะฒัะตั ัะพัะผ ะฝะฐัััะตะฝะธะน ัะฟะตัะผะฐัะพะณะตะฝะตะทะฐ ะธย ะฟะพะดะณะพัะพะฒะบะธ ะผัะถัะธะฝ ะบย ะฟัะพัะพะบะพะปั ะญะะ.
ะกะพะฒัะตะผ ะฝะตะดะฐะฒะฝะพ ะฒย ะ ะพััะธะธ ััะฐะปะฐ ะดะพัััะฟะฝะฐ ะฝะพะฒะฐั ัะพัะผะฐ ะบะพะผะฟะปะตะบัะฐย โ ะกะฟะตัะผะฐะบัะธะฝ ะคะพััะตยฎ. ะย ะฝะตะน ััะฐะดะธัะธะพะฝะฝัะน ัะพััะฐะฒ ััะธะปะตะฝ ะตัะต ะพะดะฝะธะผ ะผะพัะฝัะผ ะฐะฝัะธะพะบัะธะดะฐะฝัะพะผ-ะฐะฝัะธะณะธะฟะพะบัะฐะฝัะพะผ ัย ะดะพะบะฐะทะฐะฝะฝัะผ ะผะตัะฐะฑะพะปะธัะตัะบะธะผ ัััะตะบัะพะผ (ะฒย ัะพะผ ัะธัะปะต ะฟัะธ ะผัะถัะบะพะผ ะฑะตัะฟะปะพะดะธะธ)ย โ ะฐะปััะฐ-ะปะธะฟะพะตะฒะพะน ะบะธัะปะพัะพะน (100 ะผะณ ะฝะฐย ัะฐัะต) [120โ123]. ะญัะพ ะฟะพะทะฒะพะปะธะปะพ ะพะฟัะธะผะธะทะธัะพะฒะฐัั ะธย ััะธะปะธัั ะฐะฝัะธะพะบัะธะดะฐะฝัะฝัะต ัะฒะพะนััะฒะฐ ะบะพะผะฟะปะตะบัะฐ, ะบะพัะพััะน, ะบะฐะบ ะธย ะกะฟะตัะผะฐะบัะธะฝยฎ, ะธะผะตะตั ัะปะตะดัััะธะต ะพัะธัะธะฐะปัะฝัะต ะฟะพะบะฐะทะฐะฝะธั ะบย ะฟัะธะผะตะฝะตะฝะธั:
- ะฟะพะดะณะพัะพะฒะบะฐ ะผัะถัะธะฝ ะบย ะทะฐัะฐัะธั;
- ัะฝะธะถะตะฝะธะต ะฟะพะดะฒะธะถะฝะพััะธ ะธย ะพะฟะปะพะดะพัะฒะพััััะตะน ัะฟะพัะพะฑะฝะพััะธ ัะฟะตัะผะฐัะพะทะพะธะดะพะฒ;
- ะพะปะธะณะพะฐััะตะฝะพะทะพะพัะฟะตัะผะธั IIIโIV ััะฐะดะธะธ;
- ะฟะพะดะณะพัะพะฒะบะฐ ะบย ะฟัะพะฒะตะดะตะฝะธั ะฒัะฟะพะผะพะณะฐัะตะปัะฝัั ัะตะฟัะพะดัะบัะธะฒะฝัั ัะตั ะฝะพะปะพะณะธะน (ะญะะ, ะธะฝััะฐัะธัะพยญะฟะปะฐะทะผะฐัะธัะตัะบะฐั ะธะฝัะตะบัะธั ัะฟะตัะผะฐัะพะทะพะธะดะฐ ะฒย ัะนัะตะบะปะตัะบั ะธย ะดั.);
- ัะปัััะตะฝะธะต ะฟะพะบะฐะทะฐัะตะปะตะน ััะบัะปััะฐ ะฟัะธ ะดะพะฝะพัััะฒะต ะธย ะบัะธะพะบะพะฝัะตัะฒะฐัะธะธ;
- ัะฝะธะถะตะฝะธะต ัะตััะธะปัะฝะพััะธ, ะฒัะทะฒะฐะฝะฝะพะต ะะก.
