–Я—А–Њ–≥–љ–Њ—Б—В–Є—З–µ—Б–Ї–Њ–µ –Є¬†–њ—А–µ–і–Є–Ї—В–Є–≤–љ–Њ–µ –Ј–љ–∞—З–µ–љ–Є–µ –Ґ-–Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є –≤¬†–њ–µ—А–Є—Д–µ—А–Є—З–µ—Б–Ї–Њ–є –Ї—А–Њ–≤–Є —Г¬†–±–Њ–ї—М–љ—Л—Е –љ–µ–Њ–њ–µ—А–∞–±–µ–ї—М–љ–Њ–є –Є–ї–Є –Љ–µ—В–∞—Б—В–∞—В–Є—З–µ—Б–Ї–Њ–є –Љ–µ–ї–∞–љ–Њ–Љ–Њ–є
- –Р–љ–љ–Њ—В–∞—Ж–Є—П
- –°—В–∞—В—М—П
- –°—Б—Л–ї–Ї–Є
- English
–Т–≤–µ–і–µ–љ–Є–µ
–Ш–Љ–Љ—Г–љ–Њ—В–µ—А–∞–њ–Є—П —П–≤–ї—П–µ—В—Б—П –Њ–і–љ–Є–Љ –Є–Ј –Ї–ї—О—З–µ–≤—Л—Е —Н–ї–µ–Љ–µ–љ—В–Њ–≤ –ї–µ—З–µ–љ–Є—П –Љ–µ–ї–∞–љ–Њ–Љ—Л. –Ч–љ–∞—З–Є—В–µ–ї—М–љ—Л–µ —Г—Б–њ–µ—Е–Є –≤¬†—Н—В–Њ–Љ –љ–∞–њ—А–∞–≤–ї–µ–љ–Є–Є –±—Л–ї–Є –і–Њ—Б—В–Є–≥–љ—Г—В—Л –±–ї–∞–≥–Њ–і–∞—А—П —А–∞–Ј—А–∞–±–Њ—В–Ї–µ –Є–љ–≥–Є–±–Є—В–Њ—А–Њ–≤ –Є–Љ–Љ—Г–љ–љ–Њ–≥–Њ —Б–Є–љ–∞–њ—Б–∞¬†вАУ –њ—А–µ–њ–∞—А–∞—В–Њ–≤, –±–ї–Њ–Ї–Є—А—Г—О—Й–Є—Е —В–Њ—А–Љ–Њ–Ј–љ–Њ–µ –≤–ї–Є—П–љ–Є–µ –љ–∞ –Є–Љ–Љ—Г–љ–љ—Г—О —Б–Є—Б—В–µ–Љ—Г, –њ—А–µ–њ—П—В—Б—В–≤—Г—О—Й–µ–µ –∞–Ї—В–Є–≤–∞—Ж–Є–Є –Ґ-–Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є –Є¬†—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—О –Є–Ј –љ–Є—Е —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л—Е —Б—Г–±–њ–Њ–њ—Г–ї—П—Ж–Є–є [1]. –Ф–∞–љ–љ—Л–µ –Љ–µ—В–Њ–і—Л –Њ–Ї–∞–Ј–∞–ї–Є—Б—М –≤–µ—Б—М–Љ–∞ —Н—Д—Д–µ–Ї—В–Є–≤–љ—Л. –Ю–і–љ–∞–Ї–Њ —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–∞—П –і–Њ–ї—П –њ–∞—Ж–Є–µ–љ—В–Њ–≤ –љ–µ –Њ—В–≤–µ—З–∞–µ—В –љ–∞ –њ–Њ–і–Њ–±–љ–Њ–µ –ї–µ—З–µ–љ–Є–µ, –Є¬†–ї–Є—И—М –≤¬†–љ–µ–±–Њ–ї—М—И–Њ–Љ –њ—А–Њ—Ж–µ–љ—В–µ —Б–ї—Г—З–∞–µ–≤ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ –Є–Ј–ї–µ—З–µ–љ–Є–µ. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –Њ—З–µ–≤–Є–і–љ–∞ –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ—Б—В—М –њ–Њ–Є—Б–Ї–∞ –±–Є–Њ–Љ–∞—А–Ї–µ—А–Њ–≤ –і–ї—П –њ–µ—А—Б–Њ–љ–∞–ї–Є–Ј–∞—Ж–Є–Є –ї–µ—З–µ–љ–Є—П –±–Њ–ї—М–љ—Л—Е.
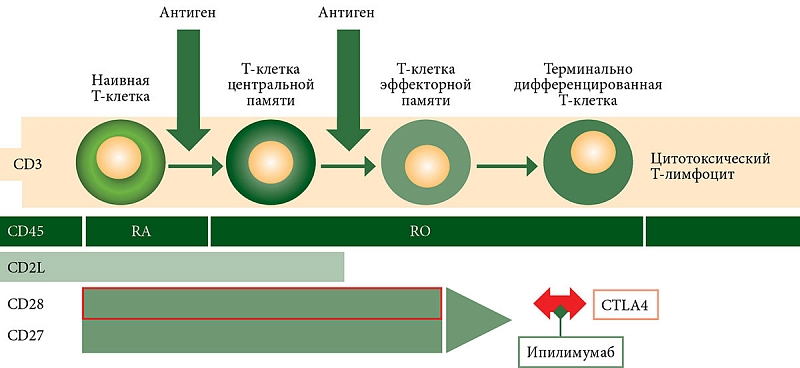
–Ґ-–Ї–ї–µ—В–Ї–Є –њ–∞–Љ—П—В–Є –њ—А–µ–і—Б—В–∞–≤–ї—П—О—В —Б–Њ–±–Њ–є –≥–µ—В–µ—А–Њ–≥–µ–љ–љ—Г—О –њ–Њ–њ—Г–ї—П—Ж–Є—О –Ї–ї–µ—В–Њ–Ї, –Ї–Њ—В–Њ—А—Л–µ –≤¬†—Б–≤–Њ–µ–Љ —А–∞–Ј–≤–Є—В–Є–Є –Њ—В –љ–∞–Є–≤–љ—Л—Е –і–Њ —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л—Е –њ—А–µ—В–µ—А–њ–µ–≤–∞—О—В —А—П–і –Є–Ј–Љ–µ–љ–µ–љ–Є–є [2]. –Э–∞ —Б–µ–≥–Њ–і–љ—П—И–љ–Є–є –і–µ–љ—М —Б—Г—Й–µ—Б—В–≤—Г–µ—В –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –Ї–Њ–љ—Ж–µ–њ—Ж–Є–є –Є¬†–њ—А–µ–і–њ–Њ–ї–Њ–ґ–µ–љ–Є–є —А–∞–Ј–≤–Є—В–Є—П —Н—В–Є—Е –Ї–ї–µ—В–Њ–Ї, –љ–Њ –≤¬†—Ж–µ–ї—П—Е –і–∞–љ–љ–Њ–≥–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –Њ—Б—В–∞–љ–Њ–≤–Є–Љ—Б—П —В–Њ–ї—М–Ї–Њ –љ–∞ –Љ–Њ–і–µ–ї–Є —Б–љ–Є–ґ–∞—О—Й–µ–≥–Њ—Б—П –њ–Њ—В–µ–љ—Ж–Є–∞–ї–∞ (—А–Є—Б. 1).
–Т–∞–ґ–љ–Њ –њ–Њ–і—З–µ—А–Ї–љ—Г—В—М, —З—В–Њ –Є–Љ–µ–љ–љ–Њ –љ–∞ T-–Ї–ї–µ—В–Ї–∞—Е –њ–∞–Љ—П—В–Є —Н–Ї—Б–њ—А–µ—Б—Б–Є—А—Г–µ—В—Б—П —А–µ—Ж–µ–њ—В–Њ—А CD28, –Ї–Њ—В–Њ—А—Л–є –Ї–Њ–љ–Ї—Г—А–Є—А—Г–µ—В —Б¬†—А–µ—Ж–µ–њ—В–Њ—А–Њ–Љ CTLA4 –Ј–∞ —Б–≤—П–Ј—Л–≤–∞–љ–Є–µ —Б¬†CD80 [3]. –Ф–∞–љ–љ–∞—П —Б–≤—П–Ј—М —Б–ї—Г–ґ–Є—В —Б–Є–≥–љ–∞–ї–Њ–Љ –≤—В–Њ—А–Њ–≥–Њ —В–Є–њ–∞ –≤¬†–Є–Љ–Љ—Г–љ–љ–Њ–Љ —Б–Є–љ–∞–њ—Б–µ –њ—А–Є –≤—Б—В—А–µ—З–µ –љ–∞–Є–≤–љ–Њ–є –Ґ-–Ї–ї–µ—В–Ї–Є –Є–ї–Є –Ґ-–Ї–ї–µ—В–Ї–Є —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –њ–∞–Љ—П—В–Є, —Б–њ–Њ—Б–Њ–±–љ–Њ–є –Ї¬†–і–ї–Є—В–µ–ї—М–љ–Њ–є –њ–µ—А—Б–Є—Б—В–µ–љ—Ж–Є–Є –Є¬†–њ—А–Њ–ї–Є—Д–µ—А–∞—Ж–Є–Є, —Б¬†–∞–љ—В–Є–≥–µ–љ–Њ–Љ –Є¬†–Њ–±–µ—Б–њ–µ—З–Є–≤–∞–µ—В –њ—А–Њ–ї–Є—Д–µ—А–∞—В–Є–≤–љ—Л–є –Є¬†–∞–љ—В–Є–∞–њ–Њ–њ—В–Њ—В–Є—З–µ—Б–Ї–Є–є —Б–Є–≥–љ–∞–ї [2]. –Э–∞–њ—А–Њ—В–Є–≤, –∞–Ї—В–Є–≤–∞—Ж–Є—П CTLA4, –Є–Љ–µ—О—Й–µ–≥–Њ –±–Њ–ї—М—И—Г—О –∞—Д—Д–Є–љ–љ–Њ—Б—В—М –Ї¬†CD80, –Є–љ–і—Г—Ж–Є—А—Г–µ—В –∞–њ–Њ–њ—В–Њ–Ј –Ї–ї–µ—В–Њ–Ї –Є¬†–љ–µ –њ—А–Є–≤–Њ–і–Є—В –Ї¬†–Є—Е –∞–Ї—В–Є–≤–∞—Ж–Є–Є. –Ь—Л –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є–ї–Є, —З—В–Њ —Г—А–Њ–≤–µ–љ—М –Ґ-–Ї–ї–µ—В–Њ–Ї –≤¬†–њ–µ—А–Є—Д–µ—А–Є—З–µ—Б–Ї–Њ–є –Ї—А–Њ–≤–Є –Љ–Њ–ґ–µ—В –±—Л—В—М –Љ–∞—А–Ї–µ—А–Њ–Љ —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є —В–µ—А–∞–њ–Є–Є –Є¬†–≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В–Є –±–Њ–ї—М–љ—Л—Е –Љ–µ–ї–∞–љ–Њ–Љ–Њ–є. –Ф–ї—П –њ–Њ–і—В–≤–µ—А–ґ–і–µ–љ–Є—П –≤—Л–і–≤–Є–љ—Г—В–Њ–є –љ–∞–Љ–Є –≥–Є–њ–Њ—В–µ–Ј—Л –±—Л–ї–Њ –њ—А–Њ–≤–µ–і–µ–љ–Њ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ.
–Ь–∞—В–µ—А–Є–∞–ї –Є¬†–Љ–µ—В–Њ–і—Л
–Т –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –≤–Ї–ї—О—З–µ–љ—Л –±–Њ–ї—М–љ—Л–µ –љ–µ–Њ–њ–µ—А–∞–±–µ–ї—М–љ–Њ–є –Є–ї–Є –Љ–µ—В–∞—Б—В–∞—В–Є—З–µ—Б–Ї–Њ–є –Љ–µ–ї–∞–љ–Њ–Љ–Њ–є, –њ–Њ–ї—Г—З–∞–≤—И–Є–µ –Є–Љ–Љ—Г–љ–Њ—В–µ—А–∞–њ–Є—О –Є–њ–Є–ї–Є–Љ—Г–Љ–∞–±–Њ–Љ —Б¬†2012 –њ–Њ 2015 –≥. –Ю—Б–љ–Њ–≤–љ—Л–µ –Ї—А–Є—В–µ—А–Є–Є –≤–Ї–ї—О—З–µ–љ–Є—П:
- –њ–Њ–і–њ–Є—Б–∞–љ–Є–µ –Є–љ—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–љ–Њ–≥–Њ —Б–Њ–≥–ї–∞—Б–Є—П;
- –≤–Њ–Ј—А–∞—Б—В —Б—В–∞—А—И–µ 18 –ї–µ—В;
- –љ–∞–ї–Є—З–Є–µ –Љ–Њ—А—Д–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є –њ–Њ–і—В–≤–µ—А–ґ–і–µ–љ–љ–Њ–є –љ–µ–Њ–њ–µ—А–∞–±–µ–ї—М–љ–Њ–є –Є–ї–Є –Љ–µ—В–∞—Б—В–∞—В–Є—З–µ—Б–Ї–Њ–є –Љ–µ–ї–∞–љ–Њ–Љ—Л;
- –Њ—В—Б—Г—В—Б—В–≤–Є–µ –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є –Ј–љ–∞—З–Є–Љ—Л—Е –љ–∞—А—Г—И–µ–љ–Є–є —Д—Г–љ–Ї—Ж–Є–Є –≤–љ—Г—В—А–µ–љ–љ–Є—Е –Њ—А–≥–∞–љ–Њ–≤;
- –Њ—В—Б—Г—В—Б—В–≤–Є–µ –∞—Г—В–Њ–Є–Љ–Љ—Г–љ–љ—Л—Е –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є–є.
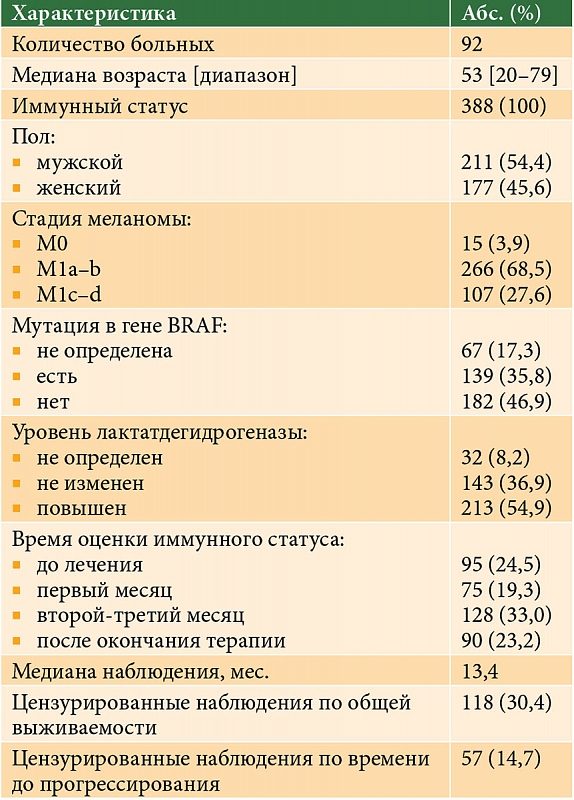
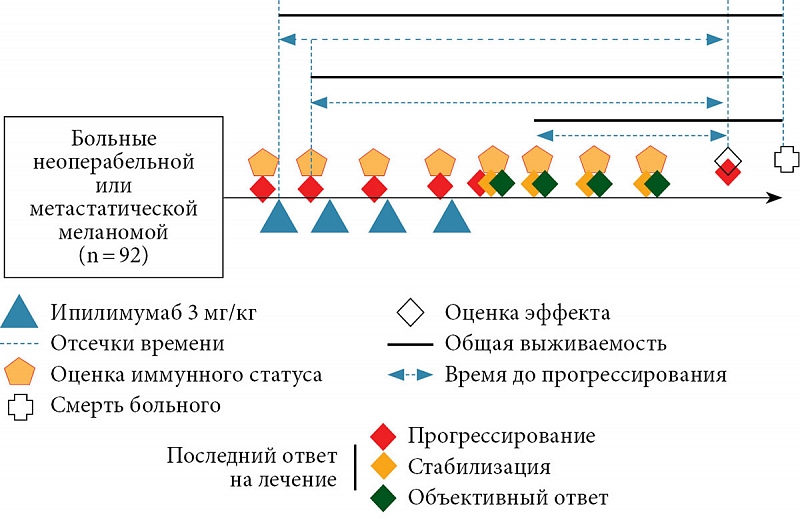
–•–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–∞ –њ–∞—Ж–Є–µ–љ—В–Њ–≤ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–∞ –≤¬†—В–∞–±–ї. 1, —Б—Е–µ–Љ–∞ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П¬†вАУ –љ–∞ —А–Є—Б. 2. –Ф–ї—П –Њ—Ж–µ–љ–Ї–Є –Є–Љ–Љ—Г–љ–љ–Њ–≥–Њ —Б—В–∞—В—Г—Б–∞ (–Ш–°) –Є–Ј–Љ–µ—А—П–ї–Є —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ —Ж–Є—В–Њ—В–Њ–Ї—Б–Є—З–µ—Б–Ї–Є—Е –ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ (CTL, CD3+CD8+), –љ–∞–Є–≤–љ—Л—Е –Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ (Tn, CD3+CD45RA+CD62L+), —Ж–µ–љ—В—А–∞–ї—М–љ—Л—Е –Ґ-–Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є (Tcm, CD3+CD45R0+CD62L+), –Ї–ї–µ—В–Њ–Ї —Н—Д—Д–µ–Ї—В–Њ—А–љ–Њ–є –њ–∞–Љ—П—В–Є (Tem, CD3+CD45R0+CD62L-).