ะะฐะบะปััะตะฝะธะต
ะย ะฝะฐััะพััะตะต ะฒัะตะผั ะฟะพะปััะตะฝั ัะฑะตะดะธัะตะปัะฝัะต ะดะพะบะฐะทะฐัะตะปัััะฒะฐ ัััะตััะฒะตะฝะฝะพะณะพ ะฝะตะณะฐัะธะฒะฝะพะณะพ ะฒะบะปะฐะดะฐ ะะก ะฒย ะฟะฐัะพะณะตะฝะตะท ะฟัะฐะบัะธัะตัะบะธ ะฒัะตั ะทะฐะฑะพะปะตะฒะฐะฝะธะน ัะพะฒัะตะผะตะฝะฝะพะณะพ ัะตะปะพะฒะตะบะฐ, ะฐย ัะฐะบะถะต ัััะฐะฝะพะฒะปะตะฝั ะผะฝะพะณะธะต ะผะพะปะตะบัะปััะฝะพ-ะบะปะตัะพัะฝัะต ะผะตั ะฐะฝะธะทะผั ะะก. ยซะัะปะธ ัะฝะตัะณะตัะธัะตัะบะธะน ะฑะฐะปะฐะฝั ะพัะณะฐะฝะธะทะผะฐ ะฝะธะถะต ััะตะดะฝะตะณะพ, ะพัะณะฐะฝะธะทะผ ะฝะตย ะผะพะถะตั ัะพะฟัะพัะธะฒะปััััั ะฑะพะปะตะทะฝะตะฝะฝัะผ ะฐะณัะตััะธัะผ ะธย ะฑะตะทะฝะฐะดะตะถะฝะพ ะทะฐะฑะพะปะตะตั. ะะตะดะธัะธะฝะฐ ะบะฐะปะตัะฐัะฐั ะดะพะปะถะฝะฐ ััััะฟะธัั ะผะตััะพ ะผะตะดะธัะธะฝะต, ััะฐัะฐััะตะนัั ะฟะพะฒััะธัั ัะฝะตัะณะตัะธัะตัะบะธะน ะฑะฐะปะฐะฝัยป,ย โ ัะฐะบ ะฟัะพัะพัะตัะบะธ ะฟะธัะฐะป ะตัะต ะฒย 1963ย ะณ.ย ะฒย ะบะฝะธะณะต ยซะขะฐะนะฝะฐั ะผัะดัะพััั ัะตะปะพะฒะตัะตัะบะพะณะพ ะพัะณะฐะฝะธะทะผะฐยป ะฒะตะปะธะบะธะน ััััะบะธะน ะฒัะฐั ะธย ัะธะปะพัะพั ะ.ะก. ะะฐะปะผะฐะฝะพะฒ.
ะกะตะณะพะดะฝั, ะฟะพะถะฐะปัะน, ะฝะธะบะพะผั ะฝะตย ะฝะฐะดะพ ะดะพะบะฐะทัะฒะฐัั, ััะพ ะะกย โ ััะพ ัะฝะธะฒะตััะฐะปัะฝัะน ะผะตั ะฐะฝะธะทะผ ะฟะฐัะพะณะตะฝะตะทะฐ ะฒัะตั ะฒะพะทัะฐัั-ะฐััะพัะธะธัะพะฒะฐะฝะฝัั ะทะฐะฑะพะปะตะฒะฐะฝะธะน, ะฟัะธ ััะพะผ ะฒััะฐะถะตะฝะฝะพััั ะะก ััะธะปะธะฒะฐะตััั ัย ะฒะพะทัะฐััะพะผ. ะกะฐะผัะผะธ ะฟะตัะฒัะผะธ ะฟะฐัะพัะธะทะธะพะปะพะณะธัะตัะบะธะผะธ ะผะธัะตะฝัะผะธ ะปัะฑะพะณะพ ะะก ััะฐะฝะพะฒัััั ะฒััะพะบะพัะฟะตัะธะฐะปะธะทะธัะพะฒะฐะฝะฝัะต ะธย ะฟะพัะพะผั ะฝะฐะธะฑะพะปะตะต ัะฝะตัะณะพะฟะพััะตะฑะปัััะธะต ะบะปะตัะบะธ ะพัะณะฐะฝะธะทะผะฐ, ะบย ะบะพัะพััะผ ะพัะฝะพััััั ะฒย ัะพะผ ัะธัะปะต ะฟะพะปะพะฒัะต ะบะปะตัะบะธ (ัะฟะตัะผะฐัะพะทะพะธะดั ัย ะผัะถัะธะฝ ะธย ัะนัะตะบะปะตัะบะธ ัย ะถะตะฝัะธะฝ). ะะพั ะฟะพัะตะผั ะะก ัะตะณะพะดะฝัย โ ะฝะตะธะทะฑะตะถะฝัะน ะฐััะธะฑัั ะฑะตัะฟะปะพะดะธั ะธย ะตะณะพ ัะฝะธะฒะตััะฐะปัะฝัะน ะฟะฐัะพะณะตะฝะตัะธัะตัะบะธะน ะผะตั ะฐะฝะธะทะผ ะฝะตะทะฐะฒะธัะธะผะพ ะพัย ะฟะตัะฒะพะฟัะธัะธะฝั ะฝะฐัััะตะฝะธั ัะตััะธะปัะฝะพััะธ. ะะก ะฐะฟัะธะพัะธ ัััะตััะฒัะตั ะฒย ะพัะณะฐะฝะธะทะผะต ะผัะถัะธะฝั (ัะธะทะธะพะปะพะณะธัะตัะบะธะน ะะก), ะพะดะฝะฐะบะพ ะฟัะธ ะฑะตัะฟะปะพะดะธะธ ะะก ะฟัะธะพะฑัะตัะฐะตั ะฟะฐัะพะปะพะณะธัะตัะบะธะน ั ะฐัะฐะบัะตั. ะะฐัะพะปะพะณะธัะตัะบะธะน ะะก ัััะตััะฒะตะฝะฝะพ ัั ัะดัะฐะตั ัะตะทัะปััะฐัั ะปัะฑะพะน ะฟัะพัะธะปัะฝะพะน ัะตัะฐะฟะธะธ ะผัะถัะบะพะณะพ ะฑะตัะฟะปะพะดะธั, ะตัะปะธ ะฟะฐัะธะตะฝัั ะพะดะฝะพะฒัะตะผะตะฝะฝะพ ะฝะตย ะฝะฐะทะฝะฐัะฐะตััั ะผะตัะฐะฑะพะปะธัะตัะบะฐั ะฐะฝัะธะพะบัะธะดะฐะฝัะฝะฐั ัะตัะฐะฟะธั, ัะฟะพัะพะฑะฝะฐั ัััะตะบัะธะฒะฝะพ ะฝะธะฒะตะปะธัะพะฒะฐัั ัะธััะตะผะฝัะต ะธย ะปะพะบะฐะปัะฝัะต (ัะตััะธะบัะปััะฝัะต) ะผะตั ะฐะฝะธะทะผั ะะก.
ะย ะฝะฐััะพััะตะต ะฒัะตะผั ะฝะฐย ัะฐัะผะฐัะตะฒัะธัะตัะบะพะผ ััะฝะบะต ะ ะพััะธะธ ะธะผะตะตััั ะตะดะธะฝััะฒะตะฝะฝัะน ะบะพะผะฟะปะตะบั ัย ะดะพะบะฐะทะฐะฝะฝัะผ ะฐะฝัะธะพะบัะธะดะฐะฝัะฝัะผ ะดะตะนััะฒะธะตะผย โ ะกะฟะตัะผะฐะบัะธะฝ ะคะพััะตยฎ. ะะณะพ ััะธะปะตะฝะฝะฐั ัะพัะผัะปะฐ ะฟะพะทะฒะพะปัะตั ะพัััะตััะฒะปััั ัััะตะบัะธะฒะฝะพะต ัะฐัะผะฐะบะพะปะพะณะธัะตัะบะพะต ัะพะฟัะพะฒะพะถะดะตะฝะธะต ะปัะฑัั ะฒะฐัะธะฐะฝัะพะฒ ะปะตัะตะฝะธั ะผัะถัะบะพะณะพ ะฑะตัะฟะปะพะดะธั ะฝะตะทะฐะฒะธัะธะผะพ ะพัย ะธั ะฟัะธัะธะฝั. ะกะฟะตัะผะฐะบัะธะฝ ะคะพััะตยฎ ะบะฐะบ ะฑะฐะทะพะฒัะน ะผะตัะฐะฑะพะปะธัะตัะบะธะน ะฐะฝัะธะพะบัะธะดะฐะฝัะฝัะน ะบะพะผะฟะพะฝะตะฝั ะบะพะผะฟะปะตะบัะฝะพะน ัะตัะฐะฟะธะธ ะปัะฑัั ัะพัะผ ะผัะถัะบะพะณะพ ะฑะตัะฟะปะพะดะธั ะพะฑะปะฐะดะฐะตั ะฒััะพะบะพะน ัััะตะบัะธะฒะฝะพัััั ะธย ะฑะตะทะพะฟะฐัะฝะพัััั ะธย ะฟะพะทะฒะพะปัะตั ัะตะฐะปะธะทะพะฒะฐััย ะณะปะฐะฒะฝัะน ะฟัะธะฝัะธะฟ ะผะตะดะธัะธะฝั ยซะปะตัะธัั ะฑะพะปัะฝะพะณะพ, ะฐย ะฝะต ะฑะพะปะตะทะฝัยป. ะัะตะฒะธะดะฝะพ, ััะพ ะฑะพะปะตะต ัะธัะพะบะพะต ะฟัะธะผะตะฝะตะฝะธะต ะกะฟะตัะผะฐะบัะธะฝะฐ ะคะพััะตยฎ ะฒย ัััะธะฝะฝะพะน ะบะปะธะฝะธัะตัะบะพะน ะฟัะฐะบัะธะบะต ะฟะพะทะฒะพะปะธั ะดะตะนััะฒะธัะตะปัะฝะพ ะพัััะตััะฒะธัั ะฟัะพััะฒ ะฒย ะบะพะฝัะตัะฒะฐัะธะฒะฝะพะน ัะตัะฐะฟะธะธ ะปัะฑัั ัะพัะผ ะผัะถัะบะพะน ัะตะฟัะพะดัะบัะธะธ, ะฒย ัะพะผ ัะธัะปะต ะธะดะธะพะฟะฐัะธัะตัะบะพะณะพ ะผัะถัะบะพะณะพ ะฑะตัะฟะปะพะดะธั, ะฒััะพะบะฐั ัะฐััะพัะฐ ะบะพัะพัะพะณะพ ะฒย ะ ะพััะธะธ, ะดะพััะธะณะฐััะฐั 60โ70%, ะฝะตย ะผะพะถะตั ะฝะตย ะฒัะทัะฒะฐัั ะพะฟะฐัะตะฝะธั.