–Ф–ї—П –Њ—Ж–µ–љ–Ї–Є —Б–≤—П–Ј–Є –њ–Њ–Ї–∞–Ј–∞—В–µ–ї—П —Б¬†—Н—Д—Д–µ–Ї—В–Њ–Љ –ї–µ—З–µ–љ–Є—П —Б—А–∞–≤–љ–Є–≤–∞–ї–Є —З–∞—Б—В–Њ—В—Г –Њ—В–і–µ–ї—М–љ—Л—Е –≤–Є–і–Њ–≤ –Њ—В–≤–µ—В–∞ –њ–Њ —Б–Є—Б—В–µ–Љ–µ RECIST (–њ—А–Њ–≥—А–µ—Б—Б–Є—А–Њ–≤–∞–љ–Є–µ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П, —Б—В–∞–±–Є–ї–Є–Ј–∞—Ж–Є—П –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П, –Њ–±—К–µ–Ї—В–Є–≤–љ—Л–є –Њ—В–≤–µ—В) —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–љ–∞–ї–Є—З–Є–µ–Љ –Є–ї–Є –Њ—В—Б—Г—В—Б—В–≤–Є–µ–Љ –Њ—В–Ї–ї–Њ–љ–µ–љ–Є–є —Г—А–Њ–≤–љ—П –∞–љ–∞–ї–Є–Ј–Є—А—Г–µ–Љ—Л—Е –њ–Њ–њ—Г–ї—П—Ж–Є–є –Ї–ї–µ—В–Њ–Ї –Њ—В —А–µ—Д–µ—А–µ–љ—Б–љ—Л—Е –Ј–љ–∞—З–µ–љ–Є–є (–њ—А–Є –Є—Е –љ–∞–ї–Є—З–Є–Є). –Ю—В–≤–µ—В –Њ–њ—А–µ–і–µ–ї—П–ї–Є –Ї–∞–Ї –њ–Њ—Б–ї–µ–і–љ–Є–є —А–µ–Ј—Г–ї—М—В–∞—В –Њ—Ж–µ–љ–Ї–Є —Б–Њ—Б—В–Њ—П–љ–Є—П –Њ–њ—Г—Е–Њ–ї–Є –њ–µ—А–µ–і –Њ—Ж–µ–љ–Ї–Њ–є –Ш–° (–≥—А—Г–±–∞—П –Њ—Ж–µ–љ–Ї–∞) –Є¬†–≤ –њ—А–µ–і–µ–ї–∞—Е –і–≤—Г—Е –љ–µ–і–µ–ї—М –Њ—В –Њ—Ж–µ–љ–Ї–Є –Ш–° (—В–Њ—З–љ–∞—П –Њ—Ж–µ–љ–Ї–∞). –Ф–ї—П —Б—А–∞–≤–љ–µ–љ–Є—П —А–∞—Б–њ—А–µ–і–µ–ї–µ–љ–Є–є –Њ—В–≤–µ—В–Њ–≤ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–Є –Љ–µ—В–Њ–і —Е–Є-–Ї–≤–∞–і—А–∞—В –Є¬†—Б—В–∞—В–Є—Б—В–Є–Ї—Г –Њ—В–љ–Њ—И–µ–љ–Є—П –њ—А–∞–≤–і–Њ–њ–Њ–і–Њ–±–Є—П. –Ф–ї—П –Њ—Ж–µ–љ–Ї–Є –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В–Є –±—Л–ї–Є –њ–Њ—Б—В—А–Њ–µ–љ—Л –њ—А–Њ–≥–љ–Њ—Б—В–Є—З–µ—Б–Ї–Є–µ –Љ–Њ–і–µ–ї–Є, –≤–Ї–ї—О—З–∞–≤—И–Є–µ –і–µ–Љ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Є–µ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є (–њ–Њ–ї, –≤–Њ–Ј—А–∞—Б—В), —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є –≤—А–µ–Љ–µ–љ–Є –ї–µ—З–µ–љ–Є—П (–і–Њ –љ–∞—З–∞–ї–∞ —В–µ—А–∞–њ–Є–Є, –љ–∞—З–∞–ї–Њ –ї–µ—З–µ–љ–Є—П, –њ—А–Њ–і–Њ–ї–ґ–µ–љ–Є–µ –ї–µ—З–µ–љ–Є—П, –њ–Њ—Б–ї–µ –ї–µ—З–µ–љ–Є—П, –≤—А–µ–Љ—П –љ–∞ —В–µ—А–∞–њ–Є–Є –≤¬†–Љ–µ—Б—П—Ж–∞—Е), —Б—В–∞–і–Є—О (III –њ—А–Њ—В–Є–≤ IV) –Є¬†–њ–Њ–і—Б—В–∞–і–Є—О (M0 –Є¬†M1aвАУb –њ—А–Њ—В–Є–≤ M1c) –њ—А–Њ—Ж–µ—Б—Б–∞ –љ–∞ –Љ–Њ–Љ–µ–љ—В –љ–∞—З–∞–ї–∞ —В–µ—А–∞–њ–Є–Є, –љ–∞–ї–Є—З–Є–µ –њ–Њ–≤—Л—И–µ–љ–љ–Њ–≥–Њ —Г—А–Њ–≤–љ—П –ї–∞–Ї—В–∞—В–і–µ–≥–Є–і—А–Њ–≥–µ–љ–∞–Ј—Л –Є¬†–Љ—Г—В–∞—Ж–Є–є –≤¬†–≥–µ–љ–µ BRAF, –Љ–µ–ї–∞–љ–Њ–Љ—Л —Б–ї–Є–Ј–Є—Б—В–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є –Є–ї–Є —Г–≤–µ–∞–ї—М–љ–Њ–є –Љ–µ–ї–∞–љ–Њ–Љ—Л, –њ–µ—А–≤–Є—З–љ–Њ-–Љ–љ–Њ–ґ–µ—Б—В–≤–µ–љ–љ—Л—Е –Њ–њ—Г—Е–Њ–ї–µ–є, –Є–Љ–Љ—Г–љ–Њ–Њ–њ–Њ—Б—А–µ–і–Њ–≤–∞–љ–љ—Л—Е –љ–µ–ґ–µ–ї–∞—В–µ–ї—М–љ—Л—Е —П–≤–ї–µ–љ–Є–є, –ї–Є–љ–Є—О —В–µ—А–∞–њ–Є–Є, –≤–Є–і –њ—А–Њ–≤–µ–і–µ–љ–љ–Њ–≥–Њ —А–∞–љ–µ–µ –ї–µ—З–µ–љ–Є—П. –Т—А–µ–Љ—П –і–Њ –њ—А–Њ–≥—А–µ—Б—Б–Є—А–Њ–≤–∞–љ–Є—П (–Т–Ф–Я) –Є–Ј–Љ–µ—А—П–ї–Њ—Б—М –Њ—В –≤—А–µ–Љ–µ–љ–Є –Њ—Ж–µ–љ–Ї–Є –Ш–° (–і–ї—П –Ш–°, –Є—Б—Б–ї–µ–і—Г–µ–Љ—Л—Е –і–Њ –љ–∞—З–∞–ї–∞ —В–µ—А–∞–њ–Є–Є,¬†вАУ –Њ—В –≤—А–µ–Љ–µ–љ–Є –љ–∞—З–∞–ї–∞ –ї–µ—З–µ–љ–Є—П) –і–Њ –≤—Л—П–≤–ї–µ–љ–Є—П –њ–µ—А–≤—Л—Е –њ—А–Є–Ј–љ–∞–Ї–Њ–≤ –њ—А–Њ–≥—А–µ—Б—Б–Є—А–Њ–≤–∞–љ–Є—П –њ—А–Њ—Ж–µ—Б—Б–∞. –Ю–±—Й–∞—П –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В—М (–Ю–Т) —А–∞—Б—Б—З–Є—В—Л–≤–∞–ї–∞—Б—М –Њ—В —В–Њ–є –ґ–µ —В–Њ—З–Ї–Є, —З—В–Њ –Є¬†–Т–Ф–Я, –і–Њ —Б–Љ–µ—А—В–Є –њ–Њ –ї—О–±–Њ–є –њ—А–Є—З–Є–љ–µ. –Ф–ї—П —Б–Њ–±–ї—О–і–µ–љ–Є—П –њ—А–µ–і–њ–Њ–ї–Њ–ґ–µ–љ–Є–є –Љ–Њ–і–µ–ї–Є –Њ¬†–љ–µ–Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –њ—А–Є–Ј–љ–∞–Ї–Њ–≤ –Є–Ј –љ–µ–µ –Є—Б–Ї–ї—О—З–∞–ї–Є—Б—М –њ–µ—А–µ–Љ–µ–љ–љ—Л–µ —Б¬†–≤—Л—Б–Њ–Ї–Є–Љ (> 0,7) —Г—А–Њ–≤–љ–µ–Љ –Ї–Њ—А—А–µ–ї—П—Ж–Є–Є —А–µ–≥—А–µ—Б—Б–Є–Њ–љ–љ—Л—Е –Њ—Б—В–∞—В–Ї–Њ–≤. –Я—А–Є –њ–Њ—Б—В—А–Њ–µ–љ–Є–Є –Љ–Њ–і–µ–ї–Є –Є–Ј—Г—З–∞–µ–Љ—Л–µ —Д–∞–Ї—В–Њ—А—Л –і–Њ–±–∞–≤–ї—П–ї–Є—Б—М –Ї¬†–Є–Ј–≤–µ—Б—В–љ—Л–Љ –Љ–µ—В–Њ–і–∞–Љ —Г—Б–ї–Њ–≤–љ–Њ–≥–Њ –≤–Ї–ї—О—З–µ–љ–Є—П. –Т¬†–і–∞–ї—М–љ–µ–є—И–µ–Љ –±—Л–ї–∞ —В–∞–Ї–ґ–µ –њ–Њ—Б—В—А–Њ–µ–љ–∞ –Љ–Њ–і–µ–ї—М, –≤–Ї–ї—О—З–∞–≤—И–∞—П –≤—Б–µ –Є–Ј—Г—З–∞–µ–Љ—Л–µ —Д–∞–Ї—В–Њ—А—Л. –†–∞–Ј–Љ–µ—А —Н—Д—Д–µ–Ї—В–∞ –Њ–њ—А–µ–і–µ–ї—П–ї—Б—П –і–ї—П –≤—Б–µ—Е —Б–ї—Г—З–∞–µ–≤ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ—Л–Љ —А–Є—Б–Ї–Њ–Љ (–Ю–†). –Т¬†–љ–∞—И–µ–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї—Б—П —Г—А–Њ–≤–µ–љ—М –Ј–љ–∞—З–Є–Љ–Њ—Б—В–Є p < 0,05.
–†–µ–Ј—Г–ї—М—В–∞—В—Л
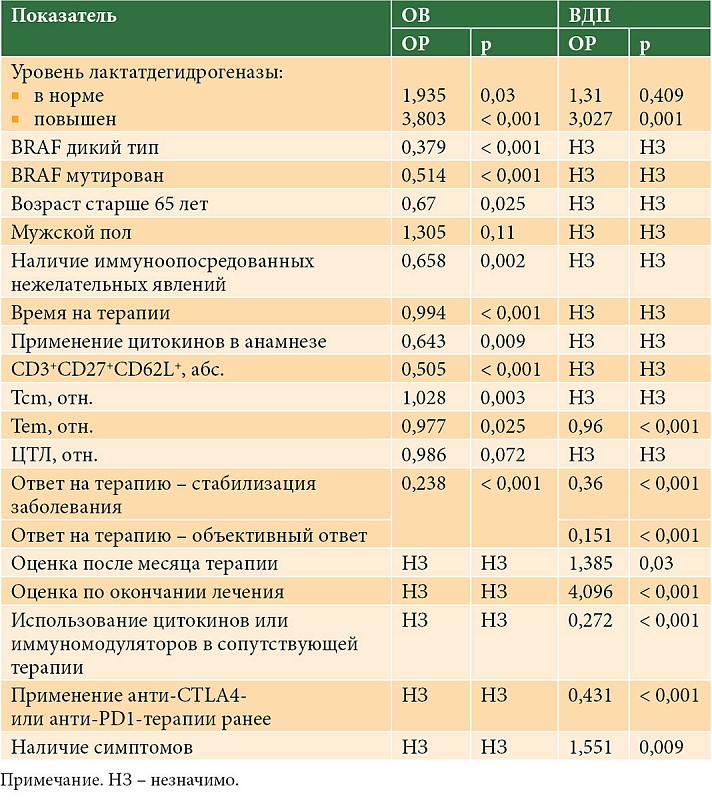
–Я—А–Є –Њ—Ж–µ–љ–Ї–µ —Г—А–Њ–≤–љ—П –љ–∞–Є–≤–љ—Л—Е –Ґn –і–Њ –љ–∞—З–∞–ї–∞ —В–µ—А–∞–њ–Є–Є –љ–µ —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ —Б–≤—П–Ј–Є —Б¬†–Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ—Л–Љ —Н—Д—Д–µ–Ї—В–Њ–Љ –ї–µ—З–µ–љ–Є—П (—А >0,05). –Я—А–Є –∞–љ–∞–ї–Є–Ј–µ –≤—Б–µ—Е –Ш–° –Њ–±–љ–∞—А—Г–ґ–µ–љ–Њ, —З—В–Њ —Б–љ–Є–ґ–µ–љ–Є–µ –і–∞–љ–љ–Њ–є —Б—Г–±–њ–Њ–њ—Г–ї—П—Ж–Є–Є –ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ (–Ї–∞–Ї –≤¬†–∞–±—Б–Њ–ї—О—В–љ—Л—Е, —В–∞–Ї –Є¬†–≤ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ—Л—Е –Ј–љ–∞—З–µ–љ–Є—П—Е) —З–∞—Й–µ –∞—Б—Б–Њ—Ж–Є–Є—А–Њ–≤–∞–ї–Њ—Б—М —Б¬†–њ—А–Њ–≥—А–µ—Б—Б–Є—А–Њ–≤–∞–љ–Є–µ–Љ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П (–Ю–† 1,236 –Є¬†–Ю–† 1,173, —А = 0,003 –Є¬†—А = 0,006 —Б–Њ–Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ). –Я—А–Є –±–Њ–ї–µ–µ –і–µ—В–∞–ї—М–љ–Њ–Љ —А–∞—Б—Б–Љ–Њ—В—А–µ–љ–Є–Є –і–Є–љ–∞–Љ–Є–Ї–Є —Б–Њ—Б—В–Њ—П–љ–Є—П –Њ–њ—Г—Е–Њ–ї–Є —Г¬†–±–Њ–ї—М–љ—Л—Е, –њ–Њ–ї—Г—З–∞–≤—И–Є—Е –Є–њ–Є–ї–Є–Љ—Г–Љ–∞–±, –≤—Л—П–≤–ї–µ–љ–∞ –∞–љ–∞–ї–Њ–≥–Є—З–љ–∞—П —В–µ–љ–і–µ–љ—Ж–Є—П (p = 0,024). –Я—А–Є —Н—В–Њ–Љ —А–∞–Ј–ї–Є—З–Є—П –≤¬†—З–∞—Б—В–Њ—В–µ —Б—В–∞–±–Є–ї–Є–Ј–∞—Ж–Є–є –њ—А–Њ—Ж–µ—Б—Б–∞ –і–Њ—Б—В–Є–≥–∞–ї–Є —Б—В–∞—В–Є—Б—В–Є—З–µ—Б–Ї–Є –Ј–љ–∞—З–Є–Љ–Њ–≥–Њ —Г—А–Њ–≤–љ—П (—А < 0,05). –Ю—В–љ–Њ—Б–Є—В–µ–ї—М–љ—Л–µ —Г—А–Њ–≤–љ–Є –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–µ–є —В–∞–Ї–ґ–µ —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –≤–ї–Є—П–ї–Є –љ–∞ —З–∞—Б—В–Њ—В—Г –Њ—В–і–µ–ї—М–љ—Л—Е –Ї–∞—В–µ–≥–Њ—А–Є–є –і–Є–љ–∞–Љ–Є–Ї–Є —Б–Њ—Б—В–Њ—П–љ–Є—П –Њ–њ—Г—Е–Њ–ї–Є. –С—Г—А–љ–Њ–µ –њ—А–Њ–≥—А–µ—Б—Б–Є—А–Њ–≤–∞–љ–Є–µ —З–∞—Й–µ —А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–ї–Њ—Б—М –њ—А–Є —Б–љ–Є–ґ–µ–љ–љ–Њ–Љ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ–Љ –Ј–љ–∞—З–µ–љ–Є–Є –љ–∞–Є–≤–љ—Л—Е –Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ (p < 0,05). –Я—А–Є –∞–љ–∞–ї–Є–Ј–µ –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В–Є —Б–≤—П–Ј–Є —Г—А–Њ–≤–љ—П –љ–∞–Є–≤–љ—Л—Е –Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ –Ї–∞–Ї –і–Њ –ї–µ—З–µ–љ–Є—П, —В–∞–Ї –Є¬†–≤ –њ—А–Њ—Ж–µ—Б—Б–µ —В–µ—А–∞–њ–Є–Є —Б¬†–Ю–Т –Є¬†–Т–Ф–Я –љ–µ –Ј–∞—Д–Є–Ї—Б–Є—А–Њ–≤–∞–љ–Њ (—А > 0,05).
–Я—А–Є –∞–љ–∞–ї–Є–Ј–µ –њ—А–Њ–≥–љ–Њ—Б—В–Є—З–µ—Б–Ї–Њ–є —А–Њ–ї–Є Tcm –Є¬†–Ґem –Њ–Ї–∞–Ј–∞–ї–Њ—Б—М, —З—В–Њ –њ—А–Є –њ–Њ–≤—Л—И–µ–љ–Є–Є –њ–µ—А–≤—Л—Е —А–Є—Б–Ї —Б–Љ–µ—А—В–Є —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В—Б—П (–Ю–† 1,02; —А = 0,02), —В–Њ–≥–і–∞ –Ї–∞–Ї —Б¬†—А–Њ—Б—В–Њ–Љ –≤—В–Њ—А—Л—Е —Г–Љ–µ–љ—М—И–∞–µ—В—Б—П (–Ю–† 0,401; —А = 0,008). –Ю—В–љ–Њ—Б–Є—В–µ–ї—М–љ—Л–є —Г—А–Њ–≤–µ–љ—М Tem —Б–ї—Г–ґ–Є–ї —В–∞–Ї–ґ–µ –њ—А–Њ–≥–љ–Њ—Б—В–Є—З–µ—Б–Ї–Є–Љ —Д–∞–Ї—В–Њ—А–Њ–Љ –њ—А–Є –∞–љ–∞–ї–Є–Ј–µ –Т–Ф–Я (–Ю–† 0,961; —А = 10-5). –Я—А–Є –Њ—Ж–µ–љ–Ї–µ —Г—А–Њ–≤–љ–µ–є –Ґ-–Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є –і–Њ –ї–µ—З–µ–љ–Є—П —Б–≤—П–Ј–Є —Б¬†–њ–Њ–Ї–∞–Ј–∞—В–µ–ї—П–Љ–Є –≤—Л–ґ–Є–≤–∞–µ–Љ–Њ—Б—В–Є –љ–µ –Њ–±–љ–∞—А—Г–ґ–µ–љ–Њ (—А > 0,05).
–Р–±—Б–Њ–ї—О—В–љ—Л–є —Г—А–Њ–≤–µ–љ—М CD3+CD27+CD62L+ —П–≤–ї—П–ї—Б—П –±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л–Љ —Д–∞–Ї—В–Њ—А–Њ–Љ –њ—А–Њ–≥–љ–Њ–Ј–∞ –Ю–Т (–Ю–† 0,529, —А = 0,001), —В–Њ–≥–і–∞ –Ї–∞–Ї –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ—Л–є —Г—А–Њ–≤–µ–љ—М CD3+CD28+CD62L+ –ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤¬†вАУ –љ–µ–±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л–Љ (–Ю–† 1,012; —А = 0,009). –Я—А–Є —Н—В–Њ–Љ –і–∞–љ–љ—Л–µ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є –љ–µ –≤–ї–Є—П–ї–Є –љ–∞ —А–Є—Б–Ї –њ—А–Њ–≥—А–µ—Б—Б–Є—А–Њ–≤–∞–љ–Є—П –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П (—А > 0,05). –£—А–Њ–≤–µ–љ—М CD3+CD27+CD62L+ –Є¬†CD3+CD28+CD62L+ –ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ –і–Њ –ї–µ—З–µ–љ–Є—П –љ–µ –±—Л–ї —Б–≤—П–Ј–∞–љ —Б¬†–њ–Њ–Ї–∞–Ј–∞—В–µ–ї—П–Љ–Є –Ю–Т –Є¬†–Т–Ф–Я (p > 0,05).
–£—А–Њ–≤–µ–љ—М CTLA4-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ —Б–ї—Г–ґ–Є–ї –љ–µ–±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л–Љ —Д–∞–Ї—В–Њ—А–Њ–Љ –њ—А–Њ–≥–љ–Њ–Ј–∞ –Т–Ф–Я —В–Њ–ї—М–Ї–Њ –њ—А–Є –µ–≥–Њ –Њ—Ж–µ–љ–Ї–µ –і–Њ –љ–∞—З–∞–ї–∞ —В–µ—А–∞–њ–Є–Є (–Ю–† 1,045; —А = 0,037).
–Я—А–Є –њ–Њ—Б—В—А–Њ–µ–љ–Є–Є –Њ–±—Й–µ–є –Љ–љ–Њ–≥–Њ—Д–∞–Ї—В–Њ—А–љ–Њ–є –Љ–Њ–і–µ–ї–Є, —Г—З–Є—В—Л–≤–∞–≤—И–µ–є –≤—Б–µ –∞–љ–∞–ї–Є–Ј–Є—А—Г–µ–Љ—Л–µ –љ–∞–Љ–Є —Б—Г–±–њ–Њ–њ—Г–ї—П—Ж–Є–Є (—В–∞–±–ї. 2), –Њ–Ї–∞–Ј–∞–ї–Њ—Б—М, —З—В–Њ —Б¬†–њ–Њ–≤—Л—И–µ–љ–Є–µ–Љ —Г—А–Њ–≤–љ—П —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л—Е —Б—Г–±–њ–Њ–њ—Г–ї—П—Ж–Є–є —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б–љ–Є–ґ–∞–ї—Б—П —А–Є—Б–Ї –њ—А–Њ–≥—А–µ—Б—Б–Є—А–Њ–≤–∞–љ–Є—П –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П –Є¬†—Б–Љ–µ—А—В–Є, —В–Њ–≥–і–∞ –Ї–∞–Ї –њ—А–Є –љ–∞–Ї–Њ–њ–ї–µ–љ–Є–Є –Ї–ї–µ—В–Њ–Ї Tcm, –љ–∞–Њ–±–Њ—А–Њ—В, –≤–Њ–Ј—А–∞—Б—В–∞–ї. –Ь–Њ–ґ–љ–Њ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ —Б–Ї–∞–Ј–∞–љ–љ–Њ–µ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г–µ—В –Њ¬†–љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ—Б—В–Є –Њ–±–µ—Б–њ–µ—З–µ–љ–Є—П —Г—Б–њ–µ—И–љ–Њ–є –∞–Ї—В–Є–≤–∞—Ж–Є–Є –Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤, –і–Њ—Б—В–Є–ґ–µ–љ–Є–µ –Ї–Њ—В–Њ—А–Њ–є –љ–∞ —Д–Њ–љ–µ –ї–µ—З–µ–љ–Є—П —В—А–∞–љ—Б–ї–Є—А—Г–µ—В—Б—П –≤¬†–њ–Њ–≤—Л—И–µ–љ–Є–µ –њ—А–Њ–і–Њ–ї–ґ–Є—В–µ–ї—М–љ–Њ—Б—В–Є –ґ–Є–Ј–љ–Є –њ–∞—Ж–Є–µ–љ—В–Њ–≤.