S.Yu. Kalinchenko, I.A. Tyuzikov
Clinic of Professor Kalinchenko, Moscow
Peoplesโ Friendship University of Russia, Moscow
Contact person: Igor Adamovich Tyuzikov, phoenix-67@list.ru
At present, an impact of male factor into familial infertility reaches up to 50%. However, in most cases the results of conservatively treated male infertility still remain unsatisfactory. Frequently, certain nosologies that might be caused by impaired male fertility are detected and corrected, but general universal mechanisms resulting in infertility are not taken into account. Oxidative stress in spermatozoa represents a direct pathogenetic mechanism of impaired spermatogenesis underlying virtually all types of male infertility. It has various manifestations generally being exhibited in developing mitochondrial dysfunction and sperm DNA fragmentation negatively affecting both macro- and microscopic features of gametes: from motility up to fertilizing capacity seemingly 'healthy' spermatozoa. By now, a great body of evidence database confirming efficacy of using antioxidant drugs in a combination therapy of male infertility has been accumulated in research literature and clinical practice. Antioxidant therapy increases rate of natural frequency of natural as well as IVF conceptions. L-carnitine/acetyl-L-carnitine complex is one of few antioxidants with verified positive effect on oxidative stress in spermatozoa. It allows recommending it as a basic metabolic antioxidant therapy for all types of male infertility regardless of underlying cause and correcting methods.
ะฃะฒะฐะถะฐะตะผัะน ะฟะพัะตัะธัะตะปั uMEDp!
ะฃะฒะตะดะพะผะปัะตะผ ะะฐั ะพ ัะพะผ, ััะพ ะทะดะตัั ัะพะดะตัะถะธััั ะธะฝัะพัะผะฐัะธั, ะฟัะตะดะฝะฐะทะฝะฐัะตะฝะฝะฐั ะธัะบะปััะธัะตะปัะฝะพ ะดะปั ัะฟะตัะธะฐะปะธััะพะฒ ะทะดัะฐะฒะพะพั ัะฐะฝะตะฝะธั.
ะัะปะธ ะั ะฝะต ัะฒะปัะตัะตัั ัะฟะตัะธะฐะปะธััะพะผ ะทะดัะฐะฒะพะพั ัะฐะฝะตะฝะธั, ะฐะดะผะธะฝะธัััะฐัะธั ะฝะต ะฝะตัะตั ะพัะฒะตัััะฒะตะฝะฝะพััะธ ะทะฐ ะฒะพะทะผะพะถะฝัะต ะพััะธัะฐัะตะปัะฝัะต ะฟะพัะปะตะดััะฒะธั, ะฒะพะทะฝะธะบัะธะต ะฒ ัะตะทัะปััะฐัะต ัะฐะผะพััะพััะตะปัะฝะพะณะพ ะธัะฟะพะปัะทะพะฒะฐะฝะธั ะะฐะผะธ ะธะฝัะพัะผะฐัะธะธ ั ะฟะพััะฐะปะฐ ะฑะตะท ะฟัะตะดะฒะฐัะธัะตะปัะฝะพะน ะบะพะฝััะปััะฐัะธะธ ั ะฒัะฐัะพะผ.
ะะฐะถะธะผะฐั ะฝะฐ ะบะฝะพะฟะบั ยซะะพะนัะธยป, ะั ะฟะพะดัะฒะตัะถะดะฐะตัะต, ััะพ ัะฒะปัะตัะตัั ะฒัะฐัะพะผ ะธะปะธ ัััะดะตะฝัะพะผ ะผะตะดะธัะธะฝัะบะพะณะพ ะฒัะทะฐ.