–Ю–±—Б—Г–ґ–і–µ–љ–Є–µ
–Э–∞–Є–≤–љ—Л–µ –Ґ-–Ї–ї–µ—В–Ї–Є CD3+/CD45RA+/CD62L+ —А–∞—Б–њ–Њ–Ј–љ–∞—О—В —З—Г–ґ–µ—А–Њ–і–љ—Л–µ –∞–љ—В–Є–≥–µ–љ—Л, –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–љ—Л–µ –≤¬†–∞—Б–њ–µ–Ї—В–µ –Љ–Њ–ї–µ–Ї—Г–ї –≥–ї–∞–≤–љ–Њ–≥–Њ –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞ –≥–Є—Б—В–Њ—Б–Њ–≤–Љ–µ—Б—В–Є–Љ–Њ—Б—В–Є, —Ж–Є—А–Ї—Г–ї–Є—А—Г—О—В –≤¬†–њ–µ—А–Є—Д–µ—А–Є—З–µ—Б–Ї–Є—Е –Њ—А–≥–∞–љ–∞—Е –Є–Љ–Љ—Г–љ–љ–Њ–є —Б–Є—Б—В–µ–Љ—Л, –Ґ-–Ј–∞–≤–Є—Б–Є–Љ—Л—Е –Ј–Њ–љ–∞—Е –≤—В–Њ—А–Є—З–љ—Л—Е –ї–Є–Љ—Д–Њ–Є–і–љ—Л—Е –Њ—А–≥–∞–љ–Њ–≤ –Є¬†–њ–µ—А–Є—Д–µ—А–Є—З–µ—Б–Ї–Њ–є –Ї—А–Њ–≤–Є, —Б—З–Є—В–∞—О—В—Б—П ¬Ђ–љ–∞–Є–≤–љ—Л–Љ–Є¬ї –і–Њ –Ї–Њ–љ—В–∞–Ї—В–∞ —Б¬†–∞–љ—В–Є–≥–µ–љ–њ—А–µ–Ј–µ–љ—В–Є—А—Г—О—Й–Є–Љ–Є –Ї–ї–µ—В–Ї–∞–Љ–Є. –Ш–Ј—Г—З–∞–µ–Љ—Л–µ –Ї–ї–µ—В–Ї–Є –µ—Й–µ –љ–µ –њ—А–Є–Њ–±—А–µ–ї–Є —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л–µ —Б–≤–Њ–є—Б—В–≤–∞, –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л–µ –і–ї—П —А–µ–∞–ї–Є–Ј–∞—Ж–Є–Є –њ—А–Њ—В–Є–≤–Њ–Њ–њ—Г—Е–Њ–ї–µ–≤–Њ–≥–Њ –Є–Љ–Љ—Г–љ–љ–Њ–≥–Њ –Њ—В–≤–µ—В–∞, –Є, –њ–Њ –≤—Б–µ–є –≤–µ—А–Њ—П—В–љ–Њ—Б—В–Є, –Є–Љ–µ–љ–љ–Њ –њ–Њ—Н—В–Њ–Љ—Г –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ –љ–∞–Љ–Є —А–µ–Ј—Г–ї—М—В–∞—В—Л –њ–Њ —Г–≤–µ–ї–Є—З–µ–љ–Є—О –і–Њ–ї–Є —Н—В–Є—Е –Ї–ї–µ—В–Њ–Ї —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–њ—А–Њ–≥—А–µ—Б—Б–Є—А–Њ–≤–∞–љ–Є–µ–Љ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П –њ–Њ–Ј–≤–Њ–ї—П—О—В –љ–∞–Љ —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞—В—М –љ–∞–Ї–Њ–њ–ї–µ–љ–Є–µ –≤¬†—Ж–Є—А–Ї—Г–ї—П—Ж–Є–Є –Ґ-–Ї–ї–µ—В–Њ–Ї, –њ–Њ–ї–љ–Њ—Б—В—М—О –ї–Є—И–µ–љ–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л—Е —Б–≤–Њ–є—Б—В–≤, –≤¬†–Ї–∞—З–µ—Б—В–≤–µ –Љ–∞—А–Ї–µ—А–∞ –љ–µ–±–ї–∞–≥–Њ–њ—А–Є—П—В–љ–Њ–≥–Њ –њ—А–Њ–≥–љ–Њ–Ј–∞ [2вАУ5].
–Ш–љ—В–µ—А–µ—Б –Ї¬†–њ–Њ–њ—Г–ї—П—Ж–Є–Є –Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ —Б¬†–Є–Љ–Љ—Г–љ–Њ—Д–µ–љ–Њ—В–Є–њ–Њ–Љ ¬Ђ–љ–∞–Є–≤–љ—Л–µ¬ї –≤—Л–Ј–≤–∞–љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П–Љ–Є, –≤¬†–Ї–Њ—В–Њ—А—Л—Е –њ–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ —Б—А–µ–і–Є —Н—В–Њ–є –њ–Њ–њ—Г–ї—П—Ж–Є–Є –Љ–Њ–≥—Г—В –±—Л—В—М ¬Ђ–љ–∞–Є–≤–љ–Њ-—А–µ–≤–µ—А—В–∞–љ—В–љ—Л–µ¬ї –Ї–ї–µ—В–Ї–Є. –°—З–Є—В–∞–µ—В—Б—П, —З—В–Њ –њ–µ—А–≤–Є—З–љ–∞—П —Б—В–Є–Љ—Г–ї—П—Ж–Є—П –Ґ-–Ї–ї–µ—В–Њ–Ї –Ј–∞–њ—Г—Б–Ї–∞–µ—В –Њ–і–љ–Њ–љ–∞–њ—А–∞–≤–ї–µ–љ–љ—Г—О –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–Ї—Г –Њ—В –љ–∞–Є–≤–љ—Л—Е –Ї¬†—Н—Д—Д–µ–Ї—В–Њ—А–љ—Л–Љ —Б—Г–±–њ–Њ–њ—Г–ї—П—Ж–Є—П–Љ –Є¬†–Ї–ї–µ—В–Ї–∞–Љ –њ–∞–Љ—П—В–Є. –Т–Љ–µ—Б—В–µ —Б¬†—В–µ–Љ G. Frumento –Є¬†—Б–Њ–∞–≤—В. (2020) –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ –Є–љ—В–µ—А–ї–µ–є–Ї–Є–љ 7 –Љ–Њ–ґ–µ—В –њ—А–Є–≤–µ—Б—В–Є –Ї¬†—Д–µ–љ–Њ—В–Є–њ–Є—З–µ—Б–Ї–Њ–є —А–µ–≤–µ—А—Б–Є–Є –љ–µ–і–∞–≤–љ–Њ –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–љ—Л—Е CD8+ –Ґ-–Ї–ї–µ—В–Њ–Ї —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –Є¬†—Н—Д—Д–µ–Ї—В–Њ—А–љ–Њ–є –њ–∞–Љ—П—В–Є —З–µ–ї–Њ–≤–µ–Ї–∞ –≤¬†–љ–∞–Є–≤–љ—Л–є —Д–µ–љ–Њ—В–Є–њ. –≠—В–Є ¬Ђ–љ–∞–Є–≤–љ–Њ-—А–µ–≤–µ—А—В–∞–љ—В–љ—Л–µ¬ї –Ї–ї–µ—В–Ї–Є CD45RA+/CD62L+/CCR7+/CXCR3+ –і–µ–Љ–Њ–љ—Б—В—А–Є—А—Г—О—В —Д–µ–љ–Њ—В–Є–њ, –∞–љ–∞–ї–Њ–≥–Є—З–љ—Л–є —Д–µ–љ–Њ—В–Є–њ—Г —А–∞–љ–µ–µ –Ј–∞—А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–љ–љ—Л—Е –њ–Њ–њ—Г–ї—П—Ж–Є–є –Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є ¬Ђ—Б—В–≤–Њ–ї–Њ–≤—Л—Е –Ї–ї–µ—В–Њ–Ї¬ї, –Њ–љ–Є –њ–Њ–і–≤–µ—А–≥–∞—О—В—Б—П –±—Л—Б—В—А–Њ–є –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–Ї–µ –Є¬†—Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–Љ—Г –Њ—В–≤–µ—В—Г –њ–Њ—Б–ї–µ –≤—В–Њ—А–Є—З–љ–Њ–є —Б—В–Є–Љ—Г–ї—П—Ж–Є–Є [6].
–§–µ–љ–Њ—В–Є–њ–Є—З–µ—Б–Ї–∞—П —А–µ–≤–µ—А—Б–Є—П –њ–Њ –Ї—А–∞–є–љ–µ–є –Љ–µ—А–µ —З–∞—Б—В–Є—З–љ–Њ –Њ–±—К—П—Б–љ—П–µ—В –≥–µ–љ–µ—А–∞—Ж–Є—О CD8+ –Ґ-–Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є ¬Ђ—Б—В–≤–Њ–ї–Њ–≤—Л—Е –Ї–ї–µ—В–Њ–Ї¬ї –Є¬†–≤—Л—П–≤–ї—П–µ—В –Ї–ї–µ—В–Ї–Є –≤¬†—Д–µ–љ–Њ—В–Є–њ–Є—З–µ—Б–Ї–Є –љ–∞–Є–≤–љ–Њ–Љ –њ—Г–ї–µ CD8+ –Ґ-–Ї–ї–µ—В–Њ–Ї, –Ї–Њ—В–Њ—А—Л–µ —Н–њ–Є–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є –љ–∞—Б—В—А–Њ–µ–љ—Л –љ–∞ –≤—В–Њ—А–Є—З–љ—Г—О —Б—В–Є–Љ—Г–ї—П—Ж–Є—О. –Ґ–∞–Ї–∞—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П –і–∞–µ—В –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–Є–µ –Њ¬†–Љ–µ—Е–∞–љ–Є–Ј–Љ–∞—Е, –Ї–Њ—В–Њ—А—Л–µ –њ–Њ–і–і–µ—А–ґ–Є–≤–∞—О—В –Ґ-–Ї–ї–µ—В–Ї–Є –њ–∞–Љ—П—В–Є –Є¬†–Љ–Њ–≥—Г—В –љ–∞–њ—А–∞–≤–ї—П—В—М —В–µ—А–∞–њ–µ–≤—В–Є—З–µ—Б–Ї–Є–µ –Љ–∞–љ–Є–њ—Г–ї—П—Ж–Є–Є —Б¬†–і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–Ї–Њ–є –≤¬†—Ж–µ–љ—В—А–∞–ї—М–љ—Л–µ –Є¬†—Н—Д—Д–µ–Ї—В–Њ—А–љ—Л–µ –Ґ-–Ї–ї–µ—В–Ї–Є –њ–∞–Љ—П—В–Є [7].
–Т —А—П–і–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Ж–µ–љ—В—А–∞–ї—М–љ—Л–µ –Ґ-–Ї–ї–µ—В–Ї–Є –њ–∞–Љ—П—В–Є CD3+/CD45R0+/CD62L+ –і–µ–Љ–Њ–љ—Б—В—А–Є—А–Њ–≤–∞–ї–Є –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї—Г—О —Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ—Г—О –∞–Ї—В–Є–≤–љ–Њ—Б—В—М –Є¬†–њ—А–Њ—В–Є–≤–Њ–Њ–њ—Г—Е–Њ–ї–µ–≤—Л–є –Є–Љ–Љ—Г–љ–Є—В–µ—В –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б¬†—Н—Д—Д–µ–Ї—В–Њ—А–љ—Л–Љ–Є –Ґ-–Ї–ї–µ—В–Ї–∞–Љ–Є –њ–∞–Љ—П—В–Є CD3+/CD45R0+/CD62L-, –Ї–Њ—В–Њ—А—Л–µ —П–≤–ї—П—О—В—Б—П —В–µ—А–Љ–Є–љ–∞–ї—М–љ–Њ –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–∞–љ–љ—Л–Љ–Є –Ї–ї–µ—В–Ї–∞–Љ–Є –њ–∞–Љ—П—В–Є, —Ж–Є—А–Ї—Г–ї–Є—А—Г—О—В –≤¬†–њ–µ—А–Є—Д–µ—А–Є—З–µ—Б–Ї–Њ–є –Ї—А–Њ–≤–Є, –≤—В–Њ—А–Є—З–љ—Л—Е –ї–Є–Љ—Д–Њ–Є–і–љ—Л—Е –Њ—А–≥–∞–љ–∞—Е, –і–Њ—Б—В–Є–≥–∞—О—В —Н—Д—Д–µ–Ї—В–Њ—А–љ–Њ–є —Д—Г–љ–Ї—Ж–Є–Є –Є¬†–Є—Б—В–Њ—Й–µ–љ–Є—П —Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –≤—Б–Ї–Њ—А–µ –њ–Њ—Б–ї–µ –њ–Њ–≤—В–Њ—А–љ–Њ–є —Б—В–Є–Љ—Г–ї—П—Ж–Є–Є [8вАУ10].
–Э–∞–ї–Є—З–Є–µ –∞–і–≥–µ–Ј–Є–Њ–љ–љ–Њ–є –Љ–Њ–ї–µ–Ї—Г–ї—Л CD62L –Є¬†—Е–µ–Љ–Њ–Ї–Є–љ–Њ–≤–Њ–≥–Њ —А–µ—Ж–µ–њ—В–Њ—А–∞ CCR7 –љ–∞ –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є –Ґ-–Ї–ї–µ—В–Њ–Ї —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –њ–∞–Љ—П—В–Є –њ–Њ–Ј–≤–Њ–ї—П–µ—В –Њ—В–ї–Є—З–Є—В—М –і–∞–љ–љ—Г—О –њ–Њ–њ—Г–ї—П—Ж–Є—О –Њ—В –Ґ-–Ї–ї–µ—В–Њ–Ї —Н—Д—Д–µ–Ї—В–Њ—А–љ–Њ–є –њ–∞–Љ—П—В–Є, –ї–Є—И–µ–љ–љ—Л—Е —Г–Ї–∞–Ј–∞–љ–љ—Л—Е –Љ–Њ–ї–µ–Ї—Г–ї.
–Т –љ–∞—И–µ–Љ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–Є –∞—Б—Б–Њ—Ж–Є–∞—Ж–Є—П —Г–≤–µ–ї–Є—З–µ–љ–Є—П –Ґ-–Ї–ї–µ—В–Њ–Ї —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –њ–∞–Љ—П—В–Є —Б¬†–љ–µ–±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л–Љ –Є—Б—Е–Њ–і–Њ–Љ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П –Љ–Њ–ґ–µ—В –±—Л—В—М —В–µ—Б–љ–Њ —Б–≤—П–Ј–∞–љ–∞ —Б¬†–±–Є–Њ–ї–Њ–≥–Є–µ–є —Н—В–Є—Е –Ї–ї–µ—В–Њ–Ї. –≠—В–∞ –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В—М –Ґ-–Ї–ї–µ—В–Њ–Ї —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –њ–∞–Љ—П—В–Є, —Б¬†–Њ–і–љ–Њ–є —Б—В–Њ—А–Њ–љ—Л, –њ–Њ–Ј–≤–Њ–ї—П–µ—В –Є–Љ –і–ї–Є—В–µ–ї—М–љ–Њ–µ –≤—А–µ–Љ—П —Ж–Є—А–Ї—Г–ї–Є—А–Њ–≤–∞—В—М –≤¬†–Њ—А–≥–∞–љ–Є–Ј–Љ–µ, —Б¬†–і—А—Г–≥–Њ–є, –Њ–њ—А–µ–і–µ–ї—П–µ—В –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–µ–љ–љ—Г—О –ї–Њ–Ї–∞–ї–Є–Ј–∞—Ж–Є—О –≤–Њ –≤—В–Њ—А–Є—З–љ—Л—Е –ї–Є–Љ—Д–Њ–Є–і–љ—Л—Е –Њ—А–≥–∞–љ–∞—Е –Є¬†–љ–µ –њ–Њ–Ј–≤–Њ–ї—П–µ—В –Љ–Є–≥—А–Є—А–Њ–≤–∞—В—М –≤¬†–Њ—З–∞–≥–Є –Њ–њ—Г—Е–Њ–ї–µ–≤–Њ–≥–Њ —А–Њ—Б—В–∞, —А–∞–≤–љ–Њ –Ї–∞–Ї –Є¬†–љ–µ –њ–Њ–і—А–∞–Ј—Г–Љ–µ–≤–∞–µ—В –љ–∞–ї–Є—З–Є–µ —Г¬†–Ґ-–Ї–ї–µ—В–Њ–Ї —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –њ–∞–Љ—П—В–Є —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л—Е —Б–≤–Њ–є—Б—В–≤, –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л—Е –і–ї—П —А–∞–Ј—А—Г—И–µ–љ–Є—П –Њ–њ—Г—Е–Њ–ї–Є [11].
–Я—А–Є–љ—Ж–Є–њ–Є–∞–ї—М–љ—Л–Љ –Њ—В–ї–Є—З–Є–µ–Љ –Ґ-–Ї–ї–µ—В–Њ–Ї —Н—Д—Д–µ–Ї—В–Њ—А–љ–Њ–є –њ–∞–Љ—П—В–Є –Њ—В –Ґ-–Ї–ї–µ—В–Њ–Ї —Ж–µ–љ—В—А–∞–ї—М–љ–Њ–є –њ–∞–Љ—П—В–Є —П–≤–ї—П–µ—В—Б—П –≤—Л—А–∞–ґ–µ–љ–љ–∞—П —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –Ї¬†–њ—А–Њ–і—Г–Ї—Ж–Є–Є —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л—Е —Ж–Є—В–Њ–Ї–Є–љ–Њ–≤ –Є¬†–њ—А–Њ—П–≤–ї–µ–љ–Є—О —Ж–Є—В–Њ–ї–Є—В–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –≤¬†–Њ—В–љ–Њ—И–µ–љ–Є–Є –Ї–ї–µ—В–Њ–Ї-–Љ–Є—И–µ–љ–µ–є [10, 12, 13]. –С–Њ–ї–µ–µ —В–Њ–≥–Њ, –љ–∞ —Н—В–Њ–є –њ–Њ–њ—Г–ї—П—Ж–Є–Є –Ї–ї–µ—В–Њ–Ї –≤–Њ–Ј—А–∞—Б—В–∞—О—В —Н–Ї—Б–њ—А–µ—Б—Б–Є—П –Љ–Њ–ї–µ–Ї—Г–ї ¬Ђ—Е–Њ—Г–Љ–Є–љ–≥–∞¬ї –Є¬†–Љ–Є–≥—А–∞—Ж–Є—П –≤¬†–≤–Њ—Б–њ–∞–ї–µ–љ–љ—Л–µ —В–Ї–∞–љ–Є. –Т–Њ–Ј–Љ–Њ–ґ–љ–Њ, –Є–Љ–µ–љ–љ–Њ –њ–Њ—Н—В–Њ–Љ—Г –љ–∞–Љ–Є –≤—Л—П–≤–ї–µ–љ–∞ —В–µ—Б–љ–∞—П –≤–Ј–∞–Є–Љ–Њ—Б–≤—П–Ј—М –Љ–µ–ґ–і—Г —А–Њ—Б—В–Њ–Љ —Н—В–Є—Е –Ї–ї–µ—В–Њ–Ї –≤¬†—Ж–Є—А–Ї—Г–ї—П—Ж–Є–Є —Г¬†–њ–∞—Ж–Є–µ–љ—В–Њ–≤ —Б¬†–±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л–Љ —В–µ—З–µ–љ–Є–µ–Љ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П.
–Э–∞–ї–Є—З–Є–µ –љ–∞ –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є –Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ –Ї–Њ-—Б—В–Є–Љ—Г–ї–Є—А—Г—О—Й–Є—Е –Љ–Њ–ї–µ–Ї—Г–ї CD27 –Є¬†CD28 –њ–Њ–Ј–≤–Њ–ї—П–µ—В –Ї–ї–µ—В–Ї–∞–Љ –∞–Ї—В–Є–≤–Є—А–Њ–≤–∞—В—М—Б—П –Є¬†—А–∞–Ј–≤–Є–≤–∞—В—М –∞–љ—В–Є–≥–µ–љ—Б–њ–µ—Ж–Є—Д–Є—З–µ—Б–Ї–Є–є –Њ—В–≤–µ—В in vivo –њ—А–Є –љ–∞–ї–Є—З–Є–Є –∞–љ—В–Є–≥–µ–љ–љ–Њ–є —Б—В–Є–Љ—Г–ї—П—Ж–Є–Є –Є¬†–ї–Є–≥–∞–љ–і–Њ–≤ –і–ї—П —Г–Ї–∞–Ј–∞–љ–љ—Л—Е –≤—Л—И–µ –Љ–Њ–ї–µ–Ї—Г–ї –љ–∞ –Љ–µ–Љ–±—А–∞–љ–µ –∞–љ—В–Є–≥–µ–љ–њ—А–µ–Ј–µ–љ—В–Є—А—Г—О—Й–Є—Е –Ї–ї–µ—В–Њ–Ї, —З—В–Њ –±—Л–ї–Њ –њ–Њ–Ї–∞–Ј–∞–љ–Њ –≤¬†—Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞—Е –љ–∞ –Љ—Л—И–∞—Е [14]. –Х—Б–ї–Є —Ж–Є—А–Ї—Г–ї–Є—А—Г—О—Й–Є–µ –Ґ-—Е–µ–ї–њ–µ—А—Л –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –≤—Б–µ–≥–і–∞ —Н–Ї—Б–њ—А–µ—Б—Б–Є—А—Г—О—В –Њ–±–µ —Н—В–Є –Љ–Њ–ї–µ–Ї—Г–ї—Л –љ–∞ —Б–≤–Њ–µ–є –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є, —В–Њ —Б—А–µ–і–Є –Њ–±—Й–µ–≥–Њ –њ—Г–ї–∞ —Ж–Є—В–Њ—В–Њ–Ї—Б–Є—З–µ—Б–Ї–Є—Е CD8+ –Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ –љ–∞–±–ї—О–і–∞–µ—В—Б—П —Б—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ–µ –Є–Ј–Љ–µ–љ–µ–љ–Є–µ –≤¬†–Є—Е —Н–Ї—Б–њ—А–µ—Б—Б–Є–Є [15]. –¶–Є—В–Њ—В–Њ–Ї—Б–Є—З–µ—Б–Ї–Є–µ –Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В—Л —Б¬†—Д–µ–љ–Њ—В–Є–њ–Њ–Љ CD28+CD27high –њ–Њ–ї—Г—З–Є–ї–Є –љ–∞–Ј–≤–∞–љ–Є–µ –љ–∞–Є–≤–љ—Л—Е –љ–µ–њ—А–∞–є–Љ–Є—А–Њ–≤–∞–љ–љ—Л—Е –Ґ-–Ї–ї–µ—В–Њ–Ї. –Т¬†–Є—Е —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ–µ –љ–µ –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞–ї–Є—Б—М —Ж–Є—В–Њ—В–Њ–Ї—Б–Є—З–µ—Б–Ї–Є–µ –≥—А–∞–љ—Г–ї—Л, –љ–Њ –љ–∞ –Љ–µ–Љ–±—А–∞–љ–µ —Н–Ї—Б–њ—А–µ—Б—Б–Є—А–Њ–≤–∞–ї–Є—Б—М CD45RA –Є¬†CCR7. –Р–љ—В–Є–≥–µ–љ CCR7 —В–∞–Ї–ґ–µ –±—Л–ї –Њ–±–љ–∞—А—Г–ґ–µ–љ –љ–∞ –њ–Њ–њ—Г–ї—П—Ж–Є–Є –Ї–ї–µ—В–Њ–Ї —Б¬†—Д–µ–љ–Њ—В–Є–њ–Њ–Љ CD28+CD27high-to-posCD45RA-, –њ–Њ–ї—Г—З–Є–≤—И–Є—Е –љ–∞–Ј–≤–∞–љ–Є–µ —А–∞–љ–љ–Є—Е –∞–љ—В–Є–≥–µ–љ-–њ—А–∞–є–Љ–Є—А–Њ–≤–∞–љ–љ—Л—Е CD3+CD8+-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤. –Ш¬†–љ–∞–Ї–Њ–љ–µ—Ж, –Ї–ї–µ—В–Ї–Є, –ї–Є—И–µ–љ–љ—Л–µ CD27 –Є¬†CD28, –њ–Њ–ї—Г—З–Є–ї–Є –љ–∞–Ј–≤–∞–љ–Є–µ –њ–Њ–Ј–і–љ–Є—Е —Ж–Є—В–Њ—В–Њ–Ї—Б–Є—З–µ—Б–Ї–Є—Е –Ґ-–Ї–ї–µ—В–Њ–Ї. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –±—Л–ї–∞ –≤—Л–і–µ–ї–µ–љ–∞ –њ–Њ–њ—Г–ї—П—Ж–Є—П –њ—А–Њ–Љ–µ–ґ—Г—В–Њ—З–љ—Л—Е –Ї–ї–µ—В–Њ–Ї —Б¬†—Д–µ–љ–Њ—В–Є–њ–Њ–Љ CD28-CD27+. –Я–Њ –Љ–µ—А–µ —Г—В—А–∞—В—Л CCR7, CD27 –Є¬†CD28 –≤–Њ–Ј—А–∞—Б—В–∞–ї–∞ —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –Ї–ї–µ—В–Њ–Ї —Б–Є–љ—В–µ–Ј–Є—А–Њ–≤–∞—В—М —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л–µ —Ж–Є—В–Њ–Ї–Є–љ—Л, –њ—А–Њ—П–≤–ї—П—В—М —Ж–Є—В–Њ—В–Њ–Ї—Б–Є—З–µ—Б–Ї–Є–µ —Б–≤–Њ–є—Б—В–≤–∞ –њ–Њ –Њ—В–љ–Њ—И–µ–љ–Є—О –Ї¬†–Ї–ї–µ—В–Ї–∞–Љ-–Љ–Є—И–µ–љ—П–Љ, —Б–Є–љ—В–µ–Ј–Є—А–Њ–≤–∞—В—М –Є¬†–љ–∞–Ї–∞–њ–ї–Є–≤–∞—В—М —Ж–Є—В–Њ–ї–Є—В–Є—З–µ—Б–Ї–Є–µ –≥—А–∞–љ—Г–ї—Л, –љ–Њ –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ —Б–љ–Є–ґ–∞–ї–∞—Б—М —Б–њ–Њ—Б–Њ–±–љ–Њ—Б—В—М –Ї¬†–њ—А–Њ–ї–Є—Д–µ—А–∞—Ж–Є–Є. –Я—А–Є —Н—В–Њ–Љ –њ–Њ –Љ–µ—А–µ —Б–Њ–Ј—А–µ–≤–∞–љ–Є—П –Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤ —Г—В—А–∞—В–∞ CD28 –њ—А–µ–і—И–µ—Б—В–≤—Г–µ—В —Б–љ–Є–ґ–µ–љ–Є—О CD27, –∞¬†–њ–µ—А–µ—Е–Њ–і –Є–Ј —Б–Њ—Б—В–Њ—П–љ–Є—П CD27+CD28- —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–µ—В—Б—П –Ї–∞–Ї –Њ–і–Є–љ –Є–Ј –Ї–ї—О—З–µ–≤—Л—Е –њ–µ—А–µ—Е–Њ–і–Њ–≤ –≤¬†—Б—В–Њ—А–Њ–љ—Г —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л—Е –Ї–ї–µ—В–Њ–Ї [8, 16]. –Э–µ —Б–ї—Г—З–∞–є–љ–Њ –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ –љ–∞–Љ–Є –і–∞–љ–љ—Л–µ –Њ–± —Г–≤–µ–ї–Є—З–µ–љ–Є–Є —З–Є—Б–ї–∞ CD3+CD27+CD62L+ –Ґ-–Ї–ї–µ—В–Њ–Ї —А–∞—Б—Б–Љ–∞—В—А–Є–≤–∞–ї–Є—Б—М –Ї–∞–Ї –±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л–є —Д–∞–Ї—В–Њ—А –њ—А–Њ–≥–љ–Њ–Ј–∞ –Ю–Т. –Т¬†—В–Њ –ґ–µ –≤—А–µ–Љ—П –љ–∞–Ї–Њ–њ–ї–µ–љ–Є–µ CD3+CD28+CD62L+ –Ґ-–ї–Є–Љ—Д–Њ—Ж–Є—В–Њ–≤, –љ–∞—Е–Њ–і–Є–≤—И–Є—Е—Б—П –љ–∞ —Б–∞–Љ—Л—Е —А–∞–љ–љ–Є—Е —Б—В–∞–і–Є—П—Е –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–Ї–Є –Є¬†–ї–Є—И–µ–љ–љ—Л—Е —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л—Е —Б–≤–Њ–є—Б—В–≤, –±—Л–ї–Њ —Б–≤—П–Ј–∞–љ–Њ —Б¬†–љ–µ–±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л–Љ –њ—А–Њ–≥–љ–Њ–Ј–Њ–Љ.
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –≥–µ–љ–µ—А–∞—Ж–Є—П, –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–Ї–∞ –Є¬†–њ–Њ–і–і–µ—А–ґ–∞–љ–Є–µ –Ґ-–Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є –≤¬†—Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ–Њ –Њ–њ—В–Є–Љ–∞–ї—М–љ–Њ–Љ —Б–Њ—Б—В–Њ—П–љ–Є–Є –Є–Љ–µ—О—В —А–µ—И–∞—О—Й–µ–µ –Ј–љ–∞—З–µ–љ–Є–µ –і–ї—П —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—П –њ—А–Њ—В–Є–≤–Њ–Њ–њ—Г—Е–Њ–ї–µ–≤–Њ–≥–Њ –Є–Љ–Љ—Г–љ–Є—В–µ—В–∞, –≤¬†—В–Њ–Љ —З–Є—Б–ї–µ –љ–∞ —Д–Њ–љ–µ –њ—А–Є–Љ–µ–љ–µ–љ–Є—П –Є–Љ–Љ—Г–љ–Њ–Њ–љ–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –њ—А–Є –Љ–µ–ї–∞–љ–Њ–Љ–µ.
–Э–µ–і–∞–≤–љ–Є–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П —Б–њ–Њ—Б–Њ–±—Б—В–≤–Њ–≤–∞–ї–Є –Њ—В–Ї—А—Л—В–Є—О –Ґ-–Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є, —А–µ–Ј–Є–і–µ–љ—В–љ—Л—Е –≤¬†—В–Ї–∞–љ—П—Е, –Є¬†—А–∞—Б—И–Є—А–Є–ї–Є –љ–∞—И–Є –Ј–љ–∞–љ–Є—П –Њ¬†—В—А–∞–љ—Б–Ї—А–Є–њ—Ж–Є–Њ–љ–љ—Л—Е –Є¬†—Н–њ–Є–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є—Е –Њ—Б–љ–Њ–≤–∞—Е –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–Ї–Є –Є¬†–і–Є—Б—А–µ–≥—Г–ї—П—Ж–Є–Є CD8+ –Ґ-–Ї–ї–µ—В–Њ–Ї [3].
–Ъ —Б–Њ–ґ–∞–ї–µ–љ–Є—О, –њ—А–Њ–≥—А–µ—Б—Б –Є–Љ–Љ—Г–љ–Њ—В–µ—А–∞–њ–Є–Є –љ–∞—Е–Њ–і–Є—В—Б—П –љ–∞ –Ї—А–Є—В–Є—З–µ—Б–Ї–Њ–Љ —Н—В–∞–њ–µ, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М –Є–Љ–Љ—Г–љ–Њ–Њ–љ–Ї–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –њ—А–µ–њ–∞—А–∞—В–Њ–≤ –Њ–≥—А–∞–љ–Є—З–Є–≤–∞–µ—В—Б—П –љ–µ–±–Њ–ї—М—И–Њ–є –і–Њ–ї–µ–є –њ–∞—Ж–Є–µ–љ—В–Њ–≤ –Є¬†—З–∞—Б—В–Њ –љ–µ –Њ–±–µ—Б–њ–µ—З–Є–≤–∞–µ—В –і–Њ–ї–≥–Њ–≤—А–µ–Љ–µ–љ–љ—Г—О –њ–Њ–ї—М–Ј—Г –Њ—В –ї–µ—З–µ–љ–Є—П [3, 5, 9, 10, 12, 15]. –†–∞—Б–Ї—А—Л—В–Є–µ –њ–Њ—В–µ–љ—Ж–Є–∞–ї–∞ –Є–Љ–Љ—Г–љ–Њ—В–µ—А–∞–њ–Є–Є —В—А–µ–±—Г–µ—В —А–∞–Ј—А–∞–±–Њ—В–Ї–Є —Б—В—А–∞—В–µ–≥–Є–є, –Ї–Њ—В–Њ—А—Л–µ –Њ–і–љ–Њ–≤—А–µ–Љ–µ–љ–љ–Њ –Є–љ–і—Г—Ж–Є—А—Г—О—В –Љ–Њ—Й–љ—Л–є —Н—Д—Д–µ–Ї—В–Њ—А–љ—Л–є –Њ—В–≤–µ—В –Є¬†–љ–∞–і–µ–ґ–љ–Њ —Д–Њ—А–Љ–Є—А—Г—О—В —Б—В–∞–±–Є–ї—М–љ—Л–µ —Д—Г–љ–Ї—Ж–Є–Њ–љ–∞–ї—М–љ–Њ –∞–Ї—В–Є–≤–љ—Л–µ –њ—Г–ї—Л –Ґ-–Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є, —Б–њ–Њ—Б–Њ–±–љ—Л–µ –њ—А–µ–і–Њ—В–≤—А–∞—В–Є—В—М —А–µ—Ж–Є–і–Є–≤ –Є–ї–Є –і–∞–ї—М–љ–µ–є—И–µ–µ –њ—А–Њ–≥—А–µ—Б—Б–Є—А–Њ–≤–∞–љ–Є–µ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П. –Ъ—А–∞–є–љ–µ –≤–∞–ґ–љ–Њ, —З—В–Њ–±—Л –Њ—Б–љ–Њ–≤–љ—Л–µ –Є¬†–љ–Њ–≤—Л–µ –Ї–Њ–љ—Ж–µ–њ—Ж–Є–Є, –Њ–±–љ–Њ–≤–ї–µ–љ–љ—Л–µ –Ј–љ–∞–љ–Є—П –Њ¬†–±–Є–Њ–ї–Њ–≥–Є–Є –Ґ-–Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є –±—Л–ї–Є —Б–≤–Њ–µ–≤—А–µ–Љ–µ–љ–љ–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ—Л –≤¬†—А–∞–Ј—А–∞–±–Њ—В–Ї–∞—Е –Є–Љ–Љ—Г–љ–Њ—В–µ—А–∞–њ–Є–Є —А–∞–Ї–∞.
–Ч–∞–Ї–ї—О—З–µ–љ–Є–µ
–Я—А–Њ–≤–µ–і–µ–љ–љ–Њ–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–µ –і–Њ–њ–Њ–ї–љ—П–µ—В —Б–Є—Б—В–µ–Љ–љ–Њ–µ –њ–Њ–љ–Є–Љ–∞–љ–Є–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤, –ї–µ–ґ–∞—Й–Є—Е –≤¬†–Њ—Б–љ–Њ–≤–µ –і–Є—Д—Д–µ—А–µ–љ—Ж–Є—А–Њ–≤–Ї–Є –Ґ-–Ї–ї–µ—В–Њ–Ї –њ–∞–Љ—П—В–Є. –Т—Б–µ –±–Њ–ї—М—И–µ–µ –Ј–љ–∞—З–µ–љ–Є–µ –њ—А–Є–Њ–±—А–µ—В–∞–µ—В –Њ–њ—А–µ–і–µ–ї–µ–љ–Є–µ –Є—Е —Г—А–Њ–≤–љ—П –і–Њ –Є¬†–≤ –њ—А–Њ—Ж–µ—Б—Б–µ –Є–Љ–Љ—Г–љ–Њ—В–µ—А–∞–њ–Є–Є, —З—В–Њ —Б–ї—Г–ґ–Є—В –Ј–љ–∞—З–Є–Љ—Л–Љ —Д–∞–Ї—В–Њ—А–Њ–Љ –њ—А–Њ–≥–љ–Њ–Ј–∞ –Ј–∞–±–Њ–ї–µ–≤–∞–љ–Є—П. –Э–µ–Њ–±—Е–Њ–і–Є–Љ—Л –і–∞–ї—М–љ–µ–є—И–Є–µ –њ—А–Њ—Б–њ–µ–Ї—В–Є–≤–љ—Л–µ –Ї–ї–Є–љ–Є—З–µ—Б–Ї–Є–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –≤¬†–і–∞–љ–љ–Њ–Љ –љ–∞–њ—А–∞–≤–ї–µ–љ–Є–Є.
A.V. Novik, PhD, I.V. Kudryavtsev, PhD, T.L. Nekhayeva, PhD, N.V. Yemelyanova, A.B. Danilova, PhD, N.A. Yefremova, Ye.M. Anokhina, –Р.I. Semyonova, PhD, D.Kh. Latipova, PhD, G.M. Teletayeva, PhD, S.A. Protsenko, PhD, Prof., I.A. Balduyeva, PhD
N.N. Petrov National Medical Research Center of Oncology
Saint-Petersburg State Pediatric Medical University
Contact person: Alexey V. Novik, anovik@list.ru
The generation, differentiation and maintenance of memory T-cells in the functionally optimal state is crucial for the effectiveness of the formation of antitumor immunity, including against the background of the use of immuno-oncological drugs in melanoma. The aim of the study was to test the hypothesis that the level of T-cells in the peripheral blood can serve as a marker of the therapy and survival of patients with melanoma effectiveness.
–£–≤–∞–ґ–∞–µ–Љ—Л–є –њ–Њ—Б–µ—В–Є—В–µ–ї—М uMEDp!
–£–≤–µ–і–Њ–Љ–ї—П–µ–Љ –Т–∞—Б –Њ —В–Њ–Љ, —З—В–Њ –Ј–і–µ—Б—М —Б–Њ–і–µ—А–ґ–Є—В—Б—П –Є–љ—Д–Њ—А–Љ–∞—Ж–Є—П, –њ—А–µ–і–љ–∞–Ј–љ–∞—З–µ–љ–љ–∞—П –Є—Б–Ї–ї—О—З–Є—В–µ–ї—М–љ–Њ –і–ї—П —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–≤ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П.
–Х—Б–ї–Є –Т—Л –љ–µ —П–≤–ї—П–µ—В–µ—Б—М —Б–њ–µ—Ж–Є–∞–ї–Є—Б—В–Њ–Љ –Ј–і—А–∞–≤–Њ–Њ—Е—А–∞–љ–µ–љ–Є—П, –∞–і–Љ–Є–љ–Є—Б—В—А–∞—Ж–Є—П –љ–µ –љ–µ—Б–µ—В –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ–Њ—Б—В–Є –Ј–∞ –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–µ –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ –њ–Њ—Б–ї–µ–і—Б—В–≤–Є—П, –≤–Њ–Ј–љ–Є–Ї—И–Є–µ –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ —Б–∞–Љ–Њ—Б—В–Њ—П—В–µ–ї—М–љ–Њ–≥–Њ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –Т–∞–Љ–Є –Є–љ—Д–Њ—А–Љ–∞—Ж–Є–Є —Б –њ–Њ—А—В–∞–ї–∞ –±–µ–Ј –њ—А–µ–і–≤–∞—А–Є—В–µ–ї—М–љ–Њ–є –Ї–Њ–љ—Б—Г–ї—М—В–∞—Ж–Є–Є —Б –≤—А–∞—З–Њ–Љ.
–Э–∞–ґ–Є–Љ–∞—П –љ–∞ –Ї–љ–Њ–њ–Ї—Г ¬Ђ–Т–Њ–є—В–Є¬ї, –Т—Л –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В–µ, —З—В–Њ —П–≤–ї—П–µ—В–µ—Б—М –≤—А–∞—З–Њ–Љ –Є–ї–Є —Б—В—Г–і–µ–љ—В–Њ–Љ –Љ–µ–і–Є—Ж–Є–љ—Б–Ї–Њ–≥–Њ –≤—Г–Ј–∞